具有改善的因子VIII表达的血友病A基因编辑的制作方法
具有改善的因子viii表达的血友病a基因编辑
技术领域
1.本文提供的披露内容涉及用于离体和体内治疗血友病a的材料和方法。另外,提供用于基因编辑以调节凝血蛋白诸如因子viii(fviii)的表达、功能或活性的材料和方法。
背景技术:
2.血友病a(hema)是由fviii基因(f8)中的遗传缺陷引起的,该遗传缺陷导致血液中fviii蛋白水平低或检测不到。这导致在组织损伤部位处不能有效地形成凝块,从而导致出血失控,如果不治疗,则可能致命。替代缺失的或非功能性的fviii蛋白对于hema受试者而言是一种有效的治疗方法并且是现行护理标准。然而,蛋白替代疗法需要频繁地静脉施用fviii蛋白,这在成人中是不方便的,在儿童中成问题,费用高昂(>$200,000/年),并且如果不严格遵循治疗方案,则可能导致破裂出血事件。
3.非常需要彻底治愈血友病a。虽然使用腺相关病毒(aav)的基于病毒的基因疗法已在临床前动物模型和人类受试者中显示出一些前景,但它具有许多缺点。例如,报道的基于aav的基因疗法使用由肝脏特异性启动子驱动的fviii编码序列,该启动子包封在aav病毒衣壳内(通常使用血清型aav5、aav8或aav9或aavrh10等)。一般而言,用于基因疗法的aav病毒将包装的编码序列盒递送至转导细胞的细胞核中,在这些细胞中该盒几乎只保持为游离型并且正是治疗性编码序列的游离型拷贝产生治疗性蛋白。aav不具有将包封的dna整合到宿主细胞基因组中的机制。因为治疗性编码序列保持为游离体,当宿主细胞分裂时它不协同复制,因此可能从子细胞中丢失。已证明,当含有aav游离体的肝脏细胞被诱导分裂时,aav基因组不复制而是被稀释。因此,预计基于aav的基因疗法在肝脏尚未达到成人大小的儿童中不会有效。因为目前的疗法是不充分的,所以对于成人和儿童的hema迫切需要新的有效且持久的或长期的治疗。
4.fviii最初表达为具有结构域结构a1
‑
a2
‑
b
‑
a3
‑
c1
‑
c2的蛋白。该蛋白通过大体积、高度糖基化的b结构域的蛋白水解裂解而活化,留下重链(a1
‑
a2)和轻链(a3
‑
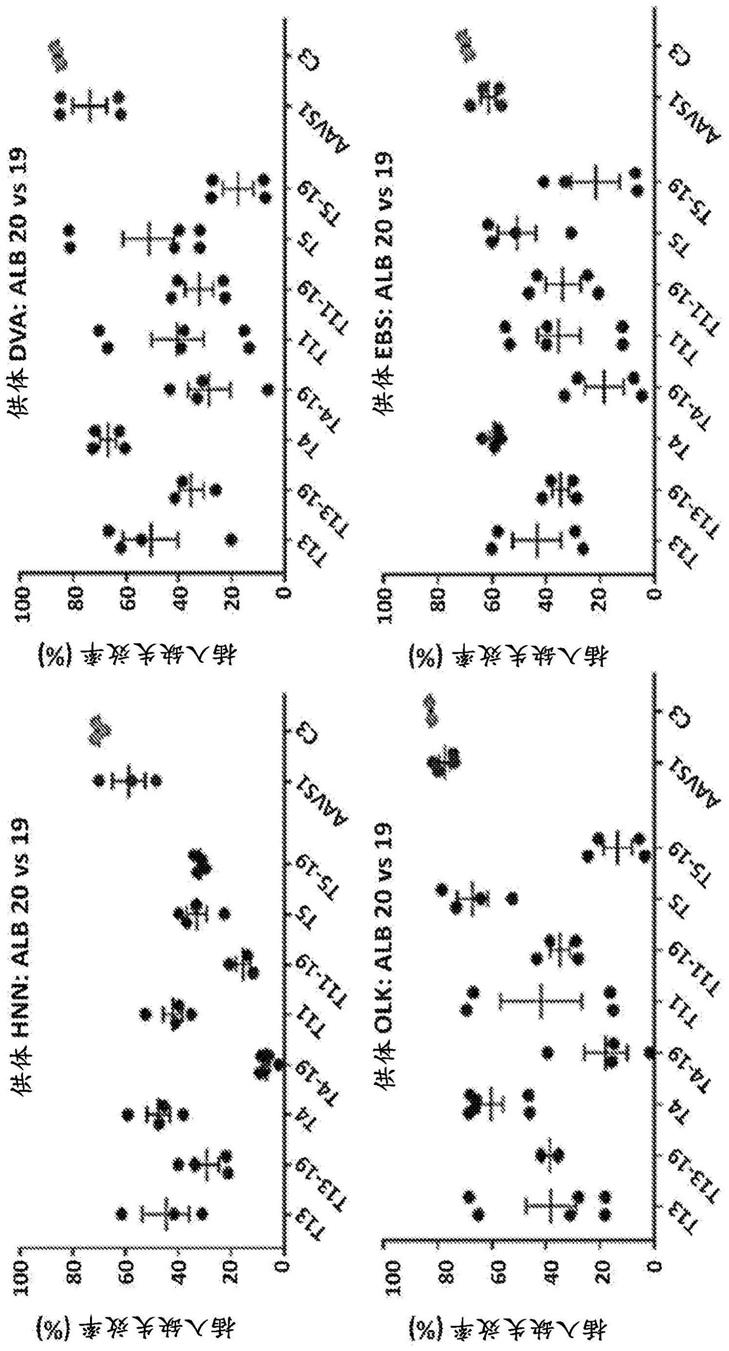
c1
‑
c2)异二聚体。fviii蛋白的b结构域不是生物活性所需的。从fviii编码序列中去除大b结构域对于实现可靠包装到用于体内递送的aav载体中是必需的。然而,去除含有多达18个n
‑
连接的糖基化位点的b结构域导致fviii蛋白的分泌受损。因此,迫切需要可以有效和高效表达的改良形式的fviii。
技术实现要素:
5.申请人已经发现了可用于补充缺陷型f8基因,从而引起功能性fviii蛋白表达的基因编辑组合物和方法。因此,本文提供的发明包括用于改变宿主细胞dna序列的系统和组合物,用于改变宿主细胞基因组的方法,用于插入提供改善的表达的合成因子viii编码序列的方法和系统,可以施用于受试者的具有提供改善的表达的合成因子viii编码序列的细胞,用于治疗血友病a的方法,以及实现任何前述方面的试剂盒。
6.在一个方面,本文提供了用于改变宿主细胞dna序列的系统,该系统具有:dna内切
核酸酶或编码该dna内切核酸酶的核酸;具有与宿主细胞基因座互补的间隔区序列的向导rna(grna)或编码该grna的核酸;以及具有编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物具有0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
7.在另一方面,提供了编辑宿主细胞中的基因组的方法,该方法包括向该细胞提供以下物质:具有与宿主细胞基因座互补的间隔区序列的grna或编码该grna的核酸;dna内切核酸酶或编码该dna内切核酸酶的核酸;以及具有编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白具有b结构域替代物,该b结构域替代物具有0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
8.在另一方面,提供了一种细胞,其中该细胞的基因组包括编码合成fviii蛋白的dna,该合成fviii蛋白具有b结构域替代物,其中该b结构域替代物具有0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
9.在另一方面,提供了通过将如上所述的具有编码合成fviii蛋白的dna的细胞施用于受试者来治疗该受试者的血友病a的方法。
10.在另一方面,提供了通过向受试者中的细胞提供以下物质来治疗该受试者的血友病a的方法:具有与宿主细胞基因座互补的间隔区序列的grna或编码该grna的核酸;dna内切核酸酶或编码该dna内切核酸酶的核酸;以及具有编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白具有b结构域替代物,该b结构域替代物具有0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
11.另一方面,本文提供了一种试剂盒,该试剂盒包含上述系统的一种或多种要素,并且进一步包含使用说明书。
12.在另一方面,本文提供了具有编码合成fviii蛋白的多核苷酸序列的核酸,其中该合成fviii蛋白具有b结构域替代物,该b结构域替代物具有0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
13.在另一方面,本文提供了通过向受试者中的细胞提供以下物质来增加受试者中的fviii量的方法,其中该受试者具有第一fviii血清水平:具有与宿主细胞基因座互补的间隔区序列的grna或编码该grna的核酸;dna内切核酸酶或编码该dna内切核酸酶的核酸;以及具有编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白具有b结构域替代物,该b结构域替代物具有0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
附图说明
14.通过参考下面的详细描述和附图,将获得对本披露某些特征和优点的理解,下面的详细描述阐述了说明性实施例,其中利用了本披露的原理,并且附图中:
15.图1描绘了流体动力学注射编码fviii供体模板的5种质粒,接着lnp递送cas9 mrna和sgrna后小鼠血液中的fviii水平。
16.图2描绘了注射包封fviii供体模板pcb099和pcb102的aav8病毒,接着在4周后施用包封spcas9 mrna和grna malbt1的lnp的小鼠血液中的fviii水平。在注射lnp后10天测量fviii水平。
17.图3描绘了用四种不同的fviii供体质粒通过hdi给药,接着用包封spcas9和
malbt1 grna的lnp给药的血友病a小鼠血液中的fviii活性。
18.图4描绘了在lnp给药后11天和28天血友病a小鼠血液中的fviii活性。小鼠在lnp给药前4周接受了2
×
10
12 vg/kg的aav8病毒。
19.图5描绘了流体动力学注射质粒pcb1007(n=7只小鼠)、pcb1019(n=7)和pcb1020(n=6)并且眶后注射包封malbt1 grna和cas9 mrna的lnp的小鼠血液中的fviii活性。在lnp给药后第6天和第9天测量fviii。
20.图6描绘了流体动力学注射质粒pcb1007(n=7只小鼠)、pcb1025(n=7)和pcb1026(n=6)并且眶后注射包封malbt1 grna和cas9 mrna的lnp的小鼠血液中的fviii活性。在lnp给药后第6天和第9天测量fviii。
21.图7描绘了来自四个供体的人原代肝细胞中向导rna t4、t5、t11和t13(靶向人白蛋白内含子1)的切割效率的结果,比较了19个碱基的靶序列与20个碱基的靶序列。
22.图8描绘了具有0至7个n
‑
连接的聚糖基序和不同密码子优化的fviii供体盒的固有表达效率(fviii活性除以靶向整合频率)。
23.图9描绘了其中b结构域替代物含有0、1、3、5或6个聚糖的fviii构建体。
24.图10描绘了在lnp给药后11天血友病a小鼠血液中的fviii活性。
25.图11描绘了在lnp给药后28天血友病a小鼠血液中的fviii活性。
26.图12描绘了具有0、1、3、5或6个n
‑
连接的聚糖基序的fviii供体盒的固有表达效率(fviii活性除以靶向整合频率)。
具体实施方式
27.rna向导内切核酸酶编辑提供优于例如慢病毒基因治疗方法的优点。然而,在编辑方案中插入大序列可能是有问题的,例如,原因是大序列可能难以包装以进行递送,或者与短序列相比,可能难以制造。一些蛋白需要n
‑
连接的糖基化位点的存在才能从表达它们的细胞中正确分泌。n
‑
糖基化位点的共有氨基酸序列是n
‑
x
‑
t/s,其中x是除脯氨酸以外的任何残基。将聚糖添加到n(天冬酰胺)残基(k.f.medzihradszky,meth mol biol[分子生物学方法](2008)446:293
‑
316)。申请人已经发现,此类蛋白中n
‑
连接的糖基化位点的数目可以大幅减小或甚至消除,从而减小蛋白编码序列的大小,而对转录、翻译或分泌没有不利影响。例如,申请人已经发现,工程化fviii编码序列的b结构域以减小或消除糖基化位点的数量可以减小用于基因编辑的fviii序列的大小,而不明显影响所得到的经工程化的(合成的)fviii的转录、翻译或分泌,同时产生具有fviii功能的经工程化的fviii蛋白。此外,最小化添加到b结构域缺失的fviii中的n
‑
聚糖位点的数目将最小化产生抗体或t细胞的新型表位的风险,从而降低新型fviii蛋白可能在受试者中诱导免疫应答的风险。本披露尤其提供了用于基因编辑以通过基因组编辑来调节细胞中凝血蛋白诸如fviii的表达、功能或活性的组合物和方法。本披露尤其还提供了用于离体和体内治疗患有血友病a的受试者的组合物和方法。具体而言,本发明提供了提供改善的整合和改善的表达的基因组编辑方法和系统,以及能够改善血友病a的合成fviii编码序列和蛋白。
[0028]
定义
[0029]
除非另有定义,否则本文使用的所有技术和科学术语具有与要求保护的主题所属领域中的技术人员通常所理解的相同含义。应当理解,详细描述仅是示例性和说明性的,并
且不限制所要求保护的任何主题。在本技术中,除非另有明确说明,否则单数的使用包括复数。如说明书中所用,除非上下文另外明确指出,否则单数形式“一”、“一种(个)”和“该”包括复数指示物。在本技术中,除非另有说明,否则“或”的使用意指“和/或”。此外,术语“包括(including)”以及其他形式(诸如“包括(include)”、“包括(includes)”和“包括(included)”)的使用不是限制性的。
[0030]
尽管可以在单个实施例的背景下描述本披露的特征,但是这些特征也可以单独地或以任何合适的组合来提供。相反,尽管为了清楚起见,本文中可以在单独实施例的背景下描述本披露,但是本披露也可以在单个实施例中实现。本文引用的任何公开的专利申请和任何其他公开的参考文献、文件、手稿和科学文献出于任何目的通过援引并入本文。在有冲突的情况下,将以本说明书(包括定义)为准。另外,材料、方法和实例仅为说明性的,并非旨在为限制性的。
[0031]
如本文所用,范围和量可以表示为“约”特定值或范围。“约”还包括确切的量。因此,“约5μl”意指“约5μl”,还意指“5μl”。通常,术语“约”包括预期在实验误差范围诸如
±
1%、
±
2%、
±
3%、
±
5%或
±
10%以内的量。
[0032]
当本文中呈现数值范围时,可以考虑到,在该范围的下限和上限之间的每个中间值、该范围的上限和下限值以及该范围内的所有规定值都涵盖在本披露的范围内。本披露还涵盖了该范围的下限和上限内的所有可能的子范围。
[0033]
术语“多肽”、“肽”和“蛋白”在本文中可互换使用,以表示通过肽键相互连接的氨基酸残基的线性系列,该系列可以包括蛋白、多肽、寡肽、肽及其片段。蛋白可以由天然存在的氨基酸和/或合成的(例如,经修饰的或非天然存在的)氨基酸组成。如本文所用,术语“氨基酸”或“肽残基”可以指天然存在的氨基酸和合成氨基酸两者。术语“多肽”、“肽”和“蛋白”包括融合蛋白,包括但不限于有或无n端甲硫氨酸残基,具有异源氨基酸序列的融合蛋白,具有异源和同源前导序列的融合蛋白;免疫标记蛋白;具有可检测的融合伴侣的融合蛋白,例如,包括荧光蛋白、β
‑
半乳糖苷酶、荧光素酶等作为融合伴侣的融合蛋白。此外,应该注意,在氨基酸序列的开始或末端的破折号表示连接一个或多个氨基酸残基的另一序列的肽键或连接端羧基或端羟基的共价键。然而,不应该将不存在破折号视为意指不存在连接端羧基或端羟基的这种肽键或共价键,因为在表示氨基酸序列时常规地将此省略。
[0034]
术语“多核苷酸”、“寡核苷酸”、“寡聚物”(oligomer)、“寡聚物”(oligo)、“编码序列”和“核酸”是指不同长度的聚合形式的核苷酸(核糖核苷酸或脱氧核糖核苷酸)。因此,这些术语包括但不限于单链、双链或多链dna或rna、基因组dna、cdna、dna
‑
rna杂合体,或具有嘌呤和嘧啶碱基或其他天然、化学或生物化学修饰的、非天然或衍生化的核苷酸碱基的聚合物。
[0035]
术语“功能等效”或作为“功能等效物”是指但不限于具有源自本文披露的化合物的结构或序列并且其结构或序列与本文披露的那些足够相似,使其具有相同或相似的活性和效用或者,基于这种相似性,本领域的技术人员预计会表现出与参考化合物相同或相似的活性和效用的任何分子诸如核酸或蛋白。获得功能等效物、“衍生物”或“变体”的修饰可以包括,例如,一个或多个核酸或氨基酸残基的添加、缺失和/或取代。
[0036]
蛋白的功能等效物或功能等效物片段可以具有一个或多个保守性氨基酸取代。术语“保守性氨基酸取代”是指将一个氨基酸取代为另一个具有与原氨基酸相似的特性的氨
基酸,即一个氨基酸被来自同一分组的另一氨基酸取代。保守性氨基酸分组如下:
[0037]
分组氨基酸名称脂肪族gly、ala、val、leu、ile含羟基或巯基/硒ser、cys、thr、met环状pro芳香族phe、tyr、trp碱性his、lys、arg酸性及其酰胺asp、glu、asn、gln
[0038]
可以在预定的肽或其片段的任何位置引入保守性取代。然而,也可能需要引入非保守性取代,特别是但不限于在任何一个或多个位置引入非保守性取代。导致形成肽的功能等效片段的非保守性取代将例如在极性、电荷、空间体积和/或与其他蛋白或核酸的结合方面不同,而保持功能等效物或变体片段的抗凝功能性。
[0039]
通过在比较窗口中比较两个最佳比对的序列来测定序列同一性的百分比,其中多核苷酸或多肽序列在比较窗口中的一部分与参考序列(没有添加或缺失)相比,可能具有添加或缺失(即空位)以便进行两个序列的最佳比对。在一些情况下,通过以下方式来计算百分比:确定两个序列中出现相同的核酸碱基或氨基酸残基的位置数目以得到匹配位置数目,将匹配位置数目除以比较窗口中的位置总数并将结果乘以100,得到序列同一性的百分比。可以使用,例如,alignx(包括在vectro nti中,基于clustalw(http://www.clustal.org/clutal2/),使用标准参数(例如:空位开放罚分=15;空位延伸罚分=6.6;空位分离罚分范围=8)确定序列同一性。
[0040]
在两个或更多个核酸或多肽序列的背景下,术语“相同”或百分比“同一性”是指正如使用以下序列比较算法之一或通过手动比对和目视检查所测量的,出于最大一致性在比较窗口或指定区域上进行比较和比对时,相同的或具有规定百分比的相同氨基酸残基或核苷酸(例如,在规定区域,例如整个多肽序列或多肽单个结构域中具有60%、65%、70%、75%、80%、85%、90%、95%、98%或99%同一性)的两个或更多个序列或子序列。于是称此类序列是“基本上相同的”。该定义也指测试序列的互补序列。
[0041]
本文可互换使用的术语“互补”或“基本上互补”意指核酸(例如dna或rna)具有使其能够与另一个核酸以序列特异性的反向平行方式非共价结合,即形成沃森
‑
克里克(watson
‑
crick)碱基对和/或g/u碱基对的核苷酸序列(即核酸与互补核酸特异性结合)。如本领域已知的,标准的沃森
‑
克里克碱基配对包括:腺嘌呤(a)与胸腺嘧啶(t)配对,腺嘌呤(a)与尿嘧啶(u)配对,以及鸟嘌呤(g)与胞嘧啶(c)配对。
[0042]“编码”特定rna的dna序列是转录为rna的dna核酸序列。dna多核苷酸可以编码翻译成蛋白的rna(mrna),或者dna多核苷酸可以编码不翻译成蛋白的rna(例如trna、rrna或向导rna;也称为“非编码”rna或“ncrna”)。“蛋白编码序列或编码特定蛋白或多肽的序列”是在体外或体内当置于适当调控序列的控制下时转录成mrna(在dna的情况下)并翻译(在mrna的情况下)成多肽的核酸序列。
[0043]
如本文所用,“密码子”是指一起形成dna或rna分子中的遗传密码单元的三个核苷酸的序列。如本文所用,术语“密码子简并性”是指遗传密码中容许核苷酸序列变化而不影响所编码的多肽的氨基酸序列的性质。
[0044]
术语“密码子优化的”或“密码子优化”是指用于转化合适宿主的核酸分子的基因或编码区,是指核酸分子的基因或编码区中反映宿主生物体的密码子使用而不改变dna编码的多肽的密码子改变。这种优化包括用该生物体的基因中使用频率更高的一个或多个密码子来替代至少一个或多于一个或大量密码子。密码子使用表可轻易获得,例如,在www.kazusa.or.jp/codon/上可用的“密码子使用数据库”(2019年1月30日访问)。通过利用关于每种生物体中的密码子使用或密码子偏好的知识,本领域的普通技术人员可以将这些频率应用于任何给定的多肽序列,并产生编码该多肽,但是使用给定物种的最佳密码子的密码子优化编码区的核酸片段。密码子优化编码区通过本领域技术人员已知的方法来设计。
[0045]
当提到例如细胞、核酸、蛋白或载体使用时,术语“重组”或“工程化”表明该细胞、核酸、蛋白或载体已经通过实验室方法进行修饰或者是实验室方法的结果。因此,例如,重组或工程化蛋白包括通过实验室方法产生的蛋白。重组或工程化蛋白可以包括在天然(非重组或野生型)形式的蛋白中未发现的氨基酸残基,并且可以包括已经过修饰(例如标记)的氨基酸残基。该术语可以包括对肽、蛋白或核酸序列的任何修饰。此类修饰包括:肽、蛋白或核酸序列的任何化学修饰;肽或蛋白中一个或多个氨基酸的添加、缺失和/或取代;以及核酸序列中一个或多个核酸的添加、缺失和/或取代。
[0046]
术语“基因组dna”或“基因组序列”是指生物体基因组的dna,包括但不限于细菌、真菌、古菌、植物或动物基因组的dna。
[0047]
如本文所用,“转基因”、“外源基因”和“外源序列”是指不存在于细胞基因组中但是人工引入基因组中(例如通过基因组编辑)的核酸序列或基因。
[0048]
如本文所用,“内源基因”或“内源序列”是指天然存在于细胞基因组中,无需经由任何人工手段引入的核酸序列或基因。
[0049]
术语“载体”或“表达载体”是指复制子,诸如质粒、噬菌体、病毒或粘粒,其上可以附着另一个dna区段,例如“插入物”,以使该附着区段在细胞中复制。
[0050]
术语“表达盒”是指具有与启动子可操作地连接的dna编码序列的载体。“可操作地连接”是指其中所述组分处于容许它们以其预期方式起作用的关系的并置。例如,如果启动子影响编码序列的转录或表达,则该启动子是与该编码序列可操作地连接的。术语“重组表达载体”和“dna构建体”在本文可互换使用,是指具有载体和至少一个插入物的dna分子。重组表达载体通常是出于使插入物表达和/或繁殖的目的或为了构建其他重组核苷酸序列而产生的。核酸可以或可以不与启动子序列可操作地连接,并且可以或可以不与dna调控序列可操作地连接。
[0051]
术语“调控序列”包括启动子、增强子和其他表达控制元件(例如,聚腺苷酸化信号)。此类调控序列是本领域已知的并且例如在goeddel;gene expression technology:methods in enzymology[基因表达技术:酶学方法]185,加州圣地亚哥学术出版社(academic press,san diego,ca)(1990)中有描述。调控序列包括那些在许多类型的宿主细胞中指导核苷酸序列组成型表达的调控序列以及那些仅在某些宿主细胞中指导核苷酸序列表达的调控序列(例如,组织特异性调控序列)。
[0052]
当外源dna例如重组表达载体已引入细胞内部时,该细胞已经被这种dna“遗传修饰”或“转化”或“转染”。外源dna的存在导致永久或暂时的遗传变化。转化dna可以或可以不
整合(共价连接)到细胞基因组中。具有治疗活性(例如治疗血友病a)的经遗传修饰(或转化或转染)的细胞可以使用并称为“治疗细胞”。
[0053]
在分子诸如肽片段的背景下使用的术语“浓度”是指在给定体积的溶液中存在的分子的量,例如分子的摩尔数。
[0054]
术语“急性期蛋白”是指响应于炎症在表达或血清浓度方面变化的蛋白。急性期蛋白的实例包括白蛋白、转铁蛋白、转甲状腺素蛋白、纤维蛋白原、抗凝血酶等。
[0055]
术语“个体”、“受试者”和“宿主”是指需要诊断、治疗或疗法的任何受试者。在一些方面,受试者是哺乳动物。在一些方面,受试者是人。在一些方面,受试者是人类患者。在一些方面,受试者患有或怀疑患有血友病a和/或具有血友病a的一种或多种症状。在一些方面,受试者是被诊断为在诊断时或之后具有血友病a风险的人。在一些情况下,可以根据基因组中内源性fviii基因或靠近fviii基因的基因组序列中存在一个或多个可以影响fviii基因表达的突变来确定血友病a风险的诊断。
[0056]
提到疾病或病状时使用的术语“治疗”意指实现与折磨个体的病状相关的症状的缓解,其中缓解在广义上用于指与所治病状(例如,血友病a)相关的参数例如症状幅度的降低。治疗还包括这样的情形,其中病理病状或至少与之相关的症状受到完全抑制,例如防止其发生或完全消除,使得宿主不再遭受该病状或至少不再遭受该病状特有的症状。因此,治疗包括:(i)预防(即,降低临床症状发展的风险,包括使临床症状不发展,例如,预防疾病进展),和(ii)抑制(即,阻止临床症状的发展或进一步发展,例如,减轻或完全抑制活动性疾病)。
[0057]
术语“有效量”、“药物有效量”和“治疗有效量”意指当施用给患有特定病状的受试者时足以提供所需效用的组合物的量。在离体治疗血友病a的背景下,术语“有效量”是指预防或缓和血友病a的至少一种或多种体征或症状所需的治疗细胞群或其后代的量,并且涉及足以提供所需作用,例如治疗受试者的血友病a症状的具有治疗细胞或其后代的组合物的量。因此,术语“治疗有效量”是指治疗细胞或具有治疗细胞的组合物,当施用于需要治疗的受试者(诸如患有血友病a或处于血友病a风险中的受试者)时,足以促进特定作用的量或数目。有效量还包括足以预防或延迟疾病症状发展,改变疾病症状过程(例如但不限于,减缓疾病症状的进展),或逆转疾病症状的量或数目。在体内治疗受试者(例如患者)的血友病a或在体外培养的细胞中进行基因组编辑的背景下,有效量是指编辑受试者体内细胞或体外培养的细胞的基因组所需的用于基因组编辑的组分,诸如grna、供体模板和/或定点多肽(诸如dna内切核酸酶)的量。应当理解,对于任何给定的情况,本领域的普通技术人员可以确定适当的“有效量”。
[0058]
如本文所用,术语“药物组合物”和“药物”是指与本发明的细胞(表达合成fviii蛋白)和/或本发明的系统的一种或多种组分(即,grna或编码grna的核酸,dna内切核酸酶或编码dna内切核酸酶的核酸,和/或编码合成因子viii蛋白的供体模板)组合的药学上可接受的赋形剂。
[0059]
如本文所用,术语“药学上可接受的赋形剂”是指提供药学上可接受的载剂、添加剂或稀释剂以将一种或多种目标化合物施用于受试者的任何合适的物质。“药学上可接受的赋形剂”可以涵盖称为药学上可接受的稀释剂、药学上可接受的添加剂和药学上可接受的载剂的物质。
[0060]
术语“合成fviii”是指与野生型人因子viii的a和c结构域具有大幅序列同一性(genbank:cad97566.1;g.a.vehar等人,nature[自然](1984)312:337
‑
42),但具有b结构域替代物而不是野生型b结构域的蛋白。在本发明的一个实施例中,合成fviii蛋白的a和c结构域的序列与a和c结构域的野生型序列具有80%、90%、95%、98%或99%的同一性。在一些实施例中,b结构域替代物是具有约10至约200个氨基酸的任何序列的多肽。在一些实施例中,b结构域替代物具有约20至约100个氨基酸。在一些实施例中,b结构域替代物可具有少于40个氨基酸(例如,具有3至40个氨基酸的任何数目的氨基酸)和1
‑
9个当表达时提供b结构域替代物糖基化的n
‑
连接的糖基化位点。b结构域替代物可进一步包括蛋白酶裂解位点,使得合成的fviii蛋白可以与野生型蛋白相同的方式裂解成重链和轻链。在一个实施例中,除了1
‑
9个n
‑
连接的糖基化(“聚糖”)位点之外,b结构域替代蛋白序列还包括来自野生型b结构域的n端和c端的1
‑
10个氨基酸。在一个实施例中,b结构域替代蛋白序列具有1
‑
6个聚糖位点。在一个实施例中,b结构域替代蛋白序列具有1
‑
5个聚糖位点。在一个实施例中,b结构域替代蛋白序列具有1
‑
4个聚糖位点。在一个实施例中,b结构域替代蛋白序列具有2
‑
4个聚糖位点。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
369、371和373中任一个的序列,或与seq id no:362
‑
369、371和373中任一个的序列具有至少80%、90%、95%、98%或99%同一性的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
366、371和373中任一个的序列,或与seq id no:362
‑
366、371和373中任一个的序列具有至少80%、90%、95%、98%或99%同一性的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
364、371和373中任一个的序列,或与seq id no:362
‑
364、371和373中任一个的序列具有至少80%、90%、95%、98%或99%同一性的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
363中任一个的序列,或与seq id no:362
‑
363中任一个的序列具有至少80%、90%、95%、98%或99%同一性的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
369中任一个的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
366中任一个的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
364中任一个的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:362
‑
363、371和373中任一个的序列。在一个实施例中,b结构域替代蛋白序列具有seq id no:371或373中任一个的序列。
[0061]
术语“安全港基因座”是指宿主细胞基因组内可以被修饰(例如,通过裂解,或通过插入供体序列)而不破坏细胞的代谢或调控(例如,通过引起细胞凋亡、增殖等),和/或不会对其他细胞(非编辑细胞)或作为整体的宿主生物体产生风险或不利影响(例如,通过无意中引起生长因子的过表达等)的基因座。在一些实施例中,安全港基因座是在宿主细胞中表达的基因座。在一些实施例中,安全港基因座是白蛋白基因座、纤维蛋白原基因座、aavs1基因座或转铁蛋白基因座。
[0062]
核酸
[0063]
靶向基因组的核酸或向导rna
[0064]
本披露提供了靶向基因组的核酸,该核酸可以将相关多肽(例如,定点多肽,诸如dna内切核酸酶)的活性指向靶核酸内的特定靶序列。在一些实施例中,靶向基因组的核酸是rna。靶向基因组的rna在本文中称为“指导rna”或“grna”。向导rna至少具有可以与目标靶核酸序列和crispr重复序列杂交的间隔区序列。在ii型系统中,grna还具有称为
tracrrna序列的第二rna。在ii型grna中,crispr重复序列和tracrrna序列彼此杂交形成双链体。在v型grna中,crrna形成双链体。在两种系统中,双链体都结合定点多肽,使得grna和定点多肽形成复合物。靶向基因组的核酸由于其与定点多肽缔合而为复合物提供了靶标特异性。因此,靶向基因组的核酸指导定点多肽的活性。
[0065]
在一些实施例中,靶向基因组的核酸是双分子grna。双分子grna具有两条rna链。第一链在5’至3’方向上具有任选的间隔区延伸序列、间隔区序列和最小crispr重复序列。第二条链具有最小tracrrna序列(与最小crispr重复序列互补)、3’tracrrna序列以及任选的tracrrna延伸序列。在一些实施例中,靶向基因组的核酸是单分子grna。ii型系统中的单分子grna(sgrna)在5’至3’方向上具有任选的间隔区延伸序列、间隔区序列、最小crispr重复序列、单分子向导接头、最小tracrrna序列、3’tracrrna序列和任选的tracrrna延伸序列。任选的tracrrna延伸序列可以具有为grna贡献附加功能(例如,稳定性)的元件。单分子指导接头将最小crispr重复序列和最小tracrrna序列连接起来以形成发夹结构。任选的tracrrna延伸序列具有一个或多个发夹。v型系统中的sgrna在5’至3’方向上具有最小crispr重复序列和间隔区序列。
[0066]
举例说明,crispr/cas/cpf1系统中使用的grna或其他较小的rna可以通过如下所述和本领域描述的化学手段容易地合成。随着化学合成程序的不断发展,通过诸如高效液相色谱法(hplc,其避免使用诸如page等凝胶)等程序纯化此类rna随着多核苷酸长度显著增加超过约一百个核苷酸而趋于更具挑战性。用于产生更大长度的rna的一种方法是产生两个或更多个连接在一起的分子。更长的rna(诸如编码cas9或cpf1内切核酸酶的那些)更容易酶促产生。如本领域所述,可以在rna的化学合成和/或酶促生成期间或之后引入rna修饰,例如,增强稳定性、降低先天免疫应答的可能性或程度、和/或增强其他属性的修饰。
[0067]
间隔区延伸序列
[0068]
在靶向基因组的核酸的一些实施例中,间隔区延伸序列可以改变活性,提供稳定性和/或提供用于修饰靶向基因组的核酸的位置。间隔区延伸序列可以改变中靶或脱靶活性或特异性。在一些实施例中,提供了间隔区延伸序列。间隔区延伸序列的长度可以大于1、5、10、15、20、25、30、35、40、45、50、60、70、80、90、100、120、140、160、180、200、220、240、260、280、300、320、340、360、380、400、1000、2000、3000、4000、5000、6000、或7000个或更多个核苷酸。间隔区延伸序列的长度可以为约1、5、10、15、20、25、30、35、40、45、50、60、70、80、90、100、120、140、160、180、200、220、240、260、280、300、320、340、360、380、400、1000、2000、3000、4000、5000、6000、或7000个或更多个核苷酸。间隔区延伸序列的长度可以小于1、5、10、15、20、25、30、35、40、45、50、60、70、80、90、100、120、140、160、180、200、220、240、260、280、300、320、340、360、380、400、1000、2000、3000、4000、5000、6000、7000个或更多个核苷酸。在一些实施例中,间隔区延伸序列的长度小于10个核苷酸。在一些实施例中,间隔区延伸序列的长度在10
‑
30个核苷酸之间。在一些实施例中,间隔区延伸序列的长度在30
‑
70个核苷酸之间。
[0069]
在一些实施例中,间隔区延伸序列具有另一个部分(例如,稳定性控制序列、内切核糖核酸酶结合序列、核酶)。在一些实施例中,该部分降低或增加靶向核酸的核酸的稳定性。在一些实施例中,该部分是转录终止子区段(即,转录终止序列)。在一些实施例中,该部分在真核细胞中起作用。在一些实施例中,该部分在原核细胞中起作用。在一些实施例中,
该部分在真核细胞和原核细胞两者中起作用。合适的部分的非限制性实例包括:5’帽(例如,7
‑
甲基鸟苷酸帽(m7g)),核糖开关序列(例如,允许蛋白和蛋白复合物调控稳定性和/或调控可及性),形成dsrna双链体的序列(即,发夹),将rna靶向亚细胞位置(例如,细胞核、线粒体、叶绿体等)的序列,提供跟踪的修饰或序列(例如,直接与荧光分子缀合,与促进荧光检测的部分缀合,允许进行荧光检测的序列等),和/或为蛋白(例如,作用于dna的蛋白,包括转录激活因子,转录阻遏因子、dna甲基转移酶、dna脱甲基化酶、组蛋白乙酰转移酶、组蛋白脱乙酰化酶等)提供结合位点的修饰或序列。
[0070]
间隔区序列
[0071]
间隔区序列可与目标靶核酸中的序列杂交。靶向基因组的核酸的间隔区经由杂交(即,碱基配对)以序列特异性方式与靶核酸相互作用。因此,间隔区的核苷酸序列根据目标靶核酸的序列而变化。
[0072]
在本文的crispr/cas系统中,间隔区序列设计成与位于系统中使用的cas9酶的pam的5’的靶核酸杂交。间隔区可以与靶序列完美匹配或者可以具有错配。每个cas9酶都有特定的pam序列,使得该酶识别靶dna。例如,酿脓链球菌(s.pyogenes)识别靶核酸中具有序列5
’‑
nrg
‑3’
的pam,其中r具有a或g,其中n是任何核苷酸并且n紧邻间隔区序列所靶向的靶核酸序列的3’。
[0073]
在一些实施例中,靶核酸序列具有20个核苷酸。在一些实施例中,靶核酸具有少于20个核苷酸。在一些实施例中,靶核酸具有超过20个核苷酸。在一些实施例中,靶核酸具有至少:5、10、15、16、17、18、19、20、21、22、23、24、25、30个或更多个核苷酸。在一些实施例中,靶核酸至多具有:5、10、15、16、17、18、19、20、21、22、23、24、25、30个或更多个核苷酸。在一些实施例中,靶核酸序列具有20个紧邻pam第一个核苷酸的5’的碱基。例如,在具有5
’‑
nnnnnnnnnnnnnnnnnnnnnrg
‑3’
的序列(seq id no:191)中,靶核酸具有对应于n的序列,其中n是任何核苷酸,并且加下划线的nrg序列(r是g或a)是酿脓链球菌cas9 pam。在一些实施例中,在本披露的组合物和方法中用作酿脓链球菌cas9所识别的pam序列是ngg。
[0074]
在一些实施例中,与靶核酸杂交的间隔区序列的长度为至少约6个核苷酸(nt)。间隔区序列可以为至少约6个nt、约10个nt、约15个nt、约18个nt、约19个nt、约20个nt、约25个nt、约30个nt、约35个nt或约40个nt、约6个nt至约80个nt、约6个nt至约50个nt、约6个nt至约45个nt、约6个nt至约40个nt、约6个nt至约35个nt、约6个nt至约30个nt、约6个nt至约25个nt、约6个nt至约20个nt、约6个nt至约19个nt、约10个nt至约50个nt、约10个nt至约45个nt、约10个nt至约40个nt、约10个nt至约35个nt、约10个nt至约30个nt、约10个nt至约25个nt、约10个nt至约20个nt、约10个nt至约19个nt、约19个nt至约25个nt、约19个nt至约30个nt、约19个nt至约35个nt、约19个nt至约40个nt、约19个nt至约45个nt、约19个nt至约50个nt、约19个nt至约60个nt、约20个nt至约25个nt、约20个nt至约30个nt、约20个nt至约35个nt、约20个nt至约40个nt、约20个nt至约45个nt、约20个nt至约50个nt、或约20个nt至约60个nt。在一些实施例中,间隔区序列具有20个核苷酸。在一些实施例中,间隔区具有19个核苷酸。在一些实施例中,间隔区具有18个核苷酸。在一些实施例中,间隔区具有17个核苷酸。在一些实施例中,间隔区具有16个核苷酸。在一些实施例中,间隔区具有15个核苷酸。
[0075]
在一些实施例中,间隔区序列与靶核酸之间的百分比互补性是至少约30%、至少约40%、至少约50%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少
约85%、至少约90%、至少约95%、至少约97%、至少约98%、至少约99%、或100%。在一些实施例中,间隔区序列与靶核酸之间的百分比互补性是至多约30%、至多约40%、至多约50%、至多约60%、至多约65%、至多约70%、至多约75%、至多约80%、至多约85%、至多约90%、至多约95%、至多约97%、至多约98%、至多约99%、或100%。在一些实施例中,间隔区序列与靶核酸之间的百分比互补性相比于靶核酸互补链的靶序列的六个连续最5’核苷酸为100%。在一些实施例中,间隔区序列与靶核酸之间的百分比互补性相比于约20个连续核苷酸为至少60%。在一些实施例中,间隔区序列和靶核酸的长度可以相差1至6个核苷酸,这可以被认为是一个或多个突起。
[0076]
在一些实施例中,使用计算机程序来设计或选择间隔区序列。计算机程序可以使用变量,诸如预测的解链温度、二级结构形成、预测的退火温度、序列同一性、基因组背景、染色质可及性、%gc、基因组发生(例如,相同或相似但由于错配、插入或缺失而在一个或多个点有所不同的序列)的频率、甲基化状态、snp的存在等。
[0077]
最小crispr重复序列
[0078]
在一些实施例中,最小crispr重复序列是与参考crispr重复序列(例如,来自酿脓链球菌的crrna;参见例如,j.j.ferretti等人,proc natl acad sci usa[美国国家科学院院刊](2001)98(8):4658
‑
63)具有至少约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%、或100%序列同一性的序列。
[0079]
