一种抗IL-17A的单域抗体及其用途的制作方法
一种抗il-17a的单域抗体及其用途
技术领域
1.本发明涉及能够与il-17a特异性结合的单域抗体(以下,其缩写为“il-17a单域抗体”),以及含有该单域抗体作为有效成分的药物组合物,及其药物治疗用途。
背景技术:
2.银屑病是一种较为常见的慢性炎症性皮肤病,俗称牛皮癣。该病在冬季易复发或加重,春秋季节多缓解,全球患病率约在2%~3%,其中有1/3的银屑病患者有银屑病关节炎(psoriaticarthritis,psa),常伴有关节肿痛、僵硬及运动障碍,部分可累及脊柱,严重者可造成残疾,对于患者身心健康产生了严重的影响。银屑病按照其临床特征进行分类,主要有以下几类:寻常型、关节型、脓疱型及红皮病型,90%以上的银屑病属于寻常型,该病的其他类型多由于患者在治疗期间使用外用刺激性药物、过度使用糖皮质激素以及免疫抑制剂过程中突然停药所导致。
3.银屑病发病机制尚不清楚。目前关于psa的治疗方案主要为非甾体抗炎药、糖皮质激素、抗风湿药物、硫唑嘌呤、维a酸类等,以及物理治疗、中医治疗等方法来缓解症状、控制病情。传统疗法由于疗效欠佳及不良反应,如长期使用激素会导致各种各样的副作用(血压升高、血糖升高、骨质疏松、消化道溃疡、皮肤萎缩等),临床上逐渐倾向于生物制剂的研究,肿瘤坏死因子抑制剂经常被选为psa患者的首选生物疗法,此外还有抗il-17类生物制剂。
4.银屑病以角质形成细胞过度增生、炎症细胞浸润、新生血管形成作为其组织病理改变的三要素。患者存在多种免疫细胞、免疫分子、细胞内信号传导系统等功能异常,发病有一定的潜伏期,而在该时期服用抗疟疾、抗精神病锂制剂、抗高血压的β受体阻滞剂以及血管紧张素转换酶抑制剂均会诱发该病的发生。严重影响患者的生活质量甚至身心健康。
5.银屑病的发病机制尚不清楚,目前认为它是多基因遗传背景下的自身免疫紊乱性疾病。其发病与t淋巴细胞,主要是cd4+th1淋巴细胞介导的免疫有关,致病过程包括初始t淋巴细胞活化为记忆-效应t淋巴细胞,记忆-效应t淋巴细胞进入循环移行至皮肤,聚集于病变部位,分泌多种细胞因子发挥多种生物学功能而致病。
6.il-17a是il-17细胞因子家族最早发现的成成员,主要由th17细胞分泌。1986年,mosmann、coffman及相关研究组成员发现辅助性t细胞的两个亚群-th1和th2(mosmannetal.,1986)。除了th17细胞外,还有其他t细胞(包括cd8+t细胞、γδt细胞、nkt细胞)和固有免疫细胞(包括nk细胞、ilc3细胞等)能够分泌il-17a。近年来,大量关于il-17a和th17细胞的研究成果显示il-17a参与了很多自身免疫性疾病(如类风湿性关节炎和脑脊髓炎)的病理进程,同时在抗细菌和抗真菌感染免疫中起着重要的保护作用。
7.银屑病是一种表皮细胞过度增生的自身免疫性皮肤疾病,研究表明il-17a缺陷的小鼠表皮增生减轻(rizzoetal.,2011);临床数据也显示th17细胞和il-17a在银屑病中的重要病理作用(hueberetal.,2010;wilsonetal.,2007)。基于以上关于il-17a在自身免疫性疾病中的研究成果,通过抑制th17细胞的分化和阻断il-17a信号通路来治疗自身免疫性疾病已成为当前热点。
8.银屑病因顽固难治,被列为当今世界皮肤科领域的重要研究课题,是全世界皮肤科重点防治疾病之一。随着一系列新生物药物的上市,银屑病患者有了更多更好的用药选择。在自身免疫领域中,tnfa拮抗剂和白细胞介素(il)类药物是目前研发的热点,il-12家族和il-17最受关注。
9.目前,现有技术中仍缺少亲和力强、具有药用价值的抗il-17a的单域抗体产品。
技术实现要素:
10.本专利的发明目的是提供一种能够与il-17a特异性结合的单域抗体及其用途。
11.本发明的第一方面提供了抗il-17a的单域抗体,所述的单域抗体由重链构成,重链包括seq id no:19-seq id no:25任意一条所示的重链cdr1、seq id no:26-seq id no:32任意一条所示的重链cdr2和seq id no:33-seq id no:41任意一条所示的重链cdr3。抗il-17a的单域抗体即针对il-17a的单域抗体。
12.优选地,所述的重链cdr1、重链cdr2和重链cdr3的氨基酸序列为下述(1)-(9)的一种:(1)seq id no:19所示的cdr1,seq id no:28所示的cdr2,seq id no:36所示的cdr3;(2)seq id no:21所示的cdr1,seq id no:29所示的cdr2,seq id no:35所示的cdr3;(3)seq id no:25所示的cdr1,seq id no:26所示的cdr2,seq id no:41所示的cdr3;(4)seq id no:23所示的cdr1,seq id no:27所示的cdr2,seq id no:37所示的cdr3;(5)seq id no:25所示的cdr1,seq id no:26所示的cdr2,seq id no:40所示的cdr3;(6)seq id no:20所示的cdr1,seq id no:30所示的cdr2,seq id no:33所示的cdr3;(7)seq id no:25所示的cdr1,seq id no:27所示的cdr2,seq id no:38所示的cdr3;(8)seq id no:24所示的cdr1,seq id no:31所示的cdr2,seq id no:34所示的cdr3;(9)seq id no:22所示的cdr1,seq id no:32所示的cdr2,seq id no:39所示的cdr3。
13.以上9种cdr组合(1)-(9)依次与seq id no.1-9对应。
14.上述所有序列,可以替换为与该序列具有“至少80%同源性”的序列或仅一个或少数几个氨基酸替换的序列;优选为“至少85%同源性”,更优选为“至少90%同源性”,更优选为“至少95%同源性”,最优选为“至少98%同源性”。
15.在一个实施方案中,其中所述重链cdr1、cdr2和cdr3的任何一个或多个cdr中,一个至五个任意氨基酸残基可以分别用其保守氨基酸取代。具体地,所述重链cdr1中,1至5个氨基酸残基可由其保守氨基酸替换;所述重链cdr2中,1至5个氨基酸残基可由其保守氨基
