检测组织样品中裂解的SNAP25的方法与流程
相关申请的交叉引用
本申请要求2014年7月7日提交的美国临时申请序号62/021,379、2015年5月8日提交的62/158,900以及2015年5月19日提交的62/163,829的权益,所述美国临时申请全部通过引用完整并入本文。
发明的技术领域
本公开涉及用于检测组织样品中裂解的snap25的方法和组合物,例如snap25的肉毒杆菌神经毒素裂解。本发明还提供了结合裂解的snap25的抗体,所述裂解的snap25包括由血清型a肉毒杆菌神经毒素产生的snap25裂解产物。
发明背景
a型肉毒杆菌神经毒素(bont/a)的治疗效用在过去数十年显著增长,并且目前,allergan的产品肉毒杆菌毒素a(onabotulinumtoxina)在全球批准了用于11种主要的治疗和美容适应症,包括治疗各种神经肌肉疾患(ramirez-castaneda,j.和jankovic,j.,(2014).“long-termefficacy,safety,andsideeffectprofileofbotulinumtoxinindystonia:a20-yearfollow-up.”toxicon90,344-348.;yablon,s.a.,brin,m.f.,vandenburgh,a.m.,zhou,j.,garabedian-ruffalo,s.m.,abu-shakra,s.,和beddingfield,f.c.,iii(2011).“doseresponsewithonabotulinumtoxinaforpost-strokespasticity:apooleddataanalysis.”movdisord.26,209-215.)、平滑肌和自主神经功能紊乱(ellsworth,p.和travis,m.,(2014).“onabotulinumtoxina:atherapeuticoptionforrefractoryneurogenicdetrusoroveractivityandidiopathicoveractivebladder.”urol.nurs.34,165-171.;grunfeld,a.,murray,c.a.,和solish,n.,(2009).“botulinumtoxinforhyperhidrosis:areview.”am.j.clin.dermatol.10,87-102.)和伤害性疼痛综合征(aoki,k.r.和francis,j.,(2011).“updatesontheantinociceptivemechanismhypothesisofbotulinumtoxina.”parkinsonism.relatdisord.17增刊1,s28-s33.;burstein,r.,zhang,x.,levy,d.,aoki,k.r.,和brin,m.f.,(2014).“selectiveinhibitionofmeningealnociceptorsbybotulinumneurotoxintypea:therapeuticimplicationsformigraineandotherpains.”cephalalgia34,853-869.)。
虽然已经确认bont/a在突触前神经末端的一般作用机制(moa)(montal,m.,(2010).“botulinumneurotoxin:amarvelofproteindesign.”annu.rev.biochem.79,591-617.),依然有许多关于细胞内运输模式和毒素一般“生命周期”的尚未解答的问题。解决这些问题部分取决于精确检测毒素在细胞内的定位、分布和移动的能力。因为其高效能并因此在神经元内极低的浓度,使用抗体直接检测bont/a是困难的。用于检测bont/a存在的可选方法曾经是通过对裂解snap25产物(snap25197)的免疫染色来跟踪其酶活性。已经使用商业和专有抗体跟踪全长snap25(snap25206)或bont/a裂解的snap25(snap25197)的表达。然而,在bont/a裂解之后产生的snap25197的末端表位难以用抗体特异性靶向而也不识别完整snap25蛋白(mort,j.s.和buttle,d.j.,(1999).“theuseofcleavagesitespecificantibodiestodelineateproteinprocessingandbreakdownpathways.”mol.pathol.52,11-18.)。因此,免疫染色结果是令人误解的,其中一些抗体是测定依赖性的,而其他抗体是组织特异性的。因此,需要非常有选择性的针对bont/a裂解的snap25的抗体,其可以识别暴露于bont/a之后的任何组织类型和多种测定中的snap25197。