在一些实施例中,最小crispr重复序列具有可与细胞中的最小tracrrna序列杂交的核苷酸。最小crispr重复序列和最小tracrrna序列形成双链体。最小crispr重复序列和最小tracrrna序列一起结合至定点多肽。最小crispr重复序列的至少一部分与最小tracrrna序列杂交。在一些实施例中,最小crispr重复序列的至少一部分与最小tracrrna序列具有至少约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%或100%的互补性。在一些实施例中,最小crispr重复序列的至少一部分与最小tracrrna序列具有至多约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%或100%的互补性。
[0080]
最小crispr重复序列可以具有约7个核苷酸至约100个核苷酸的长度。例如,最小crispr重复序列的长度是约7个nt至约50个nt、约7个nt至约40个nt、约7个nt至约30个nt、约7个nt至约25个nt、约7个nt至约20个nt、约7个nt至约15个nt、约8个nt至约40个nt、约8个nt至约30个nt、约8个nt至约25个nt、约8个nt至约20个nt、约8个nt至约15个nt、约15个nt至约100个nt、约15个nt至约80个nt、约15个nt至约50个nt、约15个nt至约40个nt、约15个nt至约30个nt、或约15个nt至约25个nt。在一些实施例中,最小crispr重复序列的长度是大约9个核苷酸。在一些实施例中,最小crispr重复序列的长度是大约12个核苷酸。
[0081]
在一些实施例中,最小crispr重复序列与参考最小crispr重复序列(例如,来自酿脓链球菌的野生型crrna;参见例如,j.j.ferretti等人,同上)在一段至少6、7、或8个连续核苷酸上具有至少约60%同一性。例如,最小crispr重复序列与参考最小crispr重复序列在一段至少6、7、或8个连续核苷酸上具有至少约65%同一性、至少约70%同一性、至少约75%同一性、至少约80%同一性、至少约85%同一性、至少约90%同一性、至少约95%同一性、至少约98%同一性、至少约99%同一性或100%同一性。
[0082]
最小tracrrna序列
[0083]
在一些实施例中,最小tracrrna序列是与参考tracrrna序列(例如,来自酿脓链球菌的野生型tracrrna;参见例如,j.j.ferretti等人,同上)具有至少约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%、或100%序列同一性的序列。
[0084]
在一些实施例中,最小tracrrna序列具有与细胞中的最小crispr重复序列杂交的核苷酸。最小tracrrna序列和最小crispr重复序列形成双链体。最小tracrrna序列和最小crispr重复序列一起结合至定点多肽。最小tracrrna序列的至少一部分可以与最小crispr重复序列杂交。在一些实施例中,最小tracrrna序列与最小crispr重复序列具有至少约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%、或100%的互补性。
[0085]
最小tracrrna序列可以具有约7个核苷酸至约100个核苷酸的长度。例如,最小tracrrna序列的长度可以是约7个nt至约50个nt、约7个nt至约40个nt、约7个nt至约30个nt、约7个nt至约25个nt、约7个nt至约20个nt、约7个nt至约15个nt、约8个nt至约40个nt、约8个nt至约30个nt、约8个nt至约25个nt、约8个nt至约20个nt、约8个nt至约15个nt、约15个nt至约100个nt、约15个nt至约80个nt、约15个nt至约50个nt、约15个nt至约40个nt、约15个nt至约30个nt或约15个nt至约25个nt。在一些实施例中,最小tracrrna序列的长度是大约9个核苷酸。在一些实施例中,最小tracrrna序列是大约12个核苷酸。在一些实施例中,最小tracrrna由m.jinek等人science[科学](2012)337(6096):816
‑
21中描述的tracrrna nt 23
‑
48组成。
[0086]
在一些实施例中,最小tracrrna序列与参考最小tracrrna序列(例如,来自酿脓链球菌的野生型tracrrna;参见例如,j.j.ferretti等人,同上)在一段至少6、7、或8个连续核苷酸上具有至少约60%同一性。例如,最小tracrrna序列与参考最小tracrrna序列在一段至少6、7、或8个连续核苷酸上具有至少约65%同一性、约70%同一性、约75%同一性、约80%同一性、约85%同一性、约90%同一性、约95%同一性、约98%同一性、约99%同一性或100%同一性。
[0087]
在一些实施例中,最小crispr rna和最小tracrrna之间的双链体具有双螺旋。在一些实施例中,最小crispr rna和最小tracrrna之间的双链体具有至少约1、2、3、4、5、6、7、8、9或10个或更多个核苷酸。在一些实施例中,最小crispr rna和最小tracrrna之间的双链体具有至多约1、2、3、4、5、6、7、8、9或10个或更多个核苷酸。
[0088]
在一些实施例中,双链体具有错配(即,双链体的两条链不是100%互补的)。在一些实施例中,双链体具有至少约1、2、3、4或5个错配。在一些实施例中,双链体具有至多约1、2、3、4或5个错配。在一些实施例中,双链体具有不超过2个错配。
[0089]
突起
[0090]
在一些实施例中,在最小crispr rna与最小tracrrna之间的双链体中存在“突起”。突起是双链体中核苷酸的未配对区域。在一些实施例中,突起有助于双链体与定点多肽的结合。突起在双链体的一侧具有未配对的5
’‑
xxxy
‑3’
,其中x是任何嘌呤,而y具有可以与相对链上的核苷酸形成摆动配对的核苷酸,并且突起在双链体的另一侧具有未配对的核苷酸区域。双链体两侧的未配对核苷酸数目可以不同。
[0091]
在一个实例中,突起在该突起的最小crispr重复链上具有未配对的嘌呤(例如,腺
嘌呤)。在一些实施例中,突起具有该突起的最小tracrrna序列链的未配对的5
’‑
aagy
‑3’
,其中y具有可以与最小crispr重复序列链上的核苷酸形成摆动配对的核苷酸。
[0092]
在一些实施例中,双链体的最小crispr重复侧上的突起具有至少1、2、3、4或5个或更多个未配对的核苷酸。在一些实施例中,双链体的最小crispr重复侧上的突起具有至多1、2、3、4或5个或更多个未配对的核苷酸。在一些实施例中,双链体的最小crispr重复序列侧上的突起具有1个未配对的核苷酸。
[0093]
在一些实施例中,双链体的最小tracrrna序列侧的突起具有至少1、2、3、4、5、6、7、8、9或10个或更多个未配对的核苷酸。在一些实施例中,双链体的最小tracrrna序列侧的突起具有至多1、2、3、4、5、6、7、8、9或10个或更多个未配对的核苷酸。在一些实施例中,双链体的第二侧(例如,双链体的最小tracrrna序列侧)上的突起具有4个未配对的核苷酸。
[0094]
在一些实施例中,突起具有至少一个摆动配对。在一些实施例中,突起具有至多一个摆动配对。在一些实施例中,突起具有至少一个嘌呤核苷酸。在一些实施例中,突起具有至少3个嘌呤核苷酸。在一些实施例中,突起序列具有至少5个嘌呤核苷酸。在一些实施例中,突起序列具有至少一个鸟嘌呤核苷酸。在一些实施例中,突起序列具有至少一个腺嘌呤核苷酸。
[0095]
发夹
[0096]
在一些实施例中,一个或多个发夹位于3’tracrrna序列中的最小tracrrna的3’。
[0097]
在一些实施例中,发夹起始于距最小crispr重复序列和最小tracrrna序列双链体中最后一个配对核苷酸的3’至少约1、2、3、4、5、6、7、8、9、10、15、或20个或更多个核苷酸处。在一些实施例中,发夹可以起始于距最小crispr重复序列和最小tracrrna序列双链体中最后一个配对核苷酸的3’至多约1、2、3、4、5、6、7、8、9或10个或更多个核苷酸处。
[0098]
在一些实施例中,发夹具有至少约1、2、3、4、5、6、7、8、9、10、15或20个或更多个连续的核苷酸。在一些实施例中,发夹具有至多约1、2、3、4、5、6、7、8、9、10、15个或更多个连续的核苷酸。
[0099]
在一些实施例中,发夹具有cc二核苷酸(即,两个连续的胞嘧啶核苷酸)。
[0100]
在一些实施例中,发夹具有双链体核苷酸(即,发夹中杂交在一起的核苷酸)。例如,发夹具有在3’tracrrna序列的发夹双链体中与gg二核苷酸杂交的cc二核苷酸。
[0101]
一个或多个发夹可以与定点多肽的向导rna相互作用区域相互作用。在一些实施例中,存在两个或更多个发夹,并且在一些实施例中,存在三个或更多个发夹。
[0102]3’
tracrrna序列
[0103]
在一些实施例中,3’tracrrna序列具有与参考tracrrna序列(例如,来自酿脓链球菌的tracrrna)具有至少约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%、或100%序列同一性的序列。
[0104]
在一些实施例中,3’tracrrna序列的长度为约6个核苷酸至约100个核苷酸。例如,3’tracrrna序列的长度可以是约6个nt至约50个nt、约6个nt至约40个nt、约6个nt至约30个nt、约6个nt至约25个nt、约6个nt至约20个nt、约6个nt至约15个nt、约8个nt至约40个nt、约8个nt至约30个nt、约8个nt至约25个nt、约8个nt至约20个nt、约8个nt至约15个nt、约15个nt至约100个nt、约15个nt至约80个nt、约15个nt至约50个nt、约15个nt至约40个nt、约15个nt至约30个nt或约15个nt至约25个nt。在一些实施例中,3’tracrrna序列的长度为大约14
个核苷酸。
[0105]
在一些实施例中,3’tracrrna序列与参考3’tracrrna序列在一段至少6、7、或8个连续核苷酸上具有至少约60%同一性。例如,3’tracrrna序列与参考3’tracrrna序列在一段至少6、7、或8个连续核苷酸上具有至少约60%同一性、至少约65%同一性、约70%同一性、约75%同一性、约80%同一性、约85%同一性、约90%同一性、约95%同一性、约98%同一性、约99%同一性或100%同一性。
[0106]
在一些实施例中,3’tracrrna序列具有多于一个双链体区域。在一些实施例中,3’tracrrna序列具有两个双链体区域。
[0107]
在一些实施例中,3’tracrrna序列具有茎环结构。在一些实施例中,3’tracrrna中的茎环结构具有至少1、2、3、4、5、6、7、8、9、10、15或20个或更多个核苷酸。在一些实施例中,3’tracrrna中的茎环结构具有至多1、2、3、4、5、6、7、8、9或10个或更多个核苷酸。在一些实施例中,茎环结构具有功能部分。例如,茎环结构可以具有适体、核酶、蛋白相互作用发夹、crispr阵列、内含子或外显子。在一些实施例中,茎环结构具有至少约1、2、3、4或5个或更多个功能部分。在一些实施例中,茎环结构具有至多约1、2、3、4或5个或更多个功能部分。
[0108]
在一些实施例中,3’tracrrna序列中的发夹具有p结构域。在一些实施例中,在发夹中p结构域具有双链区。
[0109]
tracrrna延伸序列
[0110]
在一些实施例中,无论tracrrna是在单分子向导还是双分子向导的背景下,都可以提供tracrrna延伸序列。在一些实施例中,tracrrna延伸序列的长度为约1个核苷酸至约400个核苷酸。在一些实施例中,tracrrna延伸序列的长度大于1、5、10、15、20、25、30、35、40、45、50、60、70、80、90、100、120、140、160、180、200、220、240、260、280、300、320、340、360、380、或400个核苷酸。在一些实施例中,tracrrna延伸序列的长度为约20至约5000个或更多个核苷酸。在一些实施例中,tracrrna延伸序列的长度大于1000个核苷酸。在一些实施例中,tracrrna延伸序列的长度小于1、5、10、15、20、25、30、35、40、45、50、60、70、80、90、100、120、140、160、180、200、220、240、260、280、300、320、340、360、380、400个或更多个核苷酸。在一些实施例中,tracrrna延伸序列的长度可以小于1000个核苷酸。在一些实施例中,tracrrna延伸序列的长度小于10个核苷酸。在一些实施例中,tracrrna延伸序列的长度为10
‑
30个核苷酸。在一些实施例中,tracrrna延伸序列的长度为30
‑
70个核苷酸。
[0111]
在一些实施例中,tracrrna延伸序列具有功能部分(例如,稳定性控制序列、核酶、内切核糖核酸酶结合序列)。在一些实施例中,功能部分具有转录终止子片段。在一些实施例中,功能部分的总长度为约10个nt至约100个核苷酸、约10个nt至约20个nt、约20个nt至约30个nt、约30个nt至约40个nt、约40个nt至约50个nt、约50个nt至约60个nt、约60个nt至约70个nt、约70个nt至约80个nt、约80个nt至约90个nt、或约90个nt至约100个nt、约15个nt至约80个nt、约15个nt至约50个nt、约15个nt至约40个nt、约15个nt至约30个nt、或约15个nt至约25个nt。在一些实施例中,功能部分在真核细胞中起作用。在一些实施例中,功能部分在原核细胞中起作用。在一些实施例中,功能部分在真核细胞和原核细胞两者中起作用。
[0112]
合适的tracrrna延伸功能部分的非限制性实例包括:3’聚腺苷酸化尾,核糖开关序列(例如,允许蛋白和蛋白复合物调控稳定性和/或调控可及性),形成dsrna双链体的序列,将rna靶向亚细胞位置(例如,细胞核、线粒体、叶绿体等)的序列,提供跟踪的修饰或序
no:5、6、9、8、11、15、16、12、7和10中任一个的间隔区序列或与seq id no:5、6、9、8、11、15、16、12、7和10中任一个相比具有不超过三个错配的其变体。在一些实施例中,该grna包含来自seq id no:17、18、29、76、50、54、81、96、64和51中任一个的间隔区序列或与seq id no:17、18、29、76、50、54、81、96、64和51中任一个相比具有不超过三个错配的其变体。在一些实施例中,间隔区序列长度为19个核苷酸并且不包括从中选择它的序列的位置1处的核苷酸。
[0119]
在一些实施例中,根据本披露由grna靶向的基因组位置可以处于基因组例如人类基因组中的内源纤维蛋白原α链(纤维蛋白原
‑
α)基因座处、内部或附近。靶向此类位置的示例性向导rna包括seq id no:192
‑
270中任一个列出的间隔区序列和相关的cas9或cpf1切割位点。如本领域普通技术人员所理解的,每个向导rna设计为包括与其基因组靶序列互补的间隔区序列。例如,可以将seq id no:192
‑
270中任一个列出的每个间隔区序列放入单个rna嵌合体或crrna(以及相应的tracrrna)中。参见m.jinek等人,同上,以及e.deltcheva等人,nature[自然](2011)471:602
‑
07。
[0120]
靶向白蛋白位置的示例性向导rna包括来自seq id no:271
‑
298中任一个的间隔区序列和相关的cas9或cpf1切割位点。例如,包括来自seq id no:271的间隔区序列的grna可以包括间隔区序列uaauuuucuuuugcgcacua(seq id no:299)。如本领域普通技术人员所理解的,每个向导rna设计为包括与其基因组靶序列互补的间隔区序列。例如,可以将来自seq id no:271
‑
298中任一个的每个间隔区序列放入单个rna嵌合体或crrna(以及相应的tracrrna)中。
[0121]
供体模板
[0122]
定点多肽(如,dna内切核酸酶)可以在核酸(例如,基因组dna)中引入双链断裂或单链断裂。双链断裂可以刺激细胞内源dna修复途径(例如,同源依赖性修复(hdr)、非同源末端连接或替代性非同源末端连接(a
‑
nhej)或微同源介导的末端连接(mmej))。nhej可以修复裂解的靶核酸,而无需同源模板。有时这可以在靶核酸的裂解位点处产生小缺失或插入(插入缺失),并且可以导致基因表达的破坏或改变。当同源修复模板或供体可用时,可发生hdr,也称为同源重组(hr)。
[0123]
同源供体模板具有与靶核酸裂解位点侧翼的序列同源的序列。姐妹染色单体通常被细胞用作修复模板。然而,出于基因组编辑的目的,修复模板通常作为外源核酸提供,诸如质粒、双链体寡核苷酸、单链寡核苷酸、双链寡核苷酸、或病毒核酸。对于外源供体模板,通常在具有同源性的侧翼区域之间引入另外的核酸序列(诸如转基因)或修饰(诸如单碱基或多碱基改变或缺失),使得另外的或改变的核酸序列也并入靶基因座中。mmej导致与nhej相似的遗传结果,因为在裂解位点可以发生小的缺失和插入。mmej利用位于裂解位点侧翼的几个碱基对的同源序列来驱动有利的末端连接dna修复结果。在一些情况下,可以基于核酸酶靶区域中潜在的微同源性分析来预测可能的修复结果。
[0124]
因此,在一些情况下,使用同源重组将外源多核苷酸序列插入靶核酸裂解位点。外源多核苷酸序列在本文中被称为供体模板(或供体或供体序列或供体dna模板)。在一些实施例中,将供体模板、供体模板的一部分、供体模板的拷贝、或供体模板的拷贝的一部分插入靶核酸裂解位点。在一些实施例中,供体模板是不天然存在于靶核酸裂解位点的序列。
[0125]
当在发生双链断裂的细胞核内以足够浓度提供外源dna分子时,可以在nhej修复过程中在双链断裂处插入外源dna,从而成为基因组的永久添加。如果供体模板含有目标基
因(诸如fviii基因)的编码序列,任选地还含有相关的调控序列(诸如启动子、增强子、聚a序列和/或剪接受体序列)(在本文中也称为“供体盒”),则编码序列可以从基因组中的整合拷贝表达,从而在细胞生命中永久表达。而且,当细胞分裂时,供体模板的整合拷贝可以传递到子细胞。
[0126]
在存在足够浓度的供体dna模板的情况下,该供体模板含有与双链断裂任一侧的dna序列具有同源性的侧翼dna序列(称为同源臂),可以经由hdr途径整合供体模板。同源臂充当供体模板与双链断裂任一侧的序列之间同源重组的底物。这可以导致供体模板的无错误插入,其中双链断裂任一侧的序列与未修饰的基因组中的序列相比没有改变。
[0127]
用于通过hdr进行编辑而提供的供体差异很大,但通常含有带有小或大侧翼同源臂的预期序列,以允许对基因组dna进行退火。引入的遗传变化侧翼的同源区可以是30bp或更小,或者与可以含有启动子、cdna等的几千碱基的盒一样大。可以使用单链和双链寡核苷酸供体。这些寡核苷酸的大小范围从小于100个nt到超过许多kb,但是也可以生成和使用更长的ssdna。通常使用双链供体,包括pcr扩增子、质粒和微环。一般而言,已经发现aav载体是递送供体模板非常有效的手段,但是单个供体的包装极限<5kb。供体的活跃转录使hdr增加三倍,表明包含启动子可以提高转化率。相反,供体的cpg甲基化可降低基因表达和hdr。
[0128]
在一些实施例中,可以用核酸酶或独立地通过多种不同的方法,例如通过转染、纳米颗粒、显微注射或病毒转导来提供供体dna。在一些实施例中,可以使用一系列的拴系选项来增加供体对于hdr的可用性。实例包括将供体附接至核酸酶、附接至附近结合的dna结合蛋白、或附接至参与dna末端结合或修复的蛋白。
[0129]
除了通过nhej或hdr进行基因组编辑外,还可以使用nhej途径和hr进行位点特异性基因插入。组合方法可适用于某些情形,可能包括内含子/外显子边界。nhej可以对内含子中的连接有效,而无错误hdr则更适合编码区。
[0130]
在实施例中,待插入基因组的外源序列是合成fviii编码序列,其编码在野生型b结构域将会是另外的形式的位置具有b结构域替代物的合成fviii蛋白。合成fviii编码序列可以包括编码合成fviii蛋白的核酸序列,该合成fviii蛋白具有野生型fviii蛋白的实质性活性,例如促凝血活性。合成fviii蛋白的活性程度可以是野生型fviii蛋白所表现出的活性的至少约30%、约40%、约50%、约60%、约70%、约80%、约85%、约90%、约95%或约100%。在一些实施例中,合成fviii蛋白可与fviii蛋白,例如野生型fviii蛋白具有至少约30%、约40%、约50%、约60%、约70%、约80%、约85%、约90%、约95%、约96%、约97%、约98%或约99%的氨基酸序列同一性。在一些实施例中,合成fviii蛋白可与不包括b结构域的fviii蛋白,例如b结构域裂解后的野生型fviii蛋白具有至少约30%、约40%、约50%、约60%、约70%、约80%、约85%、约90%、约95%、约96%、约97%、约98%或约99%的氨基酸序列同一性。在一些实施例中,本领域的普通技术人员可以使用本领域中已知的许多方法来测试化合物例如肽或蛋白的功能或活性。合成fviii蛋白还可以包括野生型fviii蛋白的任何片段或在全长野生型fviii蛋白的一个或多个氨基酸残基上具有保守性修饰的经修饰fviii蛋白的片段。因此,在一些实施例中,合成fviii编码序列可与fviii编码序列,例如野生型fviii编码序列具有至少约30%、约40%、约50%、约60%、约70%、约80%、约85%、约90%、约95%、约96%、约97%、约98%或约99%的核酸序列同一性。
[0131]
在本发明的实施例中,合成fviii含有一个或多个保守性或非保守性氨基酸取代,
这些取代改善蛋白的各方面而不会对蛋白的抗凝血性产生不利影响。在一个实施例中,位置309处的苯丙氨酸被丝氨酸或丙氨酸(非保守性地)替代以分别提供f309s和f309a突变蛋白。这些取代被认为会破坏a1结构域中伴侣免疫球蛋白结合蛋白(bip)的潜在结合位点,从而提高蛋白的表达和分泌(m.swaroop等人,j biol chem[生物化学杂志](1997)272:24121
‑
24)。
[0132]
本发明的b结构域替代物用小得多的肽链替代野生型fviii的b结构域,同时仍然提供蛋白酶裂解位点和一个或多个用于n
‑
连接糖基化的位点。b结构域替代物可具有约10至约200个氨基酸。在一些实施例中,b结构域替代物具有约20至约100个氨基酸。在一些实施例中,b结构域替代物具有约1至约40个氨基酸、约1至约35个氨基酸、约1至约30个氨基酸、约1至约25个氨基酸、约1至约20个氨基酸、约1至约15个氨基酸、约1至约10个氨基酸或约1至约5个氨基酸。在一些实施例中,b结构域替代物具有约5至约40个氨基酸、约10至约40个氨基酸、约15至约40个氨基酸、约20至约40个氨基酸、约25至约40个氨基酸、约30至约40个氨基酸或约35至约40个氨基酸。在一些实施例中,b结构域替代物具有1个氨基酸、2个氨基酸、3个氨基酸、4个氨基酸、5个氨基酸、6个氨基酸、7个氨基酸、8个氨基酸、9个氨基酸、10个氨基酸、11个氨基酸、12个氨基酸、13个氨基酸、14个氨基酸、15个氨基酸、16个氨基酸、17个氨基酸、18个氨基酸、19个氨基酸、20个氨基酸、21个氨基酸、22个氨基酸、23个氨基酸、24个氨基酸、25个氨基酸、26个氨基酸、27个氨基酸、28个氨基酸、29个氨基酸、30个氨基酸、31个氨基酸、32个氨基酸、33个氨基酸、34个氨基酸、35个氨基酸、36个氨基酸、37个氨基酸、38个氨基酸、39个氨基酸或40个氨基酸。在一些实施例中,编码b结构域替代物的核酸经密码子优化。在一些实施例中,b结构域替代物包含蛋白酶裂解位点,例如rhqr。
[0133]
在涉及插入其合成fviii编码序列的一些实施例中,可以将合成fviii编码序列的cdna插入具有缺陷fviii基因或其调控序列的受试者基因组中。在这种情况下,供体dna或供体模板可以是具有编码合成fviii的序列的表达盒或载体构建体。在一些实施例中,可以使用在说明书的其他地方描述的含有编码合成fviii的序列的表达载体。
[0134]
在一些实施例中,根据本文所述的包含供体盒的任何供体模板,供体盒在一侧或两侧上侧接有grna靶位点。例如,这种供体模板可以包含供体盒,在供体盒的5’具有grna靶位点和/或在供体盒的3’具有grna靶位点。在一些实施例中,供体模板包含在供体盒的5’具有grna靶位点的供体盒。在一些实施例中,供体模板包含在供体盒的3’具有grna靶位点的供体盒。在一些实施例中,供体模板包含供体盒,在供体盒的5’具有grna靶位点且在供体盒的3’具有grna靶位点。在一些实施例中,供体模板包含供体盒,在供体盒的5’具有grna靶位点且在供体盒的3’具有grna靶位点,并且两个grna靶位点包含相同的序列。在一些实施例中,供体模板包含至少一个grna靶位点,并且供体模板中的该至少一个grna靶位点包含与供体模板的供体盒要整合到的靶基因座中的grna靶位点相同的序列。在一些实施例中,供体模板包含至少一个grna靶位点,并且供体模板中的该至少一个grna靶位点包含供体模板的供体盒要整合到的靶基因座中的grna靶位点的反向互补序列。在一些实施例中,供体模板包含供体盒,在供体盒的5’具有grna靶位点且在供体盒的3’具有grna靶位点,并且供体模板中的两个grna靶位点包含与供体模板的供体盒要整合到的靶基因座中的grna靶位点相同的序列。在一些实施例中,供体模板包含供体盒,在供体盒的5’具有grna靶位点且在供体盒的3’具有grna靶位点,并且供体模板中的两个grna靶位点包含供体模板的供体盒要整
合到的靶基因座中的grna靶位点的反向互补序列。
[0135]
编码定点多肽或dna内切核酸酶的核酸
[0136]
因此,在一些实施例中,基因组编辑的方法和组合物可以使用编码定点多肽诸如dna内切核酸酶的核酸(或寡核苷酸)。编码定点多肽的核酸序列可以是dna或rna。如果编码定点多肽的核酸序列是rna,则它可以与grna序列共价连接或作为单独的序列存在。在一些实施例中,直接使用定点多肽(诸如dna内切核酸酶),而不是编码它的核酸序列。
[0137]
载体
[0138]
另一方面,本披露提供了一种核酸,该核酸具有编码本披露的靶向基因组的核酸,本披露的定点多肽和/或执行本披露方法的实施例所必需的任何核酸或蛋白分子的核苷酸序列。在一些实施例中,这种核酸是载体(例如,重组表达载体)。
[0139]
所考虑的表达载体包括但不限于基于痘苗病毒、脊髓灰质炎病毒、腺病毒、腺相关病毒、sv40、单纯疱疹病毒、人类免疫缺陷病毒、逆转录病毒(例如,鼠白血病病毒、脾坏死病毒,以及衍生自逆转录病毒(如劳氏肉瘤病毒(rous sarcoma virus)、哈维肉瘤病毒(harvey sarcoma virus)、禽类白血病病毒、慢病毒、人类免疫缺陷病毒、骨髓增生肉瘤病毒和乳腺肿瘤病毒)的载体)的病毒载体和其他重组载体。所考虑的用于真核靶细胞的其他载体包括但不限于载体pxt1、psg5、psvk3、pbpv、pmsg和psvlsv40(法玛西亚公司(pharmacia))。所考虑的用于真核靶细胞的另外的载体包括但不限于载体pctx
‑
1、pctx
‑
2和pctx
‑
3。可以使用其他载体,只要它们与宿主细胞相容即可。
[0140]
在一些实施例中,载体具有一个或多个转录和/或翻译控制元件。取决于所利用的宿主/载体系统,可以在表达载体中使用许多合适的转录和翻译控制元件中的任一种,包括组成型和诱导型启动子、转录增强子元件、转录终止子等。在一些实施例中,载体是自失活载体,其使病毒序列或crispr机制的组分或其他元件失活。
[0141]
合适的真核启动子(即,在真核细胞中有功能的启动子)的非限制性实例包括来自以下的那些:巨细胞病毒(cmv)立即早期启动子、单纯疱疹病毒(hsv)胸苷激酶、早期和晚期sv40启动子、来自逆转录病毒的长末端重复序列(ltr)、人延伸因子
‑
1(ef1)启动子、具有与鸡β
‑
肌动蛋白启动子(cag)融合的巨细胞病毒(cmv)增强子的杂交构建体、鼠干细胞病毒启动子(mscv)、磷酸甘油酸激酶1基因座启动子(pgk)和小鼠金属硫蛋白
‑
i。
[0142]
为了表达小rna(包括grna),启动子诸如rna聚合酶iii启动子(包括例如u6和h1)可以是有利的。增强此类启动子的用途的描述和参数是本领域已知的,并且定期描述另外的信息和方法;参见例如,h.ma等人,mol ther nuc acids[分子疗法
‑
核酸]3,e161(2014)doi:10.1038/mtna.2014.12。
[0143]
表达载体还可以含有用于翻译起始的核糖体结合位点和转录终止子。表达载体还可以包括用于扩增表达的适当序列。表达载体还可以包括编码与定点多肽融合的非天然标签(例如,组氨酸标签、血凝素标签、绿色荧光蛋白等)的核苷酸序列,从而产生融合蛋白。
[0144]
在一些实施例中,启动子是诱导型启动子(例如,热激启动子、四环素调节的启动子、类固醇调节的启动子、金属调节的启动子、雌激素受体调节的启动子等)。在一些实施例中,启动子是组成型启动子(例如,cmv启动子、ubc启动子)。在一些实施例中,启动子是空间受限和/或时间受限的启动子(例如,组织特异性启动子、细胞类型特异性启动子等)。在一些实施例中,如果要在载体插入基因组中之后在基因组中存在的内源性启动子下表达基
因,则载体不具有要在宿主细胞中表达的至少一个基因的启动子。
[0145]
定点多肽或dna内切核酸酶
[0146]
由于nhej和/或hdr引起的对靶dna的修饰可以导致例如突变、缺失、改变、整合、基因矫正、基因替换、基因标记、转基因插入、核苷酸缺失、基因破坏、易位和/或基因突变。将非天然核酸整合到基因组dna中的过程是基因组编辑的实例。
[0147]
定点多肽是在基因组编辑中用于裂解dna的核酸酶。能以一种或多种多肽、或一种或多种编码该多肽的mrna的方式将定点多肽施用于细胞或受试者。
[0148]
在crispr/cas或crispr/cpf1系统的背景下,定点多肽可与grna结合,该grna又指定靶dna中多肽所指向的位点。在本文的crispr/cas或crispr/cpf1系统的实施例中,定点多肽是内切核酸酶,诸如dna内切核酸酶。
[0149]
在一些实施例中,定点多肽具有多个核酸裂解(即,核酸酶)结构域。两个或更多个核酸裂解结构域可以经由接头连接在一起。在一些实施例中,该接头是柔性接头。接头的长度可为1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、30、35、40个或更多个氨基酸。
[0150]
天然存在的野生型cas9酶具有两个核酸酶结构域,即hnh核酸酶结构域和ruvc结构域。本文中,“cas9”是指天然存在的和重组的cas9两者。本文所考虑的cas9酶具有hnh核酸酶结构域或hnh样核酸酶结构域,和/或ruvc核酸酶结构域或ruvc样核酸酶结构域。
[0151]
hnh结构域和hnh样结构域具有mcra样折叠。hnh和hnh样结构域具有两条反平行的β
‑
链和α
‑
螺旋,并且具有金属结合位点(例如,二价阳离子结合位点)。hnh结构域和hnh样结构域可以裂解靶核酸的一条链(例如,crrna靶向的链的互补链)。
[0152]
ruvc和ruvc样结构域具有rna酶h或rna酶h样折叠。ruvc/rna酶h结构域涉及一系列不同的基于核酸的功能,并且作用于rna和dna两者。rna酶h结构域具有5条被多个α螺旋包围的β链。ruvc/rna酶h结构域和ruvc/rna酶h样结构域具有金属结合位点(例如,二价阳离子结合位点),并且可以裂解靶核酸的一条链(例如,双链靶dna的非互补链)。
[0153]
在一些实施例中,定点多肽具有的氨基酸序列与野生型示例性定点多肽(例如,来自酿脓链球菌的cas9,us 2014/0068797序列id no.8或r.sapranauskas等人,nuc acids res[核酸研究](2011)39(21):9275
‑
82)和其他定点多肽)具有至少10%、至少15%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少99%或100%的氨基酸序列同一性。
[0154]
在一些实施例中,定点多肽具有的氨基酸序列与野生型示例性定点多肽(例如,来自酿脓链球菌的cas9)的核酸酶结构域具有至少10%、至少15%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少99%或100%的氨基酸序列同一性。
[0155]
在一些实施例中,定点多肽是与野生型定点多肽(例如,来自酿脓链球菌的cas9)在10个连续氨基酸上具有至少70%、75%、80%、85%、90%、95%、97%、99%或100%的同一性的dna内切核酸酶。在一些实施例中,定点多肽与野生型定点多肽在10个连续氨基酸上具有至多70%、75%、80%、85%、90%、95%、97%、99%或100%的同一性。在一些实施例中,定点多肽与野生型定点多肽在定点多肽hnh核酸酶结构域的10个连续氨基酸上具有至少70%、75%、80%、85%、90%、95%、97%、99%或100%的同一性。在一些实施例中,定点
多肽与野生型定点多肽在定点多肽hnh核酸酶结构域的10个连续氨基酸上具有至多70%、75%、80%、85%、90%、95%、97%、99%或100%的同一性。在一些实施例中,定点多肽与野生型定点多肽在定点多肽ruvc核酸酶结构域的10个连续氨基酸上具有至少70%、75%、80%、85%、90%、95%、97%、99%或100%的同一性。在一些实施例中,定点多肽与野生型定点多肽在定点多肽ruvc核酸酶结构域的10个连续氨基酸上具有至多70%、75%、80%、85%、90%、95%、97%、99%或100%的同一性。
[0156]
在一些实施例中,定点多肽具有野生型示例性定点多肽的修饰形式。野生型示例性定点多肽的修饰形式具有降低定点多肽的核酸裂解活性的突变。在一些实施例中,野生型示例性定点多肽的修饰形式具有小于90%、小于80%、小于70%、小于60%、小于50%、小于40%、小于30%、小于20%、小于10%、小于5%、或小于1%的野生型示例性定点多肽的核酸裂解活性。定点多肽的修饰形式可以不具有实质性核酸裂解活性。当定点多肽是不具有实质性核酸裂解活性的修饰形式时,在本文中将其称为“酶促失活的”。
[0157]
在一些实施例中,定点多肽的修饰形式具有突变,使其可以在靶核酸上诱导单链断裂(ssb)(例如,通过切割双链靶核酸的仅一条糖
‑
磷酸骨架)。在一些实施例中,突变在野生型定点多肽(例如,来自酿脓链球菌的cas9)的多个核酸裂解结构域的一个或多个中产生小于90%、小于80%、小于70%、小于60%、小于50%、小于40%、小于30%、小于20%、小于10%、小于5%、或小于1%的核酸裂解活性。在一些实施例中,突变导致多个核酸裂解结构域中的一个或多个保留裂解靶核酸的互补链的能力,但是降低了裂解靶核酸的非互补链的能力。在一些实施例中,突变导致多个核酸裂解结构域中的一个或多个保留裂解靶核酸的非互补链的能力,但是降低了裂解靶核酸的互补链的能力。例如,使野生型示例性酿脓链球菌cas9多肽中的残基(诸如asp10、his840、asn854和asn856)突变,以使多个核酸裂解结构域(例如,核酸酶结构域)中的一个或多个失活。在一些实施例中,待突变的残基对应于野生型示例性酿脓链球菌cas9多肽中的残基asp10、his840、asn854和asn856(例如,通过序列和/或结构比对确定的)。突变的非限制性实例包括d10a、h840a、n854a和n856a。本领域技术人员将认识到,除丙氨酸取代以外的突变是合适的。
[0158]
在一些实施例中,将d10a突变与h840a、n854a或n856a突变中的一种或多种组合以产生基本上缺乏dna裂解活性的定点多肽。在一些实施例中,将h840a突变与d10a、n854a或n856a突变中的一种或多种组合以产生基本上缺乏dna裂解活性的定点多肽。在一些实施例中,将n854a突变与h840a、d10a或n856a突变中的一种或多种组合以产生基本上缺乏dna裂解活性的定点多肽。在一些实施例中,将n856a突变与h840a、n854a或d10a突变中的一种或多种组合以产生基本上缺乏dna裂解活性的定点多肽。具有一个基本上失活的核酸酶结构域的定点多肽称为“切口酶”。
[0159]
在一些实施例中,rna引导的内切核酸酶(例如cas9)的变体,可以用于增加crispr介导的基因组编辑的特异性。野生型cas9通常由设计为与靶序列(诸如内源基因组基因座)中~20个核苷酸的指定序列杂交的单个向导rna进行引导。然而,在向导rna和靶基因座之间可以容忍若干错配,从而有效地将靶位点所需的同源性长度减少到例如低至13个同源性nt,从而导致在靶基因组中其他位置处crispr/cas9复合物进行结合和双链核酸裂解(也称为脱靶裂解)的可能性增加。因为cas9的切口酶变体各自仅切割一条链,所以为了产生双链断裂,一对切口酶必须紧密结合在靶核酸的相反链上,从而产生一对切口,这相当于双链断
裂。这就要求两个单独的grna(针对每个切口酶各一个)必须紧密结合在靶核酸的相反链上。这一要求实质上使双链断裂发生所需的最小同源长度加倍,从而降低了在基因组中其他位置发生双链裂解事件的可能性,而这两个grna位点(如果存在)不太可能彼此足够靠近形成双链断裂。如本领域所述,切口酶还可以用于相对于nhej促进hdr。通过使用有效介导所希望的变化的特定供体序列,hdr可用于将所选变化引入基因组中的靶位点。对用于基因编辑的crispr/cas系统的描述可以在例如国际专利申请公开号wo 2013/176772和j.d.sander等人,nature biotechnology[自然生物技术](2014)32:347
‑
55,以及其中引用的参考文献中找到。
[0160]
在一些实施例中,定点多肽(例如,变体、突变的、酶促失活的和/或条件性酶促失活的定点多肽)靶向核酸。在一些实施例中,定点多肽靶向dna。在一些实施例中,定点多肽靶向rna。
[0161]
在一些实施例中,定点多肽具有一个或多个非天然序列(例如,定点多肽是融合蛋白)。
[0162]
在一些实施例中,定点多肽具有与来自细菌(例如,酿脓链球菌)的cas9具有至少15%氨基酸同一性的氨基酸序列,核酸结合结构域和两个核酸裂解结构域(例如,hnh结构域和ruvc结构域)。
[0163]
在一些实施例中,定点多肽具有与来自细菌的cas9具有至少15%氨基酸同一性的氨基酸序列,和两个核酸裂解结构域(即,hnh结构域和ruvc结构域)。
[0164]
