一种创制抗矮花叶病毒病玉米种质的方法及其应用
本发明属于生物,具体涉及一种创制抗矮花叶病毒病玉米种质的方法及其应用。
背景技术:
1、基因编辑是一种对基因组进行定向精确修饰的技术,主要包括锌指核酸酶(zinc-finger nucleases,zfns),类转录激活因子效应物核酸酶(transcription activator-like effector nucleases,talens)和规律成簇的间隔短回文重复系统(clusteredregularly interspaced short palindromic repeats/crispr-associated nuclease,crispr/cas)。crispr/cas系统大致分为ⅰ型、ⅱ型和ⅲ型3类,其中ii型系统的组成比较简单,只需要cas9蛋白、tracrrna和crrna 3种成分即可发挥作用,且具有高效性和易操作性,成为目前应用最广泛的基因组编辑系统。ii型crispr/cas系统的特征蛋白为cas9,cas9蛋白具有ruvc和hnh两个核酸酶结构域,该结构域负责切割靶dna的两条链,从而造成靶dna双链断裂。其中hnh结构域负责切割与crrna互补的dna链,切割位点位于原型间隔序列毗邻基序(protospacer adjacent motif,pam)上游3bp处,ruvc结构域则负责切割非互补链,切割位点位于pam上游3–8bp处。cas9蛋白同时具有加工产生crrna以及切割外源核酸的功能。crrna通过碱基配对与tracrrna结合形成tracrrna/crrna复合物,研究者可以将tracrrna和crrna作为两种向导rna(grna)或者将两者融合在一起形成单向导rna(single guiderna,sgrna)。sgrna能够与cas9核酸内切酶结合并将cas9引导至基因组上对靶位点进行切割。crispr/cas9系统使目标基因dna产生双链断裂,激活细胞内的dna损伤修复机制,进而产生缺失、敲入等变异。目前,crispr/cas9系统已被迅速地用于拟南芥、烟草、高粱、水稻、小麦、玉米等不同植物基因组的定向编辑研究中,并且获得了较高的诱导突变率和可稳定遗传的基因组编辑植株。
2、玉米(zea mays l.)是重要的粮食和饲料作物,也是现代食品和化学工业的原料,在我国农业生产和经济发展中占有重要地位。玉米矮花叶病毒((maize dwarf mosaicvirus,mdmv)是在世界范围内广泛分布的一种病毒,该病毒于1965年在美国首次发现,从20世纪90年代开始,玉米矮花叶病毒病有逐年加重的趋势,对玉米生产构成了严重威胁。生产中,若大面积种植易感病玉米品种,在对蚜虫活动有利的气候条件下,即5月至7月凉爽、降雨不多,蚜虫将迁飞到玉米田吸食传毒,大量繁殖后辗转为害,从而造成该病流行玉米矮花叶病毒病发病前期为害叶片,后期病株出现不能抽穗或虽能抽穗,但花粉粒萌芽率降低、花粉萌芽后芽管长度变短,不能进入子房进行授精,并使果穗结实不良。一般发病早的植株结穗率减少,千粒重下降。轻病田减产10%-20%,重病田减产30%~50%,部分地块甚至绝产。该病毒主要在雀麦、牛鞭草等寄主上越冬,是重要初侵染源,病毒主要靠带病毒蚜虫吸食叶片汁液而传播扩散,玉米整个生育期均可发病。感染矮花叶病毒的玉米植株在幼苗期基部细脉间出现许多椭圆形褪绿小点,呈虚线状排列,以后发展成实线,病部继续呈不规则状扩大,不受叶脉限制,在粗脉间形成许多黄色条纹,病叶褪色失绿呈斑驳花叶症状。病部可包围健部,形成许多大、小不同略呈圆形的绿斑,叶片变黄、棕、紫或干枯。重病株的苞叶、叶鞘、雄花穗有时出现褪绿斑,植株矮小,其高度有时只为健株的1/3-1/2,不能抽穗或迟抽穗而不结实。病株茎细,根部不发达或萎缩。感染此病后一般可减产10%左右,感病重的地块,产量造成严重损失。
3、mdmv是一种无包膜的单链rna病毒,属于马铃薯y病毒科(potyviridae)马铃薯y病毒属(potyvirus),病毒粒子呈弯曲线状,大小约(430~750)nm×(12~15)nm.现已报道的玉米矮花叶病毒病病毒有mdmv-a、mdmv-b、md、mv-c、mdmv-d、mdmv-e、mdmv-f、mdmv-g、mdmv-o及mdmv-ksi株系,其中mdmv-a和mdmv-b两个株系研究较多,a株系主要侵染玉米和约翰逊草,b株系仅仅侵染玉米。在自然条件下,mdmv主要由机械传播或由蚜虫以非持久性方式传播,并且mdmv还可以种传,同时,该病的发生程度与植株上的蚜量关系密切,玉米矮花叶病毒病的发生受品种抗病性、介体蚜虫数量、田间杂草数量、耕作制度以及气候条件等诸多因素影响,所以此病害防治难度较大。目前,生产上主要采取以农业措施如调整播期、加强田间管理等为主,药剂防治为辅的综合防治措施。但是,上述防治措施存在费工费时、易造成环境污染且防治效果差等不足,培育和种植抗病品种是防治玉米矮花叶病毒病的有效途径。