酸替换;所述重链cdr3中,1至5个氨基酸残基可由其保守氨基酸替换。
16.如本文所用,术语“序列同源性”表示两个(核苷酸或氨基酸)序列在比对中在相同位置处具有相同残基的程度,并且通常表示为百分数。优选地,同源性在被比较的序列的整体长度上确定。因此,具有完全相同序列的两个拷贝具有100%同源性。
17.在一些实施例中,与前述序列相比仅仅替换一个或少数几个氨基酸的序列,例如,包含1、2、3、4、5、6、7、8、9或10个保守氨基酸取代,也可以实现发明目的。这些变异形式包括(但并不限于):一个或多个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个)氨基酸的缺失、插入和/或取代,以及在c末端和/或n末端添加一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。实际上,在确定两个氨基酸序列之间的序列同源性程度或在确定单域抗体中的cdr1、cdr2和cdr3组合时,技术人员可以考虑所谓的“保守”氨基酸取代,在取代情况下,所述取代将优选为保守氨基酸取代。所述保守氨基酸,其通常可以被描述为氨基酸残基被具有相似化学结构的另一个氨基酸残基替代的氨基酸取代,并且该取代对多肽的功能、活性或其它生物学性质几乎没有或基本上没有影响。所述保守氨基酸取代在本领域是通用的,例如保守氨基酸取代是以下(a)-(d)组内的一个或少数几个氨基酸被同一组内另一个或少数几个氨基酸所取代:(a)极性带负电残基及其不带电酰胺:asp、asn、glu、gln;(b)极性带正电残基:his、arg、lys;(c)芳香族残基:phe、trp、tyr;(d)脂肪族非极性或弱极性残基:ala、ser、thr、gly、pro、met、leu、ile、val、cys。特别优选的保守氨基酸取代如下:asp被glu取代;asn被gln或his取代;glu被asp取代;gln被asn取代;his被asn或gln取代;arg被lys取代;lys被arg、gln取代;phe被met、leu、tyr取代;trp被tyr取代;tyr被phe、trp取代;ala被gly或ser取代;ser被thr取代;thr被ser取代;gly被ala或pro取代;met被leu、tyr或ile取代;leu被ile或val取代;ile被leu或val取代;val被ile或leu取代;cys被ser取代。另外,本领域技术人员知晓,单域抗体的创造性体现在cdr1-3区,而框架区序列fr1-4并非是不可改变的,fr1-4的序列可以采取本发明公开的序列的保守序列变体。
18.本发明中的“抗il-17a的单域抗体”的含义,不仅包括完整的单域抗体,还包括所述抗il-17a的单域抗体的片段、衍生物和类似物。如本文所用,术语
ꢀ“
片段”、“衍生物”和“类似物”含义相同,均是指基本上保持本发明抗体相同的生物学功能或活性的多肽。本发明的多肽片段、衍生物或类似物可以是(i)有一个或多个保守或非保守性氨基酸残基(优选保守性氨基酸残基)被取代的多肽,而这样的取代的氨基酸残基可以是也可以不是由遗传密码编码的,或(ii)在一个或多个氨基酸残基中具有取代基团的多肽,或(iii)成熟多肽与另一个化合物(比如延长多肽半衰期的化合物,例如聚乙二醇)融合所形成的多肽,或(iv)附加的氨基酸序列融合到此多肽序列而形成的多肽(如前导序列或分泌序列或用来纯化此多肽的序列或蛋白原序列,或与fc标签形成的融合蛋白)。根据本文的教导,这些片段、衍生物和类似物属于本领域熟练技术人员公知的范围。
19.在一个优选实施方案中,所述的抗体序列还包括框架区fr;所述框架区fr包括fr1、fr2、fr3和fr4的氨基酸序列;框架区fr的氨基酸序列分别为:seq id no:42-44中的任意一条所示的fr1或fr1的变体,所述fr1的变体在所述fr1中包含至多5个氨基酸的替换;seq id no:45-52中的任意一条所示的fr2或fr2的变体,所述fr2的变体在所述
fr2中包含至多5个氨基酸的替换;seq id no:53-59中的任意一条所示的fr3或fr3的变体,所述fr3的变体在所述fr3中包含至多5个氨基酸的替换;seq id no:60-62中的任意一条所示的fr4或fr4的变体,所述fr4的变体在所述fr4中包含至多5个氨基酸的替换。
20.本发明的第二方面是提供抗il-17a的单域抗体的氨基酸序列,所述单域抗体的氨基酸序列分别如seq id no.1-9所示,或者所述单域抗体与seq id no.1-9 的氨基酸序列具有至少80%序列同源性,并且能够特异性结合il-17a蛋白。
21.在一个实施方案中,所述抗il-17a的单域抗体与选自seq id no:1-9的氨基酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%序列同源性,并且能够特异性结合il-17a蛋白。
22.本发明的第三个方面是提供一种双特异性抗体,所述双特异性抗体包括第一抗原结合部分和第二抗原结合部分,第一抗原结合部分为单域抗体,所述的单域抗体的氨基酸序列分别如seq id no.1-9所示或与seq id no.1-9具有至少80%同源性。第二抗原结合部分为另一种抗体,例如可以为针对tnfα、il-6r、il-6、il-23、il-23r的抗体(可以为单抗、多抗、单域抗体或者抗体的其它任意形式);第二抗原结合部分也可以为针对其它抗原的抗体,以应用于疾病的诊断、预防或治疗或者抗原的检测。
23.本发明的第四个方面是提供前述任一的抗il-17a的单域抗体在制备双特异性抗体中的用途。
24.本发明的第五个方面是提供前述任一的抗 il-17a的单域抗体的fc融合抗体或人源化抗体。
25.本发明的第六个方面是提供编码前述的抗il-17a的单域抗体或前述的fc融合抗体或前述的人源化抗体的核苷酸分子,其核苷酸序列分别如seq id no:10-18所示,或者与seq id no:10-18 中的任意一条具有至少95%序列同源性。
26.在一个实施方案中,编码所述抗il-17a的单域抗体的核酸分子与选自seq id no:10-18的核苷酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%序列同源性,并且其编码的抗il-17a的单域抗体能够特异性结合il-17a蛋白。