在一些实施例中,定点多肽具有与来自细菌的cas9具有至少15%氨基酸同一性的氨基酸序列,和两个核酸裂解结构域,其中一个或两个核酸裂解结构域与来自细菌的cas9具有至少50%的氨基酸同一性。
[0165]
在一些实施例中,定点多肽具有与来自细菌的cas9具有至少15%氨基酸同一性的氨基酸序列,两个核酸裂解结构域(例如,hnh结构域和ruvc结构域)和非天然序列(例如,核定位信号)或连接定点多肽与非天然序列的接头。
[0166]
在一些实施例中,定点多肽具有与来自细菌的cas9具有至少15%氨基酸同一性的氨基酸序列,和两个核酸裂解结构域(例如,hnh结构域和ruvc结构域),其中定点多肽在一个或两个核酸裂解结构域中具有突变,该突变使核酸酶结构域的裂解活性降低至少50%。
[0167]
在一些实施例中,定点多肽具有与来自细菌的cas9具有至少15%氨基酸同一性的氨基酸序列,和两个核酸裂解结构域(例如,hnh结构域和ruvc结构域),其中一个核酸酶结构域具有天冬氨酸10突变,和/或其中一个核酸酶结构域具有组氨酸840突变,并且其中该突变使核酸酶结构域的裂解活性降低至少50%。
[0168]
在一些实施例中,该一种或多种定点多肽,诸如dna内切核酸酶,包括两个在基因组中的特定基因座处共同实现一个双链断裂的切口酶,或四个在基因组中的特定基因座处共同实现两个双链断裂的切口酶。替代性地,一种定点多肽影响基因组中特定基因座处的一个双链断裂。
[0169]
在一些实施例中,编码定点多肽的多核苷酸可用于编辑基因组。在一些此类实施例中,根据本领域已知的方法对编码定点多肽的多核苷酸进行密码子优化,以在含有目标靶dna的细胞中表达。例如,如果预期的靶核酸在人类细胞中,则可以使用编码cas9的人类密码子优化的多核苷酸产生cas9多肽。
[0170]
以下提供了可以在本披露的实施例中使用的定点多肽的一些实例。
[0171]
crispr内切核酸酶系统
[0172]
在许多原核生物(例如,细菌和古细菌)的基因组中都可以找到crispr(成簇规律间隔短回文重复序列)基因组基因座。在原核生物中,crispr基因座编码以下产物,这些产物起到一种类型的免疫系统的作用,以帮助原核生物防御外来入侵者(诸如病毒和噬菌体)。crispr基因座功能有三个阶段:将新序列整合到crispr基因座中,表达crispr rna(crrna),以及使外来入侵者核酸沉默。已经鉴定出五种类型的crispr系统(例如,i型、ii型、iii型、u型和v型)。
[0173]
crispr基因座包括许多短重复序列,称为“重复序列”。当表达时,重复序列可以形成二级发夹结构(例如,发夹)和/或具有非结构化的单链序列。重复序列通常成簇出现,并且经常在物种之间相异。重复序列与称为“间隔区”的独特插入序列有规律地间隔,从而形成重复序列
‑
间隔区
‑
重复序列基因座结构。间隔区与已知的外来入侵者序列相同或与其具有高度同源性。间隔区
‑
重复序列单元编码crrna,其被加工成间隔区
‑
重复序列单元的成熟形式。crrna具有参与靶向靶核酸的“种子”或间隔区序列(在原核生物中天然存在的形式,间隔区序列靶向外来入侵者核酸)。间隔区序列位于crrna的5’或3’末端。
[0174]
crispr基因座也具有编码crispr相关(cas)基因的多核苷酸序列。cas基因编码参与原核生物中crrna功能的生物发生和干扰阶段的内切核酸酶。一些cas基因具有同源二级和/或三级结构。
[0175]
ii型crispr系统
[0176]
实际上,ii型crispr系统中的crrna生物发生需要反式激活crispr rna(tracrrna)。tracrrna被内源性rna酶iii修饰,然后与前crrna阵列中的crrna重复序列杂交。募集内源性rna酶iii以裂解前crrna。裂解的crrna进行外切核糖核酸酶修剪以产生成熟的crrna形式(例如,进行5’修剪)。tracrrna保持与crrna杂交,并且tracrrna和crrna与定点多肽(例如,cas9)缔合。crrna
‑
tracrrna
‑
cas9复合物的crrna将该复合物引导至可以与crrna杂交的靶核酸。crrna与靶核酸的杂交可激活cas9进行靶向的核酸裂解。ii型crispr系统中的靶核酸称为原间隔区邻近基序(pam)。实际上,pam对促进定点多肽(例如,cas9)与靶核酸的结合至关重要。ii型系统(也称为nmeni或cass4)进一步细分为ii
‑
a型(cass4)和ii
‑
b型(cass4a)。m.jinek等人(同上)报道称crispr/cas9系统可用于rna可编程基因组编辑,并且国际专利申请公开号wo 2013/176772提供了用于位点特异性基因编辑的crispr/cas内切核酸酶系统的实例和应用。
[0177]
v型crispr系统
[0178]
v型crispr系统与ii型系统具有若干重要差异。例如,cpf1是单个rna引导的内切核酸酶,与ii型系统不同,缺少tracrrna。事实上,cpf1相关的crispr阵列无需另外的反式激活tracrrna就可以加工成成熟的crrna。将v型crispr阵列加工成长度为42
‑
44个核苷酸的短成熟crrna,其中每个成熟crrna均以19个核苷酸的正向重复序列开始,然后是23
‑
25个核苷酸的间隔区序列。相反,ii型系统中的成熟crrna以20
‑
24个核苷酸的间隔区序列开始,然后是约22个核苷酸的正向重复序列。同样,cpf1利用富含t的原间隔区邻近基序,使得cpf1
‑
crrna复合物有效地裂解前面是短的富含t的pam的靶dna,这与ii型系统中富含g的pam在靶dna之后相反。因此,v型系统在远离pam的点处裂解,而ii型系统在与pam邻近的点
处裂解。另外,与ii型系统不同,cpf1经由交错的dna双链断裂(具有4或5个核苷酸的5’突出端)裂解dna。ii型系统经由平的双链断裂裂解。与ii型系统相似,cpf1含有预测的ruvc样内切核酸酶结构域,但缺乏第二hnh内切核酸酶结构域,这与ii型系统相反。
[0179]
cas基因/多肽和原间隔区邻近基序
[0180]
示例性crispr/cas多肽包括i.fonfara等人,nucleic acids res.[核酸研究](2014)42:2577
‑
90的图1中的cas9多肽。自从发现cas基因以来,crispr/cas基因命名系统已经进行了广泛的重写。上文fonfara的图5提供了来自不同物种的cas9多肽的pam序列。
[0181]
靶向基因组的核酸和定点多肽的复合物
[0182]
靶向基因组的核酸与定点多肽(例如,核酸引导的核酸酶,诸如cas9)相互作用,从而形成复合物。靶向基因组的核酸(例如grna)将定点多肽引导至靶核酸。
[0183]
如前所述,在一些实施例中,定点多肽和靶向基因组的核酸可各自单独施用于细胞或受试者。在一些实施例中,定点多肽可以与一种或多种向导rna、或者一种或多种crrna以及tracrrna预复合。然后可以将预复合材料施用至细胞或受试者。这种预复合材料称为核糖核蛋白颗粒(rnp)。
[0184]
用于基因组编辑的系统
[0185]
本文提供了用于基因组编辑,尤其是用于将合成fviii编码序列插入细胞基因组中的系统。这些系统可以用于本文所述的方法中,诸如用于编辑细胞的基因组和用于治疗受试者,例如患有血友病a的受试者。
[0186]
在一些实施例中,本文提供了一种系统,该系统包含(a)dna内切核酸酶或编码该dna内切核酸酶的核酸;(b)靶向细胞基因组中的白蛋白基因座的grna;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板。在一些实施例中,grna靶向白蛋白基因的内含子1。在一些实施例中,grna包含来自seq id no:271
‑
298中任一个的间隔区序列。
[0187]
在一些实施例中,本文提供了一种系统,该系统包含(a)脱氧核糖核苷酸(dna)内切核酸酶或编码该dna内切核酸酶的核酸;(b)包含来自seq id no:271
‑
298中任一个的间隔区序列的向导rna(grna);以及(c)包含编码合成fviii蛋白的核酸序列的供体模板。在一些实施例中,grna包含来自seq id no:274、275、281和283中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274的间隔区序列。在一些实施例中,grna包含来自seq id no:275的间隔区序列。在一些实施例中,grna包含来自seq id no:281的间隔区序列。在一些实施例中,grna包含来自seq id no:283的间隔区序列。
[0188]
在一些实施例中,根据本文所述的任何系统,该dna内切核酸酶选自由以下项组成的组:cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4或cpf1内切核酸酶或其功能等效物。在一些实施例中,dna内切核酸酶是cas9。在一些实施例中,cas9来自酿脓链球菌(spcas9)。在一些实施例中,cas9来自路邓葡萄球菌(slucas9)。
[0189]
在一些实施例中,根据本文所述的任何系统,对编码合成fviii蛋白的核酸序列进行密码子优化以在宿主细胞中表达。在一些实施例中,对编码合成fviii蛋白的核酸序列进行密码子优化以在人类细胞中表达。
[0190]
在一些实施例中,根据本文所述的任何系统,该系统包含编码dna内切核酸酶的核酸。在一些实施例中,对编码dna内切核酸酶的核酸进行密码子优化以在宿主细胞中表达。在一些实施例中,对编码dna内切核酸酶的核酸进行密码子优化以在人类细胞中表达。在一些实施例中,编码dna内切核酸酶的核酸是dna,诸如dna质粒。在一些实施例中,编码dna内切核酸酶的核酸是rna,诸如mrna。
[0191]
在一些实施例中,根据本文所述的任何系统,供体模板在aav载体中编码。在一些实施例中,供体模板包含供体盒,该供体盒包含合成fviii编码序列,并且该供体盒在一侧或两侧上侧接有grna靶位点。在一些实施例中,供体盒在两侧上侧接有grna靶位点。在一些实施例中,grna靶位点是系统中grna的靶位点。在一些实施例中,供体模板的grna靶位点是该系统中grna的细胞基因组grna靶位点的反向互补序列。
[0192]
在一些实施例中,根据本文所述的任何系统,dna内切核酸酶或编码该dna内切核酸酶的核酸配制在脂质体或脂质纳米颗粒中。在一些实施例中,脂质体或脂质纳米颗粒还包含grna。在一些实施例中,脂质体或脂质纳米颗粒是脂质纳米颗粒。在一些实施例中,该系统包含脂质纳米颗粒,该脂质纳米颗粒包含编码dna内切核酸酶的核酸和grna。在一些实施例中,编码dna内切核酸酶的核酸是编码dna内切核酸酶的mrna。
[0193]
在一些实施例中,根据本文所述的任何系统,dna内切核酸酶与grna复合,形成rnp复合物。
[0194]
基因组编辑的方法
[0195]
本文提供了基因组编辑,尤其是将其合成fviii蛋白插入细胞基因组中的方法。该方法可用于治疗受试者,例如患有血友病a的患者,并且在这种情况下,可以从受试者或单独的供体中分离细胞。然后,使用本文所述的材料和方法编辑该细胞的染色体dna。
[0196]
本文提供了将合成fviii编码序列敲入基因组的方法。一方面,本披露提供了将合成fviii编码序列,即编码合成fviii蛋白的核酸序列插入细胞基因组中。合成fviii蛋白可以包括具有野生型fviii蛋白的大量活性,例如野生型fviii蛋白所表现出的活性的至少约30%、约40%、约50%、约60%、约70%、约80%、约90%、约95%或约100%的肽。在一些实施例中,本领域的普通技术人员可以使用本领域中已知的许多方法来测试化合物例如肽或蛋白的功能或活性。在一些实施例中,合成fviii蛋白还可以包括野生型fviii蛋白的任何片段或在全长野生型fviii蛋白的一个或多个氨基酸残基上具有保守性修饰的经修饰fviii蛋白的片段。在一些实施例中,合成fviii蛋白还可以包括基本上不会对野生型fviii蛋白的功能产生负面影响的任何修饰,例如一个或多个氨基酸的缺失、插入和/或突变。因此,在一些实施例中,合成fviii编码序列的核酸序列可与fviii编码序列具有至少约30%、约40%、约50%、约60%、约70%、约80%、约85%、约90%、约95%、约96%、约97%、约98%或约99%的核酸序列同一性。
[0197]
在一些实施例中,将合成fviii编码序列插入细胞的基因组序列中。在一些实施例中,插入位点处于细胞基因组中的白蛋白基因座、转铁蛋白基因座或纤维蛋白原α基因座处或内部。在一些实施例中,插入位点是白蛋白基因座。插入方法使用一种或多种靶向白蛋白基因第一个内含子(或内含子1)的grna。在一些实施例中,供体dna是具有合成fviii编码序列的单链或双链dna。
[0198]
在一些实施例中,基因组编辑方法利用dna内切核酸酶诸如crispr/cas系统遗传
引入(敲入)合成fviii编码序列。在一些实施例中,dna内切核酸酶是cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4或cpf1内切核酸酶、其同源物、天然存在的分子的重组物、其密码子优化或修饰形式以及任何前述物质的组合。在一些实施例中,dna内切核酸酶是cas9。在一些实施例中,cas9来自酿脓链球菌(spcas9)。在一些实施例中,cas9来自路邓葡萄球菌(slucas9)。
[0199]
在一些实施例中,经受基因组编辑的细胞在基因组中具有一个或多个突变,突变导致内源性fviii基因的表达与没有此类突变的正常细胞中的表达相比降低。正常细胞可以是来源于(或分离自)没有fviii基因缺陷的不同受试者的健康细胞或对照细胞。在一些实施例中,经受基因组编辑的细胞可以来源于(或分离自)需要治疗fviii基因相关病状或病症,例如血友病a的受试者。因此,在一些实施例中,这种细胞中内源性fviii基因的表达与正常细胞中内源性fviii基因的表达相比降低约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%或约100%。
[0200]
在一些实施例中,基因组编辑方法进行功能性fviii编码序列(例如与提供的启动子可操作地连接的fviii编码序列)(在基因组的非编码区)的靶向插入,以便在体内稳定地生成fviii蛋白。在一些实施例中,fviii编码序列的靶向整合发生于在目标细胞类型例如肝细胞或窦内皮细胞中高度表达的白蛋白基因的内含子中。
[0201]
一方面,将合成fviii编码序列的核酸序列插入细胞的基因组中。在实施例中,要插入的合成fviii编码序列是经修饰的fviii编码序列。在一些实施例中,在经修饰的fviii编码序列中,野生型fviii编码序列的b结构域缺失并且被b结构域替代物替代。在一些实施例中,合成fviii由于其尺寸较小而优于全长野生型fviii(4371bp对比7053bp)。因此,在一些实施例中,缺乏fviii信号肽并且在其5’末端(fviii编码序列的n端)含有剪接受体序列的合成fviii编码序列特异性整合到哺乳动物(包括人)的肝细胞中的基因座的内含子1中。在一个实施例中,基因座是白蛋白基因座。在另一个实施例中,基因座是转铁蛋白基因座。在另一个实施例中,基因座是纤维蛋白原α基因座。
[0202]
合成fviii编码序列从转铁蛋白启动子转录可产生含有转铁蛋白外显子1、内含子1的一部分和整合的合成fviii编码序列的前mrna。当这种前mrna经过自然剪接过程以去除内含子时,剪接机制可以将转铁蛋白外显子1的3’侧的剪接供体连接到下一个可用的剪接受体,该剪接受体将成为插入的dna供体的合成fviii编码序列的5’末端的剪接受体。这可以产生含有与合成fviii的成熟编码序列融合的转铁蛋白外显子1的成熟mrna。
[0203]
这种合成fviii编码序列从白蛋白启动子转录可产生含有白蛋白外显子1、内含子1的一部分和整合的合成fviii编码序列的前mrna。当这种前mrna经过自然剪接过程以去除内含子时,剪接机制可以将白蛋白外显子1的3’侧的剪接供体连接到下一个可用的剪接受体,该剪接受体将成为插入的dna供体的合成fviii编码序列的5’末端的剪接受体。这可以产生含有与合成fviii的成熟编码序列融合的白蛋白外显子1的成熟mrna。白蛋白的外显子1编码信号肽加上两个附加氨基酸和在人类中通常编码白蛋白n端的蛋白序列dah的密码子的1/3。因此,在一些实施例中,在从细胞分泌期间白蛋白信号肽预期裂解之后,可以产生合成fviii蛋白,该蛋白具有三个附加氨基酸残基添加至n端,从而在fviii
‑
fviii蛋白的n端
产生氨基酸序列
‑
dahatrryy(seq id no:300)。由于这三个氨基酸中的第3个(带下划线)部分由外显子1的末端编码,而部分由合成fviii dna供体模板编码,因此可以将第3个附加氨基酸残基的同一性选择为leu、pro、his、gln或arg。在这些选择中,在一些实施例中使用leu,因为leu是分子最不复杂的,因此最不可能形成新的t细胞表位,从而在合成fviii蛋白的n端产生氨基酸序列
‑
dalatrryy。替代性地,可以将dna供体模板设计为缺失第3个残基,从而在合成fviii蛋白的n端产生氨基酸序列daltrryy。在一些情况下,在天然蛋白的序列中添加附加氨基酸可增加免疫原性风险。因此,在预测合成fviii n端的两个潜在选择的潜在免疫原性的计算机模拟分析证明一个残基缺失(daltrryy)具有较低的免疫原性评分的一些实施例中,这可以是至少一些实施例中的设计。
[0204]
在一些实施例中,可以使用其中密码子使用已经优化的编码合成fviii的dna序列,以改善在哺乳动物细胞中的表达(所谓的“密码子优化”)。在该领域中不同的计算机算法也可用于进行密码子优化,并且这些算法生成不同的dna序列(v.p.mauro等人,trends mol med[分子医学趋势](2014)20:604
‑
13)。可商购的密码子优化算法的实例是atum和基因技术股份公司(geneart)(赛默飞世尔科技的一部分)采用的算法。经证明密码子优化fviii编码序列在向小鼠进行基于基因的递送后显著改善fviii的表达(a.c.nathwani等人,blood[血液](2006)107(7):2653
‑
61.;n.j.ward等人,blood[血液](2011)117(3):798
‑
807;p.a.radcliffe等人,gene ther.[基因疗法](2008)15(4):289
‑
97)。密码子优化是用于改善目标编码序列的表达的既定方法,并且主要基于用较不频繁使用的密码子取代较频繁使用的密码子,而不改变所编码的氨基酸序列。由于密码子偏倚可影响蛋白表达的初始认识,已经发展了密码子优化的方法,并且算法是可商购获得的,包括dna合成公司(诸如基因技术股份公司和atum)提供的那些。这些可商购获得的算法作为dna合成服务的一部分对用户是免费可获得的,并且也设计成去除隐蔽剪接信号并且甚至去除编码序列上的g/c内容物。递送外源核酸至体内细胞可诱导至少部分由toll受体系统识别cg二核苷酸(也称为cpg序列)所驱动的先天免疫应答,且cg二核苷酸含量的降低被提出作为降低对这些核酸的先天免疫应答的方式,尤其是当质粒dna为递送载体时。另见p.colella等人,mol ther methods clin dev[分子疗法
‑
方法与临床发展](2018)8:87
‑
104。当为了在哺乳动物物种中表达而优化基因的天然存在(天然)的编码序列时,通常增加cg二核苷酸的数目,因为更频繁使用的密码子在该密码子的第3(摆动)位置含有更高频率的g和c核苷酸。因此,编码序列中g和c核苷酸的总含量的增加将产生更高的gc二核苷酸含量。
[0205]
在一些实施例中,通过不同算法进行密码子优化的合成fviii编码序列与天然fviii序列(存在于人类基因组中)之间的序列同源性或同一性的范围可以为约30%、约40%、约50%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%或100%。在一些实施例中,密码子优化的合成fviii编码序列与天然fviii序列具有约75%至约79%的序列同源性或同一性。在一些实施例中,密码子优化的合成fviii编码序列与天然fviii序列具有约70%、约71%、约72%、约73%、约74%、约75%、约76%、约77%、约78%、约79%或约80%的序列同源性或同一性。
[0206]
在一些实施例中,将供体模板或供体构建体制备成含有编码合成fviii的dna序列。在一些实施例中,将dna供体模板设计成含有密码子优化的人合成fviii编码序列。在一些实施例中,密码子优化以这样的方式进行,使得5’末端编码fviii信号肽的序列已缺失并
且被剪接受体序列替代,另外,将聚腺苷酸化信号添加到3’末端的fviii终止密码子之后(mab8a
‑
seq id no:301)。剪接受体序列可以选自来自已知基因的已知剪接受体序列,或者可以使用源自本领域已知的许多剪接受体序列的比对的共有剪接受体序列。在一些实施例中,使用来自高度表达的基因的剪接受体序列,因为认为此类序列会提供最佳剪接效率。在一些实施例中,共有剪接受体序列由具有共有序列t/cnc/tt/ca/gac/t(seq id no:302)的分支位点(branch site)组成,其后20bp以内是10至12个碱基的多嘧啶串(c或t),接着是ag>g/a,其中>是内含子/外显子边界的位置。在一个实施例中,使用合成剪接受体序列(ctgacctcttctcttcctcccacag
‑
seq id no:303)。在另一个实施例中,使用来自人(ttaacaatccttttttttcttcccttgcccag
‑
seq id no:304)或小鼠(taaatatgttgtgtggtttttctctccctgtttccacag
‑
seq id no:305)的白蛋白基因内含子1/外显子2边界的天然剪接受体序列。
[0207]
聚腺苷酸化序列为细胞提供信号以添加聚a尾,这对于细胞内mrna的稳定性至关重要。在dna供体模板将要包装到aav颗粒中的一些实施例下,本发明的实施例将包装的dna的大小保持在aav的包装极限以内,该极限可以小于约5kb,或者不超过约4.7kb。因此,在一些实施例中,使用尽可能短的聚a序列,例如约10
‑
mer、约20
‑
mer、约30
‑
mer、约40
‑
mer、约50
‑
mer或约60
‑
mer或前述任何中间数目的核苷酸。共有合成的聚a信号序列已在文献中有所描述(n.levitt等人,genes dev[基因和发育](1989)3(7):1019
‑
25),其具有序列aataaaagatctttattttcattagatctgtgtgttggttttttgtgtg(seq id no:306),并且通常用于许多表达载体中。
[0208]
在一些实施例中,可以将附加序列元件添加至dna供体模板以提高整合频率。一种这样的元件是同源臂。双链断裂左侧的序列(lha)附加到dna供体模板的5’末端(fviii编码序列的n端),而双链断裂右侧的序列(rha)附加到dna供体模板例如mab8b(seq id no:308)的3’末端(fviii编码序列的c端)。
[0209]
在一些实施例中提供的替代性dna供体模板设计具有与将用于裂解基因组位点的sgrna的识别序列互补的序列。mab8c(seq id no:309)代表这种类型的dna供体模板的实例。通过包括sgrna识别位点,dna供体模板将被已向其中递送了dna供体模板和sgrna/cas9的细胞核内的sgrna/cas9复合物裂解。将供体模板裂解成线性片段可以提高通过非同源末端连接机制或通过hdr机制在双链断裂处整合的频率。在递送包装在aav中的供体模板的情况下,这可能特别有益,因为已知递送到细胞核中之后,aav基因组会连环化形成更大的环状双链dna分子(h.nakai等人,j virol[病毒学杂志](2001)75:6969
‑
76)。因此,在一些情况下,尤其是通过nhej机制,环状连环体可能是双链断裂处整合效率较低的供体。以前有报道称,通过在质粒中包含锌指核酸酶切割位点,可以提高使用环状质粒dna供体模板进行靶向整合的效率(s.cristea等人,biotechnol.bioeng.[生物技术与生物工程](2013)110:871
‑
80)。最近,还使用crispr/cas9核酸酶应用了这种方法(k.suzuki等人,nature[自然](2017)540:144
‑
49)。虽然sgrna识别序列在存在于双链dna供体模板的任一条链上时有活性,但预计使用基因组中存在的sgrna识别序列的反向互补序列有利于稳定整合,因为以相反方向整合会重新产生可以重新切割的sgrna识别序列,从而释放出插入的供体模板。预测通过nhej以正向方向将这种供体模板整合在基因组中不会重新产生sgrna识别序列,使得整合的供体模板不可以从基因组中切除。可以测试和确定在有或无同源臂的供体中包括sgrna识别序列在fviii供体模板的整合效率上的益处,例如在小鼠中使用aav递送供体以
及使用lnp(脂质纳米颗粒)递送crispr/cas9组件。
[0210]
在一些实施例中,供体模板在根据本文所述的任何实施例的供体盒中包含合成fviii编码序列,该供体盒在一侧或两侧上侧接有grna靶位点。在一些实施例中,供体模板包含在供体盒的5’的grna靶位点和/或在供体盒的3’的grna靶位点。在一些实施例中,供体模板包含两个侧翼grna靶位点,并且这两个grna靶位点包含相同序列。在一些实施例中,供体模板包含至少一个grna靶位点,并且供体模板中的该至少一个grna靶位点是靶向白蛋白基因的第一内含子的一个或多个grna中的至少一个的靶位点。在一些实施例中,供体模板包含至少一个grna靶位点,并且供体模板中的该至少一个grna靶位点是白蛋白基因的第一内含子中一个或多个grna中的至少一个的靶位点的反向互补序列。在一些实施例中,供体模板包含在供体盒的5’的grna靶位点和在供体盒的3’的grna靶位点,并且供体模板中的这两个grna靶位点被靶向白蛋白基因的第一内含子的一个或多个grna靶向。在一些实施例中,供体模板包含在供体盒的5’的grna靶位点和在供体盒的3’的grna靶位点,并且供体模板中的这两个grna靶位点是白蛋白基因的第一内含子中一个或多个grna中的至少一个的靶位点的反向互补序列。
[0211]
将fviii编码序列插入靶位点,即插入fviii编码序列的基因组位置,可以是在内源白蛋白基因座或其邻近序列中。在一些实施例中,以这样的方式插入fviii编码序列,使得插入的编码序列的表达受白蛋白基因的内源启动子控制。在一些实施例中,将fviii编码序列插入白蛋白基因的一个内含子中。在一些实施例中,将fviii编码序列插入白蛋白基因的一个外显子中。在一些实施例中,将fviii编码序列插入内含子:外显子(或反之亦然)的连接处。在一些实施例中,fviii编码序列的插入是在白蛋白基因座的第一内含子(或内含子1)中。在一些实施例中,fviii编码序列的插入不会明显影响(例如上调或下调)白蛋白基因的表达。
[0212]
在实施例中,用于插入fviii编码序列的靶位点是在内源白蛋白基因处、内部或附近。在一些实施例中,靶位点在基因间区中,该基因间区在基因组中白蛋白基因座的启动子上游。在一些实施例中,靶位点在白蛋白基因座内。在一些实施例中,靶位点在白蛋白基因座的一个内含子中。在一些实施例中,靶位点在白蛋白基因座的一个外显子中。在一些实施例中,靶位点在白蛋白基因座的内含子和外显子之间(或反之亦然)的一个连接处。在一些实施例中,靶位点在白蛋白基因座的第一内含子(或内含子1)中。在某些实施例中,靶位点在白蛋白基因的第一外显子下游至少、约或至多0、1、5、10、20、30、40、50、100、150、200、250、300、350、400、450或500或550或600或650bp处。在一些实施例中,靶位点在白蛋白基因的第一内含子上游至少、约或至多0.1kb、约0.2kb、约0.3kb、约0.4kb、约0.5kb、约1kb、约1.5kb、约2kb、约2.5kb、约3kb、约3.5kb、约4kb、约4.5kb或约5kb处。在一些实施例中,靶位点在白蛋白基因的第二外显子上游约0bp至约100bp,上游约101bp至约200bp,上游约201bp至约300bp,上游约301bp至约400bp,约401bp至约500bp,上游约501bp至约600bp,上游约601bp至约700bp,上游约701bp至约800bp,上游约801bp至约900bp,上游约901bp至约1000bp,上游约1001bp至约1500bp,上游约1501bp至约2000bp,上游约2001bp至约2500bp,上游约2501bp至约3000bp,上游约3001bp至约3500bp,上游约3501bp至约4000bp,上游约4001bp至约4500bp或上游约4501bp至约5000bp以内的任何位置。在一些实施例中,靶位点在基因组中人白蛋白基因的第一外显子末端(即3’末端)下游至少37bp。在一些实施例中,
靶位点在基因组中人白蛋白基因的第二外显子起点(即5’起点)上游至少330bp。
[0213]
在一些实施例中,本文提供了一种编辑细胞中的基因组的方法,该方法包括向细胞提供以下物质:(a)靶向细胞基因组中的白蛋白基因座的向导rna(grna);(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板。在一些实施例中,grna靶向白蛋白基因的内含子1。在一些实施例中,grna包含来自seq id no:271
‑
298中任一个的间隔区序列。
[0214]
在一些实施例中,本文提供了一种编辑细胞中的基因组的方法,该方法包括向细胞提供以下物质:(a)包含来自seq id no:271
‑
298中任一个的间隔区序列的grna;(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板。在一些实施例中,grna包含来自seq id no:274、275、281和283中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274的间隔区序列。在一些实施例中,grna包含来自seq id no:275的间隔区序列。在一些实施例中,grna包含来自seq id no:281的间隔区序列。在一些实施例中,grna包含来自seq id no:283的间隔区序列。在一些实施例中,细胞是人细胞,例如人肝脏细胞。
[0215]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,该dna内切核酸酶选自由以下项组成的组:cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4或cpf1内切核酸酶或其功能等效物。在一些实施例中,dna内切核酸酶是cas9。在一些实施例中,cas9是spcas9。在一些实施例中,cas9是slucas9。
[0216]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,对编码合成fviii蛋白的核酸序列进行密码子优化以在细胞中表达。在一些实施例中,细胞是人细胞。
[0217]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,该方法采用编码dna内切核酸酶的核酸。在一些实施例中,对编码dna内切核酸酶的核酸进行密码子优化以在细胞中表达。在一些实施例中,细胞是人细胞,例如人肝脏细胞。在一些实施例中,编码dna内切核酸酶的核酸是dna,诸如dna质粒。在一些实施例中,编码dna内切核酸酶的核酸是rna,诸如mrna。
[0218]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,供体模板在aav载体中编码。在一些实施例中,供体模板包含供体盒,该供体盒包含编码合成fviii蛋白的核酸序列,并且该供体盒在一侧或两侧上侧接有grna靶位点。在一些实施例中,供体盒在两侧上侧接有grna靶位点。在一些实施例中,该grna靶位点是所施用的grna的靶位点。在一些实施例中,供体模板的grna靶位点是grna的细胞基因组grna靶位点的反向互补序列。
[0219]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,dna内切核酸酶或编码该dna内切核酸酶的核酸配制在脂质体或脂质纳米颗粒中。在一些实施例中,脂质体或脂质纳米颗粒还包含grna。在一些实施例中,脂质体或脂质纳米颗粒是脂质纳米颗粒。在一些实施例中,该方法采用脂质纳米颗粒,该脂质纳米颗粒包含编码dna内切核酸酶的核酸和grna。在一些实施例中,编码dna内切核酸酶的核酸是编码dna内切核酸酶的mrna。
[0220]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,dna内切核酸
酶与grna预复合,形成rnp复合物。
[0221]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,在将供体模板提供给细胞之后,将grna和dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,在将该供体模板提供给该细胞后超过4天,将grna和dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,在将该供体模板提供给该细胞后至少14天,将grna和dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,在将该供体模板提供给该细胞后至少17天,将grna和dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,将(a)和(b)作为包含编码dna内切核酸酶的核酸和grna的脂质纳米颗粒提供给细胞。在一些实施例中,编码dna内切核酸酶的核酸是编码dna内切核酸酶的mrna。在一些实施例中,将(c)作为编码供体模板的aav载体提供给细胞。
[0222]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,在第一剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸。在一些实施例中,在第一剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸,直至达到该编码合成fviii蛋白的核酸序列的目标靶向整合水平和/或该编码合成fviii蛋白的核酸序列的目标表达水平。
[0223]
在一些实施例中,根据本文所述的编辑细胞中的基因组的任何方法,编码合成fviii蛋白的核酸序列在内源白蛋白启动子的控制下表达。
[0224]
在一些实施例中,本文提供了将合成fviii编码序列插入细胞基因组的白蛋白基因座中的方法,该方法包括将以下物质引入细胞中:(a)cas dna内切核酸酶(例如,cas9)或编码cas dna内切核酸酶的核酸,(b)grna或编码grna的核酸,其中grna能够引导cas dna内切核酸酶裂解白蛋白基因座中的靶多核苷酸序列,以及(c)根据本文所述的任何实施例的包含合成fviii编码序列的供体模板。在一些实施例中,该方法包括将编码cas dna内切核酸酶的mrna引入细胞中。在一些实施例中,该方法包括将根据本文所述的任何实施例的lnp引入细胞中,该lnp包含i)编码cas dna内切核酸酶的mrna和ii)grna。在一些实施例中,供体模板是aav供体模板。在一些实施例中,供体模板包含含有合成fviii编码序列的供体盒,其中供体盒在一侧或两侧上侧接有grna靶位点。在一些实施例中,供体盒侧翼的grna靶位点是白蛋白基因座中grna靶位点的反向互补序列。在一些实施例中,在将供体模板引入细胞后,将cas dna内切核酸酶或编码cas dna内切核酸酶的核酸以及grna或编码grna的核酸引入细胞。在一些实施例中,在将供体模板引入细胞中允许供体模板进入细胞核的足够时间后,将cas dna内切核酸酶或编码cas dna内切核酸酶的核酸以及grna或编码grna的核酸引入细胞中。在一些实施例中,在将供体模板引入细胞中允许供体模板在细胞核中由单链aav基因组转化为双链dna分子的足够时间后,将cas dna内切核酸酶或编码cas dna内切核酸酶的核酸以及grna或编码grna的核酸引入细胞中。在一些实施例中,cas dna内切核酸酶是cas9。
[0225]
在一些实施例中,根据本文所述的将合成fviii编码序列插入细胞基因组的白蛋白基因座中的任何方法,靶多核苷酸序列在白蛋白基因的内含子1中。在一些实施例中,
grna包含来自seq id no:271
‑
298中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274、275、281和283中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274的间隔区序列。在一些实施例中,grna包含来自seq id no:275的间隔区序列。在一些实施例中,grna包含来自seq id no:281的间隔区序列。在一些实施例中,grna包含来自seq id no:283的间隔区序列。
[0226]
在一些实施例中,本文提供了将合成fviii编码序列插入细胞基因组的白蛋白基因座中的方法,该方法包括将以下物质引入细胞中:(a)根据本文所述的任何实施例的lnp,其包含i)编码cas9 dna内切核酸酶的mrna和ii)grna,其中grna能够引导cas9 dna内切核酸酶裂解白蛋白基因座中的靶多核苷酸序列,以及(b)根据本文所述的任何实施例的包含合成fviii编码序列的aav供体模板。在一些实施例中,供体模板包含含有合成fviii编码序列的供体盒,其中供体盒在一侧或两侧上侧接有grna靶位点。在一些实施例中,供体盒侧翼的grna靶位点是白蛋白基因座中grna靶位点的反向互补序列。在一些实施例中,在将aav供体模板引入细胞后,将lnp引入细胞。在一些实施例中,在将aav供体模板引入细胞中允许供体模板进入细胞核的足够时间后将lnp引入细胞中。在一些实施例中,在将aav供体模板引入细胞中允许供体模板在细胞核中由单链aav基因组转化为双链dna分子的足够时间后将lnp引入细胞中。在一些实施例中,在第一次将lnp引入细胞后,进行向细胞中一次或多次(诸如2、3、4、5次或更多次)附加引入lnp。在一些实施例中,grna包含来自seq id no:271
