表达三种人源基因的SARS-CoV-2易感模型猪的构建方法及其应用与流程
表达三种人源基因的sars-cov-2易感模型猪的构建方法及其应用
技术领域
1.本发明属于生物技术领域,涉及基因编辑技术,具体涉及采用crispr/cas9系统及同源重组技术构建的在基因组的特定位置整合并表达三个人源基因(ace2基因、axl基因和tmprss2基因)的猪重组细胞,利用该重组细胞可以制备sars-cov-2易感模型猪,从而将其用于生物医药领域。
背景技术:
2.人类血管紧张素i转化酶2(hace2)为人类血管紧张素转换酶同源物,是一种锌金属蛋白酶,属于1型跨膜蛋白,其作为sars-cov-2和sars-cov等冠状病毒在人体内的受体蛋白,广泛存在于人体各个组织中,是目前已知导致sars-cov-2进入宿主细胞并造成宿主发病的关键蛋白。
3.sars-cov-2进入宿主细胞需要依赖于病毒的刺突蛋白(s)与细胞受体的结合,并通过宿主细胞蛋白酶激活s蛋白,而人丝氨酸蛋白酶2(tmprss2)在此过程中行使着激活s蛋白的关键功能。
4.此外,人axl受体酪氨酸激酶已被发现是一种不依赖于ace2的sars-cov-2新受体,其通过特异性结合sars-cov-2刺突蛋白n端的ntd区域,介导病毒在呼吸系统细胞上的吸附与内化。
5.目前,尚未有针对sars-cov-2的特效药物,世界各国都在争分夺秒地开展sars-cov-2疫苗的研发。而动物模型对于阐明sars-cov-2的感染与发病机制、传播途径以及抗病毒药物和疫苗及抗体的开发和评价至关重要。
6.现有技术中,常用的模式动物为小鼠。小鼠不论从体型、器官大小、生理、病理等方面都与人相差巨大,不能真实地模拟人类正常的生理、病理状态。而猪作为大动物,是人类长期以来主要的肉食供应动物,其体型大小和生理功能与人类近似,易于大规模繁殖饲养,而且在伦理道德及动物保护等方面要求较低,是理想的人类疾病模型动物。
7.基因编辑是近年来不断取得重大发展的一种生物技术,其包括从基于同源重组的基因编辑到基于核酸酶的zfn、talen、crispr/cas9等编辑技术,其中crispr/cas9技术是当前最先进的基因编辑技术。目前,基因编辑技术被越来越多地应用到动物模型的制作上。
技术实现要素:
8.本发明解决的技术问题为提供了采用crispr/cas9系统及同源重组技术构建的在基因组的特定位置整合并表达三个人源基因(ace2基因、axl基因和tmprss2基因)的猪重组细胞,进一步,利用该重组细胞通过体细胞核移植技术得到了sars-cov-2易感模型猪,该模型猪可用于下一步的药物、疫苗及抗体筛选和药物、疫苗及抗体效果评价以及发病机制的研究等。
9.本发明提供了一种制备重组细胞的方法,包括如下步骤:将命名为dna分子甲的
dna分子整合至猪细胞的基因组dna,得到重组细胞;所述dna分子甲表达人源ace2蛋白、人源axl蛋白和人源tmprss2蛋白。
10.具体的,人源ace2蛋白(hace2蛋白)如seq id no:15所示。
11.具体的,人源axl蛋白(haxl蛋白),如seq id no:16所示。
12.具体的,人源tmprss2蛋白(htmprss2蛋白),如seq id no:17所示。
13.人源ace2蛋白由人ace2基因编码。
14.人源axl蛋白由人axl基因编码。
15.人源tmprss2蛋白由人tmprss2基因编码。
16.具体的,人ace2基因(hace2基因)如seq id no:14中第2290-4704位核苷酸所示。
17.具体的,人axl基因(haxl基因)如seq id no:14中第4771-6648位核苷酸所示。
18.具体的,人tmprss2基因(htmprss2基因)如seq id no:14中第6712-8298位核苷酸所示。
19.所述dna分子甲中,人ace2基因、人axl基因和人tmprss2基因存在于同一表达盒。
20.所述dna分子甲中,人ace2基因、人axl基因和人tmprss2基因由同一启动子启动表达。
21.具体的,所述启动子为人hef1α启动子。
22.具体的,人hef1α启动子如seq id no:14中第1094-2271位核苷酸所示。
23.所述dna分子甲中,人ace2基因、人axl基因和人tmprss2基因由同一终止密码子终止表达。
24.所述dna分子甲中,人ace2基因、人axl基因和人tmprss2基因共用一个poly(a)信号。
25.具体的,所述poly(a)信号为ef1αpoly(a)。
26.具体的,ef1αpoly(a)如seq id no:14中第8302-8874位核苷酸所示。
27.所述dna分子甲中,人ace2基因、人axl基因和人tmprss2基因存在于同一编码框,基因之间由2a肽的编码基因间隔。2a肽又称为自剪接肽或自裂解肽或2a自剪切肽或2a自裂解肽。
28.所述2a肽具体可为p2a肽。
29.所述2a肽具体可为t2a肽。
30.具体的,所述p2a肽的编码基因如seq id no:14中第4705-4770位核苷酸所示。
31.具体的,所述t2a肽的编码基因如seq id no:14中第6649-6711位核苷酸所示。
32.具体的,所述dna分子甲中,人ace2基因、人axl基因和人tmprss2基因所在的编码框如seq id no:14中第1094-8874位核苷酸所示。
33.所述dna分子甲中还包括抗性筛选基因表达盒。所述抗性筛选基因可为编码puromycin抗性蛋白的基因。所述抗性筛选基因表达盒如seq id no:14中第8987-10262位核苷酸所示。
34.所述dna分子甲中还包括loxp序列。
35.所述dna分子甲中具体包括2个loxp序列,分别如seq id no:14中第8911-8944位核苷酸、第10307-10340位核苷酸所示。
36.所述dna分子甲中还包括绝缘子。
37.所述dna分子甲中具体包括2个绝缘子,分别如seq id no:14中第887-1087位核苷酸、第10349-10549位核苷酸所示。
38.所述dna分子甲自上游至下游依次包括如下区段:人hef1α启动子、hace2基因、编码自p2a肽的核苷酸、haxl基因、编码t2a肽的核苷酸、htmprss2基因、终止密码子、ef1αpoly(a)、loxp序列、sv40启动子、编码puromycin抗性蛋白的核苷酸、终止密码子、sv40poly(a)、loxp序列。
39.所述dna分子甲自上游至下游依次包括如下区段:绝缘子1、人hef1α启动子、hace2基因、编码p2a肽的核苷酸、haxl基因、编码t2a肽的核苷酸、htmprss2基因、终止密码子、ef1αpoly(a)、loxp序列、sv40启动子、编码puromycin抗性蛋白的核苷酸、终止密码子、sv40 poly(a)、loxp序列、绝缘子3。
40.具体的,所述dna分子甲如seq id no:14中第887-10549位核苷酸所示。
41.具体的,所述dna分子甲如seq id no:14中第881-10549位核苷酸所示。
42.所述“将命名为dna分子甲的dna分子整合至猪细胞的基因组dna”的实现方式为:将命名为dna分子乙的dna分子导入猪细胞或者将具有所述dna分子乙的重组质粒导入猪细胞;所述dna分子乙中,具有所述dna分子甲且在所述dna分子甲的上游具有上游同源臂且在所述dna分子甲的下游具有下游同源臂,所述上游同源臂和所述下游同源臂用于将所述dna分子甲整合至猪细胞的基因组dna。
43.所述同源臂为针对rosa26基因的同源臂,上游同源臂为sh1左臂,下游同源臂为sh1右臂。sh1左臂如seq id no:3中第9-339位核苷酸所示,sh1右臂如seq id no:3中第9184-10195位核苷酸所示。
44.所述同源臂为针对aavs1基因的同源臂,上游同源臂为sh2左臂,下游同源臂为sh2右臂。sh2左臂如seq id no:4所示,sh2右臂如seq id no:5所示。
45.所述同源臂为针对h11位点的同源臂,上游同源臂为sh3左臂,下游同源臂为sh3右臂。sh3左臂如seq id no:6所示,sh3右臂如seq id no:7所示。
46.所述同源臂为针对col1a1基因的同源臂,上游同源臂为sh4左臂,下游同源臂为sh4右臂。sh4左臂如seq id no:8所示,sh4右臂如seq id no:9所示。
47.所述同源臂为针对col1a1基因的同源臂,上游同源臂为sh4左臂,下游同源臂为sh4右臂。sh4左臂如seq id no:14中第9-880位核苷酸所示,sh4右臂如seq id no:14中第10550-11276位核苷酸所示。
