伊朗纤维素单胞菌菌株LG2020及其应用
伊朗纤维素单胞菌菌株lg2020及其应用
【技术领域】
1.本发明涉及微生物技术领域,具体涉及伊朗纤维素单胞菌菌株lg2020及其应用。
背景技术:
2.纤维素是由d-葡萄糖通过化学键-β-1,4糖苷键连接形成的一个长链状的高分子杂聚化合物,是植物细胞壁中含量最多的组分(30-50%),也是自然界中分布最广且结构非常复杂的一种多糖,一般由150-15000个葡萄糖基组成。在常温条件下,植物细胞壁中的纤维素分子间的氢键和范德华力这两种作用力聚合形成纤维素片层的二级结构,二级结构通过部分的疏水作用力连接-堆积形成三级的纤维结构-微纤丝,这种结构可以抵抗自由扩散的运输方式,即抵抗植物体内许多小分子物质的运输,如最常见的水分子以及酶分子甚至氢离子等,从而进一步的防止水解酶的水解或者有机试剂对结构的破坏;三级的纤维结构进一步的聚合会形成丝状基纤维晶体,使纤维素结构保持相对稳定的状态;让纤维素大分子不溶于水也不溶于大部分的有机溶剂,导致纤维素在自然界中比较难以被降解和利用,木质纤维素中主要就是以上面的这种结晶型的方式存在。然而,一般容易水解的纤维素分子,主要是以无定型纤维素链形成非晶态纤维素,水解过程首先是纤维素长链通过氧桥断裂形为纤维素短链,然后氧桥彻底断裂,导致纤维素分子完全被水解成单个的还原糖分子葡萄糖,然后葡萄糖小分子可以被微生物进一步利用,通过糖代谢中的糖酵解途径生成丙酮酸,再转化成为醇、乳酸或进入三羧酸循环生成其他的物质。目前科研者们主要采用物理、化学和生物法等方法来降解纤维素,其中,利用物理、化学方法降解纤维素不仅成本高,还会污染环境,从而破坏生态,而微生物降解木质纤维素具有能耗低、绿色环保无污染等优点,但是有周期较长,降解条件要求较高等缺点,因此我们发现,高效的降解微生物是使得生物法降解纤维素能够走上大规模应用的关键点。为了发现更多高效的降解纤维素的微生物资源,我们从不同的环境中采集样品,以期筛选到具有纤维素降解能力的微生物。同时我们对筛选得到的纤维素降解菌的降解能力和生化特性进行一些研究,为纤维素降解菌的进一步开发和利用提供一些理论依据。
技术实现要素:
3.有鉴于此,本发明的目的在于提供了一株伊朗纤维素单胞菌(cellulomonasiranensis)菌株lg2020及其应用。
4.为达到上述目的,本发明筛选得到一株伊朗纤维素单胞菌(cellulomonas iranensis)菌株lg2020,其保藏编号为gdmcc no.62533,保藏日期:2022年6月13日,保藏地址为:中国广州市先烈中路100号大院59号楼5楼,保藏单位:广东省微生物菌种保藏中心(gdmcc)。
5.进一步说明,伊朗纤维素单胞菌(cellulomonas iranensis)菌株lg2020,该菌株在lb液体培养基生长良好;菌落呈金黄色,直径约1mm,圆形,凸面,边缘整齐,不透明,表面形态平滑,有光泽,质地湿润。
6.本发明还提供一种伊朗纤维素单胞菌(cellulomonas iranensis)菌株lg2020在制备纤维素酶和/或木质素降解酶中的应用。
7.进一步说明,制备纤维素酶和/或木质素降解酶的产酶条件是:转速为200r/min、培养基装液量为200ml/500ml、培养基起始ph为6.4、接种量为15%、发酵产酶时间为3d、培养温度为37℃。
8.进一步说明,制备纤维素酶和/或木质素降解酶的产酶培养基条件:cmc-na最佳浓度2%、蛋白胨浓度为2%、dmf浓度为0.5%、牛血清蛋白浓度为1%、吐温-80浓度为0.5%、peg6000浓度为0.5%、10mm的氯化锌。
9.本发明还提供了所述的伊朗纤维素单胞菌(cellulomonas iranensis)菌株lg2020在降解木质纤维素中的应用。
10.本发明还提供了所述的伊朗纤维素单胞菌(cellulomonas iranensis)菌株lg2020在降解植物秸秆中的应用
11.综上所述,由于采用了上述技术方案,本发明的有益效果是:
12.利用本发明伊朗纤维素单胞菌cellulomonas iranensis菌株lg2020具有良好的降解纤维素、木质纤维素的能力。通过滤纸条降解试验,第7d时滤纸残余率为1.72%;秸秆降解能力试验,在第25天后,伊朗纤维素单胞菌已经将秸秆分解成非常薄的碎片,说明有很好的降解秸秆的能力。而市售的木质纤维素降解菌在降解秸秆25天后,秸秆整体形态并没有大的改变。且通过产酶条件和培养基的组分,结合响应面优化结果确定伊朗纤维素单胞菌cellulomonas iranensis菌株lg2020产酶的最佳条件:培养基装液量为200ml/500ml、培养基起始ph为6.44、接种量为15%、发酵产酶时间为3d、培养温度为37℃、转速为200r/min、最佳碳源为cmc-na、cmc-na最佳浓度为2%、最佳氮源为蛋白胨、蛋白胨最佳浓度为2.14%、牛血清蛋白最佳浓度为1.17%、dmf和吐温-80以及peg6000的最佳浓度均为0.5%、zncl2的最佳浓度为10mm。在以上最佳条件下发酵,伊朗纤维素单胞菌cellulomonas iranensis菌株lg2020的cmc酶活、fpa酶活和木聚糖酶活分别为49.8977u/ml、22.9328u/ml和90.777u/ml,较优化前酶活提高了9.712倍、6.602倍和9.634倍,其实测值与预测值的相对误差分别为1.76%、1.38%和2.00%。综上所述,本发明菌株lg2020在降解纤维素、木质纤维素领域都有着广阔的应用前景。
【附图说明】
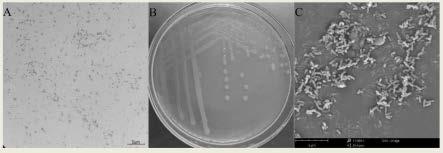
13.图1:菌株lg2020的形态学观察a:革兰氏染色观察;b:菌落形态观察;c:电子扫描电镜观察。
14.图2:lg2020菌株菌株16s rdna pcr扩增结果。
15.图3:lg2020菌株的16s rdna基因序列同源性为基础的系统发育树。
16.图4:图4(a)所示为葡萄糖标准曲线,图4(b)所示为木糖标准曲线。
17.图5:ph对cmc、fpa、木聚糖酶酶活力稳定性的影响。
18.图6:温度对cmc、fpa、木聚糖酶酶活力稳定性的影响。
19.图7:滤纸条降解滤纸残余率影响图。
20.图8:滤纸条降解滤纸影响图。图8(a)为空白组,图8(b)为处理组。
21.图9:秸秆降解效果对比图。其中,a0:伊朗纤维素单胞菌降解0天;a25:伊朗纤维素
单胞菌降解25天;b0:市售木质纤维素降解菌降解0天;b25:市售木质纤维素降解菌降解25天。
