一种基于NMR技术筛选粗枝云杉体胚萌发相关标记代谢物的方法与流程
本发明涉及组织培养环节的培养技术、仪器分析技术和数据分析技术领域,具体地说,涉及nmr技术评价干化对粗枝云杉体胚萌发影响的方法。
背景技术
代谢组学是一种通过分析生物发育过程中的大量代谢物,并研究它们在不同生理和遗传条件下的浓度差异,进而辅助基因表达和蛋白定量的新技术。任何环境胁迫、病理及其它因素对植物生命系统的影响均会引起植物体内代谢物的浓度或代谢流发生变化,而且大多数代谢途径不是孤立存在的,因此代谢组学作为反映某一植物生命过程中整体水平的代谢物质动态变化过程,正在广泛应用于植物功能基因组学、分子表型、分子病理学等领域的研究。
粗枝云杉(piceaasperatamast.)是松科云杉属植物,我国特有树种,分布范围广、材质优良且适应性强。它不仅是重要的用材树种,也是川西高原、甘肃南部和陕西西南部等高山地区森林演替的顶极群落和地带性植被的建群种。目前,粗枝云杉的体细胞胚胎发生技术已基本建立,但其中一些关键环节的内在调控机理还未知,尤其是萌发前体胚内部物质的改变和积累对体胚萌发的作用。
干化是体胚萌发前的一种处理方式,能促进体胚的萌发。研究表明,干化主要是使体胚由“形态学”成熟转化为“生理学”成熟,待转入萌发培养基后能获得一个较高的萌发率。目前,对干化促进体胚萌发的分子机理尚不清楚。基于粗枝云杉干化处理前后子叶和胚根代谢组结果分析,对揭示干化的作用机理和发现潜在的标记代谢物均具有重要的意义。但是,目前有关云杉体胚干化处理前后代谢组学的研究国内外尚无报道,尤其是基于nmr技术对云杉代谢组检测的样品处理、提取、数据分析和标记代谢物筛选缺乏可靠的方法。
技术实现要素:
本发明的目的是提供一种基于nmr技术对粗枝云杉体胚代谢组进行测定和分析,并用来评价干化处理对粗枝云杉体胚萌发能力影响的方法。
为了实现本发明目的,本发明提供了一种基于nmr技术筛选粗枝云杉体胚萌发标记代谢物的方法,包括以下步骤:用“滤纸法”对粗枝云杉体胚进行干化处理,切取未干化体胚的子叶和胚根以及干化后体胚的子叶和胚根,不同体胚组织代谢物的萃取、冻干及复溶、nmr检测,代谢组数据分析和标记代谢物的筛选。
根据本发明的方法,所述干化处理方法为:将粗枝云杉分化成熟的体细胞胚置于滤纸上,然后在24±1℃、16h光周期和15-18μmol/m2·s的光照强度条件下培养7-14天,得到干化后的体胚。
进一步的,所述干化处理方法具体为:将分化成熟的体胚平铺在1-4层滤纸上,再把带有体胚的滤纸放入小培养皿(35mm)中,每个小培养皿分散放置30-40个胚,然后将不带盖的小培养皿放入大培养皿(90mm)中,最后在大培养皿中加入10ml无菌水(不超过小培养皿三分之二的高度),加盖密封。在24±1℃、16h光周期和15-18μmol/m2·s的光照强度条件下,处理7-14天。
其中,所述的“滤纸法”干化处理中,放置粗枝云杉体细胞胚的培养容器内垫有滤纸的层数优选为2-3层,更优选为2层;光照强度优选为15μmol/m2·s;干化处理时间优选为14天。
根据本发明的方法,所述分离步骤包括在体视显微镜下切下干化处理后体胚的子叶和胚根,同时切取未干化体胚的子叶和胚根;将切取的目标组织置于冷冻管中,冷冻保存。