‑
298中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274、275、281和283中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274的间隔区序列。在一些实施例中,grna包含来自seq id no:275的间隔区序列。在一些实施例中,grna包含来自seq id no:281的间隔区序列。在一些实施例中,grna包含来自seq id no:283的间隔区序列。
[0227]
fviii编码序列插入靶位点可以在内源纤维蛋白原
‑
α基因座或其相邻序列中。在一些实施例中,以这样的方式插入fviii编码序列,使得插入的编码序列的表达受纤维蛋白原
‑
α基因的内源启动子控制。在一些实施例中,将fviii编码序列插入纤维蛋白原
‑
α基因的一个内含子中。在一些实施例中,将fviii编码序列插入纤维蛋白原
‑
α基因的一个外显子中。在一些实施例中,将fviii编码序列插入内含子:外显子(或反之亦然)的连接处。在一些实施例中,fviii编码序列的插入在纤维蛋白原
‑
α基因座的第一内含子(或内含子1)中。在一些实施例中,fviii编码序列的插入不会明显影响(例如上调或下调)纤维蛋白原
‑
α基因的表达。
[0228]
在某些实施例中,靶位点处于纤维蛋白原
‑
α基因的第一外显子下游(即,从第一外显子的最后一个碱基对或3’末端开始)至少、约或至多0、1、5、10、20、30、40、50、100、150、200、250、300、350、400、450、500、550、600、650、700、750、800、850、900、950、1000、1050、1071bp或任何介于中间的核酸长度。在一些实施例中,靶位点处于纤维蛋白原
‑
α基因的第二外显子上游(即,从第二外显子的第一个核酸或5’末端开始)至少、约或至多0.1kb、约0.2kb、约0.3kb、约0.4kb、约0.5kb、约1kb或任何介于中间的核酸长度。在一些实施例中,靶位点处于纤维蛋白原
‑
α基因的第二外显子上游(即,从第二外显子的第一个核酸或5’末端开始)约0bp至约100bp、约101bp至约200bp、约201bp至约300bp、约301bp至约400bp、约401bp至约500bp、约501bp至约600bp、约601bp至约700bp、约701bp至约800bp、约801bp至约
900bp、约901bp至约1000bp、约1001bp至约1071bp以内的任何地方。
[0229]
在一些实施例中,插入fviii编码序列的靶位点在基因组中人纤维蛋白原
‑
α基因的第一外显子的末端下游至少40bp,并且在基因组中人纤维蛋白原
‑
α基因的第二外显子的起点上游至少60bp。
[0230]
在一些实施例中,插入fviii编码序列的靶位点在基因组中人纤维蛋白原
‑
α基因的第一外显子的末端下游至少42bp,并且在基因组中人纤维蛋白原
‑
α基因的第二外显子的起点上游至少65bp。
[0231]
在一些实施例中,该插入在基因组中人纤维蛋白原
‑
α基因的第一外显子的末端下游至少12bp,并且在基因组中人纤维蛋白原
‑
α基因的第二外显子的起点上游至少52bp。
[0232]
在一些实施例中,该插入在基因组中人纤维蛋白原
‑
α基因的第一外显子的末端下游至少94bp,并且在基因组中人纤维蛋白原
‑
α基因的第二外显子的起点上游至少86bp。
[0233]
在一些实施例中,根据本文所述的任何系统,供体模板包含用于靶向整合到转铁蛋白基因的内含子1中的编码合成fviii的核酸序列,其中供体模板从5’至3’包含i)第一grna靶位点;ii)剪接受体;iii)编码合成fviii的核苷酸序列;和iv)聚腺苷酸化信号。在一些实施例中,供体模板进一步包含iv)聚腺苷酸化信号下游的第二grna靶位点。在一些实施例中,第一grna靶位点和第二grna靶位点是相同的。在一些实施例中,供体模板进一步包含在ii)剪接受体和iii)编码合成fviii蛋白的核苷酸序列之间的编码转铁蛋白基因的外显子2上编码的转铁蛋白信号肽的末端部分的序列或其变体,该变体保留内源序列的至少一些活性。在一些实施例中,供体模板进一步包含在i)第一grna靶位点与ii)剪接受体之间的多核苷酸间隔区。在一些实施例中,多核苷酸间隔区的长度为18个核苷酸。在一些实施例中,供体模板在一侧上侧接有第一aav itr(反向末端重复序列)和/或在另一侧上侧接有第二aav itr。在一些实施例中,第一aav itr是aav2 itr和/或第二aav itr是aav2 itr。在一些实施例中,iii)的核苷酸序列编码具有包含3、4、5或6个n
‑
连接的糖基化位点的b结构域替代物的合成fviii。供体模板组件的示例性序列可以在seq id no:310和/或311的供体模板序列中找到。
[0234]
靶序列选择
[0235]
在一些实施例中,5’边界和/或3’边界的位置相对于特定参考基因座的移动用于促进或增强基因编辑的特定应用,这部分取决于选择用于编辑的内切核酸酶系统,如本文进一步描述和说明的那样。
[0236]
在这种靶序列选择的第一个非限制性方面,许多内切核酸酶系统具有引导对潜在裂解靶位点的初始选择的规则或标准,诸如在crispr ii型或v型内切核酸酶的情况下在邻近dna裂解位点的特定位置需要有pam序列基序。
[0237]
在靶序列选择或优化的另一个非限制性方面,靶序列与dna内切核酸酶的特定组合的“脱靶”活性频率(即在所选靶序列以外的位点发生双链断裂的频率)相对于中靶活性的频率进行评估。在一些情况下,已在所需基因座进行正确编辑的细胞相对于其他细胞具有选择优势。选择优势的说明性但非限制性的实例包括获得诸如提高的复制率、持久性、对某些条件的抗性、引入受试者后体内成功植入率或持久性增强的属性,以及其他与此类细胞的维持或数目或活力增加相关的属性。在其他情况下,可以通过一种或多种用于鉴定、分类或以其他方式选择已正确编辑的细胞的筛选方法来正向选择已在所需基因座处正确编
辑的细胞。选择优势和定向选择方法都可以利用与矫正相关的表型。在一些实施例中,可以对细胞进行两次或更多次编辑以产生第二修饰,该第二修饰产生用于选择或纯化预期细胞群的新表型。可以通过添加选择或筛选标记的第二grna来产生这种第二修饰。在一些情况下,可以使用含有cdna还有选择标记的dna片段在所需基因座处对细胞进行正确编辑。
[0238]
在实施例中,无论在特定情况下是任何选择优势适用还是应用任何定向选择,还应通过考虑脱靶频率来引导靶序列选择,以增强应用的有效性和/或降低在除所需靶标以外的位点产生不需要的改变的可能性。如本文和本领域中进一步描述和说明的,脱靶活性的发生受多种因素的影响,这些因素包括靶位点与脱靶位点之间的相似性和相异性,以及所使用的特定内切核酸酶。可以使用协助预测脱靶活性的生物信息学工具,并且通常也可以使用此类工具来鉴定脱靶活性的最可能位点,然后可以在实验设置中对其进行评估以评价脱靶与中靶活性的相对频率,从而允许选择具有相对较高中靶活性的序列。本文提供了此类技术的实例,并且其他技术是本领域已知的。
[0239]
靶序列选择的另一方面涉及同源重组事件。共有同源区的序列可以用作导致间插序列缺失的同源重组事件的焦点。此类重组事件发生在染色体和其他dna序列的正常复制过程中,也发生在合成dna序列时,诸如在双链断裂(dsb)修复的情况下。dsb在正常细胞复制周期期间定期发生,但也可通过诸如紫外线和dna断裂的其他诱导物等因素,或诸如化学诱导物等试剂的存在而增强。许多此类诱导物导致dsb在基因组中不加选择地发生,并且dsb在正常细胞中被定期诱导和修复。在修复过程中,可以完全保真地重构原始序列,然而,在一些情况下,会在dsb位点引入小的插入缺失。
[0240]
如同在本文所述的内切核酸酶系统的情况下,还可以在特定位置特异性诱导dsb,本文所述的内切核酸酶系统可以用于在选定的染色体位置引起定向或优先的基因修饰事件。在dna修复(以及复制)的背景下同源序列易于重组的趋势可以在许多情况下加以利用,并且是基因编辑系统(诸如crispr)的一种应用的基础,其中使用同源定向修复将通过使用供体模板提供的目标序列插入所需的染色体位置。
[0241]
特定序列之间的同源区,可以是可以具有少至十个碱基对或更少的小的“微同源”区,也可以用于实现所需缺失。例如,将单个dsb引入与附近序列表现出微同源性的位点。在这种dsb的正常修复过程中,高频发生的结果是间插序列的缺失,这是由于dsb和伴随的细胞修复过程促进重组的结果。
[0242]
然而,在一些情形下,在同源区内选择靶序列也会引起更大的缺失,包括基因融合(当缺失处于编码区时),鉴于特定情形,这些缺失可能是或可能不是所需的。
[0243]
本文提供的实例进一步说明了用于产生设计用于插入fviii编码序列的dsb的靶区域的选择,以及此类区域内设计用于相对于中靶事件将脱靶事件减到最少的特定靶序列的选择。
[0244]
靶向整合
[0245]
在一些实施例中,本文提供的方法是在肝细胞基因组中的特定位置整合合成fviii编码序列,这称为“靶向整合”。在一些实施例中,通过使用序列特异性核酸酶在基因组dna中产生双链断裂来实现靶向整合。
[0246]
在一些实施例中使用的crispr/cas系统具有可以快速筛选大量基因组靶标以鉴定最佳crispr/cas设计的优点。靶向基因组任何区域的sgrna分子可通过定位与所有pam基
序相邻的20bp序列来在计算机上设计。pam基序在真核生物基因组中平均每15bp出现。然而,通过计算机模拟方法设计的sgrna将以不同效率在细胞中产生双链断裂,并且目前无法使用计算机模拟方法预测一系列sgrna分子的切割效率。由于sgrna可以在体外快速合成,因此这使得可以快速筛选给定基因组区域中所有可能的sgrna序列,以鉴定引起最有效切割的sgrna。通常,当在细胞中测试给定基因组区域内的一系列sgrna时,观察到裂解效率范围在0%至90%之间。也可以使用计算机模拟算法以及实验室实验来确定任何给定sgrna的脱靶可能性。虽然与sgrna的20bp识别序列的完美匹配主要在大多数真核基因组中仅出现一次,但在基因组中还存在许多与sgrna具有一个或多个碱基对错配的其他位点。这些位点可以按可变频率裂解,可变频率基于错配数目或位置往往是无法预测的。未通过计算机模拟分析鉴定的其他脱靶位点处的裂解也可发生。因此,筛选相关细胞类型中的许多sgrna以鉴定具有最有利的脱靶特性的sgrna是选择最佳sgrna用于治疗用途的关键组成部分。有利的脱靶特性不仅要考虑实际脱靶位点的数目和这些位点的切割频率,还要考虑这些位点在基因组中的位置。例如,与功能上重要的基因(尤其是致癌基因或抑癌基因)接近或在其内部的脱靶位点,被认为不如没有已知功能的基因间区中的位点有利。因此,无法简单地通过对生物体基因组序列的计算机模拟分析来预测对最佳sgrna的鉴定,而是需要实验测试。虽然计算机模拟分析可有助于缩小待测试的向导的数目,但无法预测具有高中靶切割的向导或预测具有低的所需脱靶切割的向导。实验数据表明,各自与基因组在目标区域(诸如白蛋白内含子1)中具有完美匹配的sgrna的切割效率从无切割到切割>90%变化,并且无法通过任何已知算法预测。给定sgrna促进cas酶裂解的能力可与基因组dna中该特定位点的可及性有关,可及性可由该区域中的染色质结构决定。静止分化细胞(诸如肝细胞)中的大多数基因组dna以高度浓缩的异染色质形式存在,而活跃转录的区域则以更开放的染色质状态存在,已知更开放的染色质是大分子更易接近的,诸如蛋白如cas蛋白。由于存在或不存在结合的转录因子或其他调节蛋白,即使在活跃转录的基因内,dna的某些特定区域也比其他区域更易接近。无法预测基因组中或特定基因组基因座或基因组基因座的区域(诸如内含子和诸如白蛋白内含子1)内的位点,因此需要在相关细胞类型中通过实验确定。一旦选择一些位点作为潜在插入位点,就可以向这种位点添加一些变异,例如在有或没有实验测试的情况下,通过将几个核苷酸移到选定位点的上游或下游来添加变异。
[0247]
在一些实施例中,可用于本文披露的方法中的grna是seq id no:271
‑
298中的一个或多个或其与seq id no:271
‑
298中的那些具有至少约85%的核苷酸序列同一性的任何功能等效物。
[0248]
核酸修饰
[0249]
在一些实施例中,引入细胞中的多核苷酸具有一种或多种修饰,这些修饰可单独或组合使用,例如以增强活性、稳定性或特异性,改变递送,减少宿主细胞中的先天免疫应答,或用于其他增强,如本文进一步描述和本领域已知的。
[0250]
在某些实施例中,在crispr/cas9/cpf1系统中使用经修饰的多核苷酸,在这种情况下,引入细胞中的向导rna(单分子向导或双分子向导)和/或编码cas或cpf1内切核酸酶的dna或rna可以是经修饰的,如下面所描述和说明的。此类经修饰的多核苷酸可以用于crispr/cas9/cpf1系统中以编辑任何一个或多个基因组基因座。
[0251]
使用crispr/cas9/cpf1系统进行此类用途的非限制性说明,可以使用对向导rna
的修饰来增强具有grna和cas或cpf1内切核酸酶的crispr/cas9/cpf1基因组编辑复合物的形成或稳定性,该grna可以是单分子向导或双分子。对grna的修饰也可以或替代性地用于增强基因组编辑复合物与基因组中靶序列之间相互作用的起始、稳定性或动力学,这可以用于例如增强中靶活性。对向导rna的修饰也可以或替代性地用于增强特异性,例如,与其他(脱靶)位点处的作用相比,中靶位点处的基因组编辑的相对速率。
[0252]
修饰也可以或替代性地用于增加指导rna的稳定性,例如通过增加其对细胞中存在的核糖核酸酶(rna酶)降解的抗性,从而导致其在细胞中的半衰期增加。增强向导rna半衰期的修饰可在其中将cas或cpf1内切核酸酶经由需要翻译以便产生内切核酸酶的rna引入待编辑的细胞的实施例中特别有用,因为增加与编码内切核酸酶的rna同时引入的向导rna的半衰期可用于增加向导rna和编码的cas或cpf1内切核酸酶在细胞中共存的时间。
[0253]
修饰也可以或替代性地用于降低引入细胞中的rna引发先天免疫应答的可能性或程度。如以下和本领域中所述,已经在rna干扰(rnai)(包括小干扰rna(sirna))的背景下充分表征的此类应答倾向于与rna的半衰期降低和/或细胞因子或与免疫应答有关的其他因子的引发相关。
[0254]
还可以对引入细胞中的编码内切核酸酶的rna进行一种或多种类型的修饰,这些修饰包括但不限于增强rna稳定性的修饰(诸如通过增加细胞中存在的rna酶的降解)、增强所得产物(即,内切核酸酶)的翻译的修饰、和/或降低引入细胞中的rna引发先天免疫应答的可能性或程度的修饰。
[0255]
类似地,可以使用诸如前述和其他的修饰的组合。在crispr/cas9/cpf1的情况下,例如,可以对向导rna进行一种或多种类型的修饰(包括上述示例的那些),和/或可以对编码cas内切核酸酶的rna进行一种或多种类型的修饰(包括以上示例的那些)。
[0256]
举例说明,crispr/cas9/cpf1系统中使用的向导rna或其他较小的rna可以通过化学手段容易地合成,这使许多修饰可以容易地并入,如下文所示和本领域所述。随着化学合成程序的不断发展,通过诸如高效液相色谱法(hplc,其避免使用诸如page等凝胶)等程序纯化此类rna随着多核苷酸长度显著增加超过约一百个核苷酸而趋于更具挑战性。用于产生更大长度的化学修饰的rna的一种方法是产生两个或更多个连接在一起的分子。更长的rna(诸如编码cas9内切核酸酶的那些)更容易酶促产生。虽然通常可用于酶促产生的rna中的修饰类型较少,但仍存在以下修饰,这些修饰可用于例如增强稳定性、降低先天免疫应答的可能性或程度、和/或增强其他属性,如以下和本领域进一步所描述的;并且定期开发新的修饰类型。
[0257]
通过举例说明修饰类型,尤其是那些经常与较小的化学合成的rna一起使用的修饰,修饰可以具有在糖的2’位置经修饰的一个或多个核苷酸,在一些实施例中为2
’‑
o
‑
烷基、2
’‑
o
‑
烷基
‑
o
‑
烷基或2
’‑
氟修饰的核苷酸。在一些实施例中,rna修饰包括在rna的3’末端的嘧啶、无碱基残基或反向碱基的核糖上的2
’‑
氟、2
’‑
氨基或2
’‑
o
‑
甲基修饰。已将此类修饰并入寡核苷酸中,并且已经报道这些寡核苷酸对给定靶标具有比2
’‑
脱氧寡核苷酸更高的t
m
(即,更高的靶结合亲和力)。
[0258]
已经报道许多核苷酸和核苷修饰使并入它们中的寡核苷酸比天然寡核苷酸对核酸酶消化更具抗性;这些经修饰的寡核苷酸比未经修饰的寡核苷酸完整地存活更长的时间。经修饰的寡核苷酸的具体实例包括具有经修饰的骨架的那些,例如硫代磷酸酯、磷酸三
酯、甲基膦酸酯、短链烷基或环烷基糖间连键或短链杂原子或杂环糖间连键。一些寡核苷酸是具有硫代磷酸酯骨架的寡核苷酸和具有杂原子骨架的寡核苷酸,尤其是ch2‑
nh
‑
o
‑
ch2、ch2‑
n(ch3)
‑
o
‑
ch2(称为亚甲基(甲基亚氨基)或mmi骨架)、ch2‑
o
‑
n(ch3)
‑
ch2、ch2‑
n(ch3)
‑
n(ch3)
‑
ch2和o
‑
n(ch3)
‑
ch2‑
ch2骨架,其中天然磷酸二酯骨架表示为o
‑
p
‑
o
‑
ch,);是具有酰胺骨架的寡核苷酸(参见a.de mesmaeker等人,ace chem res[化学研究报告](1995)28:366
‑
374);是具有吗啉代骨架结构的寡核苷酸(参见summerton和weller,美国专利号5,034,506);以及具有肽核酸(pna)骨架的寡核苷酸(下文描述)。含磷连键包括但不限于硫代磷酸酯、手性硫代磷酸酯、二硫代磷酸酯、磷酸三酯、氨基烷基磷酸三酯、具有3’亚烷基膦酸酯和手性膦酸酯的甲基和其他烷基膦酸酯、亚膦酸酯、具有3
’‑
氨基氨基磷酸酯和氨基烷基氨基磷酸酯的氨基磷酸酯、硫羰基氨基磷酸酯、硫羰基烷基膦酸酯、硫羰基烷基磷酸三酯和具有正常3
’‑5’
连键的硼烷磷酸酯,这些的2
’‑5’
连接类似物以及极性反转的那些类似物,其中核苷单元的相邻对3
’‑5’
连接至5
’‑3’
或2
’‑5’
连接至5
’‑2’
;参见美国专利号3,687,808、4,469,863、4,476,301、5,023,243、5,177,196、5,188,897、5,264,423、5,276,019、5,278,302、5,286,717、5,321,131、5,399,676、5,405,939、5,453,496、5,455,233、5,466,677、5,476,925、5,519,126、5,536,821、5,541,306、5,550,111、5,563,253、5,571,799、5,587,361和5,625,050。
[0259]
基于吗啉基的寡聚化合物描述于d.a.braasch等人,biochem[生物化学](2002)41(14):4503
‑
10;s.c.ekker等人,genesis[起源](2001)30(3):89
‑
93(和该期刊中的其他论文);j.heasman,dev biol[发育生物学](2002)243:209
‑
14;a.nasevicius等人,nat genet[自然遗传](2000)26:216
‑
20;g.lacerra等人,proc natl acad sci usa[美国国家科学院院刊](2000)97:9591
‑
96;以及美国专利号5,034,506。
[0260]
环己烯基核酸寡核苷酸模拟物在j.wang等人,j am chem.soc[美国化学学会杂志](2000)122:8595
‑
602中有描述。
[0261]
其中不包括磷原子的经修饰的寡核苷酸骨架具有由短链烷基或环烷基核苷间连键、混合杂原子和烷基或环烷基核苷间连键或者一个或多个短链杂原子核苷间连键或杂环核苷间连键形成的骨架。这些具有那些具有吗啉基连键(部分由核苷的糖部分形成)的骨架;硅氧烷骨架;硫化物、亚砜和砜骨架;甲酰基和硫代甲酰基骨架;亚甲基甲酰基和硫代甲酰基骨架;含烯烃的骨架;氨基磺酸酯骨架;亚甲基亚氨基和亚甲基肼基骨架;磺酸酯和磺胺骨架;酰胺骨架;以及具有混合的n、o、s和ch2组分的其他骨架;参见美国专利号5,034,506、5,166,315、5,185,444、5,214,134、5,216,141、5,235,033、5,264,562、5,264,564、5,405,938、5,434,257、5,466,677、5,470,967、5,489,677、5,541,307、5,561,225、5,596,086、5,602,240、5,610,289、5,602,240、5,608,046、5,610,289、5,618,704、5,623,070、5,663,312、5,633,360、5,677,437和5,677,439,每个专利通过援引并入本文。
[0262]
还可以包括一种或多种经取代的糖部分,例如,在2’位置的以下之一:oh、sh、sch3、f、ocn、och3och3、och3o(ch2)
n
ch3、o(ch2)
n
nh2或o(ch2)
n
ch3,其中n为1至约10;c1至c
10
低级烷基、烷氧基烷氧基、经取代的低级烷基、烷芳基或芳烷基;cl;br;cn;cf3;ocf3;o
‑
、s
‑
或n
‑
烷基;o
‑
、s
‑
或n
‑
烯基;soch3;so2ch3;ono2;no2;n3;nh2;杂环烷基;杂环烷芳基;氨基烷基氨基;聚烷基氨基;经取代的甲硅烷基;rna裂解基团;报告基团;嵌入剂;用于改善寡核苷酸的药代动力学特性的基团;或者用于改善寡核苷酸的药效学特性的基团和具有类似性质的
其他取代基。在一些实施例中,修饰包括2
’‑
甲氧基乙氧基(2
’‑
o
‑
ch2ch2och3,也称为2
’‑
o
‑
(2
‑
甲氧基乙基))(p.martin等人,helv chim acta[瑞士化学学报],(1995)78:486)。其他修饰包括2
’‑
甲氧基(2
’‑
o
‑
ch3)、2
’‑
丙氧基(2
’‑
och2ch2ch3)和2
’‑
氟(2
’‑
f)。还可以在寡核苷酸上的其他位置,特别是在3’端核苷酸上的糖的3’位置和5’端核苷酸的5’位置进行类似的修饰。寡核苷酸也可以具有糖模拟物,诸如代替环戊呋喃糖基的环丁基。
[0263]
在一些实施例中,核苷酸单元的糖和核苷间连键(即骨架)均被新型基团替代。维持碱基单元以与适当的核酸靶化合物杂交。一种这样的寡聚化合物,即已经报道具有优异杂交特性的寡核苷酸模拟物,被称作肽核酸(pna)。在pna化合物中,寡核苷酸的糖骨架被替换为含有酰胺的骨架,例如氨基乙基甘氨酸骨架。这些核碱基得以保持并且直接或间接地结合至该骨架的酰胺部分的氮杂氮原子。传授制备pna化合物的代表性美国专利有但不限于美国专利号5,539,082、5,714,331和5,719,262。pna化合物的其他传授内容可以在p.e.nielsen等人,science[科学],(1991)254:1497
‑
500中找到。
[0264]
在一些实施例中,向导rna还可以另外或替代性地包括核碱基(在本领域中常常简称为“碱基”)修饰或取代。如本文所用,“未修饰的”或“天然的”核碱基包括腺嘌呤(a)、鸟嘌呤(g)、胸腺嘧啶(t)、胞嘧啶(c)和尿嘧啶(u)。经修饰的核碱基包括仅很少或瞬时发现于天然核酸中的核碱基,例如次黄嘌呤、6
‑
甲基腺嘌呤、5
‑
甲基嘧啶,特别是5
‑
甲基胞嘧啶(也称为5
‑
甲基
‑2’
脱氧胞嘧啶并且常常在本领域中称为5
‑
me
‑
c)、5
‑
羟甲基胞嘧啶(hmc)、糖基hmc和龙胆二糖基hmc,以及合成的核碱基,例如2
‑
氨基腺嘌呤、2
‑
(甲基氨基)腺嘌呤、2
‑
(咪唑基烷基)腺嘌呤、2
‑
(氨基烷基氨基)腺嘌呤或其他杂取代的烷基腺嘌呤、2
‑
硫尿嘧啶、2
‑
硫胸腺嘧啶、5
‑
溴尿嘧啶、5
‑
羟甲基尿嘧啶、8
‑
氮杂鸟嘌呤、7
‑
脱氮鸟嘌呤、n6(6
‑
氨基己基)腺嘌呤和2,6
‑
二氨基嘌呤;g.gebeyehu等人,nucl acids res[核酸研究](1997)15:4513。也可以包括本领域已知的“通用”碱基,例如肌苷。已报道5
‑
me
‑
c取代使核酸双链体稳定性提高0.6℃
‑
1.2℃。(y.s.sanghvi等人,“antisense research and applications”[反义研究与应用],crc press[crc出版社],boca raton[波卡拉顿],1993,第276
‑
278页)是碱基取代的实施例。
[0265]
在一些实施例中,经修饰的核碱基包括其他合成的和天然的核碱基,诸如5
‑
甲基胞嘧啶(5
‑
me
‑
c),5
‑
羟甲基胞嘧啶,黄嘌呤,次黄嘌呤,2
‑
氨基腺嘌呤,腺嘌呤和鸟嘌呤的6
‑
甲基和其他烷基衍生物,腺嘌呤和鸟嘌呤的2
‑
丙基和其他烷基衍生物,2
‑
硫尿嘧啶,2
‑
硫胸腺嘧啶和2
‑
硫胞嘧啶,5
‑
卤尿嘧啶和胞嘧啶,5
‑
丙炔尿嘧啶和胞嘧啶,6
‑
偶氮尿嘧啶、胞嘧啶和胸腺嘧啶,5
‑
尿嘧啶(假尿嘧啶),4
‑
硫尿嘧啶,8
‑
卤代、8
‑
氨基、8
‑
硫醇、8
‑
硫代烷基、8
‑
羟基和其他8
‑
取代的腺嘌呤和鸟嘌呤,5
‑
卤代尤其是5
‑
溴、5
‑
三氟甲基和其他的5
‑
取代的尿嘧啶和胞嘧啶,7
‑
甲基鸟嘌呤和7
‑
甲基腺嘌呤,8
‑
氮杂鸟嘌呤和8
‑
氮杂腺嘌呤,7
‑
脱氮鸟嘌呤和7
‑
脱氮杂腺嘌呤,以及3
‑
脱氮鸟嘌呤和3
‑
脱氮杂腺嘌呤。
[0266]
此外,核碱基包括美国专利号3,687,808中披露的那些,在“the concise encyclopedia of polymer science and engineering[高分子科学和工程简明百科全书],第858
‑
859页,kroschwitz,j.i.编辑,约翰威立国际出版公司(john wiley&sons),1990中披露的那些,englisch等人,ange.chemie[应用化学],国际版本(int’l ed),(1991)30:613中披露的那些,以及y.s.sanghvi,第15章,“antisense research and applications[反义研究和应用]”,第289
‑
302页,crooke,s.t.和lebleu,b.编辑,crc出版
社,1993披露的那些。这些核碱基中的某些对提高本披露的寡聚化合物的结合亲和力是特别有用的。这些包括具有2
‑
氨基丙基腺嘌呤、5
‑
丙炔基尿嘧啶和5
‑
丙炔基胞嘧啶的5
‑
取代的嘧啶、6
‑
氮杂嘧啶和n
‑
2、n
‑
6和0
‑
6取代的嘌呤。已报道5
‑
甲基胞嘧啶取代使核酸双链体稳定性提高0.6℃
‑
1.2℃(y.s.sanghvi,同上,第276
‑
78页)是碱基取代的实施例,甚至更特别地当与2
’‑
o
‑
甲氧基乙基糖修饰组合时。经修饰的核碱基在以下参考文献中有描述:美国专利号3,687,808、4,845,205、5,130,302、5,134,066、5,175,273、5,367,066、5,432,272、5,457,187、5,459,255、5,484,908、5,502,177、5,525,711、5,552,540、5,587,469、5,596,091、5,614,617、5,681,941、5,750,692、5,763,588、5,830,653、6,005,096和美国专利申请公开2003/0158403。
[0267]
在一些实施例中,将编码内切核酸酶的向导rna和/或mrna(或dna)化学连接至增强寡核苷酸的活性、细胞分布或细胞摄取的一个或多个部分或缀合物。此类部分包括但不限于脂质部分,诸如胆固醇部分(letsinger等人,proc natl acad sci usa[美国国家科学院院刊],(1989)86:6553
‑
56);胆酸(manoharan等人,bioorg med chem let[生物有机化学与医药化学通讯],(1994)4:1053
‑
60);硫醚,例如己基
‑
s
‑
三苯甲基硫醇(manoharan等人,ann n y acad sci[纽约科学会年鉴](1992)660:306
‑
09)和manoharan等人,bioorg med chem let[生物有机与药物化学快报],(1993)3:2765
‑
70);巯基胆固醇(oberhauser等人,nucl acids res[核酸研究](1992)20:533
‑
538);脂肪链,例如十二烷二醇或十一烷基残基(kabanov等人,febs lett.[欧洲生化学会联合会快报],(1990)259:327
‑
330和svinarchuk等人,biochimie[生物化学](1993)75:49
‑
54);磷脂,例如二
‑
十六烷基
‑
外消旋
‑
甘油或三乙铵1,2
‑
二
‑
o
‑
十六烷基
‑
外消旋
‑
甘油基
‑3‑
h
‑
膦酸酯(manoharan等人,tetrahedron lett[四面体通讯](1995)36:3651
‑
54和shea等人,nucl acids res[核酸研究](1990)18:3777
‑
83);聚胺或聚乙二醇链(mancharan等人,nucleosides&nucleotides[核苷与核苷酸],(1995)14:969
‑
73);金刚烷乙酸(manoharan等人,tetrahedron lett[四面体通讯],(1995)36:3651
‑
54);棕榈基部分(mishra等人,biochim biophys acta[生物化学与生物物理学学报],(1995)1264:229
‑
37);或十八烷基胺或己氨基
‑
羰基
‑
t
‑
羟胆固醇部分(crooke等人,j pharmacol exp ther[药理学与实验疗法杂志](1996)277:923
‑
37)。另见美国专利号4,828,979、4,948,882、5,218,105、5,525,465、5,541,313、5,545,730、5,552,538、5,578,717;5,580,731、5,580,731、5,591,584、5,109,124、5,118,802、5,138,045、5,414,077、5,486,603、5,512,439、5,578,718、5,608,046、4,587,044、4,605,735、4,667,025、4,762,779、4,789,737、4,824,941、4,835,263、4,876,335、4,904,582、4,958,013、5,082,830、5,112,963、5,214,136、5,082,830、5,112,963、5,214,136、5,245,022、5,254,469、5,258,506、5,262,536、5,272,250、5,292,873、5,317,098、5,371,241、5,391,723、5,416,203、5,451,463、5,510,475、5,512,667、5,514,785、5,565,552、5,567,810、5,574,142、5,585,481、5,587,371、5,595,726、5,597,696、5,599,923、5,599,928和5,688,941。
[0268]
在一些实施例中,糖和其他部分可用于将具有核苷酸的蛋白和复合物(诸如阳离子多核糖体和脂质体)靶向至特定位点。例如,肝细胞定向转移可以经由脱唾液酸糖蛋白受体(asgpr)介导;参见例如,hu等人,protein pept lett[蛋白和肽快报](2014)21(10):1025
‑
30。可以使用本领域已知的其他系统来将本情况中使用的生物分子和/或其复合物靶向至特定的目标靶细胞。
[0269]
在一些实施例中,这些靶向部分或缀合物可以包括共价结合至官能团诸如伯羟基或仲羟基的缀合物基团。本披露的缀合物基团包括嵌入剂、报告分子、聚胺、聚酰胺、聚乙二醇、聚醚、增强寡聚物的药效学特性的基团,和增强寡聚物的药代动力学特性的基团。示例性缀合物基团包括胆固醇、脂质、磷脂、生物素、吩嗪、叶酸、菲啶、蒽醌、吖啶、荧光素、罗丹明、香豆素和染料。在本披露的背景下,增强药效学特性的基团包括改善摄取、增强对降解的抗性、和/或加强与靶核酸的序列特异性杂交的基团。在本披露的背景下,增强药代动力学特性的基团包括改善本披露的化合物的摄取、分布、代谢或排泄的基团。代表性缀合物基团披露于1992年10月23日提交的国际专利申请号pct/us 92/09196和美国专利号6,287,860中,将其通过援引并入本文。缀合物部分包括但不限于脂质部分(如胆固醇部分)、胆酸、硫醚(例如己基
‑5‑
三苯甲基硫醚)、巯基胆固醇、脂肪链(例如十二烷二醇或十一基残基)、磷脂(例如二
‑
十六基
‑
外消旋
‑
甘油或三乙铵l,2
‑
二
‑
o
‑
十六基
‑
外消旋
‑
甘油
‑3‑
h
‑
膦酸酯)、多胺或聚乙二醇链、或金刚烷乙酸、棕榈基部分、或十八烷基胺、或己基氨基
‑
羰基
‑
氧基胆固醇部分。参见例如,美国专利号4,828,979、4,948,882、5,218,105、5,525,465、5,541,313、5,545,730、5,552,538、5,578,717;5,580,731、5,580,731、5,591,584、5,109,124、5,118,802、5,138,045、5,414,077、5,486,603、5,512,439、5,578,718、5,608,046、4,587,044、4,605,735、4,667,025、4,762,779、4,789,737、4,824,941、4,835,263、4,876,335、4,904,582、4,958,013、5,082,830、5,112,963、5,214,136、5,082,830、5,112,963、5,214,136、5,245,022、5,254,469、5,258,506、5,262,536、5,272,250、5,292,873、5,317,098、5,371,241、5,391,723、5,416,203、5,451,463、5,510,475、5,512,667、5,514,785、5,565,552、5,567,810、5,574,142、5,585,481、5,587,371、5,595,726、5,597,696、5,599,923、5,599,928和5,688,941。
[0270]
不太适合化学合成并且通常通过酶促合成产生的更长多核苷酸也可以通进行修饰。此类修饰可以包括例如某些核苷酸类似物的引入,在分子的5’或3’端并入特定的序列或其他部分,以及其他修饰。举例说明,编码cas9的mrna的长度大约为4kb,并且可以通过体外转录来合成。对mrna的修饰可用于例如增加其翻译或稳定性(如通过增加其对被细胞降解的抗性)、或降低rna引发通常在细胞中引入外源rna(特别是较长的rna,如编码cas9的rna)后观察到的先天免疫应答的趋势。
[0271]
在本领域中已经描述了许多这样的修饰,如polya尾、5’帽类似物(例如,抗反向帽类似物(arca)或m7g(5’)ppp(5’)g(mcap))、经修饰的5’或3’非翻译区(utr)、使用经修饰的碱基(如假utp、2
‑
硫
‑
utp、5
‑
甲基胞嘧啶
‑5’‑
三磷酸酯(5
‑
甲基
‑
ctp)或n6
‑
甲基
‑
atp)、或用磷酸酶处理以去除5’末端磷酸酯。这些修饰和其他修饰是本领域已知的,并且定期开发rna的新修饰。
[0272]
已经报道,体内递送的经化学修饰的mrna可用于实现改善的治疗效果;参见例如,kormann等人,nature biotechnol[自然生物技术](2011)29:154
‑
57。此类修饰可用于例如增加rna分子的稳定性和/或降低其免疫原性。使用化学修饰(如假u、n6
‑
甲基
‑
a、2
‑
硫
‑
u和5
‑
甲基
‑
c)发现分别用2
‑
硫
‑
u和5
‑
甲基
‑
c取代尿苷和胞苷残基的四分之一导致小鼠中toll样受体(tlr)介导的mrna识别显著降低。通过减少先天免疫系统的激活,这些修饰可用于有效提高体内mrna的稳定性和寿命;参见例如,kormann等人,同上。
[0273]
还已报道重复施用合成的信使rna(并入了旨在绕过先天抗病毒应答的修饰)可以
将分化的人类细胞重新编程为具有多能性。参见例如,warren等人,cell stem cell[细胞
‑
干细胞](2010)7(5):618
‑
30。充当初级重编程蛋白的此类经修饰的mrna可成为重编程多种人类细胞类型的有效手段。此类细胞称为诱导多能干细胞(ipsc),并且发现并入5
‑
甲基
‑
ctp、假utp和抗反向帽类似物(arca)的酶促合成的rna可用于有效逃避细胞的抗病毒应答;参见例如,warren等人,同上。
[0274]
本领域描述的多核苷酸的其他修饰包括例如使用聚a尾、添加5’帽类似物(诸如m7g(5’)ppp(5’)g(mcap))、对5’或3’非翻译区(utr)的修饰,以及用磷酸酶处理以去除5’端磷酸酯。
[0275]
已经结合对rnai(包括sirna)的修饰开发了适用于产生用于本文的经修饰的rna的许多组合物和技术。sirna在体内面临特别的挑战,因为它们经由mrna干扰对基因沉默的影响通常是暂时的,可能需要重复施用。另外,sirna是双链rna(dsrna),并且哺乳动物免疫应答已经进化为检测和中和dsrna,该dsrna通常是病毒感染的副产物。因此,存在可以介导对dsrna的细胞应答的哺乳动物酶如pkr(dsrna响应激酶)和潜在视黄酸诱导型基因i(rig
‑
i),以及可响应于此类分子而触发对细胞因子的诱导的toll样受体(如tlr3、tlr7和tlr8);参见例如以下综述:angart等人,pharmaceuticals[药物](basel)(2013)6(4):440
‑
68;kanasty等人,mol ther[分子疗法](2012)20(3):513
‑
24;burnett等人,biotechnol j[生物技术杂志](2011)6(9):1130
‑
46;judge和maclachlan,hum gene ther[人类基因疗法](2008)19(2):111
‑
24;以及其中引用的参考文献。
[0276]
如本文所述,已经开发出多种修饰并将其应用于提高rna稳定性、减少先天免疫应答、和/或获得可用于结合将多核苷酸引入人类细胞中的其他益处;参见例如以下综述:k.a.whitehead等人,ann rev chem biomol eng[化学与生物分子工程年度报告](2011)2:77
‑
96;gaglione和messere,mini rev med chem[药物化学简评](2010)10(7):578