22.图10:摇床转速对cmc酶活、fpa酶活、木聚糖酶活和生长的影响图。
23.图11:装液量对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
24.图12:培养基ph对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
25.图13:接种量对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
26.图14:培养时间对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
27.图15:培养温度对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
28.图16:不同碳源(a)及最佳碳源浓度(b)对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
29.图17:不同氮源(a)及最佳氮源浓度(b)对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
30.图18:dmf对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
31.图19:牛血清蛋白对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
32.图20:吐温-80对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
33.图21:peg6000对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
34.图22:不同金属离子对cmc酶活、fpa酶活、木聚糖酶活和生长的影响。
35.图23:蛋白胨浓度和牛血清蛋白浓度对cmc酶活的等高线和响应面图。
36.图24:蛋白胨浓度和培养基ph对fpa酶活的等高线和响应面图。
37.图25:蛋白胨浓度和牛血清蛋白浓度对木聚糖酶活的等高线和响应面图。
【具体实施方式】
38.本说明书中公开的所有特征,或公开的所有方法或过程中的步骤,除了互相排斥的特征和/或步骤以外,均可以以任何方式组合。本说明书(包括任何附加权利要求、摘要)中公开的任一特征,除非特别叙述,每个特征只是一系列等效或类似特征中的一个例子而已。
39.实施例1:伊朗纤维素单胞菌的分离及鉴定
40.1.1实验材料
41.1.1.1样品的采集样品采集于广西柳州市里高镇螺蛳养殖基地的泥塘土样。
42.1.1.2培养基
43.初筛液体培养基:cmc-na 15g,k2hpo41 g,nh4no31 g,mgso4·
7h2o0.5 g,nacl 0.5g,ph自然,ddh2o定容至1l,121℃灭菌20min。
44.初筛固体培养基:在初筛液体培养基的基础上添加2%琼脂。
45.复筛培养基:配方同初筛培养基。
46.lb液体培养基:蛋白胨10g,酵母粉5g,nacl 10g,ph自然,ddh2o定容至1l,121℃灭菌20min。
47.lb固体培养基:在lb液体培养基的基础上添加2%琼脂。
48.滤纸崩解培养基:k2hpo41 g,nh4no31 g,mgso4·
7h2o 0.5g,nacl 0.5g,3g滤纸条(1cm
×
6cm),ph自然,ddh2o定容至1l,121℃灭菌20min。
49.产酶培养基:cmc-na 10g,蛋白胨20g,mgso4·
7h2o 0.3g,k2hpo4·
3h2o 1.5g,nacl 5g,cacl2·
2h2o 0.1g,ddh2o定容至1l,ph6.4,121℃灭菌20min.
50.1.1.3主要溶液
51.1mg/ml葡萄糖标准液:取100mg葡萄糖(80℃烘干至恒重),溶于ddh2o中,定容至100ml,4℃保存备用。
52.1mg/ml木糖标准液:取100mg木糖(80℃烘干至恒重),溶于ddh2o中,定容至100ml,4℃保存备用。
53.0.1mol/l柠檬酸缓冲液:取21.014g柠檬酸(分子量210.14)溶于ddh2o中,定容至1000ml,4℃保存备用。
54.0.1mol/l柠檬酸钠缓冲液:取29.412g的柠檬酸钠(分子量294.12)溶于ddh2o中,定容至1000ml,4℃保存备用。
55.ph 4.5,0.05mol/l柠檬酸-柠檬酸钠缓冲液:准确量取27.12ml柠檬酸溶液和22.88ml柠檬酸钠溶液,再定容至100ml,混合均匀后,即为ph4.5,0.05mol/l柠檬酸-柠檬酸钠缓冲液,4℃保存备用。
56.1%羧甲基纤维素钠溶液:称取1g羧甲基纤维素钠(cmc-na),加入适量ph4.5浓度为0.05mo//l柠檬酸-柠檬酸钠缓冲液,加热溶解,直至cmc-na完全溶解,冷却后用缓冲液定容至100ml,混合均匀后,4℃保存备用。
57.1%木聚糖溶液:称取1.0g木聚糖溶解于配好的ph4.5浓度为0.05mol/l柠檬酸-柠檬酸钠缓冲液中,溶解并定容到100ml,4℃保存备用。
58.3,5-二硝基水杨酸试剂(dns):称取6.3g 3,5-二硝基水杨酸,并量取262ml2 mol/l naoh到酒石酸钾钠的热溶液中(182g酒石酸钾钠溶于500mlddh2o中),再添加5g苯酚和5g亚硫酸钠溶于其中,搅拌溶解,冷却后ddh2o定容到1000ml贮于棕色瓶中。室温放置一周后使用。
59.1.2.1纤维素降解菌的筛选
60.(1)菌株的初筛
61.取一定量的土样加入250ml初筛液体培养基里,在37℃,150r/min的摇床中培养,培养3-7d,每隔12h观察一下培养基中溶液颜色的变化情况,直至培养基变浑浊。将富集液进行取样稀释,吸取900μl的培养基于1.5ml离心管,取100μl的富集悬浊液于1.5ml的离心管进行稀释,稀释梯度为10-5-10-8,分别取10μl于富集培养基进行涂布,做一组空白对照,于37℃的生化培养箱中培养,每隔12h观察一次是否长菌,72-120h将会培养出菌,选取生长旺盛、长势良好的菌株,转接到复筛培养基上,待进一步复筛确定。
62.(2)菌株的复筛
63.从初筛培养基上得到的菌株接到复筛培养基上,37℃恒温培养箱培养3-5d。然后挑取其中长势较好的菌株,接种于滤纸崩解培养基中,测试滤纸残余率;同时用dns法测定cmc酶活、fpa酶活和木聚糖酶活,综合考虑滤纸残余率和3种酶活大小的结果,最终筛选出纤维素降解能力高的菌株。