27.本发明的第七个方面是提供一种表达载体,其包含编码抗il-17a的单域抗体或fc融合抗体或人源化抗体的核苷酸分子,其核苷酸序列分别如seq id no:10-18所示。
28.在一个优选的实施方案中,所使用的表达载体为rjk-v4-3(通过基因工程手段将编码抗il-17a的单域抗体或其fc融合抗体或人源化抗体的核苷酸分子整合入rjk-v4-3中),根据需要还可以选择其他的通用型表达载体。
29.本发明的第八个方面是提供一种能够表达前述的抗il-17a的单域抗体、fc融合抗体或人源化抗体的宿主细胞,或其包含前述的表达载体。优选宿主细胞为细菌细胞、真菌细胞或哺乳动物细胞。
30.在另一个优选的实施方案中,所述的宿主细胞包括原核细胞或真核细胞,包括细菌,真菌。
31.在另一个优选的实施方案中,所述的宿主细胞选自下组:大肠杆菌、酵母细胞、哺乳动物细胞、噬菌体、或其组合。
32.在另一个优选的实施方案中,所述原核细胞选自下组:大肠杆菌、枯草杆菌、乳酸菌、链霉菌、奇异变形菌、或其组合。
33.在另一个优选的实施方案中,所述真核细胞选自下组:巴斯德毕赤酵母、酿酒酵母、裂殖酵母、木霉、或其组合。
34.在另一个优选的实施方案中,所述真核细胞选自下组:草地粘虫等昆虫细胞、烟草等植物细胞、bhk细胞、cho细胞、cos细胞、骨髓瘤细胞、或其组合。
35.在另一个优选的实施方案中,所述宿主细胞为悬浮expicho-s细胞。
36.在另一个优选的实施方案中,所述宿主细胞为悬浮293f细胞。
37.本发明的第九个方面是提供一种重组蛋白,包含前述的抗il-17a的单域抗体。所述的重组蛋白可以为前述的seq id no.1-9所示的单域抗体,也可以为与seq id no.1-9具有至少80%同源性的单域抗体,还可以是多表位抗体、多特异性抗体以及多价抗体;例如,所述的多表位抗体可由seq id no.1-9中的不止一条序列组成;所述的多价抗体可由seq id no.1-9中的其中一条序列重复排列若干次组成;所述的多特异性抗体包括但不限于前述的双特异性抗体,以及三特异性抗体;此外,重组蛋白可以为前述的抗体的片段、衍生物和类似物。
38.本发明的第十个方面是提供一种药物组合物,其包含前述的抗il-17a的单域抗体和药学上可接受的载体。通常,可将这些物质配制于无毒的、惰性的和药学上可接受的水性载体介质中,其中ph通常根据抗体的等电点确定(水性载体介质的ph需偏离该抗体的等电点,且与该抗体的等电点相差大约2)。配制好的药物组合物可以通过常规途径进行给药,其中包括(但并不限于):静脉内、透皮(在患处直接涂抹或贴敷膏药)。
39.本发明的药物组合物可直接用于结合il-17a蛋白分子,因而可用于治疗银屑病。此外,还可同时使用其他银屑病治疗剂联合使用。
40.本发明的药物组合物含有安全有效量(如0.001-99wt%,较佳地0.01-90wt%,更佳地0.1-80wt%)的前述的单域抗体以及药学上可接受的载体或赋形剂。这类载体包括(但并不限于):盐水、缓冲液、葡萄糖、水、甘油、乙醇、及其组合。药物制剂应与给药方式相匹配。本发明的药物组合物可以被制成针剂形式,例如用生理盐水或含有葡萄糖和其他辅剂的水溶液通过常规方法进行制备。药物组合物如针剂、溶液宜在无菌条件下制造。
41.本发明的第十一个方面是提供一种用于治疗银屑病的药剂,其包含前述的抗il-17a的单域抗体作为活性成分。
42.本发明的第十二个方面是提供一种检测il-17a水平的试剂盒,其含有前述的抗 il-17a的单域抗体。在本发明的一个优选例中,所述的试剂盒还包括容器、使用说明书、缓冲剂等。
43.在一个优选的实施方案中,该试剂盒包括识别il-17a蛋白的抗体,用于溶解样本的裂解介质,检测所需的通用试剂和缓冲液,如各种缓冲液、检测标记、检测底物等。该检测试剂盒可以是体外诊断装置。
44.在一个优选的实施方案中,所述的试剂盒还含有第二抗体和用于检测的酶或荧光或放射标记物,以及缓冲液。
45.在一个优选的实施方案中,所述试剂盒的第二抗体可以为前述的抗il-17a的单域抗体的抗体(作为抗抗体),可以为单域抗体、单克隆抗体、多克隆抗体或抗体的其它任意形
式。
46.本发明的第十三个方面,提供了一种产生抗il-17a的单域抗体的方法,包括步骤:(a)在适合产生单域抗体的条件下,培养本发明的第八个方面所述的宿主细胞,从而获得含所述抗il-17a的单域抗体的培养物;以及(b)从所述培养物中分离或回收所述的抗il-17a的单域抗体;以及(c)任选地,纯化和/或修饰步骤(b)中获得的il-17a的单域抗体。
47.本发明的第十四个方面是提供针对前述的抗il-17a的单域抗体或药物组合物在制备抑制il-17a基因表达的药物或抗银屑病药物中的用途。
48.本发明的第十五个方面是提供前述的抗il-17a的单域抗体或前述药物组合物在制备用于治疗疾病的药物中的用途。
49.在一个优选的实施方案中,所述疾病为自身免疫性疾病,或者所述疾病为与il-17a异常表达相关的病症。
50.在一个优选的实施方案中,所述疾病为银屑病。
51.在一个优选的实施方案中,所述疾病为关节炎。
52.本发明的第十六个方面是提供针对前述的抗il-17a的单域抗体在制备阻断il-17a与il-17r结合的药物中的用途。
53.有益效果相对于现有技术,本发明的有益效果是:(1)本发明单域抗体特异性针对具有正确空间结构的il-17a蛋白。
54.(2)本发明单域抗体具有较好的特异性,能够阻断il-17a与il-17r结合部位的相互作用,且具有高于现有药物的阻断活性,在制备治疗自身免疫性疾病的药物中具有很大的应用前景。
55.(3)本发明得到的单域抗体,表达体系选择灵活,既可以在原核系统中表达也可以在酵母细胞或哺乳动物细胞的真核系统中表达,且其在原核表达系统中的表达成本低,可降低后期生产成本。
56.(4)本发明得到的单域抗体,其抗体的多组合形式改造简单,通过基因工程的方式简单的串联即可以获得多价、多特异性的抗体,并且其免疫异质性很低,在不经过人源化改造的情况下也不会产生较强的免疫反应。
57.(5)本发明得到单域抗体,其亲和力范围更为宽泛,在未进行亲和力成熟之前,其亲和力范围可以从nm级别至pm级别,为后期不同用途的抗体提供多个选择。