‑
95;chernolovskaya等人,curr opin mol ther[分子治疗学最新观点](2010)12(2):158
‑
67;deleavey等人,curr protoc nuc acid chem[核酸化学实验操作手册],第16章:第16.3单元(2009);behlke,oligonucleotides[寡核苷酸](2008)18(4):305
‑
19;fucini等人,nucleic acid ther[核酸疗法](2012)22(3):205
‑
210;bremsen等人,front genet[基因学前沿](2012)3:154。
[0277]
存在许多经修饰的rna的商业供应商,其中许多专门从事于旨在改善sirna有效性的修饰。基于文献中报道的发现,提供了多种方法。例如,dharmacon指出,用硫(硫代磷酸酯,ps)替换非桥连氧已被广泛用于改善sirna的核酸酶抗性,如kole在nature rev drug disc[自然评论:药物发现](2012)11:125
‑
40所报道的。已经报道核糖2
’‑
位置的修饰可改善核苷酸间磷酸键的核酸酶抗性,同时增加双链体稳定性(t
m
),还已经报道这可提供保护免受免疫激活。适度的ps骨架修饰与小的、良好耐受的2
’‑
取代(2
’‑
o
‑
甲基、2
’‑
氟、2
’‑
氢)的组合与用于体内应用的高度稳定的sirna相关,如soutschek等人nature[自然](2004)432:173
‑
78所报道的;并且已经报道2
’‑
o
‑
甲基修饰有效改善稳定性,如通过volkov,oligonucleotides[寡核苷酸](2009)19:191
‑
202所报道的。关于减少对先天免疫应答的诱导,据报道用2
’‑
o
‑
甲基、2
’‑
氟、2
’‑
氢修饰特定序列可减少tlr7/tlr8相互作用,同时通常保持沉默活性;参见例如judge等人,mol ther[分子疗法](2006)13:494
‑
505;以及cekaite等人,j mol biol[分子生物学杂志](2007)365:90
‑
108。还已报道其他修饰(诸如2
‑
硫尿嘧
啶、假尿嘧啶、5
‑
甲基胞嘧啶、5
‑
甲基尿嘧啶和n6‑
甲基腺苷)可最小化由tlr3、tlr7和tlr8介导的免疫作用;参见例如k.kariko等人,immunity[免疫力](2005)23:165
‑
75。
[0278]
还如本领域中已知的并且可商购的,可以将许多缀合物应用于本文使用的多核苷酸(如rna),以增强这些缀合物的递送和/或细胞摄取,这些缀合物包括例如胆固醇、生育酚和叶酸、脂质、肽、聚合物、接头和适体;参见例如,winkler,ther.deliv.[治疗递送](2013)4:791
‑
809,及其中引用的参考文献。
[0279]
递送
[0280]
在一些实施例中,将本文提供的方法中使用的任何核酸分子,例如编码本披露的靶向基因组的核酸和/或定点多肽的核酸包装到递送载体中或表面上,以递送至细胞。递送媒介物包括但不限于纳米球、脂质体、量子点、纳米颗粒、聚乙二醇颗粒、水凝胶和胶束。如本领域中所述,多种靶向部分可用于增强此类媒介物与所需细胞类型或位置的优先相互作用。
[0281]
本披露的复合物、多肽和核酸可以通过以下方式引入细胞中:病毒或噬菌体感染、转染、缀合、原生质体融合、脂质转染、电穿孔、核转染、磷酸钙沉淀、聚乙烯亚胺(pei)介导的转染、deae
‑
葡聚糖介导的转染、脂质体介导的转染、基因枪技术、磷酸钙沉淀、直接微量注射、纳米颗粒介导的核酸递送等。
[0282]
在实施例中,向导rna多核苷酸(rna或dna)和/或内切核酸酶多核苷酸(rna或dna)通过本领域已知的病毒或非病毒递送媒介物递送。替代性地,可以通过本领域已知的病毒或非病毒递送媒介物,诸如电穿孔或脂质纳米颗粒来递送定点多肽。在一些实施例中,dna内切核酸酶作为一种或多种多肽单独地或与一种或多种grna,或一种或多种crrna与tracrrna一起预复合地递送。
[0283]
在实施例中,多核苷酸通过非病毒递送媒介物递送,非病毒递送媒介物包括但不限于纳米颗粒、脂质体、核糖核蛋白、带正电的肽、小分子rna
‑
缀合物、适体
‑
rna嵌合体和rna
‑
融合蛋白复合物。一些示例性非病毒递送媒介物在peer和lieberman,gene ther[基因疗法](2011)18:1127
‑
33(其重点是用于sirna的非病毒递送媒介物也可用于递送其他多核苷酸)中有描述。
[0284]
在实施例中,多核苷酸(诸如向导rna、sgrna和编码内切核酸酶的mrna)通过脂质纳米颗粒(lnp)递送至细胞或受试者。
[0285]
虽然已经在动物模型和人类中都测试了几种用于核酸的非病毒递送方法,但是最完善的系统是脂质纳米颗粒。lnp通常由可电离的阳离子脂质和三种或更多种附加组分组成,这些附加组分通常为胆固醇、dope和含聚乙二醇(peg)的脂质(参见例如实例1)。阳离子脂质可以与带正电的核酸结合,从而形成避免核酸被降解的致密复合物。在通过微流体系统期间,这些组分自组装以形成大小范围为50至150nm的颗粒,其中核酸包封在与阳离子脂质复合的核中并被脂质双层样结构包围。注射入受试者的循环后,这些颗粒可以与载脂蛋白e(apoe)结合。apoe是ldl受体的配体,并经由受体介导的内吞作用介导摄取到肝脏的肝细胞中。已经报道这种类型的lnp可有效地将mrna和sirna递送至啮齿动物、灵长类动物和人的肝脏的肝细胞。在内吞作用后,lnp存在于内体中。包封的核酸经历了由阳离子脂质的可电离性质介导的内体逃逸过程。这样将核酸递送到细胞质中,在细胞质中mrna可以翻译成所编码的蛋白。因此,在一些实施例中,将grna和编码cas9的mrna包封到lnp中用于在静
脉注射后将两种组分有效地递送至肝细胞。内体逃逸后,cas9 mrna翻译成cas9蛋白,并且与grna形成复合物。在一些实施例中,将核定位信号包含在cas9蛋白序列中促进cas9蛋白/grna复合物向核的易位。替代性地,小grna穿过核孔复合物,并与核中的cas9蛋白形成复合物。一旦处于核中,grna/cas9复合物就会扫描基因组中的同源靶位点,并优先在基因组中的所需靶位点处产生双链断裂。rna分子在体内的半衰期很短,大约数小时至数天。类似地,蛋白的半衰期往往很短,大约数小时至数天。因此,在一些实施例中,使用lnp递送grna和cas9 mrna可以仅引起grna/cas9复合物的瞬时表达和活性。在一些实施例中,这可以提供降低脱靶裂解频率的优点,从而使遗传毒性的风险降至最低。lnp通常免疫原性低于病毒颗粒。虽然许多人对aav有预先存在的免疫力,但对lnp却没有预先存在的免疫力。不太可能发生针对lnp的额外适应性免疫应答,这使得能够重复进行lnp给药。
[0286]
当向受试者施用基于基因编辑的基因疗法(其中将治疗性编码序列整合到宿主基因组基因座,诸如安全港基因座中)时,达到为受试者提供最佳治疗益处的基因表达水平将是有利的。例如,在血友病a中,血液中最理想的fviii蛋白水平将在正常水平的20%至100%、30%至100%、40%至100%或50%至100%的范围内。使用强启动子驱动来自aav基因组的游离型拷贝的治疗性编码序列的表达的基于aav的标准基因疗法,无法实现对所达到的表达水平的控制,因为aav病毒只可以给药一次,并且所达到的表达水平在受试者之间显著不同(s.rangarajan等人,n engl j med[新英格兰医学杂志](2017)377:2519
‑
30)。对受试者进行aav病毒给药后,他或她会产生针对病毒衣壳蛋白的高滴度抗体,基于临床前模型,这些抗体预计会阻止病毒的有效重新施用(h.petry等人,gene ther[基因疗法](2008)15:54
‑
60)。通过aav病毒而递送的治疗基因整合到基因组中的安全港基因座诸如白蛋白内含子1并且这种靶向整合经由在基因组中产生双链断裂而发生的一种方法提供了控制靶向整合水平,从而控制治疗性编码序列产物水平的机会。在通过包封含有编码合成fviii的供体dna盒的aav基因组的aav转导肝脏之后,aav基因组在转导细胞的细胞核内维持游离状态。这些游离型aav基因组随时间推移相对稳定,因此为在由crispr/cas9产生的双链断裂处进行靶向整合提供了供体模板库。
[0287]
已经开发出几种不同的可电离的阳离子脂质用于lnp。这些包括c12
‑
200(k.t.love等人,proc natl acad sci usa[美国国家科学院院刊](2010)107:1864
‑
69)、mc3(m.jayaraman等人,angew chem int ed engl[德国应用化学](2012)51:8529
‑
33)、ln16和md1(fougerolles等人,美国专利号8754062)等。c12
‑
200是1,1
’‑
((2
‑
(4
‑
(2
‑
((2
‑
(双(2
‑
羟基十二烷基)氨基)乙基)(2
‑
羟基十二烷基)氨基)乙基)哌嗪
‑1‑
基)乙基)氮烷二基)双(十二烷
‑2‑
醇)。在一种类型的lnp中,galnac部分附接到lnp的外部,并充当经由脱唾液酸糖蛋白受体摄取到肝脏中的配体。这些阳离子脂质中的任一种均可用于配制lnp,以将grna和cas9 mrna递送至肝脏。
[0288]
在一些实施例中,lnp的直径小于约1000nm、500nm、250nm、200nm、150nm、100nm、75nm、50nm或25nm。替代性地,纳米颗粒的大小范围可以是约1
‑
1000nm、1
‑
500nm、1
‑
250nm、25
‑
200nm、25
‑
100nm、35
‑
75nm或25
‑
60nm。
[0289]
lnp可以由阳离子、阴离子或中性脂质制成。中性脂质(诸如融合磷脂dope或膜组分胆固醇)可以作为“辅助脂质”包含在lnp中,以增强转染活性和纳米颗粒稳定性。阳离子脂质的局限性可以包括由于稳定性差和快速清除以及产生炎性反应或抗炎反应而导致的
功效低下。lnp还可以具有疏水性脂质、亲水性脂质,或者具有疏水性和亲水性脂质两者。
[0290]
本领域已知的任何脂质或脂质组合均可用于产生lnp。用于产生lnp的脂质的实例包括:dotma、dospa、dotap、dmrie、dc
‑
胆固醇、dotap
‑
胆固醇、gap
‑
dmorie
‑
dpype和gl67a
‑
dope
‑
dmpe
‑
聚乙二醇(peg)。阳离子脂质的实例包括98n12
‑
5、c12
‑
200、dlin
‑
kc2
‑
dma(kc2)、dlin
‑
mc3
‑
dma(mc3)、xtc、md1和7c1。中性脂质的实例包括dpsc、dppc、popc、dope和sm。peg修饰的脂质的实例包括peg
‑
dmg、peg
‑
cerc14和peg
‑
cerc20。
[0291]
在实施例中,脂质可以以任何数目的摩尔比组合以产生lnp。另外,一种或多种多核苷酸可以以宽范围的摩尔比与一种或多种脂质组合以产生lnp。
[0292]
在实施例中,定点多肽和靶向基因组的核酸可各自单独施用于细胞或受试者。定点多肽可以与一种或多种向导rna、或者一种或多种crrna以及tracrrna预复合。然后可以将预复合材料施用至细胞或受试者。这种预复合材料称为核糖核蛋白颗粒(rnp)。
[0293]
rna能够与rna或dna形成特异性相互作用。虽然在许多生物学过程中都利用了这种特性,但它也伴随着在富含核酸的细胞环境中发生混杂相互作用的风险。该问题的一种解决方案是形成核糖核蛋白颗粒(rnp),其中rna与内切核酸酶预复合。rnp的另一个益处是避免rna被降解。
[0294]
在一些实施例中,rnp中的内切核酸酶是经修饰的或未修饰的。同样,grna、crrna、tracrrna或sgrna可以是经修饰的或未修饰的。许多修饰是本领域已知的并且可以使用。
[0295]
内切核酸酶和sgrna通常以约1:1的摩尔比组合。替代性地,内切核酸酶、crrna和tracrrna可以以约1:1:1的摩尔比组合。然而,可以使用宽范围的摩尔比来生产rnp。
[0296]
在一些实施例中,将重组aav载体用于递送。用于产生raav颗粒的技术(其中向细胞提供待包装的aav基因组(其包括待递送的多核苷酸、rep和cap基因以及辅助病毒功能))在本领域是标准的。产生raav需要单个细胞(本文称为包装细胞)内存在以下组分:raav基因组,与raav基因组分开(即不在其中)的aav rep和cap基因,以及辅助病毒功能。aav rep和cap基因可以来自可衍生重组病毒的任何aav血清型,并且可以来自与raav基因组itr不同的aav血清型,包括但不限于aav血清型aav
‑
1、aav
‑
2、aav
‑
3、aav
‑
4、aav
‑
5、aav
‑
6、aav
‑
7、aav
‑
8、aav
‑
9、aav
‑
10、aav
‑
11、aav
‑
12、aav
‑
13和aav rh.74。假型raav的产生在例如国际专利申请wo 01/83692中披露。参见表1。
[0297]
表1.选定的aav的aav血清型和基因库登录号。
[0298]
aav血清型基因库登录号aav
‑
1nc_002077.1aav
‑
2nc_001401.2aav
‑
3nc_001729.1aav
‑
3baf028705.1aav
‑
4nc_001829.1aav
‑
5nc_006152.1aav
‑
6af028704.1aav
‑
7nc_006260.1aav
‑
8nc_006261.1aav
‑
9ax753250.1
aav
‑
10ay631965.1aav
‑
11ay631966.1aav
‑
12dq813647.1aav
‑
13eu285562.1
[0299]
在一些实施例中,产生包装细胞的方法,对于aav颗粒产生而言,涉及产生稳定表达必需组分的细胞系。例如,将具有以下项的质粒(或多个质粒)整合到细胞的基因组中:缺少aav rep和cap基因的raav基因组,与raav基因组分开的aav rep和cap基因,以及选择性标记(诸如新霉素耐药性基因)。已经通过诸如gc加尾(r.j.samulski等人,proc natl acad sci usa[美国国家科学院院刊](1982)79:2077
‑
81),添加含有限制性内切核酸酶裂解位点的合成接头(c.a.laughlin等人,gene[基因](1983)23:65
‑
73),以及通过直接平端连接(p.senapathy等人,j biol chem[生物化学杂志](1984)259:4661
‑
66)等程序将aav基因组引入到细菌质粒中。然后用辅助病毒(诸如腺病毒)感染包装细胞系。该方法的优点是,细胞是可选择的,并且适合raav的大规模生产。其他合适的方法采用腺病毒或杆状病毒而不是质粒来将raav基因组和/或rep和cap基因引入包装细胞中。
[0300]
产生raav的一般原理在例如b.j.carter,cur op biotechnol[生物技术当前述评](1992)3(5):533
‑
39;以及n.muzyczka,curr topics microbiol immunol[微生物学和免疫学的当前主题](1992)158:97
‑
129)中有综述。在以下参考文献中描述了一些方法:j.d.tratschin等人,mol cell biol[分子细胞生物学](1984)4:2072
‑
81;p.l.hermonat等人,proc natl acad sci usa[美国国家科学院院刊](1984)81:6466
‑
70;j.d.tratschin等人,mol cell biol[分子细胞生物学](1985)5:3251
‑
60;s.k.mclaughlin等人,j virol[病毒学杂志](1988)62:1963
‑
73;j.s.lebkowski等人,mol cell biol[分子细胞生物学](1988)8:3988
‑
96;r.j.samulski等人,j virol[病毒学杂志](1989)63:3822
‑
28);美国专利号5,173,414;wo 95/13365和对应的美国专利号5,658.776;wo 95/13392;wo 96/17947;pct/us 98/18600;wo 97/09441(pct/us 96/14423);wo 97/08298(pct/us 96/13872);wo 97/21825(pct/us 96/20777);wo 97/06243(pct/fr 96/01064);wo 99/11764;p.perrin等人,vaccine[疫苗](1995)13:1244
‑
50;r.w.paul等人,human gene ther[人类基因疗法](1993)4:609
‑
15;clark等人,gene ther[基因疗法](1996)3:1124
‑
32;美国专利号5,786,211;美国专利号5,871,982;和美国专利号6,258,595。
[0301]
aav载体血清型可以与靶细胞类型匹配。例如,以下示例性细胞类型可以用其中的指定aav血清型进行转导。例如,适于肝脏组织/细胞类型的aav载体的血清型包括但不限于aav3、aav5、aav8和aav9。
[0302]
除了腺相关病毒载体,还可以使用其他病毒载体。此类病毒载体包括但不限于慢病毒、α病毒、肠病毒、瘟病毒、杆状病毒、疱疹病毒、eb病毒、乳多空病毒(papovavirus)、痘病毒、疫苗病毒和单纯疱疹病毒。
[0303]
在一些实施例中,将cas9 mrna、靶向白蛋白基因中一个或两个基因座的sgrna和供体dna各自单独地配制为脂质纳米颗粒,或全部共同配制为一个脂质纳米颗粒,或共同配制为两个或更多个脂质纳米颗粒。
[0304]
在一些实施例中,将cas9 mrna配制为脂质纳米颗粒,而sgrna和供体dna以aav载体递送。在一些实施例中,将cas9 mrna和sgrna共同配制为脂质纳米颗粒,而将供体dna以
aav载体递送。
[0305]
可以选择以dna质粒、mrna或蛋白的形式递送cas9核酸酶。向导rna可以从相同的dna表达,或者也可以作为rna递送。可以对rna进行化学修饰以改变或改善其半衰期、或降低免疫应答的可能性或程度。内切核酸酶蛋白可以在递送前与grna复合。病毒载体允许有效递送:分离的cas9型式和较小的cas9直系同源物可以包装在aav中,用于hdr的供体也可以包装在aav中。还存在可以递送这些组分中的每一个的一系列非病毒递送方法,或者可以串联使用非病毒和病毒方法。例如,纳米颗粒可用于递送蛋白和向导rna,而aav可用于递送供体dna。
[0306]
在与递送用于治疗性治疗的基因组编辑组分有关的一些实施例中,将至少以下两种组分递送到待转化的细胞(例如肝细胞)的核中:序列特异性核酸酶和dna供体模板。在一些实施例中,将供体dna模板包装到对肝脏具有嗜性的aav中。在一些实施例中,aav选自血清型aav8、aav9、aavrh10、aav5、aav6或aav
‑
dj。在一些实施例中,将aav包装的dna供体模板施用给受试者,例如,首先通过外周静脉内注射施用给受试者,然后施用序列特异性核酸酶。先递送aav包装的供体模板的优点在于,所递送的供体模板将被稳定地保持在转导的肝细胞的核中,这允许随后施用序列特异性核酸酶。这将在基因组中产生双链断裂,随后通过hdr或nhej整合供体模板。在一些实施例中,期望序列特异性核酸酶仅在促进转基因以达到所需治疗效果的足够水平靶向整合所需的时间内在靶细胞中保持活性。如果序列特异性核酸酶在细胞中保持活性较长时间,这将导致脱靶位点处双链断裂的频率增加。具体地说,脱靶裂解的频率是脱靶切割效率乘以核酸酶具有活性的时间的函数。由于mrna和翻译的蛋白在细胞中的寿命很短,因此以mrna形式递送序列特异性核酸酶导致核酸酶活性持续时间很短,在几小时至几天的范围内。因此,预计将序列特异性核酸酶递送至已经含有供体模板的细胞中会导致靶向整合相对于脱靶整合的更佳比率。另外,在外周静脉注射后,aav介导供体模板递送至肝细胞核需要时间,通常大约为1至14天,这是因为病毒感染细胞,使内体逃逸并转运至细胞核并通过宿主组分将单链aav基因组转化为双链dna分子需要时间。因此,在一些实施例中,在提供crispr/cas9组分之前完成供体模板向细胞核的递送,因为这些核酸酶组分将仅在约1至3天内有活性。
[0307]
在一些实施例中,dna内切核酸酶是crispr/cas9,其由针对白蛋白基因内含子1内的dna序列的sgrna与cas9核酸酶一起组成。在一些实施例中,cas9内切核酸酶作为编码可操作地融合至一个或多个核定位信号(nls)的cas9蛋白的mrna递送。在一些实施例中,sgrna和cas9 mrna包装在脂质纳米颗粒中递送至肝细胞。在一些实施例中,脂质纳米颗粒含有脂质c12
‑
200(k.t.love等人,proc natl acad sci usa[美国国家科学院院刊](2010)107:1864
‑
69)。在一些实施例中,包装在lnp中的sgrna与cas9mrna的比率为1:1(质量比),在小鼠体内引起最大限度的dna裂解。在替代性实施例中,可以使用包装在lnp中的sgrna与cas9 mrna的不同质量比,例如10:1、9:1、8:1、7:1、6:1、5:1、4:1、3:1或2:1或反向比率。在一些实施例中,将cas9 mrna和sgrna包装到单独的lnp配制品中,并且在含有sgrna的lnp之前约1至约8小时将含有cas9 mrna的lnp递送至受试者,以在递送sgrna之前留出翻译cas9 mrna的最佳时间。
[0308]
在一些实施例中,将包封grna和cas9 mrna的lnp配制品(“lnp
‑
核酸酶配制品”)施用于受试者,例如先前施用了包装到aav中的dna供体模板的受试者。在一些实施例中,在施
用aav供体模板后的1天至28天内,或7天至28天内,或7天至14天内将lnp
‑
核酸酶配制品施用于受试者。相对于aav供体模板,lnp
‑
核酸酶配制品的最佳递送时间可以使用本领域已知的技术来确定,例如在包括小鼠和猴子在内的动物模型中进行的研究。
[0309]
在一些实施例中,使用非病毒递送方法将dna
‑
供体模板递送至受试者(例如受试者)的肝细胞。虽然一些受试者(通常为30%)具有针对最常用的aav血清型的预先存在的中和抗体,从而阻止了aav的有效基因递送,但是所有受试者均可通过非病毒递送方法治疗。在本领域中已知几种非病毒递送方法。具体而言,已知lnp在动物和人类中静脉注射后,有效地将其包封的货物递送至肝细胞的细胞质。这些lnp通过受体介导的内吞作用过程被肝脏主动吸收,导致优先摄取到肝脏中。
[0310]
在一些实施例中,为了促进供体模板的核定位,可以促进质粒的核定位的dna序列,例如猿猴病毒40(sv40)复制起点和早期启动子的366bp区域可以添加到供体模板上。与细胞蛋白结合的其他dna序列也可用于改善dna的核进入。
[0311]
在一些实施例中,在aav供体模板之后首次施用例如含有grna和cas9核酸酶或编码cas9核酸酶的mrna的lnp
‑
核酸酶配制品之后,测量受试者(例如受试者)血液中引入的fviii的表达或活性水平。如果fviii水平不足以治疗疾病,例如水平为正常水平的5%,则可以第二次或第三次施用lnp
‑
核酸酶配制品以促进向基因组安全港基因座的附加靶向整合。可以使用本领域已知的技术,例如使用包括小鼠和猴子在内的动物模型进行的试验来测试和优化使用多个剂量的lnp
‑
核酸酶配制品以获得fviii的所需治疗水平的可行性。
[0312]
在一些实施例中,根据本文所述的包括向受试者施用i)包含供体盒的aav供体模板和ii)lnp
‑
核酸酶配制品的任何方法,在向受试者施用aav供体模板后的约1天至约28天内向受试者施用初始剂量的lnp
‑
核酸酶配制品。在一些实施例中,在足以允许将供体模板递送至靶细胞核的时间后,向受试者施用初始剂量的lnp
‑
核酸酶配制品。在一些实施例中,在足以允许单链aav基因组在靶细胞核中转化为双链dna分子的时间后,向受试者施用初始剂量的lnp
‑
核酸酶配制品。在一些实施例中,在施用初始剂量之后,向受试者施用一个或多个(诸如两个、三个、四个、五个或更多个)附加剂量的lnp
‑
核酸酶配制品。在一些实施例中,向受试者施用一个或多个剂量的lnp
‑
核酸酶配制品,直至达到供体盒的目标靶向整合水平和/或供体盒的目标表达水平。在一些实施例中,该方法进一步包括在每次施用lnp
‑
核酸酶配制品之后测量供体盒的靶向整合水平和/或供体盒的表达水平,并且如果未达到供体盒的目标靶向整合水平和/或供体盒的目标表达水平,则施用附加剂量的lnp
‑
核酸酶配制品。在一些实施例中,这些附加剂量的lnp
‑
核酸酶配制品中的至少一个的量与初始剂量相同。在一些实施例中,这些附加剂量的lnp
‑
核酸酶配制品中的至少一个的量小于初始剂量。在一些实施例中,这些附加剂量的lnp
‑
核酸酶配制品中的至少一个的量大于初始剂量。
[0313]
经遗传修饰的细胞和细胞群
[0314]
一方面,本文的披露提供了一种编辑细胞中的基因组,从而产生经遗传修饰的细胞的方法。在一些方面,提供了经遗传修饰的细胞群。因此“经遗传修饰的细胞”是指具有至少一个通过基因组编辑(例如,使用crispr/cas9/cpf1系统)而引入的遗传修饰的细胞。在一些实施例中,经遗传修饰的细胞是经遗传修饰的肝细胞。本文考虑了具有靶向基因组的外源核酸和/或编码靶向基因组的核酸的外源核酸的经遗传修饰的细胞。
[0315]
在一些实施例中,可以通过将合成fviii编码序列的核酸序列插入细胞的基因组
序列中来编辑细胞的基因组。在一些实施例中,经受基因组编辑的细胞在基因组中具有一个或多个突变,突变导致内源性fviii基因的表达与没有此类突变的正常细胞中的表达相比降低。正常细胞可以是来源于(或分离自)没有fviii基因缺陷的不同受试者的健康细胞或对照细胞。在一些实施例中,经受基因组编辑的细胞可以来源于(或分离自)需要治疗fviii基因相关病状或病症的受试者。因此,在一些实施例中,这种细胞中内源性fviii基因的表达与正常细胞中内源性fviii基因的表达相比降低约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%或约100%。
[0316]
成功插入转基因,例如编码合成fviii编码序列的核酸后,所引入的合成fviii编码序列在细胞中的表达与细胞内源fviii基因的表达相比,可以是至少约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%、约100%、约200%、约300%、约400%、约500%、约600%、约700%、约800%、约900%、约1,000%、约2,000%、约3,000%、约5,000%、约10,000%或更多。在一些实施例中,在经基因组编辑的细胞中引入的fviii编码序列产物(包括合成fviii编码序列)的活性与细胞内源fviii基因的表达相比,可以是至少约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%、约100%、约200%、约300%、约400%、约500%、约600%、约700%、约800%、约900%、约1,000%、约2,000%、约3,000%、约5,000%、约10,000%或更多。在一些实施例中,所引入的合成fviii编码序列在细胞中的表达是细胞内源fviii基因的表达的至少约2倍、约3倍、约4倍、约5倍、约6倍、约7倍、约8倍、约9倍、约10倍、约15倍、约20倍、约30倍、约50倍、约100倍、约1000倍或更多。而且,在一些实施例中,在经基因组编辑的细胞中引入的合成fviii编码序列产物的活性可以与正常健康细胞中fviii基因产物的活性相当或更高。
[0317]
在用于治疗或缓解血友病a的实施例中,用于基因编辑的主要靶标是人细胞。在一些实施例中,在离体方法和体内方法中,人细胞是肝细胞。在一些实施例中,通过在源自有需要的受试者(并因此已经完全与受试者匹配)的自体细胞中进行基因编辑,可以产生细胞,这些细胞可以安全地重新引入受试者体内,并有效地产生在缓解与受试者疾病相关的一种或多种临床状况的方面有效的细胞群。在此类治疗的一些实施例中,根据本领域已知的任何方法分离肝细胞,并用于产生经遗传修饰的、治疗上有效的细胞。在一个实施例中,将肝脏干细胞离体进行遗传修饰,然后重新引入受试者体内,在受试者体内它们将会产生表达插入的fviii编码序列的经遗传修饰的肝细胞或窦内皮细胞。
[0318]
治疗方法
[0319]
血友病分为“轻度”(fviii蛋白血清浓度为0.40至0.05iu/ml)、“中度”(0.05至0.01iu/ml)或“重度”(<0.01iu/ml,小于正常值的1%)(g.c.white等人,thromb haemost[血栓与止血学](2001)85(3):560
‑
75)。对接受fviii替代蛋白疗法的血友病a患者的分析报告,在预测的fviii谷水平为正常水平的3%、5%、10%、15%和20%时,没有出血发生的频率分别为71%、79%、91%、97%和100%(g.spotts等人,blood[血液](2014)124:689)。这表明当fviii水平维持在15%至20%的最低水平之上时,出血事件的比率降低到接近于零。虽然治愈血友病a所需的精确fviii水平尚未定义,并且可能因受试者而异,但预计约5%至约30%的水平会引起出血事件的显著减少。
[0320]
一方面,本文提供了一种通过编辑受试者的基因组来治疗受试者的血友病a的基因治疗方法。在一些实施例中,该基因治疗方法将功能性合成fviii编码序列整合到受试者
相关细胞类型的基因组中,并且永久治愈血友病a。在一些实施例中,将合成fviii编码序列整合到肝细胞中,因为这些细胞会有效地表达许多蛋白并分泌到血液中。另外,对于肝脏未生长完全的小儿受试者,可以考虑这种使用肝细胞的整合方法,因为整合的编码序列会随着肝细胞分裂而传递给子细胞。
[0321]
另一方面,本文提供了使用基因组工程工具通过将合成fviii编码序列敲入基因座中并恢复fviii蛋白活性而对基因组产生永久性变化的细胞离体和体内方法。此类方法使用内切核酸酶,诸如crispr相关的(crispr/cas9、cpf1等)核酸酶,使任何序列从基因组中永久性缺失、插入、编辑、矫正或替代,或在基因组基因座中插入外源序列,例如合成fviii编码序列。以这种方式,本披露中阐述的实例通过单一治疗恢复了fviii基因的活性(而不是在受试者的一生中都要递送潜在治疗)。
[0322]
在一些实施例中,基于离体细胞的治疗使用从受试者分离的肝细胞。使用本文所述的材料和方法编辑这些细胞的染色体dna。最后,将经过编辑的细胞和/或其后代施用或植入受试者中。
[0323]
离体细胞治疗方法的一个优点是能够在施用前对治疗剂进行全面分析。基于核酸酶的治疗剂具有一定程度的脱靶作用。离体进行基因矫正允许人们能够在施用前表征经矫正的细胞群。本披露的方面包括对矫正细胞的基因组进行测序,以确保任何脱靶切割位于与对受试者的最小风险相关的基因组位置。此外,可以在施用或移植之前筛选或分离出特定细胞群,包括克隆群体。
[0324]
另一个实施例是基于体内的疗法。在这种方法中,使用本文所述的材料和方法矫正受试者体内细胞的染色体dna。在一些实施例中,细胞是肝细胞。
[0325]
体内基因疗法的一个优势是易于产生和施用治疗剂。可以使用相同的治疗方法和疗法来治疗多于一名受试者,例如,共有相同或相似基因型或等位基因的多名受试者。相反,离体细胞疗法通常使用受试者自身的细胞,将这些细胞分离、操纵并返回至同一受试者。
[0326]
在一些实施例中,该受试者具有血友病a的症状。在一些实施例中,该受试者是怀疑患有血友病a的人。替代性地,该受试者是被诊断为具有血友病a风险的人。在一些实施例中,需要治疗的受试者可以在内源fviii基因或其调控序列中具有一个或多个遗传缺陷(例如缺失、插入和/或突变),使得fviii蛋白的活性(包括表达水平或功能性),与正常健康受试者相比大大降低。
[0327]
在一些实施例中,本文提供了一种治疗受试者的血友病a的方法,该方法包括向受试者的细胞提供以下物质:(a)靶向细胞基因组中的白蛋白基因座的grna;(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板。在一些实施例中,grna靶向白蛋白基因的内含子1。在一些实施例中,grna包含来自seq id no:271
‑
298中任一个的间隔区序列。
[0328]
在一些实施例中,本文提供了一种治疗受试者的血友病a的方法,该方法包括向受试者的细胞提供以下物质:(a)包含来自seq id no:271
‑
298中任一个的间隔区序列的grna;(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板。在一些实施例中,grna包含来自seq id no:274、275、281和283中任一个的间隔区序列。在一些实施例中,grna包含来自seq id no:274的间隔区序列。在一
些实施例中,grna包含来自seq id no:275的间隔区序列。在一些实施例中,grna包含来自seq id no:281的间隔区序列。在一些实施例中,grna包含来自seq id no:283的间隔区序列。在一些实施例中,细胞是人细胞,例如人肝脏细胞。在一些实施例中,受试者是患有或怀疑患有血友病a的受试者。在一些实施例中,受试者被诊断为具有血友病a的风险。
[0329]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,该dna内切核酸酶选自由以下项组成的组:cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4和cpf1内切核酸酶及其功能等效物。在一些实施例中,dna内切核酸酶是cas9。在一些实施例中,cas9是spcas9。在一些实施例中,cas9是slucas9。
[0330]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,对编码合成fviii编码序列的核酸序列进行密码子优化以在细胞中表达。在一些实施例中,细胞是人细胞。
[0331]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,该方法采用编码dna内切核酸酶的核酸。在一些实施例中,对编码dna内切核酸酶的核酸进行密码子优化以在细胞中表达。在一些实施例中,细胞是人细胞,例如人肝脏细胞。在一些实施例中,编码dna内切核酸酶的核酸是dna,诸如dna质粒。在一些实施例中,编码dna内切核酸酶的核酸是rna,诸如mrna。
[0332]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,供体模板在aav载体中编码。在一些实施例中,供体模板包含供体盒,该供体盒包含编码合成fviii蛋白的核酸序列,并且该供体盒在一侧或两侧上侧接有grna靶位点。在一些实施例中,供体盒在两侧上侧接有grna靶位点。在一些实施例中,该grna靶位点是所施用的grna的靶位点。在一些实施例中,供体模板的grna靶位点是施用的grna的细胞基因组grna靶位点的反向互补序列。在一些实施例中,向该细胞提供该供体模板包括向该受试者施用该供体模板。在一些实施例中,该施用是静脉内。
[0333]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,dna内切核酸酶或编码该dna内切核酸酶的核酸配制在脂质体或lnp中。在一些实施例中,脂质体或lnp还包含grna。在一些实施例中,向该细胞提供该grna和该dna内切核酸酶或编码该dna内切核酸酶的核酸包括向该受试者施用该脂质体或lnp。在一些实施例中,该施用是静脉内。在一些实施例中,该脂质体或lnp是lnp。在一些实施例中,该方法采用lnp,该lnp包含编码dna内切核酸酶的核酸和grna。在一些实施例中,编码dna内切核酸酶的核酸是编码dna内切核酸酶的mrna。
[0334]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,dna内切核酸酶与grna预复合,形成rnp复合物。
[0335]
aav感染细胞(包括肝脏细胞)的过程涉及从内体逃逸,病毒脱壳以及aav基因组转运至细胞核。在这些研究中使用的aav的情况下,其中单链基因组包装在病毒中,单链基因组经历第二链dna合成形成双链dna基因组的过程。将单链基因组完全转化为双链基因组所需的时间尚未完全确定,但认为是限速步骤(ferrari等人,j virol[病毒学杂志](1996)70:3227
‑
34)。然后,双链线性基因组串接成多聚体环状形式,该聚体环状形式由头
‑
尾和
尾
‑
头相连的单体组成(sun等人,human gene ther.[人类基因疗法](2010)21:750
‑
62)。
[0336]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,在将供体模板提供给细胞之后,将施用的grna和施用的dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,在将该供体模板提供给该细胞后超过4天,将施用的grna和施用的dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,在将该供体模板提供给该细胞后至少14天,将grna和dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,在将该供体模板提供给该细胞后至少17天,将grna和dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。在一些实施例中,向细胞提供grna和dna内切核酸酶包括向受试者施用(诸如通过静脉内途径施用)包含编码dna内切核酸酶的核酸和grna的lnp。在一些实施例中,编码dna内切核酸酶的核酸是编码dna内切核酸酶的mrna。在一些实施例中,向细胞提供供体模板包括向受试者施用(诸如通过静脉内途径施用)在aav载体中编码的供体模板。
[0337]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,在第一剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸。在一些实施例中,在第一剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的grna和dna内切核酸酶或编码该dna内切核酸酶的核酸,直至达到该编码合成fviii蛋白的核酸序列的目标靶向整合水平和/或该编码合成fviii蛋白的核酸序列的目标表达水平。在一些实施例中,向细胞提供grna和dna内切核酸酶包括向受试者施用(诸如通过静脉内途径施用)包含编码dna内切核酸酶的核酸和grna的脂质纳米颗粒。在一些实施例中,编码dna内切核酸酶的核酸是编码dna内切核酸酶的mrna。