64.1.2.2纤维素降解菌的粗酶液的制备
65.将选出来的菌株接到lb液体培养基上,在37℃、150r/min条件下培养,做成106cfu菌悬液,以10%的接种量接种于发酵产酶培养基中,于37℃、150r/min培养3d,获得发酵液,
将发酵液10000r/min离心10min,取上清液即为粗酶液。
66.1.2.3纤维素降解菌的鉴定与保藏
67.(1)菌株的形态学观察
68.将最终筛选出来的菌株划线至lb固体培养基,倒置放在37℃的培养箱中,培养3d后从平板上观察菌落形态、颜色以及大小,并挑选其中的单菌落进行革兰氏染色镜检。同时,参考曾令杰的方法,用扫描电镜观察菌株的形态变化。
69.(2)菌株的分子生物学鉴定
70.用培养的lb菌液作为dna模板,以细菌16s rrna的一对通用引物27f(5
′‑
agagtttgatcctggctcag-3
′
)和1492r(5
′‑
tacggttaccttgttacgactt-3
′
)进行pcr扩增16s rdna序列。pcr反应体系如表1所示。
71.表1pcr反应体系(25μl)
[0072][0073]
扩增条件如下:
[0074][0075]
将反应体系混合按照扩增条件进行扩增。
[0076]
pcr扩增结束后,进行电泳实验,电泳结束将凝胶放于将紫外仪观察条带,然后根据普通琼脂糖凝胶dna回收试剂盒(北京天根生化科技有限公司)对pcr扩增产物进行回收纯化(具体操作见试剂盒操作说明书。)接着将回收好的dna片段寄至广州华大基因有限公司,获得菌株16s rdna鉴定结果,进入ncbi网站,选择blast工具,输入测得的序列,运行blast,根据查询得出结果的最高分、总体分、碱基覆盖率、e值(期望值)相符程度等,最后用软件mega 5.0完成系统发育树的绘制,确定菌株的分类学地位。
[0077]
(3)菌株的保藏
[0078]
取单菌落接种至lb液体培养基中,37℃、150r/min条件下培养1-3d后与50%
[0079]
甘油按1:1的比例,装至无菌的1.5ml离心管中-80℃冷冻保藏。
[0080]
1.2.4纤维素降解菌的酶学性质研究
[0081]
(1)葡萄糖和木糖标准曲线的绘制
[0082]
参考赵钰的方法,取0-2.0ml的1mg/ml的葡萄糖/木糖的标准液于比色管中,ddh2o补足至2ml,再加入dns 3ml(如表2)混匀,沸水浴5min,取出后冷却至室温,用ddh2o定容至25ml,颠倒混匀,用分光光度计测量od540nm。以光密度值为纵坐标,葡萄糖/木糖含量(mg)为横坐标,分别作葡萄糖和木糖的标准曲线。
[0083]
表2标准葡萄糖/木糖溶液的配制
[0084]
编号012345678910葡萄糖/木糖标准液00000111112.0ddh2o/ml21111100000dns溶液/ml33333333333
[0085]
(2)内切型-β-葡聚糖酶(cmc酶)活力的测定
[0086]
取1%cmc-na溶液1ml,加入ph4.5的柠檬酸缓冲液0.5ml,粗酶液0.5ml,于50℃水浴锅中反应30min,加入3mldns试剂,充分摇匀,沸水浴10min后取出,定容至25ml,颠倒混匀,测量其od540nm。另外把在100℃煮沸20min后的酶液作为对照。
[0087]
(3)滤纸酶(fpa酶)活力的测定
[0088]
取50mg(1cm
×
6cm)新华滤纸作为底物,加入ph 4.5的柠檬酸缓冲液1.5ml,把反应时间改为60min,余下步骤和测定cmc酶活的一致。
[0089]
(4)木聚糖酶活力的测定
[0090]
测定步骤和测定cmc酶活的一致,只需要把底物改成1%的木聚糖溶液。
[0091]
(5)酶反应的最适ph
[0092]
根据2.2.3粗酶液的制备方法,测定酶促反应在50℃下不同ph值缓冲溶液对酶活性的影响,ph为2.5-8的柠檬酸-柠檬酸钠缓冲溶液,测cmc酶活、fpa酶活和木聚糖酶活。以酶反应最适ph下的酶活为100%,计算其相对酶活。
[0093]
(6)酶反应的最适温度
[0094]
根据2.2.3粗酶液的制备方法,在最适ph值下,测定不同温度对酶活性的影响,调节酶促反应中的温度分别为30-80℃,测cmc酶活、fpa酶活和木聚糖酶活。以酶反应最适温度下的酶活为100%,计算其相对酶活。
[0095]
(7)酶活定义
[0096]
酶活定义:在温度50℃和ph为4.5的条件下,1min从底物溶液中分解产生1微克还原糖所需要的酶量即为一个酶活单位,简称为u/ml。酶活公式为:
[0097][0098]
式中:u-纤维素酶酶活力,u/ml;还原糖含量,mg;1000-转化因子;n-稀释倍数;t-反应时间,min;v-酶液体积,ml。
[0099]
1.3.1纤维素降解菌的筛选结果
[0100]
从广西柳州市里高镇螺蛳养殖基地的泥塘土样中以cmc-na为唯一碳源的培养基中进行初筛,结果涂布于复筛固体培养基平板上,获得一株生长状况良好的菌株,对该菌株命名为hf,hf菌株的cmc、fpa和木聚糖酶活分别为5.1378u/ml、3.4737u/ml和9.4223u/ml。
[0101]
1.3.2菌株的形态学观察结果
[0102]
如图1所示,在显微镜观察下,革兰氏染色呈阳性,但容易脱色;从菌落形态观察,菌落呈金黄色,直径约1mm,圆形,凸面,边缘整齐,不透明,表面形态平滑,有光泽,质地湿润。扫描电镜观察,hf菌呈不规则的、直的或稍微弯曲的杆状,0.4
×
2.5-4.0μm,偶尔表现出典型的棒状分枝。
[0103]
1.3.3菌株的分子生物学鉴定结果
[0104]
通过采用菌落pcr技术法,以hf菌株为dna模板扩增其16s rdna序列,菌株的pcr扩增结果如图2所示。由图可知,1通道为空白组,没有条带,2和3通道扩增的目的条带大小约为1500bp,和理论相同。
[0105]
hf菌株的测序结果在ncbi的genbank数据库blast序列同源性分析比对,接着用软件mega5.0进行系统发育树的构建,结果如图3所示。经序列比对后,hf菌与纤维素单胞菌属(cellulomonas sp.)的伊朗纤维素单胞菌(cellulomonas iranensis)序列相似度为100%。
[0106]
1.3.4菌株的酶学性质研究
[0107]
(1)葡萄糖和木糖标准曲线的绘制
[0108]