进一步的,本发明的方法中子叶切割的位置位于展开子叶往下0.1~0.3mm处,胚根切割的位置位于胚根尖端往上1~1.5mm处,共4组材料,每组材料6个重复。取样后准确称量单份样品并及时用液氮速冻,温度稳定后放入-80℃冰箱中保存备用。
根据本发明的方法,所述体胚组织代谢物的萃取方法为:向低温研磨后的样品中加入50%(v/v)的甲醇溶液作为提取剂,在93μm条件下进行超声波萃取,4秒超声,3秒间歇,共循环16次;离心后取上清液于离心管中并用氮气吹干;
所述冻干及复溶方法包括:向冻干的样品中加入纯水,涡旋后加入acdss试剂,再次涡旋混匀后,离心取上清液,作为nmr检测的提取液。
进一步的,体胚组织代谢物的萃取方法为:低温研磨后直接向样品管中加入1000μl提取剂(50%的甲醇溶液,v/v),涡旋1分钟;冰水浴,在93μm振幅条件下进行超声波萃取,4秒超声,3秒间歇,共循环16次;然后在4℃、13000rpm条件下离心15分钟;取澄清透明上清液600μl于离心管中用氮气吹干提取物。
所述冻干及复溶方法为:将提取物放入-80℃冰箱中冰冻1小时,再将提取物放入到冻干机中冻干1小时;加入450μl纯水,涡旋1分钟;取50μlacdss试剂(内标,4.088mm)于上述离心管中,并涡旋10秒钟(使滤液与acdss试剂充分混匀);在4℃、13000rpm条件下离心2分钟;取480μl上清液于核磁管中,作为nmr检测的样品。
nmr检测使用的nmr谱仪为agilentdd2600mhz三重共振光谱仪。具体检测参数条件如下表所示:
本发明还提供了粗枝云杉体胚材料代谢组的数据处理方法,具体如下:
(1)数据预处理:将1hnmr自由感应衰减(freeinductiondecay,fid)信号导入到chenomxnmrsuit(version8.0,chenomx,edmonton,canada)软件中,自动进行傅立叶转换,调整相位,校正基线。以dss-d6峰(0.0ppm)作为全部谱图化学位移的标准,并对其进行反转卷积操作,调整谱图峰形(csi)。根据1hnmr谱图中信号的相关信息(如化学位移、峰形、半峰宽、耦合裂分等),以dss-d6峰的浓度和谱峰面积为标准,结合chenomx自带数据库对24张样本谱图的信号逐一比对分析。然后把代谢物和对应的绝对值浓度导出到excel表格中,即得到了一个代谢物变量矩阵。
(2)化学计量学分析:把变量矩阵用样品的最初质量进行归一化后作为源数据进行后续的主成分分析(pca)和正交偏最小二乘法-判别分析(opls-da)。从复杂的数据中筛选出样品间的差异代谢标记物。通过对干化前后子叶和胚根的代谢物差异分析,筛选到精氨酸、葡萄糖和奎宁酸可作为判断粗枝云杉体胚萌发能力强弱的标记代谢物。
本发明还提供了与粗枝云杉体胚萌发能力相关的标记代谢物,所述标记代谢物包括精氨酸、葡萄糖和/或奎宁酸。
本发明还提供了nmr技术在筛选粗枝云杉体胚萌发标记代谢物中的应用,进一步的,所述代谢物为精氨酸、葡萄糖和/或奎宁酸。
本发明还提供了精氨酸、葡萄糖和/或奎宁酸在作为粗枝云杉体胚萌发标记代谢物中的应用,进一步的,上述代谢物在粗枝云杉体胚萌发过程中的含量上升。