附图说明
58.为了更清楚地说明本技术的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,显而易见地,对于本领域普通技术人员而言,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
59.图1为实施例3中靶向il-17a抗体筛选的文库富集情况;图2为实施例13中的抗体抗原结合量效曲线测定图(一部分克隆);图3为实施例13中的抗体抗原结合量效曲线测定图(另一部分克隆、tab1、higg);
图4为实施例14中的抗体阻断il-17a与il-17r结合的测定图(一部分克隆);图5为实施例14中的抗体阻断il-17a与il-17r结合的测定图(另一部分克隆、higg);图6为实施例15中抗体中和人源il-17a(真核样品)诱导hela细胞释放il-6实验结果图谱(一部分克隆、tab、higg);图7为实施例15中抗体中和人源il-17a(真核样品)诱导hela细胞释放il-6实验结果图谱(另一部分克隆);图8为实施例16中抗体中和人源il-17a(人源化样品)诱导hela细胞释放il-6实验结果图谱(tab、higg、2g3);图9为实施例16中抗体中和人源il-17a(人源化样品)诱导hela细胞释放il-6实验结果图谱(2g3、1a10);图10为实施例16中抗体中和人源il-17a(人源化样品)诱导hela细胞释放il-6实验结果图谱(1a10)。
具体实施方式
60.下面结合实施例对本发明做进一步的详细说明,以令本领域技术人员参照说明书文字能够据以实施。
61.如本文所用,“单域抗体”(sdab,也被研发者ablynx称为纳米抗体或vhh)是本领域技术人员公知的。单域抗体为其互补决定区是单域多肽的一部分的抗体。因此,单域抗体包含单个互补决定区(单个cdr1、单个cdr2和单个cdr3)。单域抗体的实例为仅有重链的抗体(该抗体天然不包含轻链)、衍生自常规抗体的单域抗体和工程化抗体。
62.单域抗体可以衍生自任何物种,包括小鼠、人、骆驼、美洲驼、山羊、兔和牛。例如,天然存在的vhh分子可以衍生自骆驼科物种(例如骆驼、单峰骆驼、美洲驼和原驼)提供的抗体。像完整的抗体一样,单域抗体能够选择性地结合特定抗原。单域抗体可以仅含有免疫球蛋白链的可变结构域,该结构域具有cdr1、cdr2和cdr3以及框架区。
63.如本文所用,术语“序列同源性”表示两个(核苷酸或氨基酸)序列在比对中在相同位置处具有相同残基的程度,并且通常表示为百分数。优选地,同源性在被比较的序列的整体长度上确定。因此,具有完全相同序列的两个拷贝具有100%同源性。
64.如本文所用,术语“fc融合抗体”指利用基因工程技术将目标抗体的fc段与某种具有生物学活性的功能蛋白分子融合而产生的新型蛋白。
65.术语“人源化抗体”是指将目标抗体(如动物抗体)的重链可变区与人抗体的恒定区融合而得到的抗体,或将目标抗体的互补决定区(cdr1~3序列)移植至人抗体的可变区内,得到的抗体,或将目标抗体根据人抗体骨架区(fr1~4)的特征进行氨基酸的突变后获得的抗体。人源化抗体可用合成法或定点突变法。
66.本发明中,与本发明公开的cdr1-3的序列同源性高的序列,也可以得到针对il-17a 的单域抗体。在一些实施例中,与seq id no.1-9中的序列具有“至少80%同源性”的序列,或者“至少85%同源性”、“至少90%同源性”、“至少95%同源性”、“至少98%同源性”的序列都可以实现发明目的。
67.在一些实施例中,与seq id no:1-9中的序列相比仅仅替换一个或少数几个氨基
酸的序列,例如,包含1、2、3、4、5、6、7、8、9或10个保守氨基酸取代,也可以实现发明目的。实际上,在确定两个氨基酸序列之间的序列同源性程度或在确定单域抗体中的cdr1、cdr2和cdr3组合时,技术人员可以考虑所谓的“保守”氨基酸取代,在取代情况下,所述取代将优选为保守氨基酸取代,所述保守氨基酸,其通常可以被描述为氨基酸残基被具有相似化学结构的另一个氨基酸残基替代的氨基酸取代,并且该取代对多肽的功能、活性或其它生物学性质几乎没有或基本上没有影响。所述保守氨基酸取代在本领域是通用的,例如保守氨基酸取代是以下(a)-(d)组内的一个或少数几个氨基酸被同一组内另一个或少数几个氨基酸所取代:(a)极性带负电残基及其不带电酰胺:asp、asn、glu、gln;(b)极性带正电残基:his、arg、lys;(c)芳香族残基:phe、trp、tyr;(d)脂肪族非极性或弱极性残基:ala、ser、thr、gly、pro、met、leu、ile、val、cys。特别优选的保守氨基酸取代如下:asp被glu取代;asn被gln或his取代;glu被asp取代;gln被asn取代;his被asn或gln取代;arg被lys取代;lys被arg、gln取代;phe被met、leu、tyr取代;trp被tyr取代;tyr被phe、trp取代;ala被gly或ser取代;ser被thr取代;thr被ser取代;gly被ala或pro取代;met被leu、tyr或ile取代;leu被ile或val取代;ile被leu或val取代;val被ile或leu取代;cys被ser取代。另外,本领域技术人员知晓,单域抗体的创造性体现在cdr1-3区,而框架区序列fr1-4并非是不可改变的,fr1-4的序列可以采取本发明公开的序列的保守序列变体。
68.本发明的优选宿主细胞为细菌细胞、真菌细胞或哺乳动物细胞。
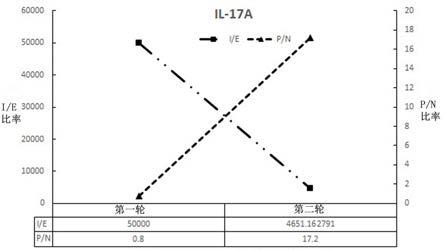
69.本专利是通过基因工程技术制备目标蛋白以及目标蛋白的截短体形式,然后将获得的抗原蛋白免疫内蒙古阿拉善双峰驼,通过多次免疫后,获得骆驼的外周血淋巴细胞或者脾细胞,通过基因工程的方式,将驼源抗体可变区编码序列重组到噬菌体展示载体中,通过噬菌体展示技术筛选出针对抗原蛋白的特异性抗体,并进一步检测其与抗原结合的能力以及在包括自身免疫疾病治疗中的应用。