根据实验结果以葡萄糖/木糖含量(mg)为横坐标,对应的吸光度od540nm值为纵坐标。如图4(a)所示为葡萄糖标准曲线,其对应的曲线回归方程为:y=0.6407x+0.0089,(r2=0.9992。)如图4(b)所示为木糖标准曲线,其对应的曲线回归方程为:y=0.7033x-0.001,(r2=0.9992。)两者都符合标准曲线的要求。
[0109]
(2)酶反应的最适ph
[0110]
为了确定3种酶的最适ph,首先用柠檬酸-柠檬酸钠缓冲液将反应体系的ph调成2.5-8.0,结果如图5所示,当ph为2.5时,cmc、fpa和木聚糖酶的相对酶活分别为28.85%、22.03%、25.84%;ph在2.5-4.5之间时,3种酶的活性呈显著上升趋势,当ph为4.5时,cmc、fpa、木聚糖的酶活达到最大;当反应体系的ph大于4.5时,酶活减小,为下降趋势。当ph达到8.0时,cmc、fpa和木聚糖酶的相对酶活分别为13.74%、10.78%、12.29%。因此,3种酶活的酶反应最适ph值为4.5。
[0111]
(3)酶反应的最适温度
[0112]
为了找到hf菌株的酶反应体系的最佳温度值,我们测定了30-80℃对酶的影响,结果如图6所示,cmc、fpa和木聚糖酶的最适反应温度均为50℃。当反应体系的温度小于50℃时,3种酶活随着反应体系温度的升高而升高,都呈现一个显著上升趋势且上升速率基本相同;而温度为45-55℃时,3种酶活的相对酶活保持在80%以上;当温度大于50℃时,3种酶活随着温度的升高而降低,可能是温度太高会使酶的结构发生改变,不能与底物相结合,甚至导致酶失活;当温度升高至80℃时,cmc、fpa和木聚糖酶的相对酶活分别为25.35%、16.21%、20.28%。
[0113]
1.3.5滤纸条降解试验结果
[0114]
为了检验hf菌株的降解滤纸的能力,我们测定了滤纸残余率。如图7所示,前面3d菌株降解滤纸的速率比较慢,可能是菌体在适应滤纸崩解培养基或者菌体在利用培养基在进行生长繁殖,当培养到第3d时,滤纸的残余率为81.56%,之后滤纸的残余率急速下降,第4d的残余率为54.79%,从第3-6d是降解速度最快的时期,到第6d时,滤纸的残余率为5.25%,之后滤纸的残余率速度明显变慢,到第7d时滤纸的残余率为1.72%;即hf菌株降解
了3g/l的滤纸。
[0115]
此外,我们设置了空白组(没有添加菌体)和处理组(添加菌体),从图8(a)我们发现空白组的滤纸条没有被降解,溶液澄清,而从图8(b)我们发现处理组中没有了滤纸条,并且培养基变黄、变浑浊,还有微小的絮状沉淀;通过滤纸残余率和实验现象说明了hf菌株具有较强的降解纤维素的能力。
[0116]
可见,本试验从螺蛳养殖基地的泥塘土样中筛选得到一株纤维素降解菌hf,该菌株的cmc、fpa和木聚糖酶活分别为5.1378u/ml、3.4737u/ml和9.4223u/ml,经鉴定为伊朗纤维素单胞菌(cellulomonas iranensis)为了可以得到hf菌株的最高酶活,我们做了部分酶学性质的实验,发现hf菌株的3种酶活的酶反应最适ph和温度均分别为4.5和50℃。最后为了验证hf菌株的降解纤维素的能力,我们做了滤纸条降解试验,发现空白组中的滤纸条没有丝毫的影响,而处理组中溶液变浑浊,滤纸的残余率为1.72%,说明了hf菌株的降解滤纸能力较强。
[0117]
秸秆降解结果:见图9,从图9可已看出,25天后,伊朗纤维素单胞菌已经将秸秆分解成非常薄的碎片,说明有很好的降解秸秆的能力。而市售的木质纤维素降解菌在降解秸秆25天后,秸秆整体形态并没有大的改变。
[0118]
经序列比对后,hf菌与纤维素单胞菌属(cellulomonas sp.)的伊朗纤维素单胞菌(cellulomonas iranensis)序列相似度为100%。
[0119]
现已经经过微生物分类学鉴定并已经经过保藏:命名为伊朗纤维素单胞菌(cellulomonas iranensis)菌株lg2020,其保藏编号为gdmcc no.62533,保藏日期:2022年6月13日,保藏地址为:中国广州市先烈中路100号大院59号楼5楼,保藏单位:广东省微生物菌种保藏中心(gdmcc)。
[0120]
1.3.6菌株lg2020对稻草秸秆和甘蔗叶的降解
[0121]
将自然晒干的稻草秸秆和甘蔗叶粉碎成长度1cm左右的丝状,80℃干燥至恒重。新鲜培养500ml伊朗纤维素菌lg2020500ml,无菌状态下准确称取10克粉碎后的稻草秸秆和甘蔗叶到菌液中,搅拌均匀,放置于30℃培养。稻草和秸秆各准备12瓶。每天各取2瓶,将稻草秸秆或甘蔗叶用清水清洗烘干后称重,取平均值后计算降解率,测定6天,结果见表3。
[0122]
表3
[0123][0124]
可见,lg2020菌液降解稻草秸秆4天降解率达到52.3%,6天达到64.5%,lg2020菌液降解甘蔗叶效果更佳,4天降解率达到58.5%,6天降解率达到72.2%。
[0125]
2.1.1菌株粗酶液的制备
[0126]
hf菌株粗酶液的制备:将选出来的菌株接到lb液体培养基上,在37℃、150r/min条件下培养,做成106cfu菌悬液,以10%的接种量接种于发酵产酶培养基中,于37℃、150r/min培养3d,获得发酵液,将发酵液10000r/min离心10min,取上清液即为粗酶液。然后测定cmc酶活、fpa酶活、木聚糖酶活和od600,每组3个平行。
[0127]
2.1.2菌株产酶外部条件的优化
[0128]
(1)摇床转速对产酶的影响
[0129]
在不同摇床转速75-250r/min条件下培养。以不同转速下最大的酶活为100%,计算其相对酶活。
[0130]
(2)培养基装液量对产酶的影响
[0131]
在500ml的三角瓶中分别装100-300ml培养基,在上一步的优化的转速下培养。以不同培养基装液量下最大的酶活为100%,计算其相对酶活。
[0132]
(3)培养基起始ph对产酶的影响在上述两步优化的条件下,调节培养基的起始ph值分别为4.0-8.4培养。以不同培养基起始ph下最大的酶活为100%,计算其相对酶活。
[0133]
(4)接种量对产酶的影响
[0134]
在上述几步的最优条件下,分别以接种量为2.5-20%的菌液接种至产酶培养基中培养。以不同接种量下最大的酶活为100%,计算其相对酶活。
[0135]
(5)培养时间对产酶的影响
[0136]
在上述几步的最优条件下,将培养基培养1-7d。以不同培养时间下最大的酶活为100%,计算其相对酶活。
[0137]
(6)培养温度对产酶的影响
[0138]