48.所述dna分子乙自上游至下游依次包括如下区段:上游同源臂、绝缘子1、人hef1α启动子、hace2基因、编码p2a肽的核苷酸、haxl基因、编码t2a肽的核苷酸、htmprss2基因、终止密码子、ef1αpoly(a)、loxp序列、sv40启动子、编码puromycin抗性蛋白的核苷酸、终止密码子、sv40 poly(a)、loxp序列、绝缘子3、下游同源臂。
49.具体的,所述dna分子乙如seq id no:14中第9-11276位核苷酸所示。
50.具体的,具有所述dna分子乙的重组质粒如seq id no:14所示。
51.dna分子甲整合至猪细胞的基因组dna的rosa26基因或者aavs1基因或者h11位点或者col1a1基因中。优选为col1a1基因。
52.dna分子甲整合至猪细胞的基因组dna的rosa26安全港插入位点或者aavs1安全港插入位点或者h11安全港插入位点或者col1a1安全港插入位点。优选为col1a1安全港插入
位点。
53.dna分子甲整合至猪细胞的基因组dna的rosa26基因中指的是将dna分子甲插入至基因组dna中sh1左臂和sh1右臂之间;sh1左臂如seq id no:3中第9-339位核苷酸所示,sh1右臂如seq id no:3中第9184-10195位核苷酸所示。
54.dna分子甲整合至猪细胞的基因组dna的aavs1基因中指的是将dna分子甲插入至基因组dna中sh2左臂和sh2右臂之间;sh2左臂如seq id no:4所示,sh2右臂如seq id no:5所示。
55.dna分子甲整合至猪细胞的基因组dna的h11位点中指的是将dna分子甲插入至基因组dna中sh3左臂和sh3右臂之间;sh3左臂如seq id no:6所示,sh3右臂如seq id no:7所示。
56.dna分子甲整合至猪细胞的基因组dna的col1a1基因中指的是将dna分子甲插入至基因组dna中sh4左臂和sh4右臂之间;sh4左臂如seq id no:8所示,sh4右臂如seq id no:9所示。
57.dna分子甲整合至猪细胞的基因组dna的col1a1基因中指的是将dna分子甲插入至基因组dna中sh4左臂和sh4右臂之间;sh4左臂如seq id no:14中第9-880位核苷酸所示,sh4右臂如seq id no:14中第10550-11276位核苷酸所示。
58.所述方法中,具有所述dna分子乙的重组质粒与两个辅助质粒共同导入猪细胞;所述两个辅助质粒为sgrna质粒和cas9质粒;
59.sgrna质粒转录得到特异sgrna;所述特异sgrna为sgrna
rosa26-g3
、sgrna
aavs1-g4
、sgrna
h11-g1
或sgrna
col1a1-g3
;sgrna
rosa26-g3
的靶序列结合区如seq id no:10中第1-20位核苷酸所示;sgrna
aavs1-g4
的靶序列结合区如seq id no:11中第1-20位核苷酸所示;sgrna
h11-g1
的靶序列结合区如seq id no:12中第1-20位核苷酸所示;sgrna
col1a1-g3
的靶序列结合区如seq id no:13中第1-20位核苷酸所示;
60.cas9质粒表达cas9蛋白。
61.所述cas9质粒具体可为质粒pkg-ge3。
62.具体来说,所述sgrna质粒是借助限制性内切酶bbsi将特异sgrna的靶序列结合区的编码序列插入pkg-u6grna载体得到的。
63.具有所述dna分子乙的重组质粒、sgrna质粒和cas9质粒的摩尔配比具体可为1:3:1。
64.猪细胞、具有所述dna分子乙的重组质粒、sgrna质粒和cas9质粒的配比具体可为:约20万个猪细胞:1.3μg具有所述dna分子乙的重组质粒:0.8μg sgrna质粒:0.9μg cas9质粒。
65.具体的,猪基因组中所述rosa26安全港插入位点及其周边区域如seq id no:18所示。
66.具体的,猪基因组中所述aavs1安全港插入位点及其周边区域如seq id no:19所示。
67.具体的,猪基因组中所述h11安全港插入位点及其周边区域如seq id no:20所示。
68.具体的,猪基因组中所述col1a1安全港插入位点及其周边区域如seq id no:21所示。
69.本发明还保护一种试剂盒,包括以上任一所述dna分子乙。
70.本发明还保护一种试剂盒,包括具有以上任一所述dna分子乙的重组质粒。
71.所述试剂盒还包括所述sgrna质粒和所述的cas9质粒。
72.具有所述dna分子乙的重组质粒、sgrna质粒和cas9质粒的摩尔配比具体可为1:3:1。
73.本发明还保护以上任一所述dna分子乙在制备试剂盒中的应用。
74.本发明还保护具有以上任一所述dna分子乙的重组质粒在制备试剂盒中的应用。
75.本发明还保护具有以上任一所述dna分子乙的重组质粒、sgrna质粒和cas9质粒在制备试剂盒中的应用。
76.以上任一所述试剂盒的用途为如下(a)或(b)或(c):(a)制备重组细胞;(b)制备sars-cov-2易感模型猪;(c)制备sars-cov-2易感猪细胞模型或sars-cov-2易感猪组织模型或sars-cov-2易感猪器官模型。
77.本发明还保护以上任一所述dna分子乙、具有以上任一所述dna分子乙的重组质粒或以上任一所述试剂盒的应用,为如下(a)或(b)或(c):(a)制备重组细胞;(b)制备sars-cov-2易感模型猪;(c)制备sars-cov-2易感猪细胞模型或sars-cov-2易感猪组织模型或sars-cov-2易感猪器官模型。
78.以上任一所述猪细胞为猪原代成纤维细胞。
79.所述猪可为任何品种的猪,优选的,所述猪可为从江香猪。
80.所述猪具体可为初生猪。
81.质粒pkg-ge3中,具有特异融合基因;所述特异融合基因编码特异融合蛋白;
82.所述特异融合蛋白自n端至c端依次包括如下元件:两个核定位信号、cas9蛋白、两个核定位信号、p2a肽、荧光报告蛋白、t2a肽、抗性筛选标记蛋白;
83.质粒pkg-ge3中,由ef1a启动子启动所述特异融合基因的表达;
84.质粒pkg-ge3中,所述特异融合基因下游具有wpre序列元件、3’ltr序列元件和bgh poly(a)signal序列元件。
85.质粒pkg-ge3中,依次具有如下元件:cmv增强子、ef1a启动子、所述特异融合基因、wpre序列元件、3’ltr序列元件、bgh poly(a)signal序列元件。
86.所述特异融合蛋白中,cas9蛋白上游的两个核定位信号为sv40核定位信号,cas9蛋白下游的两个核定位信号为nucleoplasmin核定位信号。
87.所述特异融合蛋白中,荧光报告蛋白具体可为egfp蛋白。
88.所述特异融合蛋白中,抗性筛选标记蛋白具体可为puromycin抗性蛋白。
89.p2a肽的氨基酸序列为“atnfsllkqagdveenpgp”(断裂位置为c端开始第一个氨基酸残基和第二个氨基酸残基之间)。
90.t2a肽的氨基酸序列为“egrgslltcgdveenpgp”(断裂位置为c端开始第一个氨基酸残基和第二个氨基酸残基之间)。
91.特异融合基因具体如seq id no:1中第911-6706位核苷酸所示。
92.cmv增强子如seq id no:1中第395-680位核苷酸所示。
93.ef1a启动子如seq id no:1中第682-890位核苷酸所示。
94.wpre序列元件如seq id no:1中第6722-7310位核苷酸所示。
[0095]3’
ltr序列元件如seq id no:1中第7382-7615位核苷酸所示。
[0096]
bgh poly(a)signal序列元件如seq id no:1中第7647-7871位核苷酸所示。
[0097]
质粒pkg-ge3具体如seq id no:1所示。
[0098]
质粒pkg-u6grna中,具有seq id no:2中第2280-2637位核苷酸所示的dna分子。
[0099]
质粒pkg-u6grna具体如seq id no:2所示。
[0100]
具体的,sgrna
rosa26-g3
如seq id no:10所示。
[0101]
具体的,sgrna
aavs1-g4
如seq id no:11所示。
[0102]
具体的,sgrna
h11-g1
如seq id no:12所示。
[0103]
具体的,sgrna
col1a1-g3
如seq id no:13所示。
[0104]
sgrna
rosa26-g3
(seq id no:10):
[0105]
gaaggagcaaacugacauggguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0106]
sgrna
aavs1-g4
(seq id no:11):
[0107]
ugcagugggucuuuggggacguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0108]
sgrna
h11-g1
(seq id no:12):
[0109]
uuccaggaacauaagaaaguguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0110]
sgrna
col1a1-g3
(seq id no:13):
[0111]
gcagucucagcaaccacugaguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0112]
本发明还保护以上任一所述方法制备得到的重组细胞。
[0113]
具体的,所述重组细胞可为如下重组细胞:与猪原代成纤维细胞相比,重组细胞的基因组dna的差异仅在于:在猪基因组dna中sh4左臂和sh4右臂之间插入了seq id no:14中第881-10549位核苷酸所示的dna分子。
[0114]
本发明还保护所述重组细胞在制备sars-cov-2易感模型猪中的应用。
[0115]
将所述重组细胞作为核移植供体细胞进行体细胞克隆,可以得到克隆猪,即为sars-cov-2易感模型猪。
[0116]
将所述重组细胞作为核移植供体细胞进行体细胞克隆从而制备克隆猪具体可采用如下方法:
[0117]
(1)取离体卵巢,从直径3~6mm的卵泡中抽取卵丘卵母细胞复合体(cumulus-oocyte complexes,cocs),选择至少具有三层致密卵丘细胞的cocs,接种至4孔板中,每孔装有400μl猪卵母细胞体外成熟培养基,每孔接种50个,每孔覆盖400μl的矿物油,将含cocs的培养板在38.5℃、5%co2和饱和湿度的培养箱中培养42小时;
[0118]
(2)完成步骤(1)后,用0.1%(w/v)透明质酸酶反复吹打去除cocs的扩张卵丘细胞,将具有完整膜且含排出的第一极体的卵母细胞在含有0.1mg/ml地美可辛、0.05m蔗糖和4mg/ml牛血清白蛋白的ncsu23培养基中培养0.5-1h,促使卵母细胞核突起,然后使用尖部倾斜的显微注射针在含有10μm hepes、0.3%(w/v)聚乙烯吡咯烷酮、10%fbs、0.1mg/ml地美可辛和5mg/ml细胞松弛素b的tyrode乳酸培养基中去除突起的细胞核和极体;然后,将单
个核供体细胞注入去核卵母细胞的卵周隙,使用胚胎细胞融合仪在融合培养基中用200v/mm的直流脉冲将核供体细胞与受体卵母细胞融合20μs;然后,将重构胚在pzm-3培养基中培养2h以允许细胞核重编程,然后在激活培养基中用150v/mm的单脉冲激活100μs;然后,将激活的重构胚置于含5μg/ml细胞松弛素b的pzm-3培养基中,培养2小时,以进一步激活胚胎;然后将重构胚置于pzm-3培养基中培养;
[0119]
(3)将激活后培养6h的重构胚移植到代孕母猪的输卵管中,正常饲养,得到子代,即为克隆猪。
[0120]
代孕母猪具体可为二花脸母猪。
[0121]
代孕母猪具体可为9月龄的二花脸母猪。
[0122]
本发明还保护用所述重组细胞制备的sars-cov-2易感模型猪的猪细胞、猪组织或猪器官。
[0123]
本发明还保护以上任一所述重组细胞的应用。
[0124]
本发明还保护用所述重组细胞制备的sars-cov-2易感模型猪的猪细胞、猪组织或猪器官的应用。
[0125]
本发明还保护用所述重组细胞制备的sars-cov-2易感模型猪的应用。
[0126]
所述应用为如下(d1)或(d2)或(d3):
[0127]
(d1)筛选治疗新冠肺炎的药物和/或疫苗和/或抗体;
[0128]
(d2)进行新冠肺炎的药物和/或疫苗和/或抗体的效果评价;
[0129]
(d3)研究新冠肺炎的发病机制。
[0130]
人ace2基因(hace2基因)信息:编码angiotensin-converting enzyme 2[homo sapiens];位于人x号染色体;geneid为59272。angiotensin-converting enzyme 2[homo sapiens],又称为hace2蛋白。hace2蛋白是目前sars-cov-2及sars-cov等冠状病毒公认的受体蛋白,如seq id no:15所示(np_001358344.1)。
[0131]
人axl基因(haxl基因)信息:编码axl receptor tyrosine kinase[homo sapiens];位于人19号染色体;geneid为558。axl receptor tyrosine kinase[homo sapiens],又称为haxl蛋白。haxl蛋白是近期发现的不依赖于hace2蛋白的独立介导sars-cov-2感染的蛋白,如seq id no:16所示(np_001265528.1)。
[0132]
人tmprss2基因(htmprss2基因)信息:编码transmembrane protease serine 2[homo sapiens];位于人21号染色体;geneid为7113。transmembrane protease serine 2[homo sapiens],又称为htmprss2蛋白。htmprss2蛋白能够通过激活sars-cov-2的刺突蛋白(s)从而介导sars-cov-2与宿主细胞受体结合,进而促进sars-cov-2进入宿主细胞,如seq id no:17所示(np_001128571.1)。
[0133]
与现有技术相比,本发明至少具有如下有益效果:
[0134]
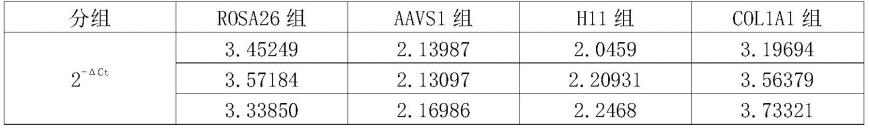
(1)本发明研究对象(猪)比其他动物(大小鼠、灵长类)具有更好的应用性。
[0135]
大小鼠等啮齿类动物不论从体型、器官大小、生理、病理等方面都与人相差巨大,无法真实地模拟人类正常的生理、病理状态。研究表明,95%以上在大小鼠中验证有效的药物在人类临床试验中是无效的。就大动物而言,灵长类是与人亲缘关系最近的动物,但其体型小、性成熟晚(6-7岁开始交配),且为单胎动物,群体扩繁速度极慢,饲养成本很高。另外,灵长类动物克隆效率低、难度大、成本高。
[0136]
而猪作为模型动物就没有上述缺点,猪是除灵长类外与人亲缘关系最近的动物,其体型、体重、器官大小等与人相近,在解剖学、生理学、免疫学、营养代谢、疾病发病机制等方面与人类极为相似。同时,猪的性成熟早(4-6个月),繁殖力高,一胎多仔,在2-3年内即可形成一个较大群体。另外,猪的克隆技术非常成熟,克隆及饲养成本也较灵长类低得多。
[0137]
(2)本发明针对猪基因组进行了4个安全港位点基因敲入后表达情况的摸索,从中筛选出了最佳的供外源基因插入的猪基因组安全港位点,可有效改善基因敲入后目的基因的表达情况。
[0138]
(3)利用本发明所得到的hace2-haxl-htmprss2纯合敲入单细胞克隆株进行体细胞核移植动物克隆可直接得到hace2-haxl-htmprss2纯合敲入的克隆猪,并且该纯合插入基因可稳定遗传。
[0139]
在小鼠模型制作中,通常采用受精卵显微注射基因编辑材料后再进行胚胎移植,因其直接获得纯合突变后代的概率非常低(低于5%),需要进行后代的杂交选育,这不太适用于妊娠期较长的大动物(如猪)模型制作。因此,本发明采用技术难度大、挑战性高的原代细胞体外编辑并筛选阳性编辑单细胞克隆的方法,后期再通过体细胞核移植动物克隆技术直接获得相应模型猪,可大大缩短模型猪制作周期,并节省人力、物力、财力。
[0140]
(4)本发明所制备的hace2-haxl-htmprss2人源化猪可用于冠状病毒动物感染模型进行临床前试验研究。
[0141]
本发明通过基因编辑手段获得了hace2、haxl和htmprss2三基因人源化模型猪,将有助于研究并揭示由hace2、haxl和htmprss2联合介导引发的sars-cov-2等冠状病毒的感染机制,并可用于进行药物筛选、药效评价、疫苗及抗体的应用效果测试等研究,能够为进一步的临床应用提供有效的实验数据,进而为预防和治疗人类sars-cov-2等冠状病毒感染疾病提供有力的实验手段。本发明对于针对sars-cov-2的药物、疫苗及抗体的研发及揭示该病毒的感染机制具有重大应用价值。
附图说明
[0142]
图1为质粒pb-1g 2r 3-puro-rosa26的结构示意图。
[0143]
图2为不同安全港位点调控gfp绿色荧光表达图。
[0144]
图3为不同安全港位点调控gfp基因转录水平荧光定量pcr结果。
[0145]
图4为不同安全港位点调控gfp蛋白表达的facs检测结果。
[0146]
图5为质粒pkg-hace2-haxl-htmprss2的结构示意图。
[0147]
图6为单细胞克隆hace2、haxl和htmprss2基因转录水平检测的结果。