本公开内容通过以下方式解决这些问题:提供用于在各种不同测定中检测肉毒杆菌毒素裂解的snap25(包括bont/a裂解的snap25和在位置‘197’处裂解snap25的任何其他bont/a相关化合物)的方法和组合物,所述测定例如但不限于使用针对snap25197的高特异性重组单克隆抗体(rmab)的免疫组织化学。这些抗体可用于检测来自不同物种的许多组织中的snap25197,所述物种包括但不限于人。这些抗体还可以作为工具来诊断已经用神经毒素,例如但不限于肉毒杆菌毒素a处理过的人组织中的活性和功效。
发明概述
本发明涵盖抗snap25抗体,其中所述抗体优先结合bont/a裂解的snap25。在一些实施方案中,抗snap25抗体优先结合已由具有血清型a肉毒杆菌毒素的酶(轻链)活性的重组肉毒杆菌毒素裂解的snap25。在其他实施方案中,抗snap25抗体不结合全长或未裂解的snap25。
在其他实施方案中,抗snap25抗体能够检测组织中bont/a裂解的snap25(或者由具有血清型a肉毒杆菌毒素的酶(轻链)活性的重组肉毒杆菌毒素裂解的snap25)。在一些实施方案中,所述组织是活检样品。在其他实施方案中,所述组织是皮肤取样(skinpunch)。
在一些实施方案中,抗snap25抗体包含seqidno:1或seqidno:2的轻链序列,并且是重组鼠抗体。在其他实施方案中,抗snap25抗体包含seqidno:3或seqidno:4的重链序列,并且是重组鼠抗体。在其他实施方案中,抗snap25抗体包含seqidno:5或seqidno:6的轻链序列,并且是重组人抗体。在其他实施方案中,抗snap25抗体包含seqidno:7或seqidno:8的重链,并且是重组人抗体。在其他实施方案中,抗snap25抗体是结合具有包含seqidno:1-8的一个或多个序列的重链和/或轻链的抗体的相同表位的抗体。
本发明还涵盖诊断组织是否已经暴露于bont/a酶活性的方法,其包括使怀疑已经暴露于bont/a酶活性的组织样品与抗snap25抗体接触,其中所述抗体优先结合bont/a裂解的snap25;和检测所述抗snap25抗体是否结合了所述组织样品,其中结合所述组织样品的抗snap25抗体的存在指示所述组织样品已经暴露于bont/a活性。在一些实施方案中,bont/a酶活性来自天然bont/a。在其他实施方案中,bont/a活性来自具有血清型a肉毒杆菌毒素的酶(轻链)活性的重组肉毒杆菌毒素。在一些实施方案中,该方法中使用的抗snap25抗体包含seqidno:1或seqidno:2的轻链序列,并且是重组鼠抗体。在其他实施方案中,该方法中使用的抗snap25抗体包含seqidno:3或seqidno:4的重链序列,并且是重组鼠抗体。在其他实施方案中,该方法中使用的抗snap25抗体包含seqidno:5或seqidno:6的轻链序列,并且是重组人抗体。在其他实施方案中,该方法中使用的抗snap25抗体包含seqidno:7或seqidno:8的重链,并且是重组人抗体。在其他实施方案中,该方法中使用的抗snap25抗体是结合具有包含seqidno:1-8的一个或多个序列的重链和/或轻链的抗体的相同表位的抗体。
另一方面,本发明涵盖用于诊断组织是否已经暴露于bont/a酶活性的试剂盒,其包括抗snap25抗体,其中所述抗体优先结合bont/a裂解的snap25。在一些实施方案中,bont/a酶活性来自天然bont/a。在其他实施方案中,bont/a活性来自具有血清型a肉毒杆菌毒素的酶(轻链)活性的重组肉毒杆菌毒素。在一些实施方案中,该试剂盒中使用的抗snap25抗体包含seqidno:1或seqidno:2的轻链序列,并且是重组鼠抗体。在其他实施方案中,该试剂盒中使用的抗snap25抗体包含seqidno:3或seqidno:4的重链序列,并且是重组鼠抗体。在其他实施方案中,该试剂盒中使用的抗snap25抗体包含seqidno:5或seqidno:6的轻链序列,并且是重组人抗体。在其他实施方案中,该试剂盒中使用的抗snap25抗体包含seqidno:7或seqidno:8的重链,并且是重组人抗体。在其他实施方案中,该试剂盒中使用的抗snap25抗体是结合具有包含seqidno:1-8的一个或多个序列的重链和/或轻链的抗体的相同表位的抗体。