70.现将上述技术方案拆分详解,以具体实施例的方式描述:实施例1:人源il-17a重组胞外结构域蛋白的制备:本专利中用到的人源重组胞外结构域蛋白为公司自己表达纯化获得,人源il-17a重组胞外结构域蛋白表达载体设计方案具体如下:(1)在ncbi中检索获得il-17a的编码序列,其收录号为nm_002190.2,该序列编码产生的氨基酸序列登录号为np_002181.1,uniprot id为q16552。
71.(2)分别通过tmhmm和smart网站对np_002181.1对应的氨基酸序列进行蛋白跨膜区和胞外端的分析。
72.(3)分析结果显示il-17a为分泌型蛋白,1
‑ꢀ
23位为该蛋白的信号肽。
73.(4)利用基因合成的方式将编码il-17a蛋白的24-155位氨基酸的核苷酸序列克隆到载体pcdna3.4中。
74.(5)将构建好的载体进行sanger测序,比对原始序列,确认无误后,将该重组质粒进行批量抽提,去除内毒素,转染悬浮293f进行目的蛋白的表达、纯化,纯化后的蛋白纯度高达90%,满足动物免疫的需求。
75.实施例2:针对il-17a蛋白的单域抗体文库的构建:将1mg实施例1中纯化获得的人源重组il-17a蛋白与等体积的弗氏完全佐剂混合,免疫一只内蒙古阿拉善双峰驼,每周免疫一次,共连续免疫7次,除首次免疫外,其余六次均
是用1mg il-17a蛋白与弗氏不完全佐剂等体积混合进行动物免疫,该免疫过程是为了集中刺激骆驼使其产生针对il-17a蛋白的抗体。
76.动物免疫结束后,抽取骆驼外周血淋巴细胞150ml,并提取细胞的rna。利用提取的总rna合成cdna,并通过套式pcr反应以cdna为模板扩增vhh(抗体重链可变区)。
77.然后利用限制性内切酶分别酶切pmecs载体和vhh片段,然后将酶切后的片段和载体链接。将连接后的片段电转化至感受态细胞tg1中,构建il-17a蛋白的噬菌体展示文库并测定库容,文库的库容大小约为1
×
109,同时,通过菌落pcr鉴定检测文库在目的片段的正确插入率。
78.结果显示,从文库中随机挑选的40个菌落进行pcr扩增后,有38个克隆可以扩增出大小为预测大小的条带,有2个克隆扩增出条带不正确,故正确插入率为38
÷ꢀ
40
×
100%≈ 95%。
79.实施例3:针对il-17a蛋白的单域抗体筛选:取200μl实施例2中的重组tg1细胞至2
×
ty培养基中培养,期间加入40μl辅助噬菌体vcsm13侵染tg1细胞,并培养过夜以扩增噬菌体,次日利用peg/nacl沉淀噬菌体,离心收集扩增噬菌体。
80.将稀释在100 mm ph8.3的nahco3中的il-17a蛋白500μg偶联在酶标板上,4℃放置过夜,同时设立阴性对照孔(培养基对照);第二天加入200μl的3%的脱脂乳,室温封闭2h;封闭结束后,加入100μl扩增后噬菌体文库(大约2
×
10
11
个噬菌体颗粒),室温作用1h;作用1小时后,用pbs+0.05%tween-20洗15遍,以洗掉未结合的噬菌体。
81.用终浓度为25mg/ml的胰蛋白酶将与il-17a蛋白特异性结合的噬菌体解离下,并感染处于对数生长期的大肠杆菌tg1细胞,37℃培养1h,产生并收集噬菌体用于下一轮的筛选,相同筛选过程重复1轮,逐步得到富集。
82.当富集倍数达到10倍以上时,富集效果如图1所示。
83.图1中,p/n=生物淘选中阳性孔洗脱下的噬菌体感染tg1细菌后生长的单克隆细菌数/阳性孔洗脱下的噬菌体感染tg1细菌后生长的单克隆细菌数,该参数在富集发生后会逐渐增大;i/e=生物淘选中每轮加入阳性孔的噬菌体总量/生物淘选中每轮从阳性孔洗脱出的噬菌体总量,该参数在富集发生后会逐渐趋近于1。
84.实施例4:用噬菌体的酶联免疫方法(elisa)筛选针对il-17a的特异性阳性克隆:根据上述实施例3中的筛选方法对抗il-17a蛋白的单域抗体进行3轮筛选,抗il-17a蛋白的噬菌体富集因子达到10以上,筛选结束后,从筛选获得的阳性克隆中挑选384个单菌落分别接种于含100 μg/ml氨苄青霉素的2
×
ty培养基的96深孔板中,并设置空白对照,37℃培养至对数期后,加入终浓度为1 mm的iptg,28℃培养过夜。
85.利用渗透涨破法获得粗提抗体;将il-17a重组蛋白分别释至100 mm ph8.3的nahco3中并将100μg蛋白在酶标板(elisa板)中4℃包被过夜。将获得的抗体粗提液取100μl转移至加入抗原的elisa板上,室温孵育1h;用pbst洗去未结合的抗体,加入100μl经1:2000稀释后的mouse anti-ha tag antibody(hrp)(鼠抗ha辣根过氧化物酶标记抗体,thermo fisher),在室温孵育1h;用pbst洗去未结合的抗体,加入辣根过氧化物酶显色液,37℃下反应15min后,加入终止液,于酶标仪上450nm波长处,读取吸收值。
86.当样品孔od值大于对照孔5倍以上时,判定为阳性克隆孔;将阳性克隆孔的菌转摇
在含有100μg/ml氨苄青霉素的lb培养基中以便提取质粒并进行测序。
87.根据序列比对软件vectornti分析各个克隆株的基因序列,把cdr1,cdr2和cdr3序列相同的株视为同一克隆株,而序列不同的株视为不同克隆株,最终获得特异性针对il-17a蛋白的单域抗体(seqidno.1-9以及未示出序列的其它单域抗体,未示出序列的其它单域抗体包括附图中的1c9、1d1等)。
88.其抗体的氨基酸序列为fr1-cdr1-fr2-cdr2-fr3-cdr3-fr4结构,构成整个vhh。获得的单域抗体重组质粒可以在原核系统中进行表达,最终获得单域抗体蛋白。
89.9种单域抗体的cdr、fr序列如表1-6所示。9种单域抗体的氨基酸序列分别如seqidno.1-9所示,编码9种单域抗体的dna序列分别如seqidno.10-18所示。
90.表19种单域抗体的cdr1序列表29种单域抗体的cdr2序列表39种单域抗体的cdr3序列
表49种单域抗体的fr1序列表59种单域抗体的fr2序列表69种单域抗体的fr3序列
表79种单域抗体的fr4序列单域抗体的氨基酸序列seqidno.1-9依次与单域抗体1a10,1d7,1e6,2e2,2g3,3d1,3e1,3h2,4a12一一对应。单域抗体的核苷酸序列seqidno.10-18也依次与单域抗体1a10,1d7,1e6,2e2,2g3,3d1,3e1,3h2,4a12一一对应。