[0148]
图7为单细胞克隆hace2、haxl和htmprss2的蛋白表达水平facs检测的结果。
[0149]
图8为体细胞克隆得到的模型猪的照片。
[0150]
图9为人源化猪的hace2、haxl和htmprss2基因转录水平检测的结果。
[0151]
图10为人源化猪的hace2、haxl和htmprss2蛋白表达水平facs检测的结果。
[0152]
图11为质粒pmd2.g-sars-c19的结构示意图。
[0153]
图12为质粒lenti-mcherry的结构示意图。
[0154]
图13为假病毒感染后的模型猪肺泡巨噬细胞的荧光信号照片。
[0155]
图14为假病毒感染后的野生型对照猪肺泡巨噬细胞的荧光信号照片。
具体实施方式
[0156]
下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。以下提供的实施例可作为本技术领域普通技术人员进行进一步改进的指南,并不以任何方式构成对本发明的限制。
[0157]
下述实施例中的实验方法,如无特殊说明,均为常规方法,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。实施例中构建的重组质粒,均已进行测序验证。完全培养液(%为体积比):15%胎牛血清(gibco)+83%dmem培养基(gibco)+1%penicillin-streptomycin(gibco)+1%hepes(solarbio)。细胞培养条件:37℃,5%co2、5%o2的恒温培养箱。
[0158]
实施例中采用的猪原代成纤维细胞均是用初生从江香猪(雌性,血型ao)的耳组织制备得到的。制备猪原代成纤维细胞的方法:
①
取猪耳组织0.5g,去除毛发及骨组织,然后用75%酒精浸泡30-40s,然后用含5%(体积比)penicillin-streptomycin(gibco)的pbs缓冲液洗涤5次,然后用pbs缓冲液洗涤一次;
②
用剪刀将组织剪碎,采用5ml 0.1%胶原酶溶液(sigma),37℃消化1h,然后500g离心5min,弃上清;
③
将沉淀用1ml完全培养液重悬,然后铺入含10ml完全培养液并已用0.2%明胶(vwr)封盘的直径为10cm的细胞培养皿中,培养至细胞长满皿底60%左右;
④
完成步骤
③
后,采用胰蛋白酶消化并收集细胞,然后重悬于完全培养液。用于进行后续电转实验。
[0159]
eef1a-mnls-hspcas9-egfp-puro简称质粒pkg-ge3(环形质粒),如seq id no:1所示。seq id no:1中,第395-680位核苷酸组成cmv增强子,第682-890位核苷酸组成ef1a启动子,第986-1006位核苷酸编码核定位信号(nls),第1016-1036位核苷酸编码核定位信号(nls),第1037-5161位核苷酸编码cas9蛋白,第5162-5209位核苷酸编码核定位信号(nls),第5219-5266位核苷酸编码核定位信号(nls),第5276-5332位核苷酸编码p2a肽(p2a肽的氨基酸序列为“atnfsllkqagdveenpgp”,断裂位置为c端开始第一个氨基酸残基和第二个氨基酸残基之间),第5333-6046位核苷酸编码egfp蛋白,第6056-6109位核苷酸编码t2a肽(t2a肽的氨基酸序列为“egrgslltcgdveenpgp”,断裂位置为c端开始第一个氨基酸残基和第二个氨基酸残基之间),第6110-6703位核苷酸编码puromycin蛋白(简称puro蛋白),第6722-7310位核苷酸组成wpre序列元件,第7382-7615位核苷酸组成3’ltr序列元件,第7647-7871位核苷酸组成bgh poly(a)signal序列元件。seq id no:1中,第911-6706形成融合基因,表达融合蛋白。由于p2a肽和t2a肽的存在,融合蛋白自发形成如下三个蛋白:具有cas9蛋白的蛋白、具有egfp蛋白的蛋白和具有puro蛋白的蛋白。记载于专利申请202011170395.1(申请公布号为cn 112522261a;申请公布日为2021.03.19)。
[0160]
pkg-u6grna载体,又称为质粒pkg-u6grna(环形质粒),如seq id no:2所示。seq id no:2中,第2280-2539位核苷酸组成hu6启动子,第2558-2637位核苷酸用于转录形成grna骨架。使用时,将20bp左右的dna分子(用于转录形成grna的靶序列结合区)插入质粒pkg-u6grna,形成重组质粒,在细胞中重组质粒转录得到grna。记载于专利申请202011170395.1(申请公布号为cn 112522261 a;申请公布日为2021.03.19)。
[0161]
实施例1、筛选供外源基因定点插入的猪基因组最佳安全港位点
[0162]
一、构建含gfp基因的不同安全港位点donor载体
[0163]
构建质粒pb-1g 2r 3-puro-rosa26、质粒pb-1g 2r 3-puro-aavs1、质粒pb-1g 2r3-puro-h11和质粒pb-1g 2r 3-puro-col1a1。四个质粒均为环形质粒。
[0164]
质粒pb-1g 2r 3-puro-rosa26如seq id no:3所示,结构示意图见图1。seq id no:3中,第9-339位核苷酸组成rosa26安全港插入位点5’端猪基因组区域(sh1左臂),第9184-10195位核苷酸组成rosa26安全港插入位点3’端猪基因组区域(sh1右臂)。seq id no:3中,第346-546、3132-3531、6506-6706、8975-9175位核苷酸分别组成4个不同的绝缘子区域。seq id no:3中,第637-1209位核苷酸组成ef-1αpoly(a)信号,第1216-1935位核苷酸编码egfp蛋白,第1954-3131位核苷酸组成ef-1α启动子,第3543-4042位核苷酸组成pgk启动子,第4059-4769位核苷酸编码mcherry蛋白,第4791-5015位核苷酸组成bgh poly(a)信号,第5054-6504位核苷酸为loxp-puro-loxp表达框区域,第6969-7233位核苷酸组成β-globin poly(a)信号,第7259-8974位核苷酸组成pcag启动子。
[0165]
质粒pb-1g 2r 3-puro-aavs1与质粒pb-1g 2r 3-puro-rosa26的差异仅在于:将sh1左臂替换为aavs1安全港插入位点5’端猪基因组区域(sh2左臂,sh2左臂如seq id no:4所示)并且将sh1右臂替换为aavs1安全港插入位点3’端猪基因组区域(sh2右臂,sh2右臂如seq id no:5所示)。
[0166]
质粒pb-1g 2r 3-puro-h11与质粒pb-1g 2r 3-puro-rosa26的差异仅在于:将sh1左臂替换为h11安全港插入位点5’端猪基因组区域(sh3左臂,sh3左臂如seq id no:6所示)并且将sh1右臂替换为h11安全港插入位点3’端猪基因组区域(sh3右臂,sh3右臂如seq id no:7所示)。
[0167]
质粒pb-1g 2r 3-puro-col1a1与质粒pb-1g 2r 3-puro-rosa26的差异仅在于:将sh1左臂替换为col1a1安全港插入位点5’端猪基因组区域(sh4左臂,sh4左臂如seq id no:8所示)并且将sh1右臂替换为col1a1安全港插入位点3’端猪基因组区域(sh4右臂,sh4右臂如seq id no:9所示)。
[0168]
二、猪rosa26、aavs1、h11、col1a1基因组安全港位点的高效切割靶点筛选
[0169]
通过前期筛选,rosa26安全港位点的高效切割靶点为sgrna
rosa26-g3
(切割效率38%),aavs1安全港位点的高效切割靶点为sgrna
aavs1-g4
(切割效率30%)、h11安全港位点的高效切割靶点为sgrna
h11-g1
(切割效率60%),col1a1安全港位点的高效切割靶点为sgrna
col1a1-g3
(切割效率56%)。
[0170]
靶点序列如下:
[0171]
sgrna
rosa26-g3
靶点:5
’‑
gaaggagcaaactgacatgg-3’;
[0172]
sgrna
aavs1-g4
靶点:5
’‑
tgcagtgggtctttggggac-3’;
[0173]
sgrna
h11-g1
靶点:5
’‑
ttccaggaacataagaaagt-3’;
[0174]
sgrna
col1a1-g3
靶点:5
’‑
gcagtctcagcaaccactga-3’。
[0175]
三、制备安全港位点grna重组载体
[0176]