附图简述
图1显示了针对snap25197的人源化和鼠重组单克隆抗体的推定表位的图示。该图显示了用于产生原始单克隆抗体的、去掉n末端半胱氨酸残基(用于缀合钥孔虫戚血兰素)的12残基肽。
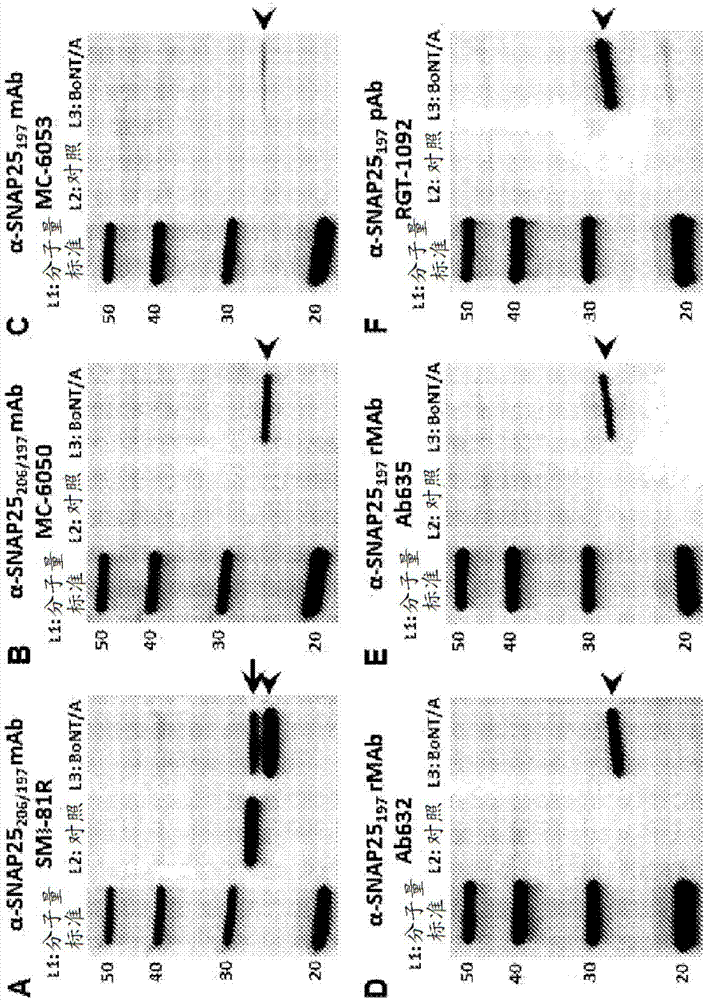
图2(a-f)显示蛋白质印迹分析,比较了用于检测用(l3)或未用(l2)3nm浓度的bont/a处理的大鼠胚胎皮层细胞裂解物中全长snap25206和裂解的snap25197的不同抗体的特异性。(a)用识别全长(206)和裂解(197)形式snap25的可商购的抗snap25mab(smi-81r)探测的印迹。在泳道2中,仅检测到snap25206,而在泳道3中,检测到snap25206(箭)和snap25197(箭头)两者。(b)用据报道识别snap25206和snap25197两者的可商购的抗snap25mab(mc-6050)探测的印迹。仅snap25197作为泳道3中的单一条带出现(箭头)。(c)用据报道对snap25197特异的可商购的抗snap25mab(mc-6053)探测的印迹。该抗体识别泳道3中细的模糊snap25197条带。(d)用ab632抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(e)用ab635抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(f)用rgt-1092抗snap25197pab探测的印迹。该抗体主要识别泳道3中的snap25197(箭头),但是仅在snap25197条带上下可见两个模糊条带。泳道1,蛋白质分子量标准;泳道2,未处理的皮层细胞裂解物;泳道3,bont/a处理的(3nm)皮层细胞裂解物。
图3(a-f)显示蛋白质印迹分析,比较了用于检测用(l3)或未用(l2)3nm浓度的bont/a处理的sima细胞裂解物中全长snap25206和裂解snap25197的不同抗体的特异性。(a)用识别全长(206)和裂解(197)形式snap25的可商购的抗snap25mab(smi-81r)探测的印迹。在泳道2中,仅检测到snap25206,而在泳道3中,检测到snap25206(箭)和snap25197(箭头)两者。(b)用据报道识别snap25206和snap25197两者的可商购的抗snap25mab(mc-6050)探测的印迹。仅snap25197作为泳道3中的单一条带出现(箭头)。(c)用据报道对snap25197特异的可商购的抗snap25mab(mc-6053)探测的印迹。该抗体没有显示识别任何条带。(d)用ab632抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(e)用ab635抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(f)用rgt-1092抗snap25197pab探测的印迹。该抗体主要识别泳道3中的snap25197(箭头),但是仅在snap25197条带上下可见两个模糊条带。泳道1,蛋白质分子量标准;泳道2,未处理的sima细胞裂解物;泳道3,bont/a处理的(0.01nm)sima细胞裂解物。
图4(a,b)显示经抗gapdhmab探测的图2中皮层细胞研究和图3中sima细胞研究的对照蛋白质印迹分析,证明了泳道2和3中的相等的样品负载。泳道1,蛋白质分子量标准;泳道2,未处理的细胞裂解物;泳道3,bont/a处理的(3nm)皮层细胞裂解物和(0.01nm)sima细胞裂解物。图4(c,d)显示证明ab632-rmab的表位特异性的蛋白质印迹分析,使用了经bont/a(l3)、bont/c(l4)、bont/e(l5)或没有毒素(l2)处理的sima细胞裂解物。(c)用识别全长(206)和裂解(bont/a的197,bont/c的198,和bont/e的180)形式snap25的可商购的抗snap25mab(smi-81r)探测的印迹。在泳道2中,仅检测到snap25206,而在泳道3中,检测到snap25206(箭)和snap25197两者。在泳道4中,检测到snap25206(箭)和snap25198两者,并且在泳道5,检测到snap25206(箭)和snap25180两者;(d)用ab632抗snap25197rmab探测的印迹。在泳道2、4和5中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带。泳道1,蛋白质分子量标准;泳道2,未处理的sima细胞裂解物;泳道3,bont/a处理的sima细胞裂解物;泳道4,bont/c处理的sima细胞裂解物;泳道5,bont/e处理的sima细胞裂解物。