本发明还提供了精氨酸、葡萄糖和/或奎宁酸作为标记代谢物在指示粗枝云杉体胚萌发强弱的应用,当上述精氨酸、葡萄糖和/或奎宁酸含量上升时,则粗枝云杉体胚萌发能力较强。
本发明首次将粗枝云杉干化前后体胚的子叶和胚根分开进行检测和分析。
本发明通过对粗枝云杉体胚材料的代谢组研究,建立一套高效、快速和重复性好的粗枝云杉体胚材料代谢组样品前处理、提取、数据处理和差异代谢物筛选的方法,能有效揭示干化处理影响粗枝云杉体胚萌发密切相关的代谢信息。本发明虽以粗枝云杉为例,但对整个针叶树种的体胚材料代谢组的检测、标记代谢物筛选和评价干化处理对其体胚乃至种胚萌发能力影响,均提供了良好的应用前景。
附图说明
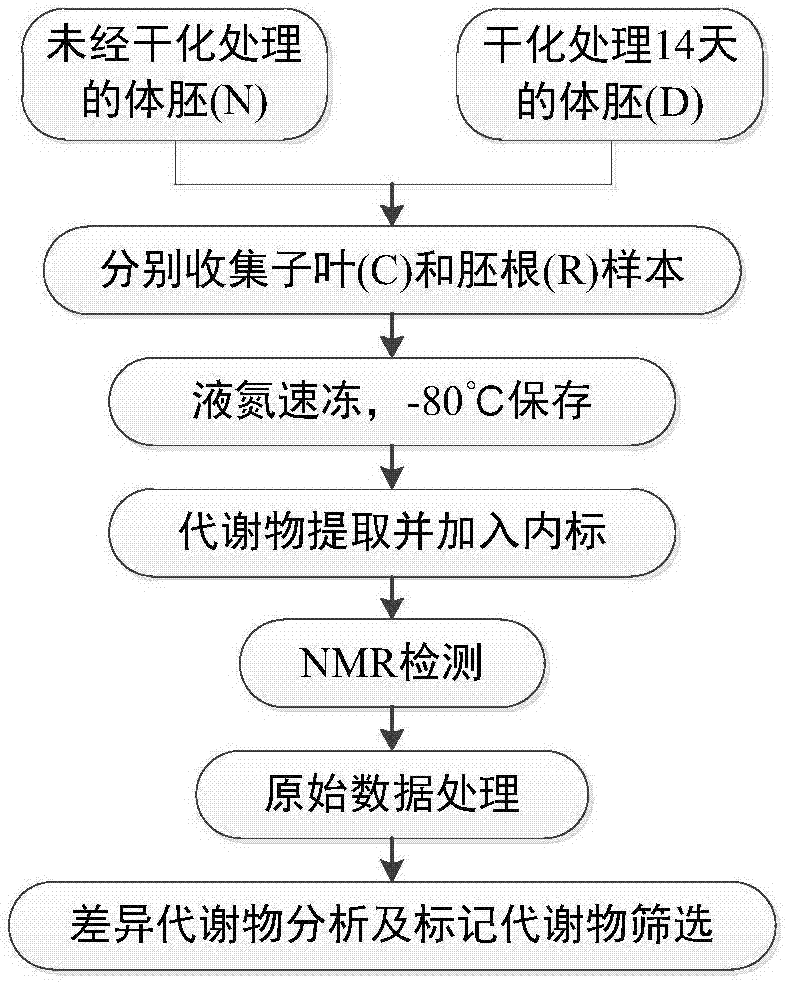
图1.本发明采用的基于nmr技术筛选粗枝云杉体胚萌发标记代谢物的工艺流程图。
图2.本发明实施例1中干化处理前后的体细胞胚。其中,a和c:未干化的体细胞胚;b和d:干化后的体细胞胚;c和d:分别标注了取样的未干化子叶(non-desiccatedcotyledon,nc)、未干化胚根(non-desiccatedradicle,nr)、干化子叶(desiccatedcotyledon,dc)和干化胚根(desiccatedradicle,dr)部位。比例尺:a和b,10mm;c和d,500μm。
图3.本发明实施例4中干化前后不同组织代谢组的pca得分图。a:四组样品未干化子叶(nc)、未干化胚根(nr)、干化子叶(dc)和干化胚根(dr)的整体pca得分图;b:dc与nc的分析结果;c:dr与nr的分析结果;d:dc与nr的分析结果;e:dr与nc的分析结果。
图4.本发明实施例4中干化前后不同组织代谢组的opls-da得分图。a:dc与nc的分析结果;b:dr与nr的分析结果;c:dc与nr的分析结果;d:dr与nc的分析结果。
图5.本发明实施例5中组间差异代谢物的汇总结果,其中精氨酸(arginine)、葡萄糖(glucose)和奎宁酸(quinicacid)的含量在干化组中均显著高于未干化组。
具体实施方式
以下实施例用于说明本发明,但不用来限制本发明的范围。若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段,所用原料均为市售商品。
实施例中涉及的标准品为acdss(anachrocertifieddssstandardsolution)–4.088mm,来自anachro;超声波破碎仪:sonicsvcx130;冻干机:labconcofreezone2.5;微型漩涡混合仪:xw-80a,购自上海泸西分析仪器厂;离心机:eppendorfcentrifuge5415r和5425r;nmr谱仪:agilentdd2600mhzspectrometerequippedwithatriple-resonancecryoprobe。
本发明的基于nmr技术筛选粗枝云杉体胚萌发标记代谢物的工艺流程图见图1。
实施例1不同体胚组织的收集
(1)粗枝云杉体胚的干化处理