取质粒pkg-u6grna,用限制性内切酶bbsi进行酶切,回收载体骨架(约3kb的线性大片段)。
[0177]
分别合成rosa26-g3-s和rosa26-g3-a,然后混合并进行退火,得到具有粘性末端的双链dna分子。将具有粘性末端的双链dna分子和载体骨架连接,得到质粒pkg-u6grna(rosa26-g3)。质粒pkg-u6grna(rosa26-g3)表达seq id no:10所示的sgrna
rosa26-g3
。
[0178]
分别合成aavs1-g4-s和aavs1-g4-a,然后混合并进行退火,得到具有粘性末端的双链dna分子。将具有粘性末端的双链dna分子和载体骨架连接,得到质粒pkg-u6grna(aavs1-g4)。质粒pkg-u6grna(aavs1-g4)表达seq id no:11所示的sgrna
aavs1-g4
。
[0179]
分别合成h11-g1-s和h11-g1-a,然后混合并进行退火,得到具有粘性末端的双链dna分子。将具有粘性末端的双链dna分子和载体骨架连接,得到质粒pkg-u6grna(h11-g1)。质粒pkg-u6grna(h11-g1)表达seq id no:12所示的sgrna
h11-g1
。
[0180]
分别合成col1a1-g3-s和col1a1-g3-a,然后混合并进行退火,得到具有粘性末端的双链dna分子。将具有粘性末端的双链dna分子和载体骨架连接,得到质粒pkg-u6grna(col1a1-g3)。质粒pkg-u6grna(col1a1-g3)表达seq id no:13所示的sgrna
col1a1-g3
。
[0181]
rosa26-g3-s、rosa26-g3-a、aavs1-g4-s、aavs1-g4-a、h11-g1-s、h11-g1-a、col1a1-g3-s和col1a1-g3-a均为单链dna分子。
[0182]
rosa26-g3-s:caccgaaggagcaaactgacatgg;
[0183]
rosa26-g3-a:aaacccatgtcagtttgctccttc。
[0184]
aavs1-g4-s:caccgtgcagtgggtctttggggac;
[0185]
aavs1-g4-a:aaacgtccccaaagacccactgcac。
[0186]
h11-g1-s:caccgttccaggaacataagaaagt;
[0187]
h11-g1-a:aaacactttcttatgttcctggaac。
[0188]
col1a1-g3-s:caccgcagtctcagcaaccactga;
[0189]
col1a1-g3-a:aaactcagtggttgctgagactgc。
[0190]
sgrna
rosa26-g3
(seq id no:10):
[0191]
gaaggagcaaacugacauggguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0192]
sgrna
aavs1-g4
(seq id no:11):
[0193]
ugcagugggucuuuggggacguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0194]
sgrna
h11-g1
(seq id no:12):
[0195]
uuccaggaacauaagaaaguguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0196]
sgrna
col1a1-g3
(seq id no:13):
[0197]
gcagucucagcaaccacugaguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu。
[0198]
四、含不同安全港插入位点两侧同源臂的荧光donor载体(即包含外源基因gfp的不同安全港位点载体)、sgrna载体和cas9载体(即质粒pkg-ge3)混合电转猪原代成纤维细胞及细胞gfp荧光强度检测
[0199]
1、共转染
[0200]
第一组(rosa26组):将质粒pb-1g 2r 3-puro-rosa26、质粒pkg-u6grna(rosa26-g3)和质粒pkg-ge3共转染猪原代成纤维细胞。配比:约20万个猪原代成纤维细胞:1.26μg质粒pb-1g 2r 3-puro-rosa26:0.82μg质粒pkg-u6grna(rosa26-g3):0.92μg质粒pkg-ge3;即3种质粒的摩尔配比依次为:1:3:1。
[0201]
第二组(aavs1组):将质粒pb-1g 2r 3-puro-aavs1、质粒pkg-u6grna(aavs1-g4)和质粒pkg-ge3共转染猪原代成纤维细胞。配比:约20万个猪原代成纤维细胞:1.26μg质粒pb-1g 2r 3-puro-aavs1:0.82μg质粒pkg-u6grna(aavs1-g4):0.92μg质粒pkg-ge3;即3种质粒的摩尔配比依次为:1:3:1。
[0202]
第三组(h11组):将质粒pb-1g 2r 3-puro-h11、质粒pkg-u6grna(h11-g1)和质粒pkg-ge3共转染猪原代成纤维细胞。配比:约20万个猪原代成纤维细胞:1.26μg质粒pb-1g 2r3-puro-h11:0.82μg质粒pkg-u6grna(h11-g1):0.92μg质粒pkg-ge3;即3种质粒的摩尔配比依次为:1:3:1。
[0203]
第四组(col1a1组):将质粒pb-1g 2r 3-puro-col1a1、质粒pkg-u6grna(col1a1-g3)和质粒pkg-ge3共转染猪原代成纤维细胞。配比:约20万个猪原代成纤维细胞:1.26μg质粒pb-1g 2r 3-puro-col1a1:0.82μg质粒pkg-u6grna(col1a1-g3):0.92μg质粒pkg-ge3;即3种质粒的摩尔配比依次为:1:3:1。
[0204]
第五组:猪原代成纤维细胞,同等电转参数不加任何质粒进行电转操作。
[0205]
共转染采用电击转染的方式,采用哺乳动物核转染试剂盒(neon kit,thermofisher)与neon tm transfection system电转仪(参数设置为:1450v、10ms、3pulse)进行转染。
[0206]
2、完成步骤1后,采用完全培养液培养12-24小时,然后更换新的完全培养液进行培养。培养总时间为48小时。
[0207]
3、完成步骤2后,更换为含1.5μg/ml嘌呤霉素的完全培养液培养3周(每2天更换新的含1.5μg/ml嘌呤霉素的完全培养液),持续观察并对gfp绿色荧光进行拍照,通过gfp荧光表达的强弱判断安全港位点表达外源基因效率的高低。
[0208]
嘌呤霉素筛选一周后,rosa26、col1a1安全港位点试验组荧光强度明显强于aavs1、h11试验组。嘌呤霉素筛选两周后,荧光强度由强到弱依次为:col1a1》rosa26》h11》aavs1,其中h11组荧光强度不太均一,rosa26组整体荧光强度较均一且荧光强度较高,aavs1组细胞荧光表达最弱,col1a1组荧光细胞数最多且荧光最强。嘌呤霉素继续筛选三周后,荧光强度由强到弱依次为:col1a1》rosa26》h11》aavs1,照片见如图2。
[0209]
五、gfp基因转录水平检测
[0210]
为了比较gfp基因整合入四个不同安全港位点后mrna转录水平的差异性,能否参与gfp的表达调控及对表达量的影响。在gfp基因外显子处设计一对引物,取步骤四中嘌呤霉素筛选三周后的细胞,提取总rna,反转录成cdna,检测原代细胞在四个不同安全港位点整合gfp基因后的转录水平,同时用第五组的细胞(无质粒的对照电转组)所得到的定量结果作为对照。以gapdh基因为内参基因按照2-δct
法进行计算。
[0211]
用于检测gfp基因的引物:f:agatccgccacaacatcgag;r:gtccatgccgagagtgatcc。
[0212]
用于检测gapdh基因的引物:f:ggtcggagtgaacggatttg;r:ccatttgatgttggcgggat。
[0213]
用spss统计学软件进行数据分析,以(平均数
±
标准差)表示,采用双因素方差分析进行统计学分析。2-δct
值结果显示嘌呤霉素筛选三周后aavs1、h11组gfp表达量较低,rosa26、col1a1组gfp表达量较高,且col1a1组和rosa26组相对于aavs1和h11组gfp转录水平差异极显著(p《0.01)。2-δct
值见表1,差异显著性分析结果见图3。
[0214]
表1 2-δct
值信息
[0215][0216]