91.实施例5:il-17a蛋白的特异性单域抗体在宿主菌大肠杆菌中的纯化及表达将实施例4中测序分析所获得不同克隆株的质粒(pmecs-vhh)电转化到大肠杆菌hb2151中,并将其涂布在lb+amp+glucose即含有氨苄青霉素和葡萄糖的培养平板上,37℃培养过夜;挑选单个菌落接种在5ml含有氨苄青霉素的lb培养液中,37℃摇床培养过夜。
92.接种1ml的过夜培养菌种至330mltb培养液中,37℃摇床培养,培养到od600nm值达到0.6-0.9时,加入1miptg,28℃摇床培养过夜;离心,收集大肠杆菌,利用渗透涨破法,获得抗体粗提液;通过镍柱亲和层析法纯化出抗体,纯化后部分的单域抗体,包括vhh1~9。
93.vhh1-9依次对应氨基酸序列seqidno.1-9的单域抗体,即vhh1-9分别对应:1a10,1d7,1e6,2e2,2g3,3d1,3e1,3h2,4a12。
94.实施例6:针对il-17a的单域抗体的人源化人源化方法采用对基于大数据分析结果而构建的抗体框架区域突变文库进行高通量筛选的方法完成。详细步骤为:(1)人源/驼源抗体数据的序列分析:对从ncbi网站批量下载的13873例nb(human)
序列进行氨基酸偏好性分析,同时对本公司2000例纳米抗体序列进行氨基酸偏好性分析,得到框架区各个位点氨基端比例数据;(2)人源驼源综合加权分析:将上述源/驼源抗体序列统一按照imgt编号规则进行编号并一一对应,结合上述两个物种中氨基酸比例分析结果,按照人源90%驼源10%的权重进行加权分析,统计出各个位点氨基酸加权后的比例,并由高到低排序;根据最终的加权结果,框架区单个位点仅保留所占比例》10%的氨基酸种类,并根据保留后比例综合为1的标准进行所占比例 》10% 氨基酸最终权重的计算,作为后续氨基酸定制文库的设计依据;(3)氨基酸定制文库的方案设计:对待进行突变的单独位点,规定其》10%的氨基酸数目为n, 规定其》10%中所占比例最高值与最低值的比值为v,对待进行突变的位点进行性质判断:若v≥3且n≤2 则认为该位点属于“高集中度位点”,否则认为该位点属于“中低集中度位点”。依据此方法将定制氨基酸文库分为“高/中低集中度文库”两个分别进行氨基酸定制文库的构建,上述(2)中的最终权重即为文库中位点氨基酸种类及比例的参考依据。
95.(4)氨基酸定制文库的高通量筛选:对抗体株,1a10,1d7,1e6,2e2,2g3,3d1,3e1,3h2, 4a12分别构建人源化抗体文库,针对构建好的文库,分别用对应抗原进行淘选,并最终获得亲和力较高且人源化程度较高的抗体序列。
96.例如可以得到以下人源化抗体:对2g3单域抗体进行人源化改造所得的8种人源化抗体(2g3-v17、2g3-v18、2g3-v24、2g3-v25、2g3-v46、2g3-v50、2g3-v31、2g3-v34,此8种2g3的人源化抗体的序列均不完全相同)、对1a10单域抗体进行人源化改造所得的8种人源化抗体(1a10-v1、1a10-v4、1a10-v9、1a10-v10、1a10-v5、1a10-v6、1a10-v15或1a10-v14,此8种1a10的人源化抗体的序列均不完全相同)。
97.实施例7:抗il-17a的单域抗体的fc融合抗体真核表达载体的构建(1)将实施例4中获得的目标序列亚克隆至真核表达载体中:将实施例4中筛选出来的抗体经sanger测序得到其核苷酸序列;(2)通过序列合成的方式将上述核苷酸序列(seq id no:10-18)合成至本公司设计改造的载体rjk-v4-3中得到重组真核表达载体,该载体的改造方法如实施例11所述;(3)将步骤(2)构建好的重组真核表达载体转化至dh5α大肠杆菌中,培养进行质粒抽提,去除内毒素;(4)将抽提后的质粒再进行序列测序鉴定;(5)将确定无误后的重组载体准备后续真核细胞转染表达,通过实施例8或9的方法将vhh的fc蛋白表达后并通过实施例10的方法纯化上述抗体。
98.实施例8:抗il-17a蛋白的单域抗体在悬浮expicho-s细胞中表达(1)转染前3天以2.5
×
105/ml细胞传代和扩大培养expicho-s
™
细胞,计算出的所需的细胞体积转移至装有新鲜的已预热的120ml(终体积)的expicho
™
表达培养基的500ml摇瓶中;使细胞浓度达到约4
×
106ꢀ‑6×
106活细胞/ ml;(2)在转染前一天,将expicho-s
™
细胞稀释浓度至3.5
×
106活细胞/ ml,使细胞过夜培养;(3)转染当天,测定细胞密度和活细胞百分比。转染之前细胞密度应达到约7
×
10
6 ‑
10
×
106活细胞/ ml;
(4)用预热至37℃新鲜的expicho
™
表达培养基将细胞稀释至6
×
106个活细胞/ ml。计算出的所需的细胞体积转移至装有新鲜的已预热的100ml(终体积)的expicho
™
表达培养基的500ml摇瓶中;(5)使轻轻颠倒混匀expifectamine
™
cho试剂,用3.7ml optipro
™
培养基稀释expifectamine
™
cho试剂,回荡或混匀;(6)用冷藏的4ml optipro
™
培养基稀释质粒dna,回荡混匀;(7)将expifectamine cho /质粒dna(质粒dna为实施例7制得的抗il-17a的单域抗体的fc融合抗体真核表达载体)复合物室温孵育1-5分钟,然后轻轻加入制备的细胞悬液中,加入过程中轻轻回荡摇瓶;(8)将细胞在37
°
c、 8%co2、加湿的空气中震荡培养;(9)转染后第1天(18-22小时后)添加600ul expifectamine
™
cho enhancer和24ml expicho feed。
99.(10)在转染后约8天(细胞活率低于70%)收集上清。
100.实施例9:抗il-17a蛋白的单域抗体在悬浮293f细胞中的表达重组单域抗体表达实验流程(以500ml摇瓶为例):(1)转染前3天以2.5
×
105/ml细胞传代和扩大培养293f细胞,计算出的所需的细胞体积转移至装有新鲜的已预热的120ml(终体积)的opm-293 cd05 medium培养基的500ml摇瓶中。使细胞浓度达到约2
×
10
6-3
×
106活细胞/ ml。
101.(2)转染当天,测定细胞密度和活细胞百分比。转染之前细胞密度应达到约2
×
10
6-3
×
106活细胞/ ml。