[0338]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,编码合成fviii蛋白的核酸序列在内源白蛋白启动子的控制下表达。在一些实施例中,编码合成fviii蛋白的核酸序列在内源转铁蛋白启动子的控制下表达。在一些实施例中,编码合成fviii蛋白的核酸序列在内源纤维蛋白原
‑
α链启动子的控制下表达。
[0339]
在一些实施例中,根据本文所述的治疗血友病a的任何方法,编码合成fviii蛋白的核酸序列在受试者的肝脏中表达。
[0340]
将细胞递送至受试者
[0341]
在一些实施例中,本披露的离体方法涉及将经过基因组编辑的细胞施用到有此需要的受试者中。这可以使用本领域已知的任何肠胃外施用方法来完成。例如,可以将遗传修饰的细胞直接注射到受试者的血液中,直接注射到肝脏中或肝脏附近(植入),或以其他方式施用于受试者。
[0342]
在一些实施例中,本文披露的方法包括通过导致引入的细胞至少部分定位在所需部位,以便产生所需效果的方法或途径,将经遗传修饰的治疗细胞植入或“移植”到受试者体内。可以通过任何适当的途径引入治疗细胞或其分化后代,该途径导致递送至受试者中的所需位置,在该位置中至少一部分植入的细胞或细胞组分保持活力。在施用于受试者后,细胞的活力期可以短至数小时(例如二十四小时)、几天,长达数年,或甚至受试者的寿命(即长期植入)。
[0343]
当以预防方式提供时,本文所述的治疗细胞在血友病a的任何症状之前施用给受
试者。因此,在一些实施例中,经遗传修饰的肝细胞群的预防性施用用于预防血友病a症状的发生。
[0344]
当在一些实施例中以治疗方式提供时,在血友病a症状或指征发作时(或之后),例如在疾病发作时,提供经遗传修饰的肝细胞。
[0345]
在一些实施例中,根据本文所述的方法施用的治疗性肝细胞群具有获自一个或多个供体的同种异体肝细胞。“同种异体”是指肝细胞或具有从相同物种的一个或多个不同供体获得的肝细胞的生物样品,其中一个或多个基因座处的基因是不相同的。例如,施用于受试者的肝细胞群可以源自一个或多个不相关的供体受试者,或源自一个或多个不相同的同胞。在一些实施例中,可以使用同基因肝细胞群,诸如从遗传上相同的动物或从同卵双胞胎获得的那些。在其他实施例中,肝细胞是自体细胞;即,这些肝细胞是从受试者获得或分离并施用给相同受试者的,即,供体和受体是相同的。
[0346]
在一个实施例中,有效量是指预防或减轻血友病a的至少一种或多种体征或症状所需的治疗细胞群的量,并且涉及足以提供所需效果,例如治疗患有血友病a的受试者的组合物的量。因此,在实施例中,治疗有效量是指治疗细胞或具有治疗细胞的组合物,当施用于受试者(诸如患有血友病a或处于血友病a风险中的受试者)时,足以促进特定作用的量。有效量还包括足以预防或延迟疾病症状发展,改变疾病症状过程(例如但不限于,减缓疾病症状的进展),或逆转疾病症状的量。应当理解,对于任何给定的情况,本领域的普通技术人员可以确定适当的有效量。
[0347]
为了用于本文所述的实施例中,治疗细胞(例如经基因组编辑的肝细胞)的有效量可以是至少约102个细胞,至少约5
×
102个细胞,至少约103个细胞,至少约5
×
103个细胞,至少约104个细胞,至少约5
×
104个细胞,至少约105个细胞,至少约2
×
105个细胞,至少约3
×
105个细胞,至少约4
×
105个细胞,至少约5
×
105个细胞,至少约6
×
105个细胞,至少约7
×
105个细胞,至少约8
×
105个细胞,至少约9
×
105个细胞,至少约1
×
106个细胞,至少约2
×
106个细胞,至少约3
×
106个细胞,至少约4
×
106个细胞,至少约5
×
106个细胞,至少约6
×
106个细胞,至少约7
×
106个细胞,至少约8
×
106个细胞,至少约9
×
106个细胞,或其倍数。治疗细胞衍生自一个或多个供体、或获得自自体来源。在本文所述的一些实施例中,治疗细胞在施用于有需要的受试者之前在培养中扩增。
[0348]
在一些实施例中,在患有血友病a的受试者的细胞中表达的功能性fviii水平的适度和增量增加有益于缓解疾病的一种或多种症状,提高长期存活率和/或减少与其他治疗相关的副作用。在将此类细胞施用于人类受试者后,存在产生更高水平的功能性fviii的治疗细胞是有益的。在一些实施例中,对受试者的有效治疗产生相对于所治受试者中的总fviii至少约1%、3%、5%或7%的功能性fviii。在一些实施例中,功能性fviii为总fviii的至少约10%。在一些实施例中,功能性fviii为总fviii的至少、约或至多20%、30%、40%、50%、60%、70%、80%、90%或100%。类似地,引入甚至相对有限的功能性fviii水平显著升高的细胞亚群在受试者中是有益的,因为在一些情况下,标准化细胞相对于患病细胞具有选择优势。然而,即使是适度水平的功能性fviii水平升高的治疗细胞,也有益于缓解受试者的血友病a的一个或多个方面。在一些实施例中,施用此类细胞的受试者中有约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%或更多的治疗剂产生水平增加的功能性fviii。
[0349]
在实施例中,通过一种方法或途径将治疗细胞组合物递送至受试者中导致该细胞组合物至少部分定位在所需部位。可以通过在受试者中导致有效治疗的任何适当途径来施用细胞组合物,即,施用导致递送至受试者中的所需位置,在该位置中递送了至少一部分组合物,即,至少约1
×
104个细胞被递送到所需部位持续一段时间。施用模式包括注射、输注、滴注或摄取。“注射”包括但不限于静脉内、肌内、动脉内、鞘内、心室内、囊内、眶内、心内、真皮内、腹膜内、经气管、皮下、表皮下、关节内、被膜下、蛛网膜下、脊柱内、脑脊髓内和胸骨内注射和输注。在一些实施例中,该途径是静脉内。为了递送细胞,可以通过注射或输注进行施用。
[0350]
在一个实施例中,全身性施用细胞,换句话说,将治疗细胞群以除直接施用至靶部位、组织或器官以外的方式施用,使其进入受试者的循环系统,从而经受代谢和其他类似过程。
[0351]
用于治疗血友病a的组合物的治疗功效可以由熟练的临床医师确定。然而,如果任何一种或多种体征或症状,例如功能性fviii水平以有益的方式发生改变(例如,增加至少10%),或其他临床上接受的症状或疾病标志物得到改善或缓解,则认为治疗是有效的治疗。功效还可以通过如通过住院治疗或需要医疗干预所评估的个体恶化的失败(例如,疾病进展停止或至少减慢)来测量。测量这些指标的方法是本领域技术人员已知的和/或本文所述的。治疗包括对个体或动物(一些非限制性实例包括人或哺乳动物)中疾病的任何治疗,并包括:(1)抑制疾病,例如阻止或减缓症状的进展;或(2)减轻疾病,例如引起症状消退;以及(3)预防或降低症状发展的可能性。
[0352]
组合物
[0353]
一方面,本披露提供了用于实施本文披露的方法的组合物。组合物可以包含以下物质中的一种或多种:靶向基因组的核酸(例如,grna);定点多肽(例如,dna内切核酸酶)或编码定点多肽的核苷酸序列;以及待插入以实现本文披露的方法的期望遗传修饰的多核苷酸(例如,供体模板)。
[0354]
在一些实施例中,组合物具有编码靶向基因组的核酸(例如grna)的核苷酸序列。
[0355]
在一些实施例中,组合物具有定点多肽(例如dna内切核酸酶)。在一些实施例中,组合物具有编码定点多肽的核苷酸序列。
[0356]
在一些实施例中,组合物具有待插入基因组中的多核苷酸(例如供体模板)。
[0357]
在一些实施例中,组合物具有(i)编码靶向基因组的核酸(例如grna)的核苷酸序列和(ii)定点多肽(例如dna内切核酸酶)或编码该定点多肽的核苷酸序列。
[0358]
在一些实施例中,组合物具有(i)编码靶向基因组的核酸(例如grna)的核苷酸序列和(ii)待插入基因组中的多核苷酸(例如供体模板)。
[0359]
在一些实施例中,组合物具有(i)定点多肽(例如dna内切核酸酶)或编码该定点多肽的核苷酸序列,和(ii)待插入基因组中的多核苷酸(例如供体模板)。
[0360]
在一些实施例中,组合物具有(i)编码靶向基因组的核酸的核苷酸序列,(ii)定点多肽或编码该定点多肽的核苷酸序列,和(iii)待插入基因组中的多核苷酸(例如供体模板)。
[0361]
在任何上述组合物的一些实施例中,该组合物具有靶向基因组的单分子向导核酸。在任何上述组合物的一些实施例中,该组合物具有靶向基因组的双分子核酸。在任何上
述组合物的一些实施例中,该组合物具有两个或更多个双分子向导或单分子向导。在一些实施例中,组合物具有编码靶向核酸的核酸的载体。在一些实施例中,靶向基因组的核酸是dna内切核酸酶,尤其是cas9。
[0362]
在一些实施例中,组合物含有一种或多种适于基因组编辑,尤其是适于将合成fviii编码序列插入细胞基因组中的grna。该组合物的grna可以靶向内源白蛋白基因处、内部或附近的基因组位点。在一些实施例中,grna在白蛋白基因处、内部或附近具有与基因组序列互补的间隔区序列。
[0363]
在一些实施例中,组合物的grna是选自seq id no:271
‑
298中任一个的序列及其变体,这些变体与seq id no:271
‑
298中任一个具有至少约50%、约55%、约60%、约65%、约70%、约75%、约80%、约85%、约90%或约95%的同一性或同源性。在一些实施例中,grna的变体与seq id no:271
‑
298中的任一个具有至少约85%的同源性。
[0364]
在一些实施例中,组合物的grna具有与基因组中的靶位点互补的间隔区序列。在一些实施例中,间隔区序列的长度为15个碱基至20个碱基。在一些实施例中,间隔区序列与基因组序列之间的互补性为至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%。
[0365]
在一些实施例中,组合物具有dna内切核酸酶或编码该dna内切核酸酶的核酸和/或具有合成fviii编码序列的核酸序列的供体模板。在一些实施例中,dna内切核酸酶是cas9。在一些实施例中,编码该dna内切核酸酶的核酸是dna或rna。
[0366]
在一些实施例中,任何寡核苷酸或核酸序列中的一个或多个在aav载体中编码。因此,在一些实施例中,grna在aav载体中编码。在一些实施例中,编码dna内切核酸酶的核酸在aav载体中编码。在一些实施例中,供体模板在aav载体中编码。在一些实施例中,两个或更多个寡核苷酸或核酸序列在单个aav载体中编码。因此,在一些实施例中,grna序列和编码dna内切核酸酶的核酸在单个aav载体中编码。
[0367]
在一些实施例中,组合物具有脂质体或脂质纳米颗粒。因此,在一些实施例中,可以将组合物的任何化合物(例如dna内切核酸酶或编码dna内切核酸酶的核酸、grna和供体模板)配制在脂质体或lnp中。在一些实施例中,一种或多种这样的化合物经由共价键或非共价键与脂质体或lnp缔合。在一些实施例中,任何化合物单独地或一起包含在脂质体或lnp中。因此,在一些实施例中,将dna内切核酸酶或编码dna内切核酸酶的核酸、grna和供体模板中的每一种单独地配制在脂质体或lnp中。在一些实施例中,将dna内切核酸酶与grna一起配制在脂质体或lnp中。在一些实施例中,将dna内切核酸酶或编码dna内切核酸酶的核酸、grna和供体模板一起配制在脂质体或lnp中。
[0368]
在一些实施例中,上述组合物还具有一种或多种附加试剂,其中此类附加试剂选自缓冲液、用于将多肽或多核苷酸引入细胞中的缓冲液、洗涤缓冲液、对照试剂、对照载体、对照rna多核苷酸、用于由dna体外产生多肽的试剂、用于测序的衔接子等。缓冲液可以是稳定化缓冲液、重构缓冲液、稀释缓冲液等。在一些实施例中,组合物还包含一种或多种组分,这些组分可用于促进或增强内切核酸酶对dna的中靶结合或裂解,或提高靶向的特异性。
[0369]
在一些实施例中,根据特定的施用模式和剂型,用药学上可接受的赋形剂诸如载剂、溶剂、稳定剂、佐剂、稀释剂等配制组合物的任何组分。在实施例中,通常将向导rna组合物配制成达到生理相容的ph,并且根据配制和施用途径,其范围是ph为约3至ph为约11,约
ph 3至约ph 7。在一些实施例中,将ph调节到约ph 5.0至约ph 8.0的范围。在一些实施例中,组合物具有治疗有效量的至少一种本文所述的化合物,以及一种或多种药学上可接受的赋形剂。任选地,该组合物可以具有本文所述的化合物的组合,可以包含可用于治疗或预防细菌生长的第二活性成分(例如但不限于,抗菌剂或抗微生物剂),并且可以包含本披露的试剂的组合。在一些实施例中,将grna与一种或多种其他寡核苷酸,例如编码dna内切核酸酶的核酸和/或供体模板一起配制。替代性地,用上述用于grna配制的方法单独地或与其他寡核苷酸组合配制编码dna内切核酸酶的核酸和供体模板。
[0370]
合适的赋形剂包括例如载剂分子,这些载剂分子包括大的、缓慢代谢的大分子(如蛋白、多糖、聚乳酸、聚乙醇酸、聚氨基酸、氨基酸共聚物和失活病毒颗粒)。其他示例性赋形剂包括抗氧化剂(例如但不限于抗坏血酸)、螯合剂(例如但不限于edta)、碳水化合物(例如但不限于糊精、羟烷基纤维素和羟烷基甲基纤维素)、硬脂酸、液体(例如但不限于油、水、盐水、甘油和乙醇)、润湿剂或乳化剂、ph缓冲物质等。
[0371]
在一些实施例中,组合物的任何化合物(例如dna内切核酸酶或编码dna内切核酸酶的核酸、grna和供体模板)经由转染诸如电穿孔来递送。在一些示例性实施例中,在提供给细胞之前,将dna内切核酸酶与grna预复合,形成rnp复合物,并且将rnp复合物进行电穿孔。在此类实施例中,供体模板可以经由电穿孔递送。
[0372]
在一些实施例中,“组合物”是指具有用于离体治疗方法中的治疗细胞的治疗性组合物。
[0373]
在实施例中,治疗性组合物含有生理上可耐受的载剂以及作为活性成分溶解或分散在其中的细胞组合物,和任选地至少一种如本文所述的附加生物活性剂。在一些实施例中,当为了治疗目的向哺乳动物或人类受试者施用治疗性组合物时,治疗性组合物基本上没有免疫原性。
[0374]
一般而言,本文所述的经遗传修饰的治疗细胞与药学上可接受的载剂一起作为悬浮液施用。本领域技术人员将认识到,将用于细胞组合物中的药学上可接受的载剂将不包括大大干扰待递送至受试者的细胞的活力的缓冲液、化合物、冷冻保存剂、防腐剂或其他试剂。具有细胞的配制品可以包括例如容许维持细胞膜完整性的渗透缓冲液,以及任选地包括营养素以在施用后维持细胞活力或增强移植。此类配制品和悬浮液是本领域技术人员已知的,和/或可以使其适于与如本文所述的细胞一起使用。
[0375]
在一些实施例中,细胞组合物也可以乳化或作为脂质体组合物存在,条件是乳化过程不会对细胞活力产生不利影响。可以将细胞和任何其他活性成分与药学上可接受的并且与活性成分相容的赋形剂混合,并以适用于本文所述的治疗方法的量混合。
[0376]
细胞组合物中所含的另外的试剂可以包括其中的组分的药学上可接受的盐。药学上可接受的盐包括与无机酸(诸如盐酸或磷酸)或有机酸(诸如乙酸、酒石酸、扁桃酸等)形成的酸加成盐(与多肽的游离氨基基团形成)。与游离羧基基团形成的盐也可以衍生自无机碱(诸如氢氧化钠、氢氧化钾、氢氧化铵、氢氧化钙或氢氧化铁)和有机碱(诸如异丙胺、三甲胺、2
‑
乙基氨基乙醇、组氨酸、普鲁卡因等)。
[0377]
生理上可耐受的载剂是本领域众所周知的。示例性液体载剂是无菌水溶液,这些无菌水溶液除了活性成分和水外不含任何材料,或者含有缓冲液(如生理ph值的磷酸钠、生理盐水或两者,如磷酸盐缓冲盐水)。更进一步,水性载剂可以含有多于一种缓冲盐,以及诸
如氯化钠和氯化钾的盐、右旋糖、聚乙二醇和其他溶质。除了并排除水之外,液体组合物还可以含有液相。此类另外的液相的实例是甘油、植物油(如棉籽油)和水油乳剂。在细胞组合物中有效治疗特定病症或病状的活性化合物的量将取决于病症或病状的性质,并且可以通过标准临床技术来确定。
[0378]
试剂盒
[0379]
一些实施例提供了一种试剂盒,该试剂盒包含任何上述组合物,例如用于基因组编辑的组合物或治疗性细胞组合物以及一种或多种附加组分。
[0380]
在一些实施例中,试剂盒可具有一种或多种附加治疗剂,这些附加治疗剂可与组合物同时或按顺序施用以达到所需目的,例如基因组编辑或细胞疗法。
[0381]
在一些实施例中,试剂盒可进一步包括用于使用试剂盒的组分来实践这些方法的说明书。通常将用于实践这些方法的说明书记录在合适的记录介质上。例如,说明书可以印刷在诸如纸或塑料等的基质上。说明书可以作为包装插页存在于试剂盒中,存在于试剂盒或其组件的容器的标签中(即,伴同包装或分包装一起)等。说明书可以作为合适的计算机可读存储介质(例如,cd
‑
rom、磁盘、闪存盘等)上存在的电子存储数据文件存在。在一些情况下,试剂盒中不存在真实的说明书,但提供了用于从远程来源(例如,经由互联网)获得说明书的方式。该实施例的实例是包括网址的试剂盒,在该网址中,可以观看该说明书和/或可以下载说明书。与说明书一样,用于获得说明书的方法可以记录在合适的基质上。
[0382]
另外的治疗方法
[0383]
可以使用工程化以靶向特定序列的定点多肽进行基因编辑。迄今为止,此类核酸酶有四种主要类型:大范围核酸酶及其功能等效物、锌指核酸酶(zfn)、转录激活因子样效应子核酸酶(talen)和crispr/cas核酸酶系统。核酸酶平台在设计难度、靶向密度和作用方式上不相同,尤其是zfn和talen的特异性是通过蛋白
‑
dna的相互作用,而rna
‑
dna的相互作用主要指导cas蛋白。cas9裂解还需要临近基序pam,该基序在不同的crispr系统之间有所不同。使用nrg pam裂解来自酿脓链球菌的cas9,而来自脑膜炎奈瑟球菌(neisseria meningitidis)的crispr可以在具有pam的位点裂解,这些位点包括nnnngatt(seq id no:312)、nnnnngttt(seq id no:313)和nnnngctt(seq id no:314)。许多其他cas9直系同源物靶向与替代性pam相邻的原间隔区。
[0384]
crispr内切核酸酶(诸如cas9)可用于本披露的方法的实施例中。此外,本文所述的传授内容(诸如治疗性靶位点)可以应用于其他形式的内切核酸酶,诸如zfn、talen、he、或megatal,或使用核酸酶的组合。然而,为了将本披露的传授内容应用于此类内切核酸酶,尤其需要工程化针对特定靶位点的蛋白。
[0385]
可以使附加结合结构域与cas9蛋白融合以增加特异性。这些构建体的靶位点映射到已鉴定的grna指定位点,但需要附加结合基序,诸如锌指结构域。在mega
‑
tal的情况下,大范围核酸酶可以与tale dna结合结构域融合。大范围核酸酶结构域可以增加特异性并提供裂解。类似地,失活或“死亡”的cas9(dcas9)可以与裂解结构域融合,并需要sgrna/cas9靶位点和融合dna结合结构域的相邻结合位点。除了催化失活之外,这可能还需要对dcas9进行一些蛋白工程化,以减少在没有附加结合位点情况下的结合。
[0386]
在一些实施例中,根据本披露的编辑基因组的组合物和方法(例如,将fviii编码序列插入白蛋白基因座中)使用以下以下任何方法。
[0387]
锌指核酸酶
[0388]
锌指核酸酶(zfn)是具有连接至ii型内切核酸酶foki的催化结构域的工程化锌指dna结合结构域的模块化蛋白。由于foki仅作为二聚体起作用,因此必须对一对zfn进行工程化以使其与相反dna链上的同源靶“半位点”序列结合,并且它们之间的精确间隔使得能够形成具有催化活性的foki二聚体。在本身没有序列特异性的foki结构域的二聚化之后,zfn半位点之间会产生dna双链断裂,作为基因组编辑的起始步骤。
[0389]
每个zfn的dna结合结构域通常具有3
‑
6个富cys2
‑
his2结构的锌指,每个指主要识别靶dna序列一条链上的核苷酸三联体,但是与第四核苷酸的跨链相互作用也可能很重要。与dna进行关键接触的位置中的指的氨基酸的改变会改变给定指的序列特异性。因此,四指锌指蛋白将选择性识别12bp的靶序列,其中该靶序列是每个指贡献的三联体偏好的合成物,但是三联体偏好可能会不同程度受到相邻指的影响。zfn的一个重要方面是,只需修饰单个指即可轻松将zfn重新靶向至几乎任何基因组地址,但是要做好这一点还需要相当多的专业知识。在zfn的大多数应用中,使用4
‑
6指的蛋白,分别识别12
‑
18bp。因此,一对zfn通常将识别24
‑
36bp的组合靶序列(不包括半位点之间5
‑
7bp的间隔区)。结合位点可以用更大的间隔区(包括15
‑
17bp)进一步隔开。假设在设计过程中排除了重复序列或基因同源物,则该长度的靶序列在人类基因组中可能是唯一的。然而,zfn蛋白
‑
dna相互作用的特异性并不是绝对的,因此脱靶结合和裂解事件的确会发生,要么是两个zfn之间的异二聚体,要么是zfn中的一个或另一个的同型二聚体。通过对foki结构域的二聚化界面进行工程化以产生“正”和“负”变体(也称为专性异二聚体变体,它们只可以与彼此二聚化,而不可以与自身二聚化),有效地消除了后者的可能性。促成专性异二聚体阻止了同二聚体的形成。这大大提高了zfn以及采用这些foki变体的任何其他核酸酶的特异性。
[0390]
在本领域中已经描述了多种基于zfn的系统,定期报告其修改,并且大量参考文献描述了用于指导zfn设计的规则和参数;参见例如segal等人,proc natl acad sci usa[美国国家科学院院刊](1999)96(6):2758
‑
63;b.dreier等人,j mol biol.[分子生物学杂志],(2000)303(4):489
‑
502;q.liu等人,j biol chem.[生物化学杂志](2002)277(6):3850
‑
6;dreier等人,j biol chem[生物化学杂志](2005)280(42):35588
‑
97;以及dreier等人,j biol chem.[生物化学杂志](2001)276(31):29466
‑
78。
[0391]
转录激活因子样效应子核酸酶(talen)
[0392]
talen代表模块化核酸酶的另一种形式,其中与zfn一样,经工程化的dna结合结构域与foki核酸酶结构域连接,并且一对talen串联作用以实现靶向dna裂解。与zfn的主要区别在于dna结合结构域的性质以及相关的靶dna序列识别特性。talen dna结合结构域源自tale蛋白,这些蛋白最初在植物细菌病原体黄单胞菌属(xanthomonas sp.)中有描述。tale具有33
‑
35个氨基酸重复序列的串联阵列,每个重复序列识别靶dna序列中的单个碱基对,该序列通常长达20bp,从而使总靶序列长度达到40bp。通过重复可变双残基(rvd)确定每个重复序列的核苷酸特异性,该重复可变双残基仅在位置12和13处包括两个氨基酸。鸟嘌呤、腺嘌呤、胞嘧啶和胸腺嘧啶碱基主要分别被以下四个rvd识别:asn
‑
asn、asn
‑
ile、his
‑
asp和asn
‑
gly。这构成了比锌指要简单得多的识别码,因此在核酸酶设计方面比锌指具有优势。然而,与zfn一样,talen的蛋白
‑
dna相互作用在其特异性上也不是绝对的,并且talen还受益于使用foki结构域的专性异二聚体变体来降低脱靶活性。
[0393]
已经产生了在其催化功能方面失活的foki结构域的另外的变体。如果talen或zfn对中的一半含有失活的foki结构域,则在靶位点处只会发生单链dna裂解(产生切口),而不会发生dsb。其结果与使用crispr/cas9/cpf1“切口酶”突变体(其中cas9裂解结构域之一已失活)相当。dna切口可以用于驱动通过hdr进行基因组编辑,但是效率要比dsb低。与dsb不同,主要益处在于脱靶切口被快速而准确地修复,而dsb容易受到nhej介导的错误修复。
[0394]
在本领域中已经描述了多种基于talen的系统,并且定期报告其修改:参见例如boch,science[科学](2009)326(5959):1509
‑
12;mak等人,science[科学](2012)335(6069):716
‑
9;以及moscou等人,science[科学](2009)326(5959):1501。已经有多个小组描述了基于“金门(golden gate)”平台或克隆方案的talen的用途:参见例如,t.cermak等人,nucleic acids res.[核酸研究](2011)39(12):e82;li等人,nucleic acids res.[核酸研究](2011)39(14):6315
‑
25;weber等人,plos one[公共科学图书馆
·
综合](2011)6(2):e16765;wang等人,j genet genomics[遗传学与基因组学杂志](2014)41(6):339
‑
47,epub 2014can 17;以及t.cermak等人,methods mol biol.[分子生物学方法](2015)1239:133
‑
59。
[0395]
归巢内切核酸酶
[0396]
归巢内切核酸酶(he)是具有长的识别序列(14
‑
44个碱基对)的位点特异性内切核酸酶,并通常在基因组中唯一的位点处以高特异性裂解dna。至少有六个按其结构分类的已知he家族,包括laglidadg(seq id no:6)、giy
‑
yig、his
‑
cis框、h
‑
n
‑
h、pd
‑
(d/e)xk和类vsr,它们源自多种宿主,包括真核生物、原生生物、细菌、古细菌、蓝细菌和噬菌体。与zfn和talen一样,he可以用于在靶基因座处产生dsb,作为基因组编辑的起始步骤。另外,一些天然的和经工程化的he仅切割dna的单链,从而作为位点特异性切口酶起作用。he的较大的靶序列以及he提供的特异性使它们成为产生位点特异性dsb的有吸引力的候选物。
[0397]
在本领域中已经描述了多种基于he的系统,并且定期报告其修改:参见例如以下文献的综述:steentoft等人,glycobiology[糖生物学](2014)24(8):663
‑
80;belfort和bonocora,methods mol biol.[分子生物学方法](2014)1123:1
‑
26;hafez和hausner,genome[基因组](2012)55(8):553
‑
69;以及其中引用的参考文献。
[0398]
megatal/tev
‑
mtalen/megatev
[0399]
作为杂合核酸酶的另外的实例,megatal平台和tev
‑
mtalen平台使用tale dna结合结构域和有催化活性的he的融合,该融合利用tale的可调dna结合和特异性,以及he的裂解序列特异性:参见例如boissel等人,nuc.acids res.[核酸研究](2014)42:2591
‑
601;kleinstiver等人,g3(2014)4:1155
‑
65;以及boissel和scharenberg,methods mol.biol.[分子生物学方法](2015)1239:171
‑
96。
[0400]
在另一种变型中,megatev架构是大范围核酸酶(mega)与衍生自giy
‑
yig归巢内切核酸酶i
‑
tevi(tev)的核酸酶结构域的融合。这两个活性位点在dna底物上相距~30bp,并产生两个具有不相容粘性末端的dsb;参见例如wolfs等人,nuc.acids res.[核酸研究](2014)42:8816
‑
29。可以预见,现有基于核酸酶的方法的其他组合将会发展,并可用于实现本文所述的靶向基因组修饰。
[0401]
dcas9
‑
foki或dcpf1
‑
fok1和其他核酸酶
[0402]
结合上述核酸酶平台的结构和功能特性提供了一种另外的基因组编辑的方法,该
方法可能克服一些固有缺陷。例如,crispr基因组编辑系统通常使用单个cas9内切核酸酶产生dsb。靶向的特异性由向导rna中的20或22个核苷酸的序列驱动,该序列与靶dna进行沃森
‑
克里克碱基配对(在来自酿脓链球菌的cas9的情况下,加上相邻的nag或ngg pam序列中的另外两个碱基)。这种序列足够长以至于在人类基因组中是唯一的,然而,rna/dna相互作用的特异性不是绝对的,有时可以耐受明显的混杂,尤其是在靶序列的5’一半处,这有效地减少了驱动特异性的碱基的数目。对此的一种解决方案是使cas9或cpf1催化功能完全失活(仅保留rna指导的dna结合功能),而将foki结构域与失活的cas9融合;参见例如tsai等人,nature biotech[自然
·
生物技术](2014)32:569
‑
76;以及guilinger等人,nature biotech.[自然
·
生物技术](2014)32:577
‑
82。由于foki必须二聚化才能变得有催化活性,因此需要两种指导rna来将两种foki融合物拴系在紧密接近的位置,以形成二聚体并裂解dna。这本质上使组合靶位点中的碱基数目加倍,从而提高被基于crispr的系统靶向的严格性。
[0403]
作为另一实例,tale dna结合结构域与具有催化活性的he(如i
‑
tevi)的融合利用tale的可调dna结合和特异性,以及i
‑
tevi的裂解序列特异性,预期可以进一步减少脱靶裂解。
[0404]
在下面的所附描述中阐述了本披露的一个或多个实施例的详情。从说明书看,本披露的其他特征、目的和优点将显而易见。在说明书中,除非上下文另外明确指出,否则单数形式也包括复数指示物。除非另外定义,否则本文使用的技术和科学术语均具有与本披露所属领域中的普通技术人员通常所理解的相同含义。在有冲突的情况下,以本说明书为准。
[0405]
应了解本文所述的实例和实施例仅出于说明的目的,并且将对本领域技术人员建议鉴于此的修改或变化且包括在本技术案的精神和范围内及所附权利要求的范围内。本文引用的所有出版物、专利和专利申请均据此通过援引整体并入以用于所有目的。
[0406]
通过以下非限制性实例进一步说明本文提供的披露的一些实施例。
[0407]
示例性实施例
[0408]
实施例1.一种系统,该系统包含:脱氧核糖核酸(dna)内切核酸酶或编码该dna内切核酸酶的核酸;包含与宿主细胞基因座互补的间隔区序列的向导rna(grna)或编码该grna的核酸;和包含编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
[0409]
实施例2.如实施例1所述的系统,其中该b结构域替代物包含0至6个n
‑
连接的糖基化位点。
[0410]
实施例3.如实施例2所述的系统,其中该b结构域替代物包含0至3个n
‑
连接的糖基化位点。
[0411]
实施例4.如实施例1所述的系统,其中该b结构域替代物包含seq id no:362
‑
369、371和373中任一个的氨基酸序列。
[0412]
实施例5.如实施例4所述的系统,其中该b结构域替代物包含seq id no:362
‑
366、371和373中任一个的氨基酸序列或其与seq id no:362
‑
366、371和373中任一个具有至少80%同一性的变体。
[0413]
实施例6.如实施例5所述的系统,其中该b结构域替代物包含seq id no:362
‑
364、371和373中任一个的氨基酸序列。
[0414]
实施例7.如实施例1
‑
6中任一项所述的系统,其中该宿主细胞基因座是在肝脏中表达的基因的基因座。
[0415]
实施例8.如实施例1
‑
7中任一项所述的系统,其中该宿主细胞基因座是编码急性期蛋白的基因的基因座。
[0416]
实施例9.如实施例8所述的系统,其中该急性期蛋白是白蛋白、转铁蛋白或纤维蛋白原。
[0417]
实施例10.如实施例1
‑
7中任一项所述的系统,其中该宿主细胞基因座是安全港基因座。
[0418]
实施例11.如实施例1
‑
10中任一项所述的系统,其中该dna内切核酸酶选自由以下项组成的组:cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4或cpf1内切核酸酶及其功能衍生物。
[0419]
实施例12.如实施例11所述的系统,其中该dna内切核酸酶是cas9。
[0420]
实施例13.如实施例1
‑
11中任一项所述的系统,其中对编码该dna内切核酸酶的核酸进行密码子优化以在宿主细胞中表达。
[0421]
实施例14.如实施例1
‑
13中任一项所述的系统,其中编码该dna内切核酸酶的核酸是脱氧核糖核酸(dna)。
[0422]
实施例15.如实施例1
‑
13中任一项所述的系统,其中编码该dna内切核酸酶的核酸是核糖核酸(rna)。
[0423]
实施例16.如实施例15所述的系统,其中编码该dna内切核酸酶的rna是mrna。
[0424]
实施例17.如实施例1
‑
16中任一项所述的系统,其中该供体模板核酸序列经过密码子优化以在该宿主细胞中表达。
[0425]
实施例18.如实施例1
‑
17中任一项所述的系统,其中与编码fviii蛋白的野生型核酸序列相比,该供体模板核酸序列包含降低含量的cpg二核苷酸。
[0426]
实施例19.如实施例18所述的系统,其中该供体模板核酸序列不含cpg二核苷酸。
[0427]
实施例20.如实施例1
‑
19中任一项所述的系统,其中该供体模板在aav载体中编码。
[0428]
实施例21.如实施例1
‑
20中任一项所述的系统,其中该供体模板包含供体盒,该供体盒包含编码合成fviii蛋白的核酸序列,并且其中该供体盒在一侧或两侧上侧接有grna靶位点。
[0429]
实施例22.如实施例21所述的系统,其中该供体盒在两侧上侧接有grna靶位点。
[0430]
实施例23.如实施例21所述的系统,其中该供体盒在其5’侧上侧接有grna靶位点。
[0431]
实施例24.如实施例21
‑
23中任一项所述的系统,其中该grna靶位点是该系统中grna的靶位点。
[0432]
实施例25.如实施例24所述的系统,其中该供体模板的grna靶位点是该系统中grna的基因组grna靶位点的反向互补序列。
[0433]
实施例26.如实施例1
‑
25中任一项所述的系统,其中该dna内切核酸酶或编码该dna内切核酸酶的核酸包含在脂质体或脂质纳米颗粒中。
[0434]
实施例27.如实施例26所述的系统,其中该脂质体或脂质纳米颗粒还包含该grna。
[0435]
实施例28.如实施例1
‑
27中任一项所述的系统,其中该dna内切核酸酶与该grna复合,从而提供核糖核蛋白(rnp)复合物。
[0436]
实施例29.一种编辑宿主细胞中的基因组的方法,该方法包括向该细胞提供以下物质:(a)包含与宿主细胞基因座互补的间隔区序列的grna或编码该grna的核酸;(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
[0437]
实施例30.如实施例29所述的方法,其中该b结构域替代物包含0至6个n
‑
连接的糖基化位点。
[0438]
实施例31.如实施例30所述的方法,其中该b结构域替代物包含0至3个n
‑
连接的糖基化位点。
[0439]
实施例32.如实施例29所述的方法,其中该b结构域替代物包含seq id no:362
‑
369、371和373中任一个的氨基酸序列。
[0440]
实施例33.如实施例32所述的方法,其中该b结构域替代物包含seq id no:362
‑
366、371和373中任一个的氨基酸序列或其与seq id no:362
‑
366、371和373中任一个具有至少80%同一性的变体。
[0441]
实施例34.如实施例33所述的方法,其中该b结构域替代物包含seq id no:362
‑
364、371和373中任一个的氨基酸序列。
[0442]
实施例35.如实施例29
‑
34中任一项所述的方法,其中该宿主细胞内源基因座是在肝脏中表达的基因的基因座。
[0443]
实施例36.如实施例29
‑
35中任一项所述的方法,其中该宿主细胞内源基因座是编码急性期蛋白的基因的基因座。
[0444]
实施例37.如实施例36所述的方法,其中该急性期蛋白是白蛋白、转铁蛋白或纤维蛋白原。
[0445]
实施例38.如实施例29
‑
34中任一项所述的方法,其中该宿主细胞内源基因座是安全港基因座。
[0446]
实施例39.如实施例29
‑
38中任一项所述的方法,其中该dna内切核酸酶选自由以下项组成的组:cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4或cpf1内切核酸酶;或其功能衍生物。
[0447]
实施例40.如实施例39所述的方法,其中该dna内切核酸酶是cas9。
[0448]
实施例41.如实施例29
‑
40中任一项所述的方法,其中对编码该dna内切核酸酶的核酸进行密码子优化以在宿主细胞中表达。
[0449]
实施例42.如实施例29
‑
41中任一项所述的方法,其中编码该dna内切核酸酶的核酸是脱氧核糖核酸(dna)。
[0450]
实施例43.如实施例29
‑
41中任一项所述的方法,其中编码该dna内切核酸酶的核酸是核糖核酸(rna)。
[0451]
实施例44.如实施例43所述的方法,其中编码该dna内切核酸酶的rna是mrna。
[0452]
实施例45.如实施例29所述的方法,其中该供体模板在aav载体中编码。
[0453]
实施例46.如实施例29
‑
45中任一项所述的方法,其中该供体模板核酸序列经过密码子优化以在该宿主细胞中表达。
[0454]
实施例47.如实施例29
‑
46中任一项所述的方法,其中与编码fviii的野生型核酸序列相比,该供体模板核酸序列包含降低含量的cpg二核苷酸。
[0455]
实施例48.如实施例47所述的方法,其中该供体模板核酸序列不含cpg二核苷酸。
[0456]
实施例49.如实施例29
‑
48中任一项所述的方法,其中该供体模板包含供体盒,该供体盒包含编码合成fviii蛋白的核酸序列,并且其中该供体盒在一侧或两侧上侧接有grna靶位点。
[0457]
实施例50.如实施例49所述的方法,其中该供体盒在两侧上侧接有grna靶位点。
[0458]
实施例51.如实施例49所述的方法,其中该供体盒在其5’侧上侧接有grna靶位点。
[0459]
实施例52.如实施例49
‑
51中任一项所述的方法,其中该grna靶位点是所施用的grna的靶位点。
[0460]