综上,根据培养细胞三周后的荧光信号强度与gfp基因实时荧光定量pcr的结果,可以得出如下结论,在rosa26、aavs1、h11、col1a1这四个基因组安全港位点中,col1a1位点插入外源基因后表达效果最好。
[0217]
六、gfp基因的蛋白表达水平facs检测
[0218]
为了比较gfp基因整合入四个不同安全港位点后gfp蛋白的表达情况。分别用胰蛋白酶消化步骤四中嘌呤霉素筛选三周后的电转细胞,400g离心4min,弃上清。以1ml完全培养液重悬细胞,并将细胞悬液分别转移至流式管内。在bd facsmelody流式细胞仪的fitc通道内检测gfp信号,收集5
×
104个细胞进行分析,结果见图4。
[0219]
结果显示,gfp荧光信号强度col1a1》rosa26》h11》aavs1。
[0220]
因此,综合上述结果,col1a1位点是rosa26、aavs1、h11、col1a1四个安全港位点中最高效表达外源基因的猪原代细胞安全港位点。
[0221]
实施例2、制备hace2-haxl-htmprss2定点插入猪col1a1安全港位点的单细胞克隆
[0222]
人ace2基因(hace2基因)信息:编码angiotensin-converting enzyme 2[homo sapiens];位于人x号染色体;geneid为59272。angiotensin-converting enzyme 2[homo sapiens],又称为hace2蛋白。hace2蛋白是目前sars-cov-2及sars-cov等冠状病毒公认的受体蛋白,如seq id no:15所示(np_001358344.1)。
[0223]
人axl基因(haxl基因)信息:编码axl receptor tyrosine kinase[homo sapiens];位于人19号染色体;geneid为558。axl receptor tyrosine kinase[homo sapiens],又称为haxl蛋白。haxl蛋白是近期发现的不依赖于hace2蛋白的独立介导sars-cov-2感染的蛋白,如seq id no:16所示(np_001265528.1)。
[0224]
人tmprss2基因(htmprss2基因)信息:编码transmembrane protease serine 2[homo sapiens];位于人21号染色体;geneid为7113。transmembrane protease serine 2[homo sapiens],又称为htmprss2蛋白。htmprss2蛋白能够通过激活sars-cov-2的刺突蛋白(s)从而介导sars-cov-2与宿主细胞受体结合,进而促进sars-cov-2进入宿主细胞,如seq id no:17所示(np_001128571.1)。
[0225]
一、构建pkg-hace2-haxl-htmprss2 donor载体
[0226]
pkg-hace2-haxl-htmprss2 donor载体即质粒pkg-hace2-haxl-htmprss2。
[0227]
质粒pkg-hace2-haxl-htmprss2如seq id no:14所示,为环形质粒,结构示意图见图5。seq id no:14中,第9-880位核苷酸为col1a1安全港插入位点5’端猪基因组区域(sh4左臂),第887-1087位核苷酸为绝缘子1(insulator 1),第1094-2271位核苷酸为人hef1α启动子,第2290-4704位核苷酸为hace2基因,第4705-4770位核苷酸编码p2a肽,第4771-6648位核苷酸为haxl基因,第6649-6711位核苷酸编码t2a肽,第6712-8298位核苷酸为htmprss2基因,第8302-8874位核苷酸为ef1αpoly(a),第8911-8944位核苷酸为loxp序列,第8987-9316位核苷酸为sv40启动子,第9365-9961位核苷酸编码puromycin蛋白(简称puror蛋白),
第10141-10262位核苷酸为sv40 poly(a),第10307-10340位核苷酸为loxp序列,第10349-10549位核苷酸为绝缘子3(insulator 3),第10550-11276位核苷酸为col1a1安全港插入位点3’端猪基因组区域(sh4右臂)。
[0228]
二、共转染
[0229]
将质粒pkg-hace2-haxl-htmprss2、质粒pkg-u6grna(col1a1-g3)和质粒pkg-ge3共转染猪原代成纤维细胞。配比:约20万个猪原代成纤维细胞:1.3μg质粒pkg-hace2-haxl-htmprss2:0.8μg质粒pkg-u6grna(col1a1-g3):0.9μg质粒pkg-ge3;即3种质粒的摩尔配比依次为:1:3:1。
[0230]
共转染采用电击转染的方式,采用哺乳动物核转染试剂盒(neon kit,thermofisher)与neon tm transfection system电转仪(参数设置为:1450v、10ms、3pulse)。
[0231]
质粒pkg-ge3和质粒pkg-u6grna(col1a1-g3)发挥的功能为在猪基因组dna中制造dna双链断裂,以提高同源重组率。质粒pkg-hace2-haxl-htmprss2与猪基因组dna发生同源重组,在猪基因组dna中sh4左臂和sh4右臂之间插入外源靶基因片段(外源靶基因片段即seq id no:14中第881-10549位核苷酸所示的dna分子)。
[0232]
三、嘌呤霉素加压筛选
[0233]
1、嘌呤霉素筛选hace2-haxl-htmprss2基因插入的阳性细胞
[0234]
(1)完成步骤二后,采用完全培养液培养电转后的细胞16-18小时,然后更换新的完全培养液进行培养。培养总时间为48小时。
[0235]
(2)完成步骤(1)后,更换为含1.5μg/ml嘌呤霉素的完全培养液进行筛选培养(每天更换新的含1.5μg/ml嘌呤霉素的完全培养液),筛选培养的时间为3周。
[0236]
筛选培养1周时,细胞出现大量死亡。
[0237]
筛选培养2周时,细胞只有零星死亡,部分阳性克隆开始分裂增殖,细胞数不断增多。
[0238]
筛选培养第3周的目的是使细胞内质粒降解完全以排除假阳性细胞克隆。
[0239]
(3)完成步骤(2)后,收集细胞,采用不含嘌呤霉素的完全培养液恢复培养2代(每2天1代),让细胞恢复至良好状态以用于下一步的单细胞分选。
[0240]
2、单细胞分选,放大培养
[0241]
(1)完成步骤1后,收集细胞,使用胰蛋白酶进行消化,然后采用完全培养液中和,然后500g离心5min,弃除上清,将沉淀用1ml完全培养液重悬并适当稀释,用口吸管挑取单细胞转移到96孔板中(每孔预先加入100μl完全培养液)(每组细胞一个96孔板,每孔一个细胞),进行培养,培养2天后更换为含1.5μg/ml嘌呤霉素的完全培养液,之后每2~3天更换新的含1.5μg/ml嘌呤霉素的完全培养液,期间用显微镜观察每孔细胞生长情况,排除无细胞及非单细胞克隆的孔。
[0242]
(2)待步骤(1)中的96孔板的孔中细胞长满孔底(大约2周左右),使用胰蛋白酶消化并收集细胞,其中2/3细胞接种到含有完全培养液的6孔板中,剩余的1/3细胞收集在1.5ml离心管中。
[0243]
(3)待步骤(2)中的6孔板的孔中细胞长至50%丰满度时使用0.25%(gibco)的胰蛋白酶消化并收集细胞,使用细胞冻存液(90%完全培养液+10%dmso,体积比)将细胞冻
存。
[0244]
四、猪col1a1安全港位点定点插入hace2-haxl-htmprss2的基因组水平鉴定
[0245]
为了检测猪col1a1安全港位点是否成功定点插入了hace2-haxl-htmprss2。取步骤三的2的(2)中的离心管,提取细胞基因组dna,采用特异引物对(特异引物对分别为:sh4-lr-jdf1414和sh4-lr-jdr5965组成的引物对、sh4-rr-jdf282和sh4-rr-jdr4723组成的引物对、sh4-wt-jdf1085和sh4-wt-jdr1560组成的引物对)进行pcr扩增,然后进行电泳。将猪原代成纤维细胞作为野生型对照(wt)。
[0246]
sh4-lr-jdf1414和sh4-lr-jdr5965组成的引物对用来鉴定猪col1a1安全港插入位点5’端hace2-haxl-htmprss2是否重组成功(靶序列为4552bp,获得约4552bp的扩增产物表示重组成功);sh4-rr-jdf282和sh4-rr-jdr4723组成的引物对用来鉴定猪col1a1安全港插入位点3’端hace2-haxl-htmprss2是否重组成功(靶序列为4442bp,获得约4442bp的扩增产物表示重组成功);sh4-wt-jdf1085和sh4-wt-jdr1560组成的引物对用来鉴定猪col1a1安全港位点定点插入的hace2-haxl-htmprss2表达框为纯合型还是杂合型(野生型对照的基因组dna可扩增出476bp片段,重组细胞由于外源插入片段太大无法实现扩增;因此,如果不显示扩增产物,说明细胞为插入hace2-haxl-htmprss2的纯合型;如果显示476bp扩增产物,说明细胞为插入hace2-haxl-htmprss2的杂合型或者是野生型)。