102.(3)用预热的opm-293 cd05 medium将细胞稀释至1
×
106个活细胞/ ml。计算出所需的细胞体积转移至装有新鲜的已预热的100ml(终体积)的培养基的500ml摇瓶中。
103.(4)用4ml opti-mem培养基稀释pei(1mg/ml)试剂,回荡或吹打混匀;用4ml opt-mem培养基稀释质粒dna(质粒dna为实施例7制得的抗il-17a的单域抗体的fc融合抗体真核表达载体),回荡混匀,并用0.22um的滤头过滤。室温孵育5min。
104.(5)将稀释的pei试剂加入稀释的dna中,颠倒混匀。将pei/质粒dna复合物室温孵育15-20分钟,然后轻轻加入制备的细胞悬液中,加入过程中轻轻回荡摇瓶。
105.(6)将细胞在37
°
c、 5%co2、120rpm震荡培养。
106.(7)转染后第24h、72h添加5ml opm-cho pff05补料。
107.(8)在转染后约7天(细胞活率低于70%)收集上清。
108.实施例10:抗il-17a蛋白的单域抗体的纯化(1)将实施例8或9中获得蛋白表达上清用0.45μm的一次性滤头过滤除掉不可溶杂质;(2)将上述滤液使用蛋白纯化仪进行亲和层析纯化,利用人源fc与protein a结合的能力,使用偶联protein a的琼脂糖填料进行纯化;(3)将滤液通过1ml/分钟的流速流穿protein a预装柱,该步骤中滤液中的目标蛋白会与填料结合;(4)通过低盐和高盐缓冲液将柱上结合的杂质蛋白洗涤;(5)用低ph缓冲液将柱上结合的目标蛋白进行系统;
(6)将洗脱液迅速加入ph9.0的tris-hcl溶液,进行中和;(7)将上述中和后的蛋白溶液透析后,进行sds-page分析,确定蛋白纯度在95%以上,且浓度在0.5mg/ml以上后,低温保存备用。
109.实施例11:单域抗体真核表达载体rjk-v4-3的构建所提及纳米抗体通用的目标载体rjk-v4-3,为本公司在invitrogen商业化载体pcdna3.4(载体资料链接:https://assets.thermofisher.com/tfs-assets/lsg/manuals/pcdna3_4_topo_ta_cloning_kit_man.pdf)的基础上融合了人源igg4的重链编码序列中的fc区段后改造而来的,即该载体包含了igg4重链的铰链区(hinge)ch2和ch3区。具体改造方案如下:(1)选取pcdna3.4上的限制性酶切位点xbai和agei;(2)在fc片段编码序列的5’端和3’端通过重叠pcr的方式分别引入多克隆位点(mcs,multiple cloning site)和6
×
his标签;(3)使用分别带有xbai和agei酶切位点的一对引物通过pcr的方式将上述片段扩增;(4)使用限制性内切酶xbai和agei分别酶切pcdna3.4和(3)中的重组dna片段;(5)将酶切后的载体和插入片段在t4连接酶的作用下连接,然后将连接产物转化至大肠杆菌,扩增,测序核实,获得重组质粒。
110.实施例12:靶向人源il-17a的工具抗体(tool antibody, tab1)的表达和纯化本文中,tab 1即tab。tab1(secukinumab)为苏金单抗,序列来自imgt。
111.将搜索到的序列委托通用生物系统(安徽)有限公司进行哺乳动物细胞表达系统密码子优化,并克隆至pcdna3.1载体。经过抗性筛选,选择质粒阳性菌扩增,使用质粒中提试剂盒(macherey nagel, cat#740412.50)抽提质粒。按照每100ml细胞加入100μg质粒(40μg重链+60μg轻链),使用pei在293f细胞(培养基:freestyle 293 expression medium, thermo, cat#12338026 + f-68, thermo, cat#24040032)中瞬转表达;转染6~24h后加入5%体积的10% peptone(sigma,cat#p0521-100g),8% co
2 130rpm培养约7~8天;细胞活率降至50%时收取表达上清,使用proteina(ge,cat#17-5438-02)重力柱纯化;pbs透析后,使用nanodrop测定浓度,sec鉴定纯度,间接elisa验证结合能力;通过本方法获得的tab,浓度不小于2 mg/ml,纯度大于95%。
112.实施例13:抗体的抗原结合量效曲线测定本实施例采用标准酶联免疫吸附测定(elisa)操作流程进行。
113.(1)包被50μl 1μg/ml il-17a&f(acro,cat#ilf-h52wf,lot#g41-88zf1-rd),4℃过夜。
114.(2)洗板;加入200μl 5%牛奶,37℃封闭2h。
115.(3)将vhh-hfc稀释至2ug/ml,然后5倍梯度稀释抗体共8个浓度梯度。这里的vhh-hfc指实施例9制得的抗il-17a蛋白的单域抗体的fc融合抗体(在293f细胞中表达)经实施例10纯化而得。此外,还分别设置higg、tab对照;tab1由实施例12制得;(4)洗板;加入50μl经步骤(3)稀释得到的单域抗体,两复孔,37℃孵育1h。
116.(5)洗板;加入50μl hrp-goat anti higg 二抗,37℃孵育30min。
117.(6)洗板(多洗几次);加入50μl预先恢复常温的tmb,避光常温反应15min。
118.(7)加入50μl终止液(1n hcl),酶标仪读数保存。
119.(8)绘制曲线,计算ec50,如图2-3所示,其中higg指同型对照,不与任何靶标结合的免疫球蛋白分子,通过商品化购买得到;可见单域抗体1a10,1d7,1e6,2e2,2g3,3d1,3e1,3h2, 4a12均对il-17a蛋白结合效力和特异性优异。
120.图2中,2a示出了单域抗体1a10、1c9、1d1、1d4、1d7、1e2对il-17a蛋白的结合力,2b示出了单域抗体1e6、1h2、2d8、2e2、2f5、2g3对il-17a蛋白的结合力。图3中,3a示出了单域抗体3a6、3a8、3b11、3d1、3e1、3e2对il-17a蛋白的结合力,3b示出了单域抗体3f8、3h2、4a1、4a12、4f4、1h3对il-17a蛋白的结合力,3c示出了单域抗体3b10、4a1、4g4、tab1、higg对il-17a蛋白的结合力。