在上述几步的最优条件下,将培养基置于27-46℃下培养。以不同培养温度下最大的酶活为100%,计算其相对酶活。
[0139]
2.1.3菌株产酶培养基条件的优化
[0140]
在基于hf菌株的产酶外部最优条件下,再对产酶培养基进行优化。
[0141]
(1)不同碳源及碳源浓度对产酶的影响
[0142]
分别以10g/l可溶性淀粉、葡萄糖、蔗糖、cmc-na、乳糖、果糖、滤纸和麦芽糖为碳源,以未添加碳源为对照,确定培养基的最优碳源。以空白组的酶活为100%,计算其相对酶活。
[0143]
在确定最佳培养碳源后,设计碳源浓度梯度为0.5-3%。以不同氮源浓度下最大的酶活为100%,计算其相对酶活。
[0144]
(2)不同氮源及氮源浓度对产酶的影响
[0145]
分别以20g/l的蛋白胨、玉米浆粉、黄豆粉、硝酸钠、硫酸铵、氯化铵、尿素、硝酸钾和硝酸铵为氮源,以未添加氮源为对照,确定培养基的最优氮源。以空白组的酶活为100%,计算其相对酶活。
[0146]
在确定最佳培养氮源后,设计氮源浓度梯度为0.5-3%。以不同氮源浓度下最大的酶活为100%,计算其相对酶活。
[0147]
(3)dmf对产酶的影响
[0148]
在上述优化后的产酶培养基里分别添加浓度为0-2%dmf,以未添加dmf为对照。以空白组的酶活为100%,计算其相对酶活。
[0149]
(4)牛血清蛋白对产酶的影响
[0150]
在上述优化后的产酶培养基里分别添加浓度为0-2%牛血清蛋白,以未添加牛血清蛋白为对照。以空白组的酶活为100%,计算其相对酶活。
[0151]
(5)吐温-80对产酶的影响
[0152]
在上述优化后的产酶培养基里分别添加浓度为0-2%吐温-80,以未添加吐温-80
为对照。以空白组的酶活为100%,计算其相对酶活。
[0153]
(6)peg6000对产酶的影响
[0154]
在上述优化后的产酶培养基里分别添加浓度为0-2%peg6000,以未添加peg6000为对照。以空白组的酶活为100%,计算其相对酶活。
[0155]
(7)不同金属离子对产酶的影响
[0156]
在上述优化后的产酶培养基分别添加浓度为10mm的zn2+、mg2+、al3+、cu2+、k+、ag+、ca2+、mn2+、fe2+、hg+、fe3+,以未添加金属离子为对照。以空白组的酶活为100%,计算其相对酶活。
[0157]
2.1.4响应面法优化产酶条件
[0158]
在单因素试验的基础上,根据中心组合设计原理,选择蛋白胨浓度、牛血清蛋白浓度、培养基起始ph、培养温度4个因素作为影响其产酶活性的自变量,进一步研究各个因素之间的关系,利用软件designexpert 10和sas 9.1做4因素3水平试验确定产酶活力的最佳培养条件。因素水平编码表见表4。
[0159]
表4box-behnken试验设计因素和水平表
[0160][0161]
2.1.5数据统计与分析
[0162]
每组试验生物学重复3次,采用origin pro 9.1软件和graphpad prism 8.0.1软件对数据进行图表的绘制
[0163]
2.2结果与分析
[0164]
2.2.1摇床转速对产酶的影响
[0165]
转速的大小不仅影响菌体生长的速度以及新陈代谢,还影响发酵过程中溶氧量的多少,直接影响了菌体的生长繁殖和酶的活性。实验结果如图10所示,当摇床转速小于200r/min时,hf菌株的3种酶活和od600随着摇床转速的增加而逐渐升高,因为转速的升高会引起培养基和空气的接触面积变大,使培养基中溶入了大量的氧气,补充了菌体在生长繁殖过程中对氧气的消耗量;当转速为200r/min时,3种酶的产量最大。当转速超过200r/min时,hf菌株的od600出现上升,而酶活呈现快速下降的趋势;可能原因是摇床的转速过高,导致菌体生长代谢过快,使菌体提前进入了凋亡期。因此,选择摇床转速为200r/min最有利于hf菌株的产酶。
[0166]
2.2.2培养基装液量对产酶的影响
[0167]
装液量的多少直接影响着三角瓶中的氧气的浓度,由菌株的培养条件优化可知,hf菌株为好氧型细菌,本实验在500ml的三角瓶中分别设定9组,装液量范围为100-300ml,结果如图11所示,装液量没有很大影响hf菌株的od600;当装液量为100ml时,3种酶活的相对酶活为49.17%、19.18%、50.64%,当装液量小于200ml时,3种酶活的相对酶活都随着装液量的递增而增大,其中相对酶活增长速率比较显著的是fpa酶活,增加了80%左右;且木
聚糖酶活的相对酶活都比cmc和fpa的高;当装液量为200ml时,3种酶活性最大;当装液量在175-225ml时,木聚糖酶的相对酶活在90%以上;随着装液量的增加,3种酶活性均减小,且木聚糖酶活的速率呈直线式的下降,可能原因是装液量越多,培养基与空气接触的面积减少,使菌体得不到氧气,导致菌体的酶活性减小。因此,选择500ml的装液量为200ml最有利于hf菌株的产酶。
[0168]
2.2.3培养基起始ph对产酶的影响
[0169]
本实验测定了培养基初始ph 4.0-8.4之间的产酶能力,结果如图12所示。在ph4.0-6.4之间,hf菌株的od600和酶活力呈上升趋势,且ph在6.0-7.2之间的相对酶活基本都在80%以上,而在ph值大于6.4时又呈下降趋势,且发现fpa酶活的相对酶活比cmc酶活和木聚糖酶活的高;在ph为8.4时,3种酶活的相对酶活分别为52.55%、56.46%和32.15%。此外,还发现在pfa酶活和cmc酶活的下降趋势基本相同。综上所述,hf菌株适宜在弱酸和中性的环境下进行产酶发酵。因此,选择培养基ph为6.4最有利于hf菌株的产酶。
[0170]
2.2.4接种量对产酶的影响
[0171]
接种量对酶的活性以及发酵的周期都会产生不同程度的影响。结果如图13所示,接种量的增大对hf菌株的生长没有太大的影响;当接种量为2.5%时,3种酶活的相对酶活分别为15.59%、18.92%和22.27%;当接种量在2.5%~15%时,3种酶的产量随着接种量的增加而逐渐增大且增长速率没有稳定的规律性产酶;当接种量为15%时,菌株的产酶活性最高。当接种量大于15%时,酶的产量随着接种量的增加而减少,可能原因是接种量过大,菌体大量繁殖,营养物质过度吸收,溶氧含量迅速下降,导致酶活下降。因此,选择接种量为15%最有利于hf菌株的产酶。
[0172]
2.2.5培养时间对产酶的影响
[0173]