实施例53.如实施例52所述的方法,其中该供体模板的grna靶位点是该细胞基因组中针对所施用的grna的grna靶位点的反向互补序列。
[0461]
实施例54.如实施例29
‑
53中任一项所述的方法,其中该dna内切核酸酶或编码该dna内切核酸酶的核酸配制在脂质体或脂质纳米颗粒中。
[0462]
实施例55.如实施例54所述的方法,其中该脂质体或脂质纳米颗粒还包含该grna。
[0463]
实施例56.如实施例29
‑
55中任一项所述的方法,其中该dna内切核酸酶和该grna作为核糖核蛋白(rnp)复合物提供给宿主细胞,该复合物包含与该grna预复合的dna内切核酸酶。
[0464]
实施例57.如实施例29
‑
56中任一项所述的方法,其中在将该供体模板提供给该细胞后超过4天,将该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。
[0465]
实施例58.如实施例29
‑
57中任一项所述的方法,其中在将该供体模板提供给该细胞后至少14天,将该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。
[0466]
实施例59.如实施例57或58所述的方法,其中在第一剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸。
[0467]
实施例60.如实施例59所述的方法,其中在第一剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸,直至达到该编码合成fviii蛋白的核酸序列的目标靶向整合水平,或达到该编码合成fviii蛋白的核酸序列的目标表达水平。
[0468]
实施例61.如实施例29
‑
60中任一项所述的方法,其中该细胞是肝脏细胞。
[0469]
实施例62.如实施例61所述的方法,其中该细胞是人肝脏细胞或人肝窦上皮细胞。
[0470]
实施例63.一种细胞,其中该细胞的基因组包含编码合成fviii蛋白的dna,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
[0471]
实施例64.如实施例63所述的细胞,其中该合成fviii蛋白与内源性白蛋白启动子、内源性转铁蛋白启动子或内源性纤维蛋白原α启动子可操作地连接。
[0472]
实施例65.如实施例63所述的细胞,其中编码该合成fviii蛋白的核酸序列经过密码子优化以在该细胞中表达。
[0473]
实施例66.如实施例63所述的细胞,其中该细胞是人肝脏细胞。
[0474]
实施例67.如实施例66所述的细胞,其中该细胞是人肝脏细胞或人肝窦上皮细胞。
[0475]
实施例68.如实施例67所述的细胞,其中该细胞通过如实施例29
‑
62中任一项所述的方法制备。
[0476]
实施例69.一种治疗受试者的血友病a的方法,该方法包括:向该受试者中的细胞提供以下物质:(a)包含与宿主细胞基因座互补的间隔区序列的grna或编码该grna的核酸;(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
[0477]
实施例70.如实施例69所述的方法,其中该b结构域替代物包含0至6个n
‑
连接的糖基化位点。
[0478]
实施例71.如实施例70所述的方法,其中该b结构域替代物包含0至3个n
‑
连接的糖基化位点。
[0479]
实施例72.如实施例29所述的方法,其中该b结构域替代物包含seq id no:362
‑
369、371和373中任一个的氨基酸序列。
[0480]
实施例73.如实施例72所述的方法,其中该b结构域替代物包含seq id no:362
‑
366、371和373中任一个的氨基酸序列或其与seq id no:362
‑
366、371和373中任一个具有至少80%同一性的变体。
[0481]
实施例74.如实施例73所述的方法,其中该b结构域替代物包含seq id no:362
‑
364、371和373中任一个的氨基酸序列。
[0482]
实施例75.如实施例69
‑
74中任一项所述的方法,其中该宿主细胞基因座是在肝脏中表达的基因的基因座。
[0483]
实施例76.如实施例69
‑
75中任一项所述的方法,其中该宿主细胞基因座是编码急性期蛋白的基因的基因座。
[0484]
实施例77.如实施例76所述的方法,其中该急性期蛋白是白蛋白、转铁蛋白或纤维蛋白原。
[0485]
实施例78.如实施例69
‑
74中任一项所述的方法,其中该宿主细胞基因座是安全港基因座。
[0486]
实施例79.如实施例69
‑
78中任一项所述的方法,其中该dna内切核酸酶选自由以下项组成的组:cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas100、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、
csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4和cpf1内切核酸酶;或其功能衍生物。
[0487]
实施例80.如实施例79所述的方法,其中该dna内切核酸酶是cas9。
[0488]
实施例81.如实施例80所述的方法,其中该cas9是spcas9或slucas9。
[0489]
实施例82.如实施例69
‑
81中任一项所述的方法,其中对编码该dna内切核酸酶的核酸进行密码子优化以在细胞中表达。
[0490]
实施例83.如实施例69
‑
82中任一项所述的方法,其中编码该dna内切核酸酶的核酸是脱氧核糖核酸(dna)。
[0491]
实施例84.如实施例69
‑
82中任一项所述的方法,其中编码该dna内切核酸酶的核酸是核糖核酸(rna)。
[0492]
实施例85.如实施例84所述的方法,其中编码该dna内切核酸酶的rna是mrna。
[0493]
实施例86.如实施例69
‑
85中任一项所述的方法,其中将该grna或编码该grna的核酸、该dna内切核酸酶或编码该dna内切核酸酶的核酸以及该供体模板中的一种或多种配制在脂质体或纳米颗粒中。
[0494]
实施例87.如实施例69
‑
86中任一项所述的方法,其中该供体模板在aav载体中编码。
[0495]
实施例88.如实施例69
‑
87中任一项所述的方法,其中该供体模板核酸序列经过密码子优化以在该宿主细胞中表达。
[0496]
实施例89.如实施例69
‑
88中任一项所述的方法,其中与编码fviii的野生型核酸序列相比,该供体模板核酸序列包含降低含量的cpg二核苷酸。
[0497]
实施例90.如实施例89所述的方法,其中该供体模板核酸序列不含cpg二核苷酸。
[0498]
实施例91.如实施例69
‑
90中任一项所述的方法,其中该供体模板包含供体盒,该供体盒包含编码合成fviii蛋白的核酸序列,并且其中该供体盒在一侧或两侧上侧接有grna靶位点。
[0499]
实施例92.如实施例91所述的方法,其中该供体盒在两侧上侧接有grna靶位点。
[0500]
实施例93.如实施例91所述的方法,其中该供体盒在其5’侧上侧接有grna靶位点。
[0501]
实施例94.如实施例91
‑
93中任一项所述的方法,其中该grna靶位点是该grna的靶位点。
[0502]
实施例95.如实施例94所述的方法,其中该供体模板的grna靶位点是该细胞基因组中针对该grna的grna靶位点的反向互补序列。
[0503]
实施例96.如实施例69
‑
95中任一项所述的方法,其中向该细胞提供该供体模板包括向该受试者静脉施用该供体模板。
[0504]
实施例97.如实施例69
‑
96中任一项所述的方法,其中该dna内切核酸酶或编码该dna内切核酸酶的核酸配制在脂质体或脂质纳米颗粒中。
[0505]
实施例98.如实施例97所述的方法,其中该脂质体或脂质纳米颗粒还包含该grna。
[0506]
实施例99.如实施例98所述的方法,其中向该细胞提供该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸包括向该受试者静脉内施用该脂质体或脂质纳米颗粒。
[0507]
实施例100.如实施例69
‑
99中任一项所述的方法,其中该dna内切核酸酶和该grna
作为核糖核蛋白(rnp)复合物提供给该宿主细胞,该rnp复合物包含与该grna复合的dna内切核酸酶。
[0508]
实施例101.如实施例69
‑
100中任一项所述的方法,其中在将该供体模板提供给该细胞后超过4天,将该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。
[0509]
实施例102.如实施例69
‑
101中任一项所述的方法,其中在将该供体模板提供给该细胞后至少14天,将该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸提供给该细胞。
[0510]
实施例103.如实施例101或102所述的方法,其中在第一剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸。
[0511]
实施例104.如实施例103所述的方法,其中在第一剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸后,向该细胞提供一个或多个附加剂量的该grna或编码该grna的核酸和该dna内切核酸酶或编码该dna内切核酸酶的核酸,直至达到该编码合成fviii蛋白的核酸序列的目标靶向整合水平和/或该编码合成fviii蛋白的核酸序列的目标表达水平。
[0512]
实施例105.如实施例101
‑
104中任一项所述的方法,其中向该细胞提供该grna和该dna内切核酸酶或编码该dna内切核酸酶的核酸包括向该受试者施用包含编码该dna内切核酸酶的核酸和该grna的脂质纳米颗粒。
[0513]
实施例106.如实施例101
‑
105中任一项所述的方法,其中向该细胞提供该供体模板包括向该受试者施用在aav载体中编码的该供体模板。
[0514]
实施例107.如实施例69
‑
106中任一项所述的方法,其中该细胞是肝细胞。
[0515]
实施例108.如实施例69
‑
107中任一项所述的方法,其中编码合成fviii蛋白的核酸序列在该受试者的肝脏中表达。
[0516]
实施例109.一种治疗受试者的血友病a的方法,该方法包括向该受试者施用如实施例63
‑
68中任一项所述的细胞。
[0517]
实施例110.如实施例109所述的方法,其中该细胞对该受试者来说是自体的。
[0518]
实施例111.如实施例110所述的方法,该方法进一步包括从该受试者获得生物样品,其中该生物样品包含肝脏细胞,并且其中该细胞由该肝脏细胞制备。
[0519]
实施例112.一种试剂盒,该试剂盒包含如实施例1
‑
28中任一项所述的系统的一种或多种要素,进一步包含使用说明书。
[0520]
实施例113.一种包含编码合成fviii蛋白的多核苷酸序列的核酸,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
[0521]
实施例114.如实施例113所述的核酸,其中该b结构域替代物包含0至6个n
‑
连接的糖基化位点。
[0522]
实施例115.如实施例113所述的核酸,其中该b结构域替代物包含0至3个n
‑
连接的糖基化位点。
[0523]
实施例116.如实施例113所述的核酸,其中该b结构域替代物包含seq id no:362
‑
369、371和373中任一个的氨基酸序列。
[0524]
实施例117.如实施例116所述的核酸,其中该b结构域替代物包含seq id no:362
‑
364、371和373中任一个的氨基酸序列或其与seq id no:362
‑
364、371和373中任一个具有至少80%同一性的变体。
[0525]
实施例118.如实施例116所述的核酸,其中该b结构域替代物包含seq id no:362
‑
363、371和373中任一个的氨基酸序列。
[0526]
实施例119.如实施例113
‑
118中任一项所述的核酸,其中编码合成fviii蛋白的该多核苷酸序列经过密码子优化以在宿主细胞中表达。
[0527]
实施例120.如实施例113
‑
119中任一项所述的核酸,其中与编码fviii的野生型核酸序列相比,编码合成fviii蛋白的该多核苷酸序列包含降低含量的cpg二核苷酸。
[0528]
实施例121.如实施例120所述的核酸,其中编码合成fviii蛋白的该多核苷酸序列不含cpg二核苷酸。
[0529]
实施例122.如实施例113
‑
121中任一项所述的核酸,其中该核酸是病毒载体。
[0530]
实施例123.如实施例122所述的核酸,其中该病毒载体是aav载体。
[0531]
实施例124.一种增加受试者中的fviii量的方法,该方法包括:向该受试者中的细胞提供以下物质,其中该受试者具有第一fviii血清水平:(a)包含与宿主细胞基因座互补的间隔区序列的grna或编码该grna的核酸;(b)dna内切核酸酶或编码该dna内切核酸酶的核酸;以及(c)包含编码合成fviii蛋白的核酸序列的供体模板,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度为3至约40个氨基酸。
[0532]
实施例125.如实施例124所述的方法,其中第一fviii血清水平低于约0.40iu/ml。
[0533]
实施例126.如实施例125所述的方法,其中第一fviii血清水平低于约0.05iu/ml。
[0534]
实施例127.如实施例125所述的方法,其中第一fviii血清水平低于约0.01iu/ml。
[0535]
实施例128.如实施例1
‑
28中任一项所述的系统用于治疗血友病a的用途。
[0536]
实施例129.如实施例1
‑
28中任一项所述的系统用于制造用于治疗血友病a的药物的用途。
[0537]
实施例130.如实施例63
‑
68中任一项所述的细胞用于治疗血友病a的用途。
[0538]
实施例131.如实施例63
‑
68中任一项所述的细胞用于制造用于治疗血友病a的药物的用途。
[0539]
实施例132.如实施例112所述的试剂盒用于治疗血友病a的用途。
[0540]
实施例133.如实施例112所述的试剂盒用于制造用于治疗血友病a的药物的用途。
[0541]
实施例134.如实施例113
‑
123中任一项所述的核酸用于治疗血友病a的用途。
[0542]
实施例135.如实施例113
‑
123中任一项所述的核酸用于制造用于治疗血友病a的药物的用途。
[0543]
实施例136.一种合成fviii蛋白,其中该合成fviii蛋白包含b结构域替代物,其中该b结构域替代物包含0至9个n
‑
连接的糖基化位点且长度不超过约40个氨基酸。
[0544]
实例
[0545]
实例1:含有n
‑
糖基化基序的氨基酸序列在crispr/cas9裂解介导的靶向整合到小
鼠白蛋白内含子1中之后改善fviii的表达
[0546]
构建体设计
[0547]
将编码fviii的核酸序列插入基因组中的挑战在于:天然fviii编码序列为7053bp,这尤其使得难以将其包装在腺相关病毒中(对于作为模板体内递送以整合在由序列特异性核酸酶诸如cas9产生的双链断裂处,aav具有4800至5000bp范围内的包装限值)。为了解决该问题,申请人设计了一组具有改变的b结构域的fviii编码序列。尽管fviii的b结构域不是功能所需的,但它会改善fviii的分泌。将这些fviii编码序列设计为表达具有短b结构域(替代b结构域)的合成fviii。为了评价具有替代b结构域的合成fviii编码序列在整合到基因组中后产生和分泌fviii蛋白的能力,将构建体设计为将fviii编码序列靶向整合到小鼠白蛋白基因的内含子1中。白蛋白基因座提供了在肝脏细胞中有活性的强启动子,使得在该基因座处插入的合适的fviii编码序列当与白蛋白启动子可操作地连接时可以表达。
[0548]
使用已知的分子生物学技术构建本文称为pcb076(seq id no:316)、pcb100(seq id no:320)、pcb1003(seq id no:324)、pcb085(seq id no:3319)或pcb080(seq id no:318)的一系列质粒。相同的基于puc19的细菌质粒骨架(含有细菌复制起点和卡那霉素抗性基因)用于所有5种质粒。用以下元件构建质粒(按顺序):grna靶位点(对于grna malbt1,seq id no:338,靶向小鼠白蛋白基因的外显子1)|18bp间隔区|剪接受体位点(“sa”)|fviii编码序列|多聚腺苷酸化信号(“spa”)。质粒的不同之处仅在于人fviii编码序列的密码子优化,以及编码b结构域替代物的序列的存在(pcb076)或不存在(pcb100、pcb1003、pcb085和pcb080)。本实例中使用的b结构域替代物由来自人fviii b结构域n端的前6个n
‑
糖基化基序组成。
[0549]
质粒pcb100、pcb1003、pcb085和pcb080全部含有b结构域缺失的人fviii的编码序列,其中b结构域被“sq接头”(其编码氨基酸sfsqnppvlkrhqr,seq id no:337)替代。sq接头包括蛋白酶裂解位点(rhqr),但缺少n
‑
连接的糖基化位点。质粒pcb076(seq id no:316)含有与pcb100相同的密码子优化的b结构域缺失的人fviii编码序列(“co1”,参见下面的实例4),和编码17个氨基酸的附加dna序列,与插入sq接头中代替b结构域(从而形成b结构域替代物)的人fviii b结构域n端的前6个n
‑
糖基化基序相对应。其他质粒具有以下密码子优化:pcb100
‑
co1(seq id no:320)、pcb1003
‑
co2(seq id no:324)、pcb085
‑
co3(seq id no:319)和pcb080
‑
co4(seq id no:318)(参见下面的实例4)。将质粒设计为用于靶向整合到双链断裂中的供体,该双链断裂是使用crispr/cas9系统,利用grna malbt1(tgccagttcccgatcgttac,seq id 338)在小鼠白蛋白基因的内含子1中产生的。肝脏是这种靶向整合的靶器官,特别是肝细胞。体内肝细胞大部分是静止的,并且已知修复非分裂细胞中dna双链断裂的主要细胞机制是非同源末端连接(nhej)(z.mao等人,cell cycle[细胞周期](2008)7:2902
‑
06)。在基因组中存在线性双链dna分子(供体)和双链断裂的情况下,供体dna可通过nhej机制插入双链断裂处。
[0550]
替代性地,基因组中双链断裂的末端可通过相同的nhej机制彼此重新连接,这是通常比插入供体模板更频繁的事件。nhej修复是易错过程,这导致在双链断裂位点引入插入或缺失。在细胞基因组的双链断裂处作为质粒递送的供体模板的靶向整合可以通过在供体质粒中包括核酸酶的切割位点来增强。因为质粒是环状分子,它们不是在双链断裂处整
合的模板。在质粒中包含单个向导rna切割位点导致质粒在cas9/grna复合物的存在下线性化。因此,在小鼠基因组中存在的序列的反向互补序列中的fviii盒的5’末端插入malbt1向导的单个向导rna切割位点。
[0551]
当使用位于盒侧该翼的两个向导位点时,在基因组中使用向导序列的反向互补序列理论上有利于正向整合。然而,当仅使用一个向导切割位点时,不可能保持该优点。包含在编码序列侧翼的向导切割位点产生由编码序列盒和细菌质粒骨架(编码抗生素抗性基因和复制起点)组成的两个线性片段,在这种情况下,细菌骨架片段竞争在基因组中的双链断裂处整合。由于这个原因,申请人将质粒设计成使用单个向导切割位点。合成fviii编码序列盒从5’末端开始依次由以下元件组成;malbt1 grna靶位点、18bp间隔区序列、剪接受体序列(actaaagaattattcttttacatttcag,seq id no:307)、其中信号肽被二核苷酸tg替代的b结构域缺失的人fviii编码序列和多聚腺苷酸化信号(aataaaagatctttattttcattagatctgtgtgttggttttttgtgtg,seq id no:306)。
[0552]
将构建体设计成使得在整合到白蛋白的内含子1中之后,产生杂交前mrna,其含有白蛋白的外显子1、白蛋白的内含子1的一部分和fviii编码序列盒。在整合到白蛋白内含子1中后,预期细胞的剪接机制以一定频率剪接出内含子1,从而产生成熟mrna,其中白蛋白外显子1与成熟fviii的编码序列框内融合。tg二核苷酸包含在构建体中以维持翻译阅读框。预计该mrna的翻译会产生一种蛋白,其中白蛋白的信号肽和前肽与fviii的成熟编码序列融合。当通过细胞的分泌机制时,预计信号肽和前肽被裂解掉,留下三个添加到成熟fviii的天然n端的氨基酸(glu
‑
ala
‑
leu)。使用该方法产生的fviii蛋白在小鼠中具有活性,尽管存在这些额外的三个氨基酸。
[0553]
grna
[0554]
这些实验中使用的grna是化学合成的,并入了经化学修饰的核苷酸以提高对核酸酶的抗性。一个实例中的grna由以下结构组成:5’usgscscaguucccgaucguuacguuuuagagcuagaaauagcaaguuaaa auaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcusususu
‑3’
(seq id no:339),其中“a、g、u、c”是天然rna核苷酸,“a、g、u、c”是2
’‑
o
‑
甲基核苷酸,并且“s”是硫代磷酸酯骨架。grna的小鼠白蛋白靶向序列带有下划线,grna序列的其余部分是共同的支架序列。
[0555]
mrna
[0556]
可以通过本领域已知的方法产生该mrna。本文所用的一种这样的方法是使用t7聚合酶进行体外转录,其中mrna的序列在含有t7聚合酶启动子的质粒中编码。简言之,将质粒在含有t7聚合酶和核糖核苷酸的适当缓冲液中孵育后,产生了编码所需蛋白的氨基酸序列的rna分子。天然核糖核苷酸或经化学修饰的核糖核苷酸可用于反应混合物中以产生具有天然mrna的天然化学结构或具有经修饰的化学结构的mrna分子。在本文所述的研究中,使用天然(未修饰的)核糖核苷酸。另外,在转录反应中包括加帽组分,使得mrna的5’末端加帽。
[0557]
将spcas9 mrna设计为编码与核定位结构域(nls)融合的spcas9蛋白,核定位结构域是将spcas9蛋白转运到可发生基因组dna裂解的核区室中所必需的。cas9 mrna另外的组分是在5’末端第一个促进核糖体结合的密码子之前的kozak序列,以及在3’末端由一系列a残基组成的聚a尾。具有nls序列的spcas9 mrna实例示于seq id no:340中。另外,通过利用
每个氨基酸最常用的密码子,针对密码子使用对spcas9编码序列的序列进行优化。另外,为了促进mrna有效翻译为spcas9蛋白,优化编码序列以去除隐蔽核糖体结合位点和上游开放阅读框。
[0558]
lnp
[0559]
这些研究中使用的lnp的主要组分是脂质c12
‑
200(love等人,2010同上)。c12
‑
200与带负电荷的rna分子形成复合物。一般而言,将c12
‑
200与1,2
‑
二油酰基
‑
sn
‑
甘油
‑3‑
磷酸乙醇胺(dope)、dmpe
‑
mpeg2000和胆固醇组合。当在受控条件下,例如在设备(卑诗省温哥华市精密纳米系统公司(precision nanosystems,vancouver,bc))中与核酸诸如grna和mrna混合时,发生lnp自组装,其中核酸被包封在lnp内部。为了将grna和cas9 mrna组装在lnp中,视情况而定将乙醇和脂质原液吸取到玻璃小瓶中。示例性比率由摩尔比为50:10:38.5:1.5的c12
‑
200、dope、胆固醇和mpeg2000
‑
dmg组成。在无rna酶的试管中,将grna和mrna稀释于100mm柠檬酸钠(ph 3.0)和300mm nacl中。将筒柱(精密纳米系统公司(precision nanosystems))在脂质侧用乙醇洗涤,在rna侧用水洗涤。将脂质工作原液抽入注射器中,从注射器中去除空气,然后将注射器插入筒柱中。使用相同的程序将grna和cas9 mrna的混合物装入注射器。然后在制造商推荐的条件下进行运行。使用10k截留分子量(mwco)透析筒柱将lnp悬浮液在4升pbs中透析4小时,然后通过100k mwco离心筒柱(艾米康(amicon))离心浓缩,包括在离心过程中在pbs中洗涤三次。最后,将lnp悬浮液通过0.2μm针头式过滤器进行无菌过滤。使用市售内毒素试剂盒(鲎变形细胞溶解物(lal)测定法)检查内毒素水平,并通过动态光散射测定粒度分布。
[0560]
使用测定法(赛默飞世尔(thermo fisher))测定包封的rna的浓度。替代性地,将grna和cas9 mrna单独配制到lnp中,然后混合在一起,之后处理培养中的细胞或注入动物体内。使用单独配制的grna和cas9 mrna,可以测试grna和cas9 mrna的特定比率。
[0561]
利用替代性阳离子脂质分子的替代性lnp配制品也用于体内递送grna和cas9 mrna。
[0562]
构建体的体内测试
[0563]
使用鼠类模型测试设计的构建体产生fviii的能力。血友病a的小鼠模型是本领域已知的(例如,l.bi等人,nat genet.[自然
·
遗传学](1995)10:119
‑
21,doi:10.1038/ng0595
‑
119)。使用qiagen 质粒大提试剂盒(目录号12362)纯化质粒pcb076、pcb100、pcb1003、pcb085和pcb080,然后在0.9%盐水中稀释到15μg/ml的最终浓度。血友病a小鼠(品系b6;129s
‑
f8
tm1kaz
/j),一种缺乏小鼠fviii蛋白的小鼠品系,获自杰克逊实验室(缅因州巴尔港(bar harbor,me))。通过流体动力学注射(“hdi”),在5至6秒的时间内,经由尾静脉向血友病a小鼠队列每只小鼠注射2ml经稀释的质粒dna。已有报道,hdi过程导致将质粒dna递送到肝脏细胞(包括肝细胞)的细胞核中(参见例如,f.niola等人,meth mol biol[分子生物学方法](2019)1961:329
‑
41)。注射后一天,给予小鼠眶后(“ro”)注射包封spcas9 mrna和向导rnamalbt1的lnp配制品。施用给小鼠的lnp剂量为1mg/kg体重的spcas9 mrna加上1mg/kg体重的grna。
[0564]
三天后处死一组lnp单独给药的小鼠,从整个肝脏中提取dna并使用tide分析
(e.k.brinkman等人,nuc acid res[核酸研究](2014)42:e168)测定malbt1 grna的预期切割位点处的插入缺失(indel)。在tide分析中,通过pcr从经过处理的细胞的基因组dna扩增预期crispr/cas9切割位点的基因组区域,然后进行sanger测序。使用tide软件程序分析测序色谱图,确定预测的切割位点周围区域中的插入和缺失的频率。
[0565]
在这些实验中,确定中靶位点处插入缺失的频率为25.4%。用lnp对注射了质粒的小鼠给药六天后,通过ro放血取血样到柠檬酸钠中(柠檬酸钠与血液的比率为1:9),并且通过离心收集血浆。使用fviii活性测定(diapharma,chromogenix sp因子fviii,目录号k824086)测量血浆中的fviii活性。使用(拜耳(bayer))(一种重组人fviii)作为标准品,并且将血液中每ml的fviii活性单位转化为正常活性的百分比(1u/ml=100%)。结果汇总在图1中。注射含有6个n
‑
聚糖b结构域替代序列代替b结构域的质粒pcb076的小鼠的平均合成fviii水平相当于正常人fviii水平的20%。相反,注射pcb100质粒的小鼠在其血液中没有可检测的fviii水平,pcb100质粒除了不存在6个n
‑
聚糖b结构域替代序列外与pcb076相同。当与未经基因编辑的(原初)血友病a小鼠相比时,注射了质粒pcb1003、pcb085或pcb080的小鼠在其血液中具有低的或不可测量的fviii活性,这些质粒含有缺乏6个n
‑
聚糖b结构域替代序列的不同密码子优化的b结构域缺失的fviii编码序列。注射了pcb1003和pcb080的一些小鼠在其血液中具有可检测的fviii,在正常值的1%
‑
3%的范围内,表明密码子优化co2(pcb1003)和co4(pcb080)可能比密码子优化co1(pcb100)和co3(pcb085)活性更高。
[0566]
本研究中小鼠血液中产生的fviii水平取决于正向(能够产生fviii蛋白的方向)靶向整合到白蛋白内含子1中的频率和fviii编码序列的固有表达效率。fviii编码序列的固有表达效率是转录效率、翻译效率(其随采用的密码子优化的类型而变化)和分泌过程的效率的函数。在fviii蛋白的情况下,已经报道蛋白的分泌可以是限速步骤,并且与当fviii在细胞中以高水平表达时可诱导的未折叠蛋白反应相关。(m.swaroop等人,j biol chem[生物化学杂志](1997)272:24121
‑
24;r.j.kaufman,blood[血液](2009)114:sci
‑
19)。
[0567]
为了区分靶向整合频率(由于hdi或其他因素在供体递送效率方面的可变性而在小鼠之间可能有所不同)与合成fviii编码序列的固有表达效率,使用微滴式数字pcr(dd
‑
pcr)对靶向整合频率进行定量。dd
‑
pcr是一种用于定量样品中核酸序列的绝对拷贝数的方法。为了仅定量插入白蛋白内含子1中的合成fviii编码序列盒的正向,设计一对pcr引物,其中正向引物位于白蛋白内含子1中grnamalbt1切割位点的位点5’,而反向引物位于fviii编码序列的5’末端。设计与两个引物之间的序列互补的荧光探针。针对远离malbt1 grna位点的位点处的天然小鼠白蛋白基因序列,设计参考引物/探针组。使用参考引物探针归一化每次测定中输入小鼠基因组dna的量。
[0568]
为了进行该分析,在将小鼠用lnp给药后8天处死来自上述实验的小鼠。将整个肝脏匀化,并使用qiagen 组织试剂盒纯化总基因组dna。然后使用上述dd
‑
pcr测定法测定相等质量基因组dna的靶向整合频率。每只小鼠的结果汇总在表2中。正向的靶向整合频率范围为0.09%至0.95%(每100个单倍体基因组0.09至0.95个拷贝)。血液中的峰值fviii水平与整合频率正相关,表明fviii的水平取决于整合到白蛋白内含子1中的fviii盒的拷贝数。在注射pcb076的小鼠中的平均靶向整合频率是0.47
±
0.26,相比而言在注射
pcb100的小鼠中为0.28
±
0.15,表明在注射含有b结构域替代物代替sq接头的pcb076的小鼠中有整合频率更高的趋势,但是这种差异在统计上不显著。
[0569]
表2:与血液中的峰值fviii水平相比小鼠肝脏中的靶向整合频率
[0570][0571]
将每只小鼠血液中的fviii水平归一化为整合频率提供了fviii编码序列的固有表达效率的量度。fviii水平除以靶向整合频率得到的比率的平均值对于pcb076是42且对于pcb100是5.3,并且如使用双尾学生t检验所确定的,这种差异具有统计学显著性(p=0.0004)。这些结果证明pcb076中的合成fviii编码序列的固有表达效率是pcb100中的编码序列的约8倍。这证明,用编码b结构域替代物的序列代替sq接头使该密码子优化的fviii编码序列的固有表达效率提高约8倍。当fviii编码序列在其中fviii编码序列由强肝脏特异性启动子驱动的非整合aav病毒中递送时,这种改善的幅度显著高于对于相同的6个聚糖的基序序列所报告的两倍改善(j.mcintosh等人,blood[血液](2013)121:3335
‑
44)。
[0572]
实例2:用b结构域替代物替代sq接头增加了来自通过aav递送并整合到白蛋白内含子1中的fviii供体盒的fviii表达。
[0573]
为了确定当使用aav将合成fviii编码序列递送到小鼠肝脏时是否发生b结构域替代肽的相同有益作用,构建质粒pcb099(seq id no:311)和pcb102(seq id no:341)并包装在aav8(宾夕法尼亚州莫尔文的载体生物实验室(vector biolabs,malvern,pa)或宾夕法尼亚州费城的赛博泰科科技有限公司(sabtech,philadelphia,pa))中。用以下元件构建质粒(按顺序):itr|grna靶位点(对于malbt1)|18bp间隔区|剪接受体位点(“sa”)|fviii编码序列|聚腺苷酸化信号(“spa”)|grna靶位点|itr。pcb099和pcb102的fviii编码序列分别与pcb076(具有b结构域替代物)和pcb100(仅具有sq接头)的fviii编码序列相同。这些fviii盒缺乏启动子,因此不能作为非整合aav游离基因组表达fviii。通过这些aav病毒递送的
fviii的表达需要邻近适当启动子的整合。
[0574]
在这些实验中,向血友病a小鼠静脉注射2
×
10
12
个载体基因组(“vg”)/千克体重的aav8
‑
pcb099或aav8
‑
pcb102。四周后,给小鼠静脉注射两种lnp的1:1混合物,一种lnp包封malbt1 grna,另一种lnp包封spcas9 mrna。如实例1所述制备lnp,总剂量为2mg rna/kg体重。使用实例1中所述的方法在lnp给药后10天测量小鼠血液中的fviii活性。lnp给药后10天小鼠血液中的fviii水平(图2)平均为接受aav9
‑
pcb099的小鼠的正常人fviii水平的20%,但在接受aav8
‑
pcb102(缺乏b结构域替代物)的小鼠中为本底水平。
[0575]
在lnp给药后第24天,处死小鼠,将整个肝脏匀化,并从一部分肝脏裂解物中提取总基因组dna。使用实例1中描述的dd
‑
pcr测定法定量正向靶向整合到白蛋白内含子1中的频率。每只小鼠的结果汇总在表3中。
[0576]
结果显示注射aav8
‑
pcb099的小鼠中的平均靶向整合频率(%每单倍体基因组)是1.86(
±
0.25),而对于注射aav8
‑
pcb102的小鼠而言平均靶向整合频率是0.46(
±
0.2)。使用双尾学生t检验,这种差异具有统计学显著性(p<0.01)。这些结果证明,包含b结构域替代物导致靶向整合频率高4倍,鉴于先前已经证实包含聚糖代替fviii的b结构域会提高fviii的表达水平,该结果是无法预测的。注射aav8
‑
pcb099的小鼠的血液中的平均fviii水平是正常值的18.6(
±
2.2)%,而对于注射aav8
‑
pcb102的小鼠而言平均fviii水平是正常值的1.7(
±
1.1)%。使用双尾学生t检验,这种11倍的差异具有统计学显著性(p<0.01)。通过将各小鼠中的fviii水平除以靶向整合频率,而将fviii水平归一化为靶向整合频率(表3)。fviii活性除以靶向整合频率得到的比率的平均值对于注射aav8
‑
pcb099的小鼠是10.2(
±
1.7),而对于注射aav8
‑
pcb102的小鼠是3.1(
±
1.7)。使用双尾学生t检验,这种差异具有统计学显著性(p<0.01)。
[0577]
这些结果证明aav8
‑
pcb099中的fviii编码序列的固有表达效率是aav8
‑
pcb102中的约3倍。因为aav8
‑
pcb099与aav8
‑
pcb102的区别仅在于含有n
‑
聚糖基序的序列的存在,这些数据证明aav8
‑
pcb099中的n
‑
聚糖基序赋予固有表达效率三倍的提高。因此,小鼠血液中fviii水平整体提高11倍是由于整合的fviii编码序列的靶向整合提高4倍和表达效率提高3倍的组合。
[0578]
表3:与注射aav8病毒然后注射包封cas9 mrna和malbt1 grna的lnp的小鼠的血液中的峰值fviii水平相比,小鼠肝脏中的靶向整合频率
[0579][0580]
实例3:b结构域替代物中n
‑
聚糖数目的优化
[0581]
实例1和实例2的数据证明插入含有6个n
‑
连接的聚糖基序的b结构域替代物提高了fviii的表达以及靶向整合的频率。然而,这种提高对b结构域替代物中n
‑
聚糖序列数目的依赖性是未知的。因此,我们设计了探测fviii表达这一方面的实验。具体而言,希望确定fviii表达提高所需的n
‑
连接的聚糖基序的最小数目。
[0582]
质粒构建体
[0583]
为了研究不同数目的n
‑
聚糖基序对表达的影响,构建了一系列含有1至9个n
‑
聚糖基序的供体质粒。这些汇总在表4中。所有质粒按5’至3’的顺序由以下序列元件组成:malbt1 grna的靶序列|18bp间隔区|剪接受体|b结构域缺失的fviii编码序列,其中信号肽被tg二核苷酸|聚腺苷酸化信号序列替代。在这些质粒的每一个中,fviii编码序列基于pcb076中使用的密码子优化序列(参见实例1),其中信号肽被tg二核苷酸替代,但在b结构域替代物中具有1至9个n
‑
连接的糖基化位点。所有质粒均含有相同的基于puc19的细菌质粒骨架(含有细菌复制起点和卡那霉素抗性基因)。
[0584]
表4:在b结构域替代物中含有不同数目的n
‑
糖基化位点三联体的fviii供体质粒
[0585]
质粒idn
‑
聚糖基序数目seq id no.蛋白seq id:pcb10301370371pcb10292372373pcb10183331362pcb10174330363pcb10075326364pcb0776317365pcb10066(在c端三联体中s变为t)325366pcb10087327367pcb10158328368pcb10169329369
[0586]
构建体的体内测试:5个、6个和7个聚糖
[0587]
使用实例1的方法,通过流体动力学注射向血友病a小鼠给药每只小鼠30μg的质粒
pcb077、pcb1006、pcb1007或pcb1008。一天后,为相同小鼠眶后注射包封spcas9 mrna和malbt1 grna的lnp的1:1混合物,总rna剂量为2mg/kg体重。lnp如实例1中所述而制备。六天后使用实例1中所述的方法测量小鼠血液中的fviii活性。结果汇总在图3中,并证明由四种质粒供体产生的fviii水平相似。
[0588]