技术实现思路
1、本发明所要解决的技术问题是如何调控植物对矮花叶病毒病的抗性。
2、为了解决现有技术存在的问题,本发明提供了蛋白质或调控基因的表达物质或调控所述蛋白质活性或含量的物质在调控植物对矮花叶病毒病抗性中的应用。
3、本发明提供的蛋白质或调控基因的表达物质或调控所述蛋白质活性或含量的物质在下述任一种中的应用:
4、1)蛋白质或调控基因的表达物质或调控所述蛋白质活性或含量的物质在调控植物矮花叶病毒病抗性中的应用;
5、2)蛋白质或调控基因的表达物质或调控所述蛋白活性或含量的物质在制备调控植物矮花叶病毒病抗性的产品中的应用;
6、3)蛋白质或调控基因的表达物质或调控所述蛋白活性或含量的物质在培育对矮花叶病毒病抗性改变的植物中的应用;
7、4)蛋白质或调控基因的表达物质或调控所述蛋白活性或含量的物质在制备培育对矮花叶病毒病抗性改变的植物的产品中的应用;
8、5)蛋白质或调控基因的表达物质或调控所述蛋白活性或含量的物质在植物育种中的应用;
9、所述蛋白质为如下任一所示蛋白质:
10、a1)氨基酸序列为seq id no.2的蛋白质;
11、a2)将seq id no.2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的蛋白质;
12、a3)a1)-a2)中任一所限定的氨基酸序列具有75%以上同一性且具有相同功能的蛋白质;
13、a4)a1)-a3)中任一所限定的蛋白质的末端连接标签后得到的融合蛋白。
14、上述a1)所述蛋白质的名称为rab gdp解离抑制因子(rab gdp dissociationinhibitor alpha,本文简称zmgdiα)。
15、为了使a1)中的蛋白质便于纯化或检测,可在由序列表中seq id no.2所示的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接标签蛋白。
16、上述蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
17、所述标签蛋白包括但不限于:gst(谷胱甘肽巯基转移酶)标签蛋白、his6标签蛋白(his-tag)、mbp(麦芽糖结合蛋白)标签蛋白、flag标签蛋白、sumo标签蛋白、ha标签蛋白、myc标签蛋白、egfp(增强型绿色荧光蛋白)、ecfp(增强型青色荧光蛋白)、eyfp(增强型黄绿色荧光蛋白)、mcherry(单体红色荧光蛋白)或avitag标签蛋白。
18、本领域普通技术人员可以很容易地采用已知的方法,例如定向进化或点突变的方法,对本发明的编码蛋白质zmgdiα的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的蛋白质zmgdiα的核苷酸序列75%或75%以上同一性的核苷酸,只要编码蛋白质zmgdiα且具有蛋白质zmgdiα功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
19、上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。
20、本文中,同一性是指氨基酸序列或核苷酸序列的同一性。可使用国际互联网上的同源性检索站点测定氨基酸序列的同一性,如ncbi主页网站的blast网页。例如,可在高级blast2.1中,通过使用blastp作为程序,将expect值设置为10,将所有filter设置为off,使用blosum62作为matrix,将gap existence cost,per residue gap cost和lambda ratio分别设置为11,1和0.85(缺省值)并进行检索一对氨基酸序列的同一性进行计算,然后即可获得同一性的值(%)。
21、本文中,所述80%以上的同一性可为至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。