用镊子挑选分化成熟、形态饱满的粗枝云杉体胚平放至干化容器中,按“滤纸法”进行干化处理,具体为:将分化成熟的体胚平铺在两层滤纸上,再把带有体胚的滤纸放入小培养皿(35mm)中,每个小培养皿中分散放置30个胚,然后将所述小培养皿放置于更大的培养皿(90mm)中,在大培养皿中加入10ml无菌水,最后加盖封口,在24℃、16h光周期和15μmol/m2·s的光照强度下干化处理14天,得到干化后的体胚(图2)。
(2)干化前后粗枝云杉体胚不同组织的收集和保存
在体视显微镜下,用无菌解剖刀分别切取未干化体胚和干化体胚的子叶和胚根,子叶切割的位置位于展开子叶往下约2mm处,胚根切割的位置位于胚根尖端往上约1.2mm处,切取的子叶和胚根分别置于冻存管中并准确称量,每组样品6个生物学重复。用液氮彻底速冻后放入-80℃冰箱中保存备用。
实施例2基于nmr对粗枝云杉体胚材料代谢组样品的前处理
(1)粗枝云杉体胚材料代谢物的萃取
低温研磨后,向样品管中加入1000μl提取剂(50%的甲醇溶液,v/v),涡旋1分钟;冰水浴,在93μm振幅条件下进行超声波萃取,4秒超声,3秒间歇,共循环16次;在4℃、13000rpm条件下,离心15分钟;取澄清透明上清液600μl于离心管中用氮气吹干样品。
(2)代谢物的冻干及复溶
将样品放入-80℃冰箱中冰冻1小时,再将样品放入到冻干机中继续冻干1小时;加入450μl纯水,涡旋1分钟;取50μlacdss试剂(内标,4.088mm)于上述离心管中,并涡旋10秒钟(使滤液与acdss试剂充分混匀);在4℃、13000rpm条件下离心2分钟;取480μl上清液于核磁管中,作为nmr检测的粗枝云杉体胚不同组织代谢组样品。
实施例3基于nmr对粗枝云杉体胚代谢组样品的检测
nmr检测使用的nmr谱仪为agilentdd2600mhz三重共振光谱仪。具体检测条件见表1:
表1核磁数据采集参数
实施例4粗枝云杉体胚不同组织代谢组数据分析
(1)数据预处理
将1hnmr自由感应衰减(freeinductiondecay,fid)信号导入到chenomxnmrsuit(version8.0,chenomx,edmonton,canada)软件中,自动进行傅立叶转换,调整相位,校正基线。以dss-d6峰(0.0ppm)作为全部谱图化学位移的标准,并对其进行反转卷积操作,调整谱图峰形(csi)。根据1hnmr谱图中信号的相关信息(如化学位移、峰形、半峰宽、耦合裂分等),以dss-d6峰的浓度和谱峰面积为标准,结合chenomx自带数据库对24张样本谱图的信号逐一比对分析。然后把代谢物和对应的绝对值浓度导出到excel表格中,即得到了一个代谢物浓度变量矩阵。最后对数据使用paretoscaling法进行归一化处理。
(2)主成分分析(pca)
使用omicsharetools对归一化得到的四组样品的数据进行模式识别多变量分析,对数据的第一和第二主成分进行模型拟合。得到了四组样品(nc、nr、dc和dr)的整体分析结果和两两比较(dcvsnc、drvsnr、dcvsnr和drvsnc)的分析结果(图3)。
(3)化学计量学分析