为了找到hf菌株的最佳产酶时间,我们测定了1-7d的酶活。实验结果如图14所示,在培养前期,由于发酵时间短,菌体还在对数生长期,没有进入稳定期,使得hf菌株数量不够,而且大部分菌体还没有到达产酶期,在第1d时,3种酶活的相对酶活均在50%左右。随着发酵时间的不断增加,菌体产生的酶量不断积累,当发酵时间为第3d时酶的产量达到最大,且菌体进入了稳定期。随着培养时间的增长,第3d之后,菌体开始进入凋亡期,菌体慢慢的衰老自溶,再加上产酶培养基中环境的一些代谢产物的积累已不利于菌体生长与产酶,使发酵液中的酶活开始降低。因此,选择培养时间为第3d最有利于hf菌株的产酶。
[0174]
2.2.6培养温度对产酶的影响
[0175]
温度的高低影响着菌体的生长繁殖的速度,当温度较低时,菌体生长速率缓慢,难以吸收营养物质来产酶,使产酶量较低;而当温度过高时,菌体生长代谢较快,导致菌体提前进入稳定期和凋亡期,使菌体的数量减少,也就导致菌体产酶能力降低。本实验设定7个温度梯度,hf菌株的产酶情况如图15所示,当温度小于37℃时,hf菌株的od600和酶活都随着温度的升高而升高;当温度范围在34-40℃时,菌株的产酶活性都较大,且cmc酶和木聚糖酶活的相对酶活在80%以上;在当温度大于37℃时,od600和酶活都下降;当温度为46℃时,cmc酶活、fpa酶活、木聚糖酶的相对酶活分别为13.94%、13.46%
[0176]
和33.34%。可能原因是温度过高,菌株体内酶的活性降低从而影响代谢过程。同时我们发现,温度对fpa酶活的影响更大。因此,选择培养温度为37℃最有利于hf菌株的产酶。
[0177]
2.2.7不同碳源及碳源浓度对产酶的影响
[0178]
碳源不仅仅可以为hf菌株的生长提供营养物质,还可以对纤维素酶起诱导作用。碳源的优化结果如图16(a)所示,结果表明可溶性淀粉、葡萄糖、蔗糖、cmc-na、滤纸、麦芽糖和果糖作为唯一碳源时,菌体均可以大量的生长繁殖,且均可产生cmc酶、fpa酶和木聚糖酶,都比空白组的高。以空白组的相对酶活为100%,发现所有的碳源都可以促进菌株hf产生酶活;其中以cmc-na为碳源时,菌株的所产酶活最高,此时cmc酶、fpa酶和木聚糖酶的相对酶活分别为289.35%、383.02%、285.50%,远远高于其他的几种碳源。同时,我们发现fpa酶活的相对酶活在除麦芽糖的碳源中,其比cmc酶活和木聚糖酶活的相对都要高,且cmc-na和滤纸中的pfa酶活的相对酶活比另外两种酶的相对酶活都高了80%左右,可能是这两种碳源更有利于pfa酶活的生产。因此我们选择cmc-na作为碳源进行下一步优化。
[0179]
同时,我们还对cmc-na的浓度做了优化,结果如图16(b)所示,当cmc-na浓度在0.5%-1.0%之间时,随着浓度的增加3种酶活不断升高,只是浓度在1%-2.5%之间时,3种酶活的相对酶活大小相差不大,基本都在90%以上;在cmc-na浓度为2.0%时,3种酶活的相对酶活为100%;当cmc-na浓度高于2.0%时,3种酶活的相对酶活在缓慢减小,可能原因是cmc-na的浓度过高,抑制了菌体的生长繁殖。因此,选择cmc-na浓度为2%最有利于hf菌株的产酶。
[0180]
2.2.8不同氮源及氮源浓度对产酶的影响
[0181]
氮源是微生物细胞核酸和酶的重要组成元素,实验一共选取了9种不同的氮源分别对hf菌株进行发酵产酶培养,各氮源所测得的3种酶活力的相对酶活如图17(a)所示。由图可知,以空白组的相对酶活为100%,在以蛋白胨作为唯一氮源所测得的酶活力最高,且明显高于其他氮源,cmc酶、fpa酶和木聚糖酶的相对酶活分别为323.34%、410.12%、324.29%;其次是玉米浆粉对酶活也比较显著,3种酶活约为空白组的1.73、1.49和1.93倍。黄豆粉对木聚糖活有促进作用,且相对酶活为192.44%,约为空白组的2倍;其却对cmc酶活和fpa酶活有抑制的作用。因此,在不同氮源对酶活力的影响的单因素优化中,最适氮源为蛋白胨。接着我们对蛋白胨的浓度进行了产酶优化,实验结果如图17(b)所示,蛋白胨浓度促进了菌体的生长繁殖,说明选用蛋白胨作为氮源是合理的;蛋白胨浓度在2%时可达到其最大酶活;当浓度为0.5%时,3种酶活的相对酶活分别为53.50%、30.14%和56.33%。在浓度小于1.5%时,随着蛋白胨浓度的增加,相对酶活不断上升,并且在浓度1.5-2%之间,3种的相对酶活相差不大,而在浓度大于2%时,随着蛋白胨浓度的增加,3种酶的相对酶活呈下降的趋势。因此,选择蛋白胨浓度为2%最有利于hf菌株的产酶。
[0182]
2.2.9dmf对产酶的影响
[0183]
dmf作为高产纤维素酶的一种优良诱导剂,主要是诱导细胞内ca2+水平显著升高,并触发ca2+-crz1信号转导通路诱导酶基因转录,促进了酶活的生成。通过在产酶培养基中添加不同浓度的dmf来提高酶的产量,实验结果如图18所示,dmf明显的促进了hf菌株的生长,且在dmf浓度为0.5%时可达到其最高酶活,cmc酶和木聚糖酶的相对酶活为123.42%、110.72%;而dmf浓度在1.0%时,fpa酶的相对酶活达到最大为
[0184]
115.06%;当浓度大于1.0%时,随着dmf浓度的增加,相对酶活不断降低,且相对酶活小于空白组,可能原因是dmf过高的浓度会抑制了菌体的产酶能力。因此,选择dmf浓度为0.5%最有利于hf菌株的产酶。
[0185]
2.2.10牛血清蛋白对产酶的影响
[0186]
牛血清蛋白常常被作为酶的稳定剂,且较强的疏水性,它可以在溶液中形成分子有序聚集体-胶束以加速底物、酶和产物的运动,促进酶系的解吸附,提高底物和酶的有效吸附,从而使酶活明显提高。结果如图19所示,产酶培养基中添加牛血清蛋白对hf菌株的生长没有太大的影响;当牛血清蛋白浓度小于1%时,3种酶的酶活随着牛血清蛋白浓度的增加而逐渐增大;在浓度为1%时,菌株的产酶活性最高,cmc酶、fpa酶和木聚糖酶的相对酶活为215.57%、206.97%和192.23%,和空白组比较,发现3种酶活约增加了2倍。当浓度大于1%时,酶的活性随着牛血清蛋白浓度的增加而减少,且fpa酶活的下降趋势比cmc酶活和木聚糖酶活的下降趋势要缓。因此,选择牛血清蛋白浓度为1%最有利于hf菌株的产酶。
[0187]
2.2.11吐温-80对产酶的影响
[0188]