22、本文中,所述90%以上的同一性可为至少90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。
23、上述应用中,所述蛋白质来源于玉米(zea mays l.)。
24、本文中,调控所述蛋白质活性和/或含量的物质可为调控基因表达的物质,所述基因编码所述蛋白质zmgdiα。
25、本文中,所述调控基因表达的物质可为进行如下6种调控中至少一种调控的物质:1)在所述基因转录水平上进行的调控;2)在所述基因转录后进行的调控(也就是对所述基因的初级转录物的剪接或加工进行的调控);3)对所述基因的rna转运进行的调控(也就是对所述基因的mrna由细胞核向细胞质转运进行的调控);4)对所述基因的翻译进行的调控;5)对所述基因的mrna降解进行的调控;6)对所述基因的翻译后的调控(也就是对所述基因翻译的蛋白质的活性进行调控)。
26、本文中,所述调控可为上调或增强或提高;所述调控也可为下调或减弱或降低。
27、本文中,所述调控所述蛋白质的编码基因的表达可为抑制或降低或下调所述编码基因表达。所述抑制或降低或下调所述编码基因表达可通过基因敲除或基因沉默实现。
28、所述基因敲除(gene knock-out)是指通过基因编辑技术使特定靶基因失活的现象。基因敲除通过dna序列的改变使特定靶基因失活,包括且不限于基于锌指核酸酶(zinc-finger nucleases,zfn),转录激活因子样效应因子核酸酶(transcription activator-like effector nucleases,talen)和crispr/cas系统,crispr(clustered regulatoryinterspaced short palindromic repeat)即成簇的规律间隔的短回文重复序列,是基因组中一个含有多个短重复序列的位点,cas9蛋白在rna的介导下能够对crrna–tracrrna识别的靶序列进行切割。
29、所述基因沉默是指在不损伤原有dna的情况下使基因不表达或低表达的现象。基因沉默以不改变dna序列为前提,使基因不表达或低表达。基因沉默可发生在两种水平上,一种是由于dna甲基化、异染色质化以及位置效应等引起的转录水平的基因沉默,另一种是转录后基因沉默,即在基因转录后的水平上通过对靶标rna进行特异性抑制而使基因失活,包括反义rna、共抑制(co-suppression)、基因压抑(quelling)、rna干扰(rnai)和微小rna(mirna)介导的翻译抑制等。
30、上述应用中,所述调控基因的表达物质或调控所述蛋白质活性或含量的物质可为与前文所述蛋白质相关的生物材料,所述生物材料可为下述任一种:
31、c1)编码前文所述蛋白的核酸分子;
32、c2)含有c1)所述核酸分子的表达盒;
33、c3)含有c1)所述核酸分子的重组载体、或含有c2)所述表达盒的重组载体;
34、c4)含有c1)所述核酸分子的重组微生物、或含有c2)所述表达盒的重组微生物、或含有c3)所述重组载体的重组微生物;
35、c5)含有c1)所述核酸分子的转基因植物细胞系、或含有c2)所述表达盒的转基因植物细胞系;
36、c6)含有c1)所述核酸分子的转基因植物组织、或含有c2)所述表达盒的转基因植物组织;
37、c7)含有c1)所述核酸分子的转基因植物器官、或含有c2)所述表达盒的转基因植物器官;
38、e1)抑制或降低或沉默前文所述蛋白编码基因表达的核酸分子;
39、e2)含有e1)所述核酸分子的表达盒;
40、e3)含有e1)所述核酸分子的重组载体、或含有e2)所述表达盒的重组载体;
41、e4)含有e1)所述核酸分子的重组微生物、或含有e2)所述表达盒的重组微生物、或含有e3)所述重组载体的重组微生物;
42、e5)含有e1)所述核酸分子的转基因植物细胞系、或含有e2)所述表达盒的转基因植物细胞系;
43、e6)含有e1)所述核酸分子的转基因植物组织、或含有e2)所述表达盒的转基因植物组织;
44、e7)含有e1)所述核酸分子的转基因植物器官、或含有e2)所述表达盒的转基因植物器官。
45、上述应用中,c1)所述核酸分子可为如下任一所示的dna分子,
46、d1)核苷酸序列是seq id no.3所示的dna分子;
47、d2)编码区序列是序列表中seq id no.1所示的dna分子;
48、d3)与d1)或d2)限定的核苷酸序列具有90%或90%以上同一性,且编码前文所述蛋白的dna分子;