把变量矩阵做为源数据进行组间(dcvsnc、drvsnr、dcvsnr和drvsnc)正交偏最小二乘法-判别分析(opls-da),得到组间的区分结果(图4)。
实施例5干化前后粗枝云杉体胚不同组织的差异代谢物分析和标记代谢物筛选
(1)干化前后不同组织间差异代谢物分析
从复杂的数据中判别出组间(dcvsnc、drvsnr、dcvsnr和drvsnc)差异代谢物。本发明采用opls-da模型中第一主成分的vip(variableimportanceintheprojection)值,并结合学生氏t检验(student’st-test)的p值筛选差异代谢物。vip≥1,p<0.05的被认为是差异代谢物。
①干化子叶与未干化子叶的差异代谢物(dcvsnc)
与未干化子叶(nc)相比,干化子叶(dc)中有8个代谢物的含量发生了显著变化(表2),且均为上调。如表2所示,其中差异倍数最高的是奎宁酸,上调至原来的46.71倍;有3种糖类物质也显著上升,其中葡萄糖上调最多,上调至原来的16.09倍;精氨酸作为植物体内n/c最高的氨基酸之一,也显著上调,上调至原来的6.95倍。
表2干化子叶(dc)和未干化子叶(nc)之间的显著差异代谢物
②干化胚根与未干化胚根的差异代谢物(drvsnr)
与未干化胚根(nr)相比,干化胚根(dr)中有5种代谢物的含量发生了显著变化(表3)。精氨酸、果糖、葡萄糖和奎宁酸显著上调,分别上调至原来的6.64、4.02、5.78和3.10倍;蔗糖显著下调,降至原来的0.25。
表3干化胚根(dr)和未干化胚根(nr)之间的显著差异代谢物
③干化子叶与未干化胚根的差异代谢物(dcvsnr)
与未干化胚根相比,干化子叶(dc)中有7种物质显著增加,1种物质显著降低(表4)。如表4所示,奎宁酸、葡萄糖、精氨酸、蔗糖、肌醇、谷氨酸盐和4-氨基丁酸分别上调至原来的3.21、2.05、3.81、2.38、1.97、1.58和4.80倍;而谷氨酰胺下调至原来的0.40。
表4干化子叶(dc)和未未干化胚根(nr)之间的显著差异代谢物
④干化胚根与未干化子叶的差异代谢物(drvsnc)
与未干化子叶(nc)相比,干化胚根(dr)中有7中物质的含量发生了显著变化(表5)。其中,有6种物质的含量上调,精氨酸、胆碱、果糖、葡萄糖、奎宁酸和肌醇分别上调至原来的12.12、1.83、19.84、45.37、45.09和2.00;而蔗糖的含量下调至原来的0.29。
表5干化胚根(dr)和未干化子叶(nc)之间的显著差异代谢物
(2)体胚萌发能力强弱的标记代谢物筛选
通过对干化组(dc和dr)与未干化组(nc和nr)代谢组的比较分析,发现精氨酸、葡萄糖和奎宁酸的含量在干化组中均高于未干化组(图5)。因此,筛选到精氨酸、葡萄糖和奎宁酸这三种差异代谢物可作为判断粗枝云杉体胚萌发能力强弱的标记代谢物,且当干化处理后含量上升,则表明粗枝云杉体胚的萌发能力增强。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之作一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
- 还没有人留言评论。精彩留言会获得点赞!