[0247]
sh4-lr-jdf1414:cctgctgtaagtgccgtagt;
[0248]
sh4-lr-jdr5965:ctaggggcacagcacgtc;
[0249]
sh4-rr-jdf282:aagttattaggtctgaagaggagttt;
[0250]
sh4-rr-jdr4723:cccatcattccgtcccagag;
[0251]
sh4-wt-jdf1085:tgctgagttctggcttcctg;
[0252]
sh4-wt-jdr1560:tctaccaagagagtgaccagcag。
[0253]
根据鉴定结果,编号为1-20号的单细胞克隆均为成功在猪col1a1安全港位点定点插入hace2-haxl-htmprss2的克隆,其中10、15号单细胞克隆为纯合定点插入,其他单细胞克隆为杂合定点插入。见表2。
[0254]
表2 hace2-haxl-htmprss2定点插入猪col1a1安全港位点单细胞克隆的基因型
[0255]
[0256][0257]
表2中编号为hace2-haxl-htmprss2-1的重组细胞(杂合定点插入型)命名为1#重组细胞。
[0258]
表2中编号为hace2-haxl-htmprss2-10的重组细胞(纯合定点插入型)命名为10#重组细胞。经全基因组测序,与同一来源的猪原代成纤维细胞相比,10#重组细胞的基因组dna的差异仅在于:在猪基因组dna中sh4左臂和sh4右臂之间插入了seq id no:14中第881-10549位核苷酸所示的dna分子。
[0259]
五、单细胞克隆的hace2、haxl和htmprss2基因转录水平检测
[0260]
供试细胞:1#重组细胞和10#重组细胞。
[0261]
取供试细胞,提取总rna,反转录得到cdna。将cdna作为模板,通过荧光定量pcr检测hace2基因、haxl基因和htmprss2基因的相对表达水平(以β-actin为内参基因按照2-δct
法进行计算)。用猪原代成纤维细胞作为对照(wt)。
[0262]
荧光定量pcr的引物如下:
[0263]
hace2-f:ctaaccagccccctgtttcc;
[0264]
hace2-r:ggaggcataaggattttctccac;
[0265]
haxl-f:tcaactcctgccttctcgtg;
[0266]
haxl-r:acctctttccactgttggttca;
[0267]
htmprss2-f:tggaagttcatgggcagcaa;
[0268]
htmprss2-r:gtagaggcgaacacaccgat;
[0269]
β-actin-f:cacgccatcctgcgtctgga;
[0270]
β-actin-r:agcaccgtgttggcgtagag。
[0271]
用spss统计学软件进行数据分析,以(平均数
±
标准差)表示,采用独立样本t检验统计分析。结果见图6。2-δct
值结果显示,供试细胞的hace2基因、haxl基因和htmprss2基因的表达量均显著高于对照,且纯合定点插入的单细胞克隆比杂合定点插入的单细胞克隆表达量高。结果表明,hace2基因、haxl基因和htmprss2基因在供试细胞中均有较高程度的表达。
[0272]
六、单细胞克隆hace2、haxl和htmprss2的蛋白表达水平facs检测
[0273]
取10#重组细胞(用猪原代成纤维细胞作为对照,wt),用pbs缓冲液充分洗涤,然后用-20℃预冷的90%甲醇重悬并固定20min。固定结束后,离心,弃去固定液。加入3%bsa封闭1h。封闭结束后,离心,弃去封闭液,并用完全培养基洗涤。洗涤结束后,分别以人特异性的ace2抗体(novus biologicals,nbp2-80038)(1:100稀释)、haxl抗体(santa cruz biotechnology,sc-166269)(1:50稀释)和htmprss2抗体(santa cruz biotechnology,sc-515727)(1:50稀释)重悬细胞,室温孵育1h。抗体孵育结束后,用完全培养基充分洗涤,加入500μl完全培养基重悬细胞,并将细胞悬液转移至流式管内。在bd facsmelody流式细胞仪的相关通道内分别检测hace2、haxl和htmprss2抗体荧光信号,收集5
×
104个细胞进行分析。
[0274]
结果见图7。结果显示,10#重组细胞中检测到hace2、haxl和htmprss2的抗体荧光信号,而对照wt细胞中没有检测到hace2、haxl和htmprss2的抗体荧光信号,说明所插入的hace2基因、haxl基因和htmprss2基因在猪成纤维细胞中有显著的表达。
[0275]
实施例3、模型猪的克隆生产及所插入的目标基因在模型猪中的表达分析
[0276]
一、利用体细胞核移植技术克隆生产模型猪
[0277]
1、卵母细胞体外成熟
[0278]
从屠宰场采集新鲜的离体猪卵巢(卵巢来源于杜长大三元杂交母猪),卵巢在含75mg/ml青霉素和50mg/ml链霉素的0.9%氯化钠溶液中保存,于25-30℃温度下运输至实验室。
[0279]
取卵巢,从直径3~6mm的卵泡中抽取卵丘卵母细胞复合体(cumulus-oocyte complexes,cocs),选择至少具有三层致密卵丘细胞的cocs,接种至4孔板中,每孔装有400μl猪卵母细胞体外成熟培养基,每孔接种50个,每孔覆盖400μl的矿物油(sigma,m8410)。每次移植需培养300-400个cocs。将含cocs的培养板在38.5℃、5%co2和饱和湿度的培养箱中培养42-44小时。
[0280]
猪卵母细胞体外成熟培养基(ivm培养基):含0.1mg/ml丙酮酸、0.1mg/ml盐酸半胱氨酸、10ng/ml表皮生长因子、10%(v/v)猪卵泡液、75mg/ml青霉素、50mg/ml链霉素、10iu/ml ecg和10iu/ml hcg,余量为tcm-199培养基。
[0281]
2、体细胞核移植与胚胎移植
[0282]
(1)体细胞核移植(scnt)
[0283]
核供体细胞为实施例2获得的10#重组细胞。
[0284]
完成步骤1后,用0.1%(w/v)透明质酸酶反复吹打去除cocs的扩张卵丘细胞。将具有完整膜且含排出的第一极体的卵母细胞在含有0.1mg/ml地美可辛、0.05m蔗糖和4mg/ml牛血清白蛋白的ncsu23培养基中培养0.5-1h,促使卵母细胞核突起,然后使用尖部倾斜的显微注射针(直径约20μm)在含有10μm hepes、0.3%(w/v)聚乙烯吡咯烷酮、10%fbs、0.1mg/ml地美可辛和5mg/ml细胞松弛素b的tyrode乳酸培养基中去除突起的细胞核和极体。将单个核供体细胞注入去核卵母细胞的卵周隙,使用胚胎细胞融合仪(et3,fuj ihira industry)在融合培养基中用200v/mm的直流脉冲将核供体细胞与受体卵母细胞融合20μs。然后,将重构胚在pzm-3培养基(配方见表3)中培养2h以允许细胞核重编程,然后在激活培养基中用150v/mm的单脉冲激活100μs。然后,将激活的重构胚置于含5μg/ml细胞松弛素b的pzm-3培养基中,于38.5℃、5%co2、5%o2、90%n2和饱和湿度的培养箱中培养2小时,以进一
步激活;然后将重构胚置于pzm-3培养基中,于38.5℃、5%co2、5%o2、90%n2和饱和湿度的培养箱中培养。大部分重构胚在激活后培养6h即可用于后续的胚胎移植。
[0285]
融合培养基:含0.25m d-山梨醇、0.05mm mg(c2h3o2)2,20mg/ml bsa和0.5mm hepes[acid-free],余量为水。
[0286]
激活培养基:含0.25m d-山梨醇,0.01mm ca(c2h3o2)2,0.05mm mg(c2h3o2)2和0.1mg/ml bsa,余量为水。
[0287]
表3 pzm-3培养基的配方
[0288] 成分g/50ml1nacl0.3156g2kcl0.0373g3kh2po40.0024g4mgso4·
7h2o0.0024g5nahco30.1055g6na-pyruvate0.0011g7ca-(lactate)2·
5h2o0.0308g8myo-inositol0.02509phenol red(母液10mg/ml)0.5ml母液10l-glutamine*0.0073g11hypotaurine*0.0273g12bme essential amino acid(50