49、d4)在严格条件下与d1)或d2)限定的核苷酸序列杂交,且编码前文所述蛋白的dna分子。
50、本文所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如grna、mrna、sirna、shrna、sgrna、mirna或反义rna。
51、c2)和e2)所述的表达盒是指能够在宿主细胞中表达所述基因的dna,该dna不但可包括启动基因转录的启动子,还可包括终止基因转录的终止子。进一步,所述表达盒还可包括增强子序列。
52、可用于本发明的启动子包括但不限于:组成型启动子,组织、器官和发育特异的启动子和诱导型启动子。启动子的例子包括但不限于:花椰菜花叶病毒的组成型启动子35s;来自西红柿的创伤诱导型启动子,亮氨酸氨基肽酶("lap",chao等人(1999)plant physiol120:979-992);来自烟草的化学诱导型启动子,发病机理相关1(pr1)(由水杨酸和bth(苯并噻二唑-7-硫代羟酸s-甲酯)诱导);西红柿蛋白酶抑制剂ii启动子(pin2)或lap启动子(均可用茉莉酮酸曱酯诱导);热休克启动子(美国专利5,187,267);四环素诱导型启动子(美国专利5,057,422);种子特异性启动子,如谷子种子特异性启动子pf128(cn101063139b(中国专利2007 1 0099169.7)),种子贮存蛋白质特异的启动子(例如,菜豆球蛋白、napin,oleosin和大豆beta conglycin的启动子(beachy等人(1985)embo j.4:3047-3053))。它们可单独使用或与其它的植物启动子结合使用。此处引用的所有参考文献均全文引用。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子(nos终止子)、花椰菜花叶病毒camv 35s终止子、tml终止子、豌豆rbcs e9终止子和胭脂氨酸和章鱼氨酸合酶终止子(参见,例如:odell等人(i985)nature 313:810;rosenberg等人(1987)gene,56:125;guerineau等人(1991)mol.gen.genet,262:141;proudfoot(1991)cell,64:671;sanfacon等人genes dev.,5:141;mogen等人(1990)plant cell,2:1261;munroe等人(1990)gene,91:151;ballad等人(1989)nucleic acids res.17:7891;joshi等人(1987)nucleic acidres.,15:9627)。
53、本文所述载体是本领域技术人员公知的,包括但不限于:质粒、噬菌体(如λ噬菌体或m13丝状噬菌体等)、黏粒(即柯斯质粒)、ti质粒或病毒载体。
54、可用现有的植物表达载体构建含有zmgdiα基因的重组表达载体。所述植物表达载体包括但不限于如双元农杆菌载体和可用于植物微弹轰击的载体等。所述植物表达载体还可包含外源基因的3'端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3'端,如包括但不限于农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂合成酶nos基因)、植物基因(如大豆贮藏蛋白基因)3'端转录的非翻译区均具有类似功能。
55、为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入包括但不限于可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、荧光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除草剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
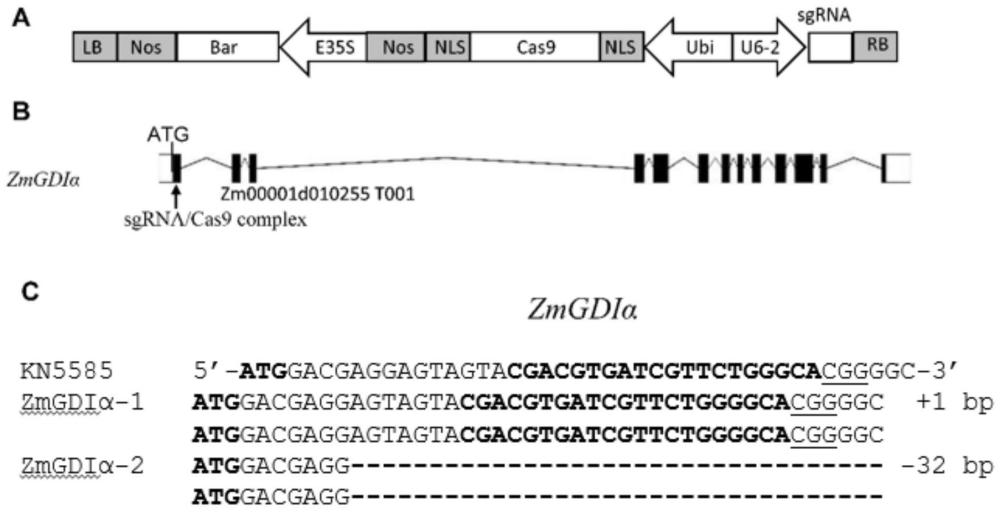
56、上述应用中,所述重组载体为重组载体cpb-dm,是由19720bp组成的双链dna。
57、重组表达载体cpb-dm的结构描述如下:在出发载体cpb—cas9的限制性内切酶hindiii插入含有sgrna的表达盒序列(seq id no.4),保持空载体cpb—cas9的其他序列不变得到的重组载体。