吐温-80的亲水端可以吸附纤维素酶,增加酶与底物的结合和可及性,使得纤维素酶组分体系更稳定。结果如图20所示,产酶培养基中添加吐温-80对hf菌株的生长繁殖有显著的作用;当吐温-80浓度小于0.5%时,3种酶的产量随着浓度的增大而逐渐增大;在浓度为0.5%时,菌株的产酶活性最高,cmc酶、fpa酶和木聚糖的相对酶活为123.42%、114.02%、110.70%。当浓度大于0.5%时,酶的活性随着吐温-80浓度的增加而减少;同时发现木聚糖酶的相对酶活比cmc酶和fpa酶的相对酶活都高,可以猜测吐温-80更有利于hf菌株产木聚糖酶。因此,选择吐温浓度为0.5%最有利于hf菌株的产酶。
[0189]
2.2.12peg6000对产酶的影响
[0190]
peg6000能改变底物的结构,增加纤维素酶的可及性,同时还可以使酶更稳定,阻止它们在产生过程中发生变性;如图21所示,peg6000对hf菌株生长没有太大的影响;在浓度为0.5-1.5%之间时,3种酶的相对酶活均相差不大,但都比空白组的相对酶活要高。当浓度大于1.5%时,3种酶的相对酶活都呈下降的趋势,并且fpa酶活下降最明显。当peg6000浓度为2%时,3种酶的相对酶活分别为89.42%、61.59%和74.86%。因此,选择peg6000浓度为0.5%最有利于hf菌株的产酶。
[0191]
2.2.13不同金属离子对产酶的影响
[0192]
有些金属离子可能是酶的激活剂而能够促进酶活,而有的金属离子可与酶活性基团作用而使酶失活。如图22所示为在终浓度为10mm金属离子的条件下对hf菌株的粗酶液中3种酶活性的影响。由图可知,zn
2+
、mg
2+
、cu
2+
对hf菌株的cmc酶活有促进作用,其中zn
2+
对3种酶活均有明显的促进作用,cmc酶活、fpa酶活和木聚糖酶活的相对酶活分别达到了118.65%、117.49%和122.18%,而al
3+
、ca
2+
、k
+
、fe
3+
、fe
2+
对酶活有抑制作用,但是促进了菌体的生长,其中hg
+
、ag
+
完全抑制了酶的活性和菌体的生长。mn
2+
对酶活的影响不明显。最后,在产酶培养基中加入了10mm的氯化锌。
[0193]
2.2.14响应面法优化产酶条件
[0194]
运用design expert 10创建box-behnken试验组,共进行29组试验,其结果见表5。
[0195]
表5box-behnken试验结果一览表
[0196][0197][0198]
通过对box-behnken试验结果一览表的29组数据进行二次多项拟合,获得3种酶活力值与4个因素之间关系的二次多项式回归方程为:
[0199]
y1=50.40+1.61x1+1.77x
2-0.22x3+0.81x4+1.18x1x2+0.69x1x
3-1.67x1x4+0.27x2x3+0.54x2[0200]
x4+0.12x3x
4-4.17x
12-4.88x
22-1.42x
32-2.55x
42
(1)
[0201]
y2=22.82+1.38x1+0.74x2+0.43x
3-0.038x
4-0.62x1x
2-0.42x1x
3-0.083x1x4+0.035x2x3+0.29
[0202]
x2x4+0.30x3x
4-1.49x
12-0.62x
22-1.38x
32-170x
42
(2)
[0203]
y3=91.76+2.34x1+2.61x2+1.15x
3-0.52x4+1.59x1x
2-0.61x1x
3-0.80x1x4+5.75
×
10-4
x2x
3-0.3
[0204]
8x2x4+1.36x3x
4-5.10x
12-2.23x
22-4.10x
32-5.45x
42
(3)
[0205]
式中:y1为cmc酶活(u/ml),y2为fpa酶活(u/ml),y3为木聚糖酶活(u/ml),x1、x2、x3、
x4分别为蛋白胨浓度、牛血清蛋白浓度、培养基起始ph、培养温度。
[0206]
由方差分析cmc酶活结果表的表6可知,该模型p<0.0001,表明该试验所选的模型差异极度显著,说明数据合理;该模型的f值为43.74,且仅有不足1%的概率不能用此模型来解释,说明该模型显著;该模型的均方差为23.64,方差为330.92,说明该模型显著;失拟项(lack of fit)p=0.1917>0.05,说明该模型的失拟项差异不显著,说明回归模型与试验数据的拟合度比较高,符合响应面优化的要求;从表7可看到,该模型的相关系数r2=0.9776,std.dev为0.74且偏差系数为1.63,说明该模型与实际拟合较好;校正决定系数radj=0.9553和pred r-squared=0.8840,说明二次项方程的可信度较高。
[0207]
表6box-behnken试验方差分析cmc酶活结果表
[0208][0209][0210]
通过方差分析f值可知,4个因素在实验过程对实验的影响顺序为:x2》x1》x4》x3,即牛血清蛋白浓度》蛋白胨浓度》培养温度》培养基起始ph,其中牛血清蛋白浓度和蛋白胨浓度起主要作用,培养温度和培养基起始ph起次要作用。若考虑二次项和交互项,则各因素对实验的影响顺序为:x
22
》x
12
》x
42
》x
32
》x1x4》x1x2》x1x3》x2x4》x2x3》x3x4。
[0211]
表7box-behnken试验方差分析cmc酶活的其他参数
[0212][0213]
接着对cmc酶活响应面结果进行分析,得到相应交互作用的等高线和响应面图。结果如图23所示。
[0214]
由图23可知,在ph为6.4,培养温度为37℃时,蛋白胨浓度和牛血清蛋白浓度的作用随添加量的增大,cmc酶活在一定范围内随其增大而升高,蛋白胨浓度超过2%,牛血清蛋白浓度超过1%后cmc酶活开始平稳减小。蛋白胨浓度和牛血清蛋白浓度的作用显著,曲面较陡。响应曲面等高线中可以看出最优条件取值范围在蛋白胨浓度1.9-2.3%,牛血清蛋白浓度0.9-1.3%之间。
[0215]
通过软件sas对二次多项式回归方程进行分析,首先求该方程的一阶偏导且要求一阶偏导等于0,就可以得到4个因素的最佳水平值,分别为x1=0.199207、x2=0.211417、x3=-0.003275、x4=0.115813,即蛋白胨浓度、牛血清蛋白浓度、培养基起始ph、培养温度分别是2.0996(%)、1.1057(%)、6.399、37.3474(℃),所得数据代入(1)式中可得最大cmc酶活的预测值y1=50.7932u/ml。
[0216]