本研究中小鼠血液中产生的fviii水平取决于正向(能够产生fviii蛋白的方向)靶向整合到白蛋白内含子1中的频率和fviii编码序列的固有表达效率。fviii编码序列的固有表达效率是转录速率、翻译效率(其受所用的密码子优化的类型的影响)和分泌过程的效率的函数。在fviii蛋白的情况下,已经提出蛋白的分泌可以是限速步骤(m.swaroop等人,同上)并且与当fviii在细胞中以高水平表达时发生的未折叠蛋白反应相关(r.j.kaufman,同上)。为了区分预期在各小鼠之间变化的靶向整合频率与整合的合成fviii编码序列的固有表达效率,如实例1所述使用微滴式数字pcr(dd
‑
pcr)定量靶向整合频率。
[0589]
在将小鼠用lnp给药后8天,处死小鼠,将整个肝脏匀化,并使用qiagen组织试剂盒纯化总基因组dna。然后使用dd
‑
pcr测定相等质量基因组dna的靶向整合频率。每只小鼠的结果汇总在表5中。各小鼠中正向靶向整合频率范围为0.17%至0.70%,但每组小鼠内四种质粒的平均值相似,对于pcb077、pcb1006、pcb1007和pcb1008而言分别为0.49%、0.47%、0.52%和0.38%。注射pcb077、pcb1006、pcb1007和pcb1008的小鼠的fviii活性与ti的比率的平均值分别为51.33、48.54、48.9和38.9,并且质粒之间的差异不具有统计学显著性。这些结果证明含有5个n
‑
聚糖位点(pcb1007)或7个聚糖位点(pcb1008),或其中一个聚糖三肽基序从nds改变为ndt(pcb1006)的合成fviii编码序列与编码6个n
‑
聚糖位点的合成fviii编码序列(pcb077)相比具有相似的固有表达效率。
[0590]
用质粒pcb1015(seq id no:328)和pcb1016(seq id no:329)进行相同的小鼠研究,其中n
‑
聚糖基序的数目分别变为8和9。另外,构建除了仅具有一个或两个n
‑
聚糖基序之外与pcb077相同的质粒,并使用在lnp中递送的相同grna和spcas9 mrna测试它们在靶向整合到小鼠白蛋白内含子1中之后表达fviii的能力。
[0591]
表5:与血液中的峰值fviii水平相比小鼠肝脏中的靶向整合频率
[0592]
[0593][0594]
构建体的体内测试:3个、4个和5个聚糖
[0595]
纯化质粒pcb1007、pcb1017和pcb1018并如上所述施用于血友病a小鼠。一天后,给予小鼠眶后(ro)注射基于c12
‑
200的包封spcas9 mrna(1mg/kg)和向导rna(grna)malbt1(1mg/kg)的lnp。在lnp给药后5天和7天通过ro放血取血样到柠檬酸钠中(柠檬酸钠与血液的比率为1:9),并通过离心收集血浆。使用实例1中所述的方法测量血浆中的fviii活性。
[0596]
在注射pcb1007、pcb1018和pcb1018的小鼠中,第5天血液中的fviii活性分别平均为8.1%、5.0%和23.5%。第7天,在注射pcb1007、pcb1018和pcb1018的小鼠中fviii活性分别平均为7.9%、3.0%和13.5%。因此,注射具有4个n
‑
聚糖基序(pcb1017)或3个n
‑
聚糖基序(pcb1018)的质粒的小鼠中的fviii表达与接受在b结构域替代物中含有5个n
‑
聚糖基序的质粒(pcb1007)的小鼠中的fviii表达相似。
[0597]
在lnp施用后第7天取血样后,处死小鼠并取出整个肝脏并储存在rnalater
tm
缓冲液(凯杰(qiagen))中。使用基于珠粒的匀浆器匀化整个肝脏,并使用qiagen dna/rna小提试剂盒(目录号80204)从匀浆的等分试样中纯化dna。如实例1所述,通过dd
‑
pcr分析肝脏基因组dna中fviii供体盒正向整合的频率。对于注射pcb1007、pcb1017和pcb1018的小鼠而言,平均靶向整合频率分别为0.27%、0.27%和0.55%,并且这些值没有统计学差异(双尾学生t检验)。
[0598]
将每只小鼠血液中的fviii水平归一化为整合频率提供了fviii编码序列的固有表达效率的量度。fviii水平除以靶向整合频率得到的比率的平均值对于注射pcb1007(5个n
‑
聚糖)的小鼠是23.6,对于注射pcb1017(4个n
‑
聚糖)的小鼠是11.6,并且对于注射pcb1018(3个n
‑
聚糖)的小鼠是23.3。注射pcb1017和pcb1018的小鼠的fviii除以靶向整合的比率与注射pcb1007的小鼠的fviii除以靶向整合的比率在统计学上没有差异。
[0599]
这些数据证明,使用含有具有四个n
‑
聚糖基序或三个n
‑
聚糖基序的b结构域替代
物的合成fviii编码序列,当整合到白蛋白内含子1中时,导致与含有五个n
‑
聚糖基序的fviii编码序列相似的表达。因此,具有带三个n
‑
聚糖基序的b结构域替代物的合成fviii构建体提供了与由带五个n
‑
聚糖基序的b结构域替代物提供的fviii表达相当的提高的fviii表达。据推断,因为含有五个n
‑
聚糖基序的b结构域替代物与含有六个n
‑
聚糖基序的b结构域替代物等效,所以我们推断三个n
‑
聚糖基序与六个n
‑
聚糖基序效力相等。
[0600]
构建体的体内测试:一个和两个聚糖
[0601]
将质粒pcb1018(包含具有带三个n
‑
聚糖基序的b结构域替代物的fviii供体)、pcb1029(包含具有带两个n
‑
聚糖基序的b结构域替代物的fviii供体)和pcb1030(包含具有带一个n
‑
聚糖基序的b结构域替代物的fviii供体)纯化并如上所述通过流体动力学注射施用于血友病a小鼠。一天后,给予小鼠眶后(ro)注射基于c12
‑
200的包封spcas9 mrna(1mg/kg)和grna malbt1(1mg/kg)的lnp。在lnp给药后5天和8天通过ro放血取血样到柠檬酸钠中(1:9柠檬酸钠:血液),并通过离心收集血浆。如上所述测量血浆中的fviii活性,并表示为正常活性的百分比(1u/ml=100%)。
[0602]
在注射pcb1018、pcb1029和pcb1030的小鼠中,第5天血液中的fviii活性分别平均为12.8%、15.8%和13.4%。第八天,在注射pcb1018、pcb1029和pcb1030的小鼠中fviii活性分别平均为13.8%、14.5%和16.0%。因此,注射含有带3个n
‑
聚糖基序(pcb1018)、2个n
‑
聚糖基序(pcb1029)或1个n
‑
聚糖基序(pcb1030)的b结构域替代物的质粒的小鼠中的fviii表达彼此相似。
[0603]
在lnp施用后第7天取血样后,处死小鼠并取出整个肝脏并储存在rnalater
tm
缓冲液(凯杰)中。将整个肝脏匀化,并且如实例1所述,通过dd
‑
pcr分析肝脏基因组dna中fviii供体盒正向整合的频率。对于注射pcb1018、pcb1029和pcb1030的小鼠,平均靶向整合频率分别为0.29%、0.47%和0.36%:这些值没有统计学差异(双尾学生t检验)。
[0604]
将每只小鼠血液中的fviii水平归一化为整合频率提供了fviii编码序列的固有表达效率的量度。fviii水平除以靶向整合频率得到的比率的平均值对于注射pcb1018(3个n
‑
聚糖)的小鼠是41.9,对于注射pcb1029(2个n
‑
聚糖)的小鼠是31.4,并且对于注射pcb1030(1个n
‑
聚糖)的小鼠是40.2。注射pcb1029(3个n
‑
聚糖)和pcb1030(2个n
‑
聚糖)的小鼠的固有表达效率与注射pcb1018(3个n
‑
聚糖)的小鼠的固有表达效率没有统计学差异。这些数据证明,包含含有2个n
‑
聚糖基序(氨基酸序列natnvs)或1个n
‑
聚糖基序(氨基酸序列nat)的b结构域替代物的fviii供体盒以与含有具有3个n
‑
聚糖基序的b结构域替代物的fviii供体盒相同的效率表达。
[0605]
表6:在注射fviii供体pcb1018、pcb1029和pcb1030的小鼠中的fviii活性、靶向整合频率和相对于整合频率归一化的fviii活性
[0606][0607]
包含含有0、1、2、3、4、5、6或7个n
‑
连接的聚糖基序的b结构域替代物的fviii供体盒的体内fviii表达结果的比较
[0608]
比较以上测试的不同fviii盒的固有表达效率。使用相同品系的小鼠(血友病a小鼠)和相同的实验方案,在总共5项研究中产生实例3中描述的数据集。在第5天或第6天测量fviii活性,并在第8天或第9天再次测量fviii活性。在从最后一次fviii活性测量当天(第8或9天)处死的小鼠整个肝脏中提取的dna中测量靶向整合频率。固有表达效率的汇编示于图8。该比较中包括具有不同密码子优化的fviii盒。可对称为“co1”的具有密码子优化的供体进行不同数目的聚糖对归一化fviii表达的影响的比较,这些供体是图8中的前九个柱。这些供体含有fviii盒,其区别仅在于b结构域替代物中n
‑
聚糖基序的数目。含有1
‑
7个n
‑
聚糖基序的聚糖变体的固有表达效率没有显著差异。虽然具有2个n
‑
聚糖基序的供体(“co1
‑
2”)显示出归一化fviii活性较低的趋势(值为30,相比而言具有5、6或7个n
‑
聚糖的变体的值为约45),这种差异在统计学上不显著。没有n
‑
聚糖基序代替b结构域的供体(“co1
‑
0”)表现出显著较低的归一化fviii活性(值为7.4,相比而言具有聚糖和相同密码子优化的变体为40至50)。具有5个聚糖和密码子优化的fviii供体co2与具有5个n
‑
聚糖基序的co1等效,而具有5个n
‑
聚糖的co3以具有5个n
‑
聚糖的co1的效率的约50%表达。这些数据证明,含有包含单个n
‑
聚糖基序的b结构域替代物的fviii编码序列足以赋予与用包含2至7个n
‑
聚糖基序的b结构域替代物获得的fviii表达水平相当的fviii表达水平。含有包含单个n
‑
聚糖基序的b结构域替代物的fviii编码序列(图8中的“co1
‑
1”/pcb1030)是缺少b结构域替代物的相同fviii编码序列(“co1
‑
0”/pcb100)表达效率的约5.4倍(40.1/7.4)。因此,含有具有少于6个n
‑
聚糖(例如5个n
‑
聚糖、4个n
‑
聚糖、3个n
‑
聚糖、2个n
‑
聚糖或1个n
‑
聚糖)的b结构域替代物的fviiii编码序列具有用于基因编辑方法的优点,这是由于添加到fviii蛋白的
非天然氨基酸的数目减少,以及dna供体的大小减少。
[0609]
实例4:在小鼠中靶向整合到安全港基因座(例如白蛋白基因座)中后用于表达的fviii编码序列的最佳密码子优化的鉴定
[0610]
质粒构建体
[0611]
进行实验以确定不同形式的密码子优化对合成fviii表达的影响。通过应用可在基因技术股份公司(co3)获得的市售算法,对含有14个氨基酸的sq接头代替b结构域(1438个氨基酸的总编码序列)的成熟(缺乏信号肽)b结构域缺失的人fviii编码序列进行密码子优化,这将cg二核苷酸的数目从天然序列中存在的54个增加到198个。b结构域缺失的fviii的co3形式的变体(“co4”)通过以下方式产生:通过人工消除全部198个cg二核苷酸,通过根据公开的智人密码子表选择作为次最频繁密码子或更频繁使用的密码子的替代密码子(h.c.brown等人,mol ther meth&clin dev[分子疗法
‑
方法和临床开发](2018)9:57
‑
69(doi:10.1016/j.omtm.2018.01.004)。b结构域缺失的fviii(“fviii
‑
bdd”)编码序列使用基于在肝脏中高度表达的基因的密码子偏倚的算法(h.c.brown等人,同上)进行密码子优化以产生含有176个cg二核苷酸的fviii
‑
bdd co2。还合成了这种经进一步修饰以去除所有cg二核苷酸的构建体,这里称为fviii
‑
bdd co5。还构建了j.mcintosh等人,blood[血液](2013)121(17):3335
‑
44和us 9,393,323的经密码子优化的fviii
‑
bdd编码序列(其中的seq id no:1),在本文中称为“co1”。合成了wo 2011/005968中公开的含有245个cg二核苷酸的b结构域缺失的fviii编码序列(其中的seq id no:5)的进一步密码子优化的变体(本文中的“fviii
‑
bdd co6”)。质粒构建如下:puc19质粒骨架|itr|grnamalbt1的靶位点|18bp间隔区|剪接受体(sa)|tg二核苷酸|b结构域缺失的fviii序列|聚a(spa)|grna malbt1的靶位点|itr,其中供体序列密码子优化为co2(pcb1002,seq id no:323)、co3(pcb1001,seq id no:322)、co4(pcb1000,seq id no:321)或co5(pcb103,seq id no:336)。
[0612]
每个质粒中的fviii供体盒侧接有aav2 itr,并用于使用基于hek293的包装系统将这些盒包装到aav8中,并使用氯化铯密度离心纯化。将所得aav8病毒(命名为aav8
‑
pcb103、aav8
‑
pcb1002、aav8
‑
pcb1001和aav8
‑
pcb1000)使用q
‑
pcr或dd
‑
pcr,用位于fviii基因编码序列内的引物/探针组滴定。这些fviii供体盒被设计为仅在靶向整合到白蛋白内含子1中后表达fviii。供体盒缺乏启动子,因此不能从非整合游离型病毒基因组转录成mrna。另外,所有fviii供体盒在fviii编码序列的n端都缺少信号肽序列,因此可能由非整合游离型病毒拷贝表达的任何蛋白都不能分泌到循环中。在整合到白蛋白内含子1中之后,从基因组中的白蛋白启动子转录产生包含小鼠白蛋白外显子1、内含子1的一部分和合成fviii编码序列的杂交前mrna,终止于fviii供体盒的5’末端所包含的多聚腺苷酸化信号。在白蛋白外显子1的剪接供体与fviii供体盒的5’末端所包含的剪接受体之间剪接这种前mrna产生mrna,其中白蛋白的外显子1编码与成熟fviii编码序列框内融合的信号肽和前原肽。由这种杂交mrna编码的蛋白通过细胞的分泌机制进行加工,在此期间,白蛋白的信号肽和前原肽应被裂解掉,产生预测的双链fviii分子,其中在重链的n端包括在fviii中通常不存在的三个氨基酸。
[0613]
构建体的体内测试
[0614]
为了测试这些配制品,为4或5只血友病a小鼠的队列经由尾静脉注射每种aav8病毒(aav8
‑
pcb103、aav8
‑
pcb1002、aav8
‑
pcb1001和aav8
‑
pcb1000),剂量为2
×
10
12 vg/kg。四
周后,所有小鼠以2mg/kg的总rna剂量静脉注射包封malbt1 grna和spcas9 mrna的lnp的1:1混合物。根据实例1中描述的方法配制lnp。使用实例1中所述的方法测量血液中的fviii活性。结果汇总在图4中。
[0615]
在这些实验中,接受aav8
‑
pcb103和aav8
‑
pcb1002(含有分别具有密码子优化co5和co2的fviii
‑
bdd)的小鼠在其血液中没有可检测的fviii活性。接受病毒pcb1001(密码子优化的co3)的小鼠在lnp给药后第11天具有平均8%的fviii活性并且在第28天具有平均20%的fviii活性。接受病毒aav8
‑
pcb1000(密码子优化co4)的5只小鼠中有3只的fviii活性水平是正常值的1%至3%。这些数据证明,使用geneart算法密码子优化的fviii
‑
bdd dna序列(aav8
‑
pcb1001,co3)导致比基于肝脏中高度表达的基因的最频繁的密码子进行密码子优化的fviii
‑
bdd(aav8
‑
pcb103和aav8
‑
pcb1002)水平更高的fviii表达。与其中使用保留cg二核苷酸的geneart算法对fviii
‑
bdd进行密码子优化的相同盒相比,修饰geneart密码子优化的fviii
‑
bdd序列以去除cg二核苷酸(aav8
‑
pcb1000,co4)导致fviii表达降低。与co2和co5密码子优化不同,具有co4密码子优化的fviii
‑
bdd能够产生可测量的fviii活性。接受aav8
‑
pcb102(co1密码子优化的fviii
‑
bdd dna序列,参见实例2)的小鼠并未产生当在aav8中以2
×
10
12 vg/kg的相同剂量递送并且使用相同剂量的lnp时(实例2,图2,aav8
‑
pcb102)在血友病a小鼠中的fviii活性。这证明在小鼠中靶向整合到白蛋白内含子1中后,对于fviii的表达,co1劣于co3和co4密码子优化的fviii
‑
bdd序列。
[0616]
实例5:在靶向整合到编码具有五个n
‑
聚糖和替代性密码子优化co4和co5的合成fviii的供体模板的白蛋白内含子1中之后fviii在小鼠中的表达。
[0617]
为了测试使用具有b结构域替代物的合成fviii的不同密码子优化的影响,使用命名为co1、co4和co5,并且进一步含有b结构域替代物代替b结构域的三种经密码子优化的dna序列构建缺乏信号肽的fviii
‑
bdd编码序列。b结构域替代物含有五个n
‑
聚糖基序(序列:atnvsnnsntsnds,seq id no:343)。这些编码序列在5’侧上侧接有malbt1 grna的靶位点、18bp间隔区、剪接受体和两个核苷酸(tg)。tg二核苷酸在剪接到小鼠白蛋白外显子1后保持正确的阅读框。在编码序列的3’末端包含短的多聚腺苷酸化信号(spa)。这三种质粒中的合成fviii编码序列编码具有相同氨基酸序列的fviii蛋白,但由于密码子优化不同,编码序列由不同的dna序列编码。在血友病a小鼠中测试命名为pcb1007(co1,seq id no:326)、pcb1019(co4,seq id no:332)和pcb1020(co5,seq id no:333)的这些质粒在通过malbt1 grna靶位点处的crispr/cas9裂解介导靶向整合到白蛋白内含子1中之后表达活性fviii蛋白的能力。
[0618]
实验方案与实例1中的相同。使用qiagen 大提试剂盒(目录号12362)纯化质粒pcb1007、pcb1019和pcb1029的质粒dna,然后在0.9%盐水中稀释到最终浓度为15μg/ml。通过hdi向血友病a小鼠队列每只小鼠注射2ml经稀释的质粒dna。一天后给予小鼠眶后注射基于c12
‑
200的包封spcas9 mrna(1mg/kg体重)和grna malbt1(1mg/kg体重)的lnp。在给药后3天处死仅注射包封spcas9 mrna和malbt1 grna的lnp的五只血友病a小鼠的队列,并分析从整个肝脏提取的基因组dna在白蛋白内含子1中的靶标位点处的插入缺失。平均插入缺失频率为52.9%,表明在肝脏中的中靶位点有效裂解。
[0619]
用lnp对注射了质粒的小鼠给药六天和九天后,通过ro放血取血样到柠檬酸钠中(柠檬酸钠与血液的比率为1:9),并且通过离心收集血浆。使用实例1中所述的方法测量血
浆中的fviii活性。结果汇总在图5中。
[0620]
在lnp给药后第6天,接受质粒pcb1007、pcb1019或pcb1020的小鼠中的平均fviii活性为正常值的22.3%、17.6%和17.8%。在lnp给药后第9天,接受质粒pcb1007、pcb1019或pcb1020的小鼠中的平均fviii活性为正常值的19.7%、14.1%和14.9%。当使用同方差(2样本等方差)双尾t检验(全部p值>0.28)评价时,以三种质粒给药的小鼠中的fviii水平在第6天或第9天没有统计学显著差异。
[0621]
这些结果证明,在编码具有含5个n
‑
聚糖基序的b结构域替代物代替b结构域的合成fviii的供体模板的情况下,密码子优化co1、co4和co5(所有这些都缺少cg二核苷酸)在靶向整合到白蛋白内含子1中之后产生相似水平的fviii。因此,特定密码子优化没有明显优势,并且任何无cpg的密码子优化(例如,co1、co4和co5)在靶向整合后提供相似水平的合成fviii蛋白。
[0622]
实例6:b结构域替代物和f309突变为s或a的组合
[0623]
据报道,a1结构域中伴侣免疫球蛋白结合蛋白(bip)的潜在结合位点内的点突变(f309s)使培养的细胞中b结构域缺失的fviii的分泌提高约3倍(m.swaroop等人,j biol chem[生物化学杂志](1997)272:24121
‑
24)。fviii的f309a突变蛋白类似地提高了分泌。据报道,与b结构域缺失的fviii相比,b结构域的f309s和226个氨基酸的n端部分的组合使小鼠体内的fviii水平提高20至30倍,而添加b结构域的226个氨基酸的n端使fviii水平仅提高5倍(h.z.miao等人,blood[血液](2004)103(9):3412
‑
19)。
[0624]
为了评价b结构域替代物与用丝氨酸或丙氨酸取代309处苯丙氨酸残基的组合是否引起靶向整合后fviii表达的进一步改善,构建了质粒pcb1025(seq id no:334)和pcb1026(seq id no:335)。两种质粒均含有co4密码子优化的fviii dna序列,该序列具有含5个n
‑
连接的糖基化位点的b结构域替代物。这些质粒具有以下元件:puc19质粒骨架|grna malbt1的靶位点|18bp间隔区|剪接受体(sa)|tg二核苷酸|具有五个位点b结构域替代物的fviii序列(co4)|聚a(spa)。质粒pcb1007与pcb1025和pcb1026相同,除了pcb1025在位置309处具有ala而不是phe,而pcb1026在位置309处具有ser而不是phe以外。质粒pcb1007在该研究中用作比较物。
[0625]
实验方案与实例1中的相同。使用qiagen 大提试剂盒(目录号12362)纯化质粒pcb1007、pcb1025和pcb1026,然后在0.9%盐水中稀释到最终浓度为15μg/ml。通过hdi向血友病a小鼠队列每只小鼠注射2ml经稀释的质粒dna。一天后,给予小鼠ro注射基于c12
‑
200的包封spcas9 mrna(1mg/kg)和grna malbt1(1mg/kg)的lnp。lnp(pcb1025、pcb1026)给药后5天或lnp(pcb1019)给药后6天,通过ro放血取血样到柠檬酸钠中(柠檬酸钠与血液的比率为1:9),并且通过离心收集血浆。使用实例1中所述的方法测量血浆中的fviii活性。在注射pcb1019、pcb1025或pcb1026的三组小鼠中fviii活性相似,平均fviii活性分别为17.6%、27.2%和24.5%。
[0626]
在lnp后第9天(注射pcb1019的小鼠)或lnp后第7天(注射pcb1025和pcb1026的小鼠)还测定了相同血友病a小鼠的血液中的fviii活性。然后处死小鼠,准备整个肝脏并如上述实例1所述分析整合频率。三组之间的靶向整合频率相似,注射pcb1019的小鼠的平均频率为0.42,注射pcb1025的小鼠的平均频率为0.47,注射pcb1026的小鼠的平均频率为0.36。
[0627]
将每只小鼠血液中的fviii水平归一化为整合频率提供了fviii编码序列的固有
表达效率的量度。fviii水平除以靶向整合频率得到的比率的平均值对于注射pcb1019的小鼠是37.4,对于注射pcb1025的小鼠是41.5,并且对于注射pcb1026的小鼠是49.9。与注射pcb1019的小鼠相比,针对注射pcb1025和pcb1026的小鼠的靶向整合归一化的fviii活性的差异在统计学上不显著(双尾学生t检验),这证明将氨基酸f309改变为丝氨酸或丙氨酸(在含有5个n
‑
聚糖基序代替b结构域的fviii
‑
bdd盒的情况下)并未提高fviii表达。因此,并非对fviii蛋白进行的所有氨基酸改变在靶向整合到白蛋白内含子1中后都对fviii表达具有影响。
[0628]
实例7:通过crispr/cas核酸酶将合成fviii靶向整合到转铁蛋白内含子1中引起治疗水平的人fviii的表达。
[0629]
dna构建体
[0630]
为了检查向转铁蛋白基因座的整合和从转铁蛋白基因座的表达,作为白蛋白基因座的替代,如下按5’至3’的顺序用序列元件构建人fviii供体盒(seq id no:224):aav2的反向末端重复序列(itr)|grna mtf
‑
t2的靶位点|18bp间隔区|剪接受体|编码小鼠转铁蛋白的信号肽的最后4个氨基酸的序列(ggctgtgtctggct,seq id no:225)|合成fviii编码序列|多聚腺苷酸化信号(spa)|grna mtf
‑
t2的靶位点|和aav2的反向末端重复序列(itr)。grna mtf
‑
t2的靶位点的序列是小鼠基因组中靶序列的反向互补序列,可能有利于正向整合。聚腺苷酸化信号是一个49bp的短序列,已报道该序列会有效地指导聚腺苷酸化(n.levitt等人,genes dev[基因与发育](1989)3:1019
‑
25)。合成fviii编码序列编码含有氨基酸序列的代替b结构域的b结构域替代物,并且包括替代b结构域的31个氨基酸的异源序列。该序列含有六个对应于n
‑
连接的糖基化位点的三肽(以粗体表示),并且表明其可提高fviii的表达(j.mcintosh等人,blood[血液](2013)121:3335
‑
44)。
[0631]
在用三种质粒转染的hek293细胞中使用已确立的病毒包装方法实现将pcb1009 fviii供体dna包装到aav8中;一种编码aav包装蛋白,第二种编码腺病毒辅助蛋白,以及第3种含有侧翼是aav itr序列的fviii供体dna序列。转染的细胞产生由第一质粒上编码的aav衣壳蛋白的组成所指定的血清型的aav颗粒。从细胞上清液或从上清液和溶解的细胞中收集这些aav颗粒,并通过cscl梯度纯化。通过微滴式数字pcr(dd
‑
pcr)测量供体dna的基因组拷贝数来定量纯化的病毒颗粒。
[0632]
构建体的体内测试
[0633]
五只血友病a小鼠的队列以2
×
10
12 vg/kg体重的剂量向尾静脉中静脉(i.v.)注射aav8
‑
pcb1009。aav8病毒优先转导肝细胞。四周后,以2mg/kg体重的总rna剂量,向相同小鼠静脉注射两种lnp的1:1(按rna质量计)混合物,一种lnp包封spcas9 mrna,一种lnp包封向导rna mtf
‑
t2。lnp主要被肝细胞吸收。lnp给药后10天,获得血样并如上所述进行测定。fviii活性平均为正常人fviii水平的954%(
±
251%)(图6),相当于9.54iu/ml或是无血友病的人中的平均水平的9.5倍。原初血友病a小鼠的fviii活性不可检测(<正常值的0.5%)。
[0634]
这些数据证明将fviii编码序列靶向整合到转铁蛋白的内含子1中可引起高水平的fviii表达和活性,证明了该方法用于治疗具有缺陷型fviii的病状诸如血友病a的效用。
[0635]
实例8:另外的递送模式
[0636]
在另一个实例中,使用非病毒lnp递送系统在体内递送供体模板。将dna分子包封
到类似于上述那些的lnp颗粒中,并通过静脉注射递送至肝脏。虽然dna从内体向细胞质的逃逸相对有效地发生,但是带电的dna大分子向细胞核的易位中却效率不高。在一种情况下,通过将aav itr序列并入供体模板中来模拟aav基因组,从而改善dna向细胞核的递送。在这种情况下,itr序列使dna稳定或改善核易位。从供体模板序列中去除cg二核苷酸(cpg序列)也改善了核递送。含有cg二核苷酸的dna被先天免疫系统识别并消除。去除人工dna序列中存在的cpg序列改善了通过非病毒载体和病毒载体递送的dna的持久性。密码子优化的过程通常会增加cg二核苷酸的含量,这是因为在许多情况下最常见的密码子在第3位置具有c残基,这在下一个密码子以g开头时会增加产生cg的概率。在血友病a小鼠中评价lnp递送供体模板并于一小时至五天后接着递送含grna和cas9 mrna的lnp的组合。
[0637]
grna和cas9 mrna的体内递送可以通过已知方法完成。在一种方法中,从aav病毒载体表达grna和cas9蛋白。在这种情况下,grna的转录由u6启动子驱动,而cas9 mrna的转录则由普遍存在的启动子(例如ef1
‑
α)或肝脏特异性启动子/增强子(诸如甲状腺素运载蛋白启动子/增强子)驱动。spcas9编码序列的大小(4.4kb)妨碍了在单个aav中包含spcas9和grna盒,因而需要单独的aav来递送grna和spcas9。在第二种情况下,使用具有促进病毒基因组自我灭活的序列元件的aav载体。在这种情况下,在载体dna中包含grna的裂解位点导致体内载体dna的裂解。通过在裂解时阻断cas9表达的位置中包含裂解位点,将cas9表达限制在较短的时间段内。在将grna和cas9递送至体内细胞的第三种替代方法中,使用非病毒递送方法。在一个实例中,将lnp用作非病毒递送方法。有几种不同的可电离的阳离子脂质可用于lnp。这些包括c12
‑
200、mc3、ln16和md1等。在一种类型的lnp中,galnac部分附接到lnp的外部,并充当经由脱唾液酸糖蛋白受体摄取到肝脏中的配体。这些阳离子脂质中的任一种均可用于配制lnp,以将grna和cas9 mrna递送至肝脏。
[0638]
实例9:治疗性编码序列在小鼠纤维蛋白原α内含子1处的靶向整合
[0639]
为了检查纤维蛋白原α基因座的整合和表达,作为白蛋白或转铁蛋白基因座的替代,构建了aav8病毒(aav8
‑
pcb1010,seq id no:361),其具有带以下元件的盒:grna mfga
‑
t6的靶位点、18bp间隔区、fix剪接受体、成熟人fviii编码序列(n端经修饰以在剪接至内源fga外显子1后完成fga信号肽)(其中b结构域被6个n
‑
聚糖基序替代)、多聚腺苷酸化序列和grna mfga
‑
t6的靶位点。
[0640]
向血友病a小鼠注射aav8
‑
pcb1010,28天后注射包封t6 grna(靶向小鼠纤维蛋白原α内含子1)和cas9 mrna的lnp。lnp给药后10天,通过眶后放血取血样到装有柠檬酸钠(柠檬酸钠与血液的比率为1:9)的毛细管中,并通过离心收集血浆。然后如上所述测定血浆样品的fviii。测定结果报告为正常人fviii活性的百分比(正常值定义为1iu/ml)。fviii活性平均为正常人fviii水平的1124%(
±
527%),相当于11.24iu/ml或是无血友病的人中的平均水平的11倍。原初血友病a小鼠的fviii活性不可检测(<正常值的0.5%)。因为aav8
‑
pcb1010病毒含有fviii盒,其中编码序列缺乏信号肽并且也缺乏启动子,所以单独的这种病毒不能产生分泌型fviii蛋白。
[0641]
这些数据证明纤维蛋白原作为编码序列的插入位点的适用性。此外,他们证明b结构域替代的fviii序列可用于表达有用量的fviii。因此,此类构建体和方法可用于治疗与缺陷型fviii相关的病症。
[0642]
实例10:在培养的原代人肝脏细胞中在人白蛋白内含子1处有效裂解的向导rna的
鉴定和选择
[0643]
为了证明本发明的系统在人肝脏细胞中的操作,制备了四种grna(t4
‑
seq id no:357、t5
‑
seq id no:358、t11
‑
seq id no:359和t13
‑
seq id no:360),其基于在人和非人灵长类动物之间具有完美的同一性并针对huh7和hepg2细胞中的切割效率进行筛选,用于评价在原代人肝脏细胞中的切割效率。将原代人肝脏细胞(从bioivt获得)解冻,转移至冷冻保存的肝细胞恢复培养基(chrm)(gibco)中,低速沉淀,然后以0.7
×
106个细胞/ml的密度接种在预先涂有胶原iv(康宁)的24孔板中的invitrogro
tm
cp培养基(bioivt)加上torpedo
tm
抗生素混合物(bioivt)中。将板在5%co2中于37℃孵育。细胞粘附后(接种后3
‑
4小时),用新鲜温热的完全培养基洗去未粘附到板上的死细胞,添加另外的培养基,并将细胞在5%co2中于37℃孵育。为了转染细胞,将cas9 mrna(trilink)和向导rna在冰上解冻,然后以每孔0.6μg mrna和0.2μg向导rna添加到30μl opti
‑
mem
tm
培养基(gibco)中。将以30μl按2:1体积:总核酸重量稀释于opti
‑
mem
tm
中的messengermax
tm
(赛默飞世尔)与cas9 mrna/grna opti
‑
mem
tm
溶液在室温下孵育20分钟。将该混合物滴加到24孔板中每孔培养的肝细胞的500μl肝细胞接种培养基中,并将细胞在5%co2中于37℃孵育。第二天早上,洗涤细胞并重新饲养。转染后48小时,通过向每个孔中添加200μl温热的0.25%胰蛋白酶
‑
edta(gibco)并在37℃孵育5至10分钟,收集细胞用于基因组dna提取。一旦细胞脱落,就添加200μl fbs(gibco)使胰蛋白酶失活。添加1ml pbs(gibco)之后,将细胞以1200rpm沉淀三分钟,然后重悬于50μl pbs中。按照试剂盒说明书,使用magmax
tm
dna multi
‑
sample ultra 2.0试剂盒(应用生物系统公司(applied biosytems))提取基因组dna。使用分光光度计分析基因组dna的质量和浓度。对于tide分析,使用在预测的中靶裂解位点侧翼的引物(albf:ccctccgtttgtcctagcttttc,seq id no:353,和albr:ccagatacagaatatcttcctcaacgcaga,seq id no:354)和pcr supermix high fidelity(invitrogen
tm
),使用35个pcr循环和55℃的退火温度对基因组dna进行pcr扩增。通过琼脂糖凝胶电泳分析pcr产物,以确认已生成合适大小的产物(1053bp),然后纯化并使用引物(正向引物:cctttggcacaatgaagtgg,seq id no:355,反向引物:gaatctgaaccctgatgacaag,seq id no:356)进行测序。使用命名为tsunami的tides算法修改版本(e.k.brinkman等人,nuc acids res[核酸研究](2014)42(22):e168)分析序列数据,该算法确定grna/cas9复合物的预测切割位点处存在的插入缺失的频率。
[0644]
测试含有t4(seq id no:357)、t5(seq id no:358)、t11(seq id no:359)和t13(seq id no:360)向导的20个核苷酸靶的序列或19个核苷酸的靶序列(在5’末端的1bp较短序列)的向导rna。19个核苷酸的grna可具有更高的序列特异性,但较短的向导可具有较低的效力(双链裂解的效率,作为插入缺失进行测量)。将靶向人aavs1基因座和人补体因子的对照向导包括在内以在供体之间进行比较。转染后48小时,使用tides方法测量白蛋白内含子1中靶位点处的插入缺失频率。图7汇总了来自四个不同人类供体的原代肝细胞转染的结果。
[0645]
结果证明,不同向导的切割效率范围在20%至80%之间。每种白蛋白grna的20个核苷酸型式始终比19个核苷酸的变体更有效。20个核苷酸的grna的优良效力可抵消19个核苷酸的grna就脱靶切割较低而言可能具有的任何潜在益处。向导rna t4在四个细胞供体上表现出最一致的切割,其插入缺失频率为约60%。
[0646]
实例11:评价在具有靶向转铁蛋白基因座的grna的单一lnp剂量后包封具有由不同数目的n
‑
聚糖组成的b结构域替代物的密码子优化的fviii编码序列(无cpg)的aav8病毒的fviii表达
[0647]
该研究评价了编码fviii的aav8病毒的fviii表达,其中b
‑
结构域替代物含有0、1、3、5或6个聚糖。对fviii编码序列进行密码子优化,然后人工消除cpg。该研究中使用的构建体如图9所示。
[0648]
第0天,通过尾静脉注射用相应的病毒对血友病a小鼠(8
‑
10周龄)给药。第28天,向血友病a小鼠眶后注射包封cas9 mrna(411μg/ml)和向导rnamtf
‑
t2(379μg/ml)的脂质纳米颗粒(lnp)。研究组和剂量示于表7。
[0649]
表7.研究组和剂量。
[0650][0651][0652]
lnp给药后11天,获得血样并如上所述进行测定。然后,在lnp给药后18天,经由心脏末端放血获得血样并如上所述进行测定。
[0653]
第11天测量的fviii活性水平示于图10。第18天测量的fviii活性水平示于图11。表8和表9中提供了fviii活性水平。
[0654]
表8.第11天的fviii活性水平。
[0655][0656]
表9.第18天的fviii活性水平。
[0657][0658]
处死小鼠后,将整个肝脏匀化,并从一部分肝脏裂解物中提取总基因组dna。使用实例1中描述的dd
‑
pcr测定法定量正向靶向整合到白蛋白内含子1中的频率。结果示于图12和表10中。
[0659]
表10.fviii靶向整合频率。
[0660][0661]
这些数据证明含有0、1、3、5或6个聚糖的fviii编码序列可引起高水平的fviii表达和活性,证明了该方法用于治疗具有缺陷型fviii的病状诸如血友病a的效用。
[0662]
虽然已经相对于描述的几个实施例相当详细地以一定特殊性描述了本披露,但是并不意图应该将本披露限于任何此类细节或实施例或任何特定的实施例,而是参考所附权利要求来对其进行解释,以便按照本领域的观点来提供对此类权利要求的最广泛的可能解
释,以有效地涵盖本披露的预期范围。
[0663]
序列表
[0664]
除了本披露中其他地方披露的序列之外,还提供了以下序列,因为在本披露的示例性实施例中提到或使用了以下序列,这些序列是为了说明的目的而提供的。
[0665]
[0666]
[0667]
[0668]
[0669]
[0670]
[0671]
[0672]
[0673]
[0674]
[0675]
[0676]
[0677]
[0678]
[0679]
[0680]
[0681]
[0682]
[0683]
[0684]
[0685]
[0686]
[0687]
[0688]
[0689]
[0690]
[0691]
[0692]
[0693]
[0694]
[0695]
[0696]
[0697]
[0698]
[0699]
[0700]
[0701]
[0702]
[0703]
[0704]
[0705]
[0706]
[0707]
[0708]
[0709]
[0710]
[0711]
[0712]
[0713]
[0714]
[0715]
[0716]
[0717]
[0718]
[0719]
[0720]
[0721]
[0722]
[0723]
[0724]
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1