121.实施例14:抗体阻断il-17a与il-17r结合的测定本实施例采用标准酶联免疫吸附测定(elisa)操作流程进行。
122.(1)包被50μl 1μg/ml il-17ra & il-17rc(acro,cat#ilc-h5257,lot#g173a-2085f1-te),4℃过夜。
123.(2)洗板;加入200μl5%牛奶,37℃封闭2h。
124.(3)将biotin-il-17a(acro,cat#ila-h82q1,lot#cbv296p1-97rf1-qj))与梯度稀释的vhh-hfc混合,得到混合液(biotin-il-17a在混合液中的浓度为3.8ng/ml)。这里的vhh-hfc指实施例9制得的抗il-17a蛋白的单域抗体的fc融合抗体(在293f细胞中表达)经实施例10纯化而得。此外,还设置higg对照; higg指同型对照,不与任何靶标结合的免疫球蛋白分子,通过商品化购买得到;(4)洗板;加入50μl上述混合物,两复孔,37℃孵育1h。
125.(5)洗板;加入50μl streptavidin-hrp,37℃孵育30min。
126.(6)洗板(多洗几次);加入50μl预先恢复常温的tmb,避光常温反应15min。
127.(7)加入50μl终止液(1n hcl),酶标仪读数保存。
128.(8)绘制曲线,计算ec50。结果如图4-5所示,图4-5中横坐标表示经梯度稀释的vhh-hfc的具体浓度。可见,单域抗体1a10,1d7,1e6,2e2,2g3,3d1,3e1,3h2,4a12均有效阻断了il-17a与il-17r的结合。
129.图4中,4a示出了1a10、1c9、1d7、1e2、1d1、1d4的测定结果,4b示出了单域抗体3a6、3a8、3e1、3e2、3b11、3d1的测定结果,4c示出了3f8、3h2、4f4、1h3、4a1、4a12的测定结果。图5中,5a示出了3b10、4a1、higg、4g4的测定结果,5b示出了单域抗体1e6、1h2、2f5、2g3、2d8、2e2的测定结果。
130.实施例15:抗体(真核样品)中和人源il-17a诱导hela细胞释放il-6实验按照本领域技术人员通用的方法进行如下操作:(1)将复苏后传代3次及以上的hela细胞按10000个,50μl每孔铺入96孔板;(2)将梯度稀释的抗体与4*55ng/ml human il-17a(novo,cat#c774,lot#0331872-4147)等体积混合,将此混合物按每孔50μl加入细胞培养孔;这里的抗体为实施例9制得的抗il-17a蛋白的单域抗体的fc融合抗体(在293f细胞中表达)经实施例10纯化而得;此外,还分别设置higg、tab对照;higg指同型对照,不与任何靶标结合的免疫球蛋白分子,通过商品化购买得到;tab由实施例12制得;(3)细胞培养箱37℃孵育24h后,用human il-6 elisa 试剂盒检测上清中il-6浓
度;根据检测结果计算抗体中和il-17a诱导hela细胞释放il-6的ec50, 结果如图6-7所示,其中横坐标表示前述的梯度稀释的抗体的摩尔浓度;纵坐标表示上清中il-6浓度(单位pg/ml);从图6-7可知,真核抗体能有效中和il-17a并诱导hela细胞释放il-6。
131.图6中,6a示出了tab、higg的实验结果,6b示出了2g3、3d1、1d4、2e2的实验结果,6c示出了1a10、1d1、3e1、3h2的实验结果。图7中,7a示出了4a12、4f4的实验结果,7b示出了1c9、1d7、1e2、1e6的实验结果,7c示出了3e2、3f8、2f5、3a6的实验结果,7d示出了4a1、1h3的实验结果。
132.实施例16:抗体(人源化样品)中和人源il-17a诱导hela细胞释放il-6实验按照本领域技术人员通用的方法进行如下操作:(1)将复苏后传代3次及以上的hela细胞按10000个,50μl每孔铺入96孔板;(2)将梯度稀释的抗体与4*55ng/ml human il-17a等体积混合,将此混合物按每孔50μl加入细胞培养孔;这里的抗体为:人源化抗体2g3-v17、2g3-v18、2g3-v24、2g3-v25、2g3-v46、2g3-v50、2g3-v31、2g3-v34、1a10-v1、1a10-v4、1a10-v9、1a10-v10、1a10-v5、1a10-v6、1a10-v15或1a10-v14通过实施例7构建抗il-17a的人源化单域抗体的fc融合抗体真核表达载体(将人源化抗体的核苷酸序列构建入载体rjk-v4-3中),再通过实施例9表达及实施例10纯化而得。此外,还分别设置higg、tab对照;higg指同型对照,不与任何靶标结合的免疫球蛋白分子,通过商品化购买得到;tab由实施例12制得;(3)细胞培养箱37℃孵育24h后,用human il-6 elisa 试剂盒检测上清中il-6浓度;根据检测结果计算抗体中和il-17a诱导hela细胞释放il-6的ec50, 结果如图8-10所示,其中横坐标表示前述的梯度稀释的抗体的具体浓度;纵坐标表示上清中il-6浓度(单位pg/ml);从图8-10可知,人源化抗体能有效中和il-17a并诱导hela细胞释放il-6,且其技术效果优于抗体(真核样品)。
133.图8中,8a示出了tab、higg的实验结果,8b示出了2g3-v17、2g3-v18的实验结果,8c示出了2g3-v24、2g3-v25的实验结果。图9中,9a示出了2g3-v46、2g3-v50的实验结果,9b示出了2g3-v31、2g3-v34的实验结果,9c示出了1a10-v1、1a10-v4的实验结果。图10中,10a示出了1a10-v9、1a10-v10的实验结果,10b示出了1a10-v5、1a10-v6的实验结果,10c示出了1a10-v15、1a10-v14的实验结果。
134.尽管本发明的实施方案已公开如上,但其并不仅仅限于说明书和实施方式中所列运用,它完全可以被适用于各种适合本发明的领域,对于熟悉本领域的人员而言,可容易地实现另外的修改,因此在不背离权利要求及等同范围所限定的一般概念下,本发明并不限于特定的细节。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1