×
)*1ml13mem non-essential amino acid(100
×
)*0.5ml14超纯水补足至50ml
[0289]
*使用前添加
[0290]
(2)胚胎移植
[0291]
选择4头处于发情期的9月龄二花脸母猪二花脸母猪作为重构胚的代孕母猪,将激活后培养6h的重构胚移植到代孕母猪的输卵管中,每头母猪移植300-350个重构胚。在胚胎移植后约23天,使用超声波扫描仪(hs-101v,日本本田电子)检查妊娠情况,确认受体母猪是否怀孕。4头代孕母猪中,2头怀孕。
[0292]
克隆猪在胚胎移植后第116-117天左右出生。2头成功怀孕的母猪共生产获得5头克隆猪。克隆猪即为模型猪(照片见图8)。
[0293]
3、野生型对照猪的制备
[0294]
将同一来源的猪原代成纤维细胞代替重组细胞作为核供体细胞进行步骤2,得到4头克隆猪,即为野生型对照猪。野生型对照猪的遗传背景除不具有外源插入dna外,其余与模型猪完全一致。
[0295]
二、人源化模型猪的外源基因表达分析
[0296]
1、从猪肺脏中分离肺泡巨噬细胞
[0297]
(1)分离模型猪(或野生型对照猪)的完整气管和肺脏,并用含0.3%链霉素/青霉素的pbs缓冲液充分清洗,然后用pbs缓冲液清洗;然后向气管中灌入50ml无菌pbs缓冲液,轻轻拍打肺后倒出液体;然后向气管中灌入50ml无菌pbs缓冲液,轻轻拍打肺后倒出液体;
然后向气管中灌入50ml无菌pbs缓冲液,轻轻拍打肺后倒出液体;合并三次获得的液体,1500g离心4分钟,弃上清,收集沉淀。
[0298]
(2)加入10ml红细胞裂解液重悬步骤(1)获得的细胞沉淀,静置4分钟,然后加入20ml pbs缓冲液,1500g离心10分钟,弃上清,沉淀即为肺泡巨噬细胞。
[0299]
2、人源化猪的hace2、haxl和htmprss2基因转录水平检测
[0300]
引物和方法均同实施例2的步骤五。
[0301]
结果见图9。模型猪(hace2-haxl-htmprss2-pig)的肺泡巨噬细胞中,hace2基因的表达量为管家基因β-actin表达量的0.11倍,haxl基因的表达量为管家基因β-actin表达量的0.06倍,htmprss2基因的表达量为管家基因β-actin表达量的0.04倍,均显著高于野生型对照猪(wt-pig)肺泡巨噬细胞的相应基因的表达量。
[0302]
综上,hace2基因、haxl基因和htmrss2基因在模型猪的肺泡巨噬细胞中有较强的表达。
[0303]
3、人源化猪的hace2、haxl和htmprss2蛋白表达水平facs检测
[0304]
方法同实施例2的步骤六。
[0305]
结果见图10。结果显示,模型猪的肺泡巨噬细胞中分别检测到hace2、haxl和htmprss2的抗体荧光信号,而在野生型对照猪的肺泡巨噬细胞中没有检测到相应基因的抗体荧光信号。说明,所插入的hace2基因、haxl基因和htmprss2基因在模型猪的在肺泡巨噬细胞中被成功表达。
[0306]
实施例4、模型猪的肺泡巨噬细胞对sars-cov-2假病毒的感染性研究
[0307]
假病毒不具备复制能力,可以最大程度降低sars病毒研究过程中的各种风险。另外,由于假病毒的感染过程与真病毒相同,因此可以模拟病毒感染的早期过程,且假病毒内携带有报告基因,可以方便地进行各种检测分析。
[0308]
一、sars-cov-2-spike假病毒的制备
[0309]
1、构建重组质粒
[0310]
质粒pmd2.g-sars-c19的结构示意图见图11。初始质粒为pmd2.g商品质粒;改造过程:将pmd2.g质粒的vsv-g区域删除,并将sars-cov-2病毒的刺突蛋白(spike,为sars-cov-2的膜蛋白)进行胞内c端19个氨基酸缺失突变,然后插入已删除vsv-g区域的pmd2.g载体中。质粒pmd2.g-sars-c19如seq id no:22所示,为环形质粒。seq id no:22中,第161-540位核苷酸为cmv增强子,第541-744位核苷酸为cmv启动子,第878-1353位核苷酸为β-globin内含子,第1415-5188位核苷酸为sars-cov-2刺突蛋白(spike)的编码序列,第5264-5648位核苷酸为β-globinpoly(a)signal。
[0311]
质粒lenti-mcherry的结构示意图见图12。初始质粒为商品质粒lenti-crispr v2;改造过程:去除该质粒中的grna骨架与编码cas9蛋白的区域,并将报告基因(mcherry基因)插入相应区域,同时保留原质粒所携带的嘌呤霉素抗性基因。这将使利用该质粒联合配套质粒所构建的假病毒的基因组带有mcherry荧光标签及嘌呤霉素抗性标签。质粒lenti-mcherry如seq id no:23所示,为环形质粒。seq id no:23中,第2602-2813位核苷酸为ef1a核心启动子元件,第2844-3551位核苷酸编码mcherry荧光蛋白,第3567-3623位核苷酸编码p2a肽(p2a肽的氨基酸序列为“atnfsllkqagdveenpgp”,断裂位置为c端开始第一个氨基酸残基和第二个氨基酸残基之间),第3624-4220位核苷酸编码puromycin抗性蛋白(简称
puror蛋白),第5161-5385位核苷酸为bghpoly(a)signal。
[0312]
2、sars-cov-2-spike假病毒的制备
[0313]
pspax2慢病毒包装质粒:addgene,#12260。
[0314]
(1)将质粒pmd2.g-sars-c19、质粒lenti-mcherry与pspax2慢病毒包装质粒按6μg:4μg:5μg的比例混合,用真空浓缩仪除去水分。
[0315]
(2)完成步骤(1)后,加入750μl opti-medium,轻柔混合。
[0316]
(3)完成步骤(2)后,加入24μl lipo8000
tm
(碧云天,st483)转染试剂,轻柔混合。
[0317]
(4)hek293t细胞接种至直径为10cm的细胞培养盘中,培养至细胞密度为70%-80%。
[0318]
(5)将步骤(3)得到的混合物滴加至完成步骤(4)的细胞培养盘中培养6小时,然后更换新的完全培养液进行培养。培养总时间为48小时。
[0319]
(6)完成步骤(5)后,收集上清液,用0.45μm滤膜过滤,收集滤液。
[0320]
(7)取10ml步骤(6)得到的滤液,加入3.3ml lenti-x concentrator(clontech,631231),轻柔混合,4℃放置过夜。
[0321]
(8)完成步骤(7)后,4℃,1500g离心45分钟,弃上清,加入100μl dmem培养基溶解沉淀,即为浓缩后的sars-cov-2假病毒病毒液,简称浓缩病毒原液。
[0322]
二、假病毒感染猪肺泡巨噬细胞
[0323]
供试细胞:模型猪的肺泡巨噬细胞(实施例3中制备得到)或野生型对照猪的肺泡巨噬细胞(实施例3中制备得到)。
[0324]
(1)于感染实验24小时前,取供试细胞接种于96孔板中(3
×
104个细胞/孔)。
[0325]
(2)向96孔板细胞中加入500μl浓缩病毒原液,同时加入0.8μl polybrene试剂。
[0326]
(3)感染6小时后换液为新的完全培养液进行培养。
[0327]
(4)感染48小时后,在倒置荧光显微镜下观察细胞的mcherry荧光信号。
[0328]
结果显示,在模型猪的肺泡巨噬细胞中观察到了整合至细胞基因组中的病毒mcherry荧光信号(图13),表明sars-cov-2假病毒能够感染模型猪的肺泡巨噬细胞,而野生型对照猪的肺泡巨噬细胞中则未见有荧光信号(图14)。结果表明,模型猪的肺泡巨噬细胞具有感染sars-cov-2假病毒的能力,模型猪可作为sars-cov-2病毒感染动物模型。
[0329]
进一步的,本发明所制备的模型猪(hace2-haxl-htmprss2人源化猪)可用于下一步的药物筛选、药效评价、疫苗和抗体效果测试、病毒感染机制等生物医药领域的相关研究。
[0330]
以上对本发明进行了详述。对于本领域技术人员来说,在不脱离本发明的宗旨和范围,以及无需进行不必要的实验情况下,可在等同参数、浓度和条件下,在较宽范围内实施本发明。虽然本发明给出了特殊的实施例,应该理解为,可以对本发明作进一步的改进。总之,按本发明的原理,本技术欲包括任何变更、用途或对本发明的改进,包括脱离了本技术中已公开范围,而用本领域已知的常规技术进行的改变。按以下附带的权利要求的范围,可以进行一些基本特征的应用。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1