58、本发明还提供了一种提高植物对矮花叶病毒病抗性的方法,所述方法包括步骤p,所述步骤p为抑制或降低或沉默目的植物中前文所述蛋白的活性和/或含量,或/和,抑制或降低或沉默前文所述蛋白的编码基因的表达量,来提高植物矮花叶病毒病抗性。
59、本发明还提供了一种抑制植物矮花叶病毒病抗性的方法,所述方法包括步骤m,所述步骤m为增强、提高或上调目的植物中前文所述蛋白的活性和/或含量,或/和,增强、提高或上调前文所述蛋白的编码基因的表达量,来抑制植物对矮花叶病毒病抗性。
60、上述方法中,所述降低目的植物中所述蛋白质zmgdiα的编码基因的表达量和/或活性可为利用基因突变、基因敲除、基因编辑或基因敲减技术使目的植物基因组中所述蛋白质zmgdiα的编码基因活性下降或失活。
61、在一个具体的实施例中,所述抑制植物矮花叶病毒病抗性的方法可包括如下步骤:
62、a)用crispr/cas9系统对受体植物中的所述zmgdiα蛋白的编码基因zmgdiα进行基因编辑,且使所述zmgdiα基因发生突变导致翻译蛋白提前终止,得到目的植物;
63、b)所述目的植物自交,得到纯合植物,即为目的植物,所述矮花叶病毒病抗性提高的植物对矮花叶病毒病抗性能力高于所述受体植物。
64、所述zmgdiα基因是如下的dna分子:
65、d1)核苷酸序列是seq id no.3所示的dna分子;
66、d2)编码区序列是序列表中seq id no.1所示的dna分子;
67、d3)与d1)或d2)限定的核苷酸序列具有90%或90%以上同一性,且编码前文所述蛋白的dna分子;
68、d4)在严格条件下与d1)或d2)限定的核苷酸序列杂交,且编码前文所述蛋白的dna分子。
69、本发明提供一种培育矮花叶病毒病抗性提高的植物的方法,包括抑制或降低或沉默目的植物中上述蛋白的编码基因的表达和/或上述蛋白的含量和/或活性,或/和抑制或降低或沉默上述蛋白质的编码基因的活性和/或含量,得到矮花叶病毒病抗性提高的植物,所述矮花叶病毒病抗性提高的植物矮花叶病毒病抗性高于所述受体植物。
70、在本发明的一个实施方案中,所述培育矮花叶病毒病抗性提高植物的育种方法包括如下步骤:
71、(1)构建抑制或降低或沉默前文所述蛋白质的编码基因的重组表达载体;
72、(2)将步骤(1)构建的重组表达载体转入受体植物中,获得矮花叶病毒病抗性提高的植物。
73、上述方法中,所述抑制或降低或沉默目的植物中所述蛋白质zmgdiα的编码基因的表达量和/或活性可为利用基因突变、基因敲除、基因编辑或基因敲减技术使目的植物基因组中所述蛋白质zmgdiα的编码基因活性下降或失活。
74、上述方法中,所述基因敲除通过crispr/cas9系统实现。
75、上述方法中,所述crispr/cas9系统基因编辑的靶点为序列1第214到第233位。
76、上述方法中,敲除目的玉米所述蛋白质的编码基因可为将玉米基因组中的序列3所示的所述蛋白质的编码基因进行下述至少一种突变:
77、1)用5’-atggacgaggagtacgacgtgatcgttctggggcacggggctcaagga-3’替换玉米基因组dna中的所述蛋白质的编码基因中的5’-atggacgaggagtacgacgtgatcgttctgggcacggggctcaagga-3’,从而将编码zmgdiα蛋白的基因敲除。
78、2)用5’-acgaggaagga-3’替换玉米基因组dna中的所述蛋白质的编码基因中的5’-atggacgaggagtacgacgtgatcgttctgggcacggggctcaagga-3’,从而将编码zmgdiα蛋白的基因敲除。
79、本发明中,所述植物育种的目的可包括培育矮花叶病毒病抗性提高的植物。
80、前文所述的蛋白质和/或所述的生物材料也属于本发明要求保护的范围。
81、本文中,所述植物可为如下任一种:
82、n1)单子叶植物:
83、n2)禾本目植物;
84、n3)禾本科植物;
85、n4)玉蜀黍属植物;
86、n5)玉米。
87、本研究利用基因编辑技术,敲除玉米感矮花叶病毒病的基因zmgdiα,快速创制抗矮花叶病毒病的玉米种质。通过基因编辑技术敲除感病基因节约了成本,缩短了育种年限,高效精准的定向改造玉米抗性。通过基因编辑技术获得了两个zmgdiα纯合突变体,通过人工接种mdmv,发现突变体的抗病性显著高于野生型;同时,在未接种条件下,wt和突变体植株在生长或发育方面没有可见的表型差异。鉴于该基因在水稻、小麦、高梁等禾本科作物中的保守机制,为其抗病育种策略提供了重要借鉴。
- 还没有人留言评论。精彩留言会获得点赞!