由方差分析fpa酶活结果表表8和表9可知,该模型p《0.0001,表明试验所选模型差异极度显著;该模型f值为32.00,且仅有不足1%的概率不能用此模型来解释,也说明该模型显著;失拟项p=0.7248》0.05,失拟项的方差为1.33,表明失拟项差异不显著,说明回归模型与试验数据的拟合度比较高;该模型的均方根误差为0.39,响应均值为20.67,且r2=0.9697,说明该模型与实际拟合较好,数据准确度较高;偏差系数为1.88且校正决定系数radj=0.9394,说明方程可信度较高。
[0217]
表8box-behnken试验方差分析fpa酶活结果表
[0218][0219]
表9box-behnken试验方差分析fpa酶活的其他参数
[0220][0221]
由方差分析比较f值可知,4个因素在产酶培养基的发酵过程中对酶活的影响顺序为:x1》x2》x3》x4,即蛋白胨浓度》牛血清蛋白浓度》培养基起始ph》培养温度,其中蛋白胨浓度和牛血清蛋白浓度起主要作用,培养基起始ph和培养温度起次要作用。同时若考虑二次项和交互项的结果,发现二次项的i型平方和为33.14远远大于交互项的i型平方和为2.93;综上所述,则4个因素对实验的影响顺序为:x
42
》x
12
》x
32
》x
22
》x1x2》x1x3》x3x4》x2x4》x1x4》x2x3。对fpa酶活响应面结果进行分析,得到相应交互作用的等高线和响应面图。结果如图24所示。
[0222]
观察图24可知,在牛血清蛋白浓度为1%,培养温度为37℃时,蛋白胨浓度在1.5-2.1%范围内随着添加量的增大,cmc酶活随其升高。蛋白胨浓度和培养基ph的作用显著,曲面较陡。响应曲面等高线中可以看出最优条件取值范围在蛋白胨浓度2.0-2.4%,培养基ph6.3-6.5%之间。通过利用软件sas对回归方程基于编码的响应曲面进行典型分析,求(2)式的导并令结果等于0,可以得到4个因素的最佳临界值,分别为x1=0.3599、x2=0.431425、x3=0.110514、x4=0.026381,既蛋白胨浓度、牛血清蛋白浓度、培养基起始ph、培养温度分别是2.1780(%)、1.2157(%)、6.444、37.0791(℃),所得数据代入(2)式中可得最大fpa酶活的预测值y2=23.2528u/ml。由方差分析木聚糖酶活结果表的表10和11可得,该模型p《0.0001,表明试验所选模型差异极度显著;该模型f值为26.76,且仅有不足1%的概率不能用此模型来解释,也说明该模型显著;失拟项p=0.2025》0.05,且方差为18.43,表明失拟项差异不显著,说明回归模型与试验数据的拟合度比较高;模型的相关系数r2=0.9640,响应均值为83.87,均方根误差为1.24,说明该模型与实际拟合较好;校正决定系数radj=0.9280和预测决定系数为0.8138,说明方程可信度较高。
[0223]
表10box-behnken试验方差分析木聚糖酶活结果表
[0224][0225]
表11box-behnken试验方差分析木聚糖酶活的其他参数
[0226][0227]
由方差分析f值可知,4个因素在实验过程对实验的影响顺序为:x2》x1》x3》x4,即牛血清蛋白浓度》蛋白胨浓度》培养基起始ph》培养温度,其中牛血清蛋白浓度和蛋白胨浓度起主要作用,培养基起始ph和培养温度起次要作用。若考虑二次项和交互项的i型平方和以及f值,则4个因素对实验产酶的影响顺序为:x
42
》x
12
》x
22
》x
32
》x1x2》x3x4》x1x4》x1x3》x2x4》x2x3。对木聚糖酶活响应面结果进行绘图分析,得到相应的交互作用等高线和响应面图。结果如图25所示。由图25可知,在ph为6.4,培养温度为37℃时,蛋白胨浓度和牛血清蛋白浓度的作用随添加量的增大,cmc酶活在一定范围内呈梯度式的升高;当蛋白胨浓度超过2.4%,牛血清蛋白浓度超过1.3%后cmc酶活开始平稳减小。蛋白胨浓度和牛血清蛋白浓度的作用显著,曲面较陡。响应曲面等高线中可以看出最优条件取值范围在蛋白胨浓度2.0-2.3%,牛血清蛋白浓度1.0-1.4%之间。
[0228]
最后基于sas软件的编码数据整合分析求导,可以得到影响木聚糖酶活的4个最佳水平值,分别为x1=0.283208、x2=0.348730、x3=0.107619、x4=-0.066976,既蛋白胨浓度、牛血清蛋白浓度、培养基起始ph、培养温度分别是2.1416(%)、1.1744(%)、6.4430、36.7991(℃),所得数据代入(3)式中可得木聚糖酶活的最大预测值y3=92.63u/ml。综上所
述,在结合3种酶的最佳条件下,实际发酵产酶过程中选择蛋白胨浓度为2.14%、牛血清蛋白浓度为1.17%、培养基起始ph为6.44和培养温度为37℃,测得cmc酶活、fpa酶活和木聚糖酶活的平均值分别为49.8977u/ml、22.9328u/ml和90.7770u/ml,它们与预测值的相对误差分别为1.76%、1.38%和2.00%;且与优化产酶条件前的酶活相比,3种酶活分别约是优化前的9.712倍、6.602倍和9.634倍。
[0229]
(1)确定hf菌株产酶外部的最佳条件:转速为200r/min、培养基装液量为200ml/500ml、培养基起始ph为6.4、接种量为15%、发酵产酶时间为3d、培养温度为37℃。(2)优化了hf菌株的产酶培养基条件,最佳碳源为cmc-na、cmc-na最佳浓度为2%、最佳氮源为蛋白胨、蛋白胨最佳浓度为2%、dmf最佳浓度为0.5%、牛血清蛋白最佳浓度为1%、吐温-80最佳浓度为0.5%、peg6000最佳浓度为0.5%。zn
2+
、mg
2+
、cu
2+
对cmc酶活有促进作用,其中zn
2+
对3种酶活的促进作用最明显,而al
3+
、ca
2+
、k
+
、fe
3+
、fe
2+
对酶活有抑制作用,其中hg
+
、ag
+
完全抑制了酶的活性。mn
2+
对酶活没有影响。(3)在单因素的条件下,确定了以蛋白胨浓度、牛血清蛋白浓度、起始ph和培养温度作4因素3水平的响应面法优化,结果表明,选择蛋白胨浓度为2.14%、牛血清蛋白浓度为1.17%、培养基起始ph为6.44和培养温度为37℃,在此条件下,测得cmc、fpa和木聚糖酶活分别为49.8977u/ml、22.9328u/ml和90.7770u/ml,实测值与预测值的相差不大。与优化产酶条件前的酶活相比,3种酶活约是优化前的9.712倍、6.602倍和9.634倍。
[0230]
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之作一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1