图5(a-j)显示免疫组织化学分析,比较了抗体针对用肉毒杆菌毒素a(10u/kg)或媒介物处理之后大鼠膀胱切片中snap25的特异性。(a-e)用肉毒杆菌毒素a注射并用以下抗体探测的膀胱逼尿肌(dm)的共聚焦图像:(a)针对全长(206)和裂解的(197)snap25的商业mab(smi-81r),(b)针对snap25197和snap25206的第二商业mab(mc-6050),(c)针对snap25197的商业mab(mc-6053),(d)针对snap25197的ab632-rmab,和(e)针对snap25197的rgt-1092pab。(f-j)用媒介物注射和用相同的五种抗体探测的对照大鼠膀胱的共聚焦图像。箭(f和j)指示来自媒介物处理的膀胱的神经纤维内的ir信号。dm,逼尿肌;比例尺=50μm。
图6(a-j)显示免疫组织化学分析,比较了抗体针对用肉毒杆菌毒素a(30u/kg)或媒介物处理之后大鼠无毛皮肤切片中snap25的特异性。(a-e)用肉毒杆菌毒素a注射并用以下抗体探测的大鼠皮肤的共聚焦图像:(a)针对全长(206)和裂解的(197)snap25的商业mab(smi-81r),(b)针对snap25197和snap25206的第二商业mab(mc-6050),(c)针对snap25197的商业mab(mc-6053),(d)针对snap25197的ab632-rmab,和(e)针对snap25197的rgt-1092pab。(f-j)用媒介物注射和用相同的五种抗体探测的对照大鼠皮肤的共聚焦图像。箭(f、g、h和j)指示来自媒介物大鼠皮肤的神经纤维内的ir信号。星号(b、c、g和h)指示血管腔内非特异性ir信号。bv,血管;ct,结缔组织;比例尺=50μm。
图7(a-h)显示免疫组织化学分析,比较了抗体针对用肉毒杆菌毒素a(30u/kg)或媒介物处理之后大鼠无毛皮肤下骨骼肌中snap25的特异性。(a–d)显示用肉毒杆菌毒素a注射并用以下抗体探测的大鼠爪的下面肌肉内运动神经末梢(mnt,箭)的共聚焦图像:(a)针对全长(206)和裂解的(197)snap25的商业mab(smi-81r),(b)针对snap25197和snap25206的第二商业mab(mc-6050),(c)针对snap25197的商业mab(mc-6053),和(d)针对snap25197的ab632-rmab;(e–h)来自用媒介物注射并用相同四种抗体探测的对照大鼠爪的共聚焦图像。snap25-ir信号呈绿色(在黑白图中显示为灰色),并且使用dic照明来描绘下面的肌肉纤维(mf)。箭(e)指示来自媒介物处理的大鼠爪的mnt内的ir信号;星号(b、c、f和g)指示肌肉内非特异性ir信号。比例尺=50μm。
图8(a-t)显示免疫组织化学分析,比较了用
图9(a-h)显示用肉毒杆菌毒素a(10u)或媒介物处理之后人背部皮肤切片中针对snap25197的可商购的(mc-6053)mab与ab635-rmab的免疫组织化学比较。用(a,e)mc-6053mab或(b,f)ab635-rmab探测的肉毒杆菌毒素a(a,b)和媒介物处理的(e,f)人皮肤中血管的共聚焦图像。用(c,g)mc-6053mab或(d,h)ab635-rmab探测的肉毒杆菌毒素a(c,d)和媒介物处理的(g,h)人皮肤中的汗腺。箭指示来自媒介物处理的人皮肤的神经纤维内的ir信号。bv,血管;ct,结缔组织;sg,汗腺;比例尺=50μm。
图10(a-f)显示用针对不同形式snap25的抗体探测的bont/a(3nm)或媒介物处理的背根神经节(drg)培养物的共聚焦图像。(a-c)暴露于bont/a3小时并之后用以下抗体染色的drg培养物:(a)针对全长(206)和裂解的(197)snap25的商业(smi-81r)mab,(b)针对snap25197的商业(mc-6053)mab,和(c)针对snap25197的ab632-rmab。(d-f)暴露于媒介物并用相同三种抗体探测的对照drg培养物。e中箭头指示表现出背景标记的神经元胞体。
发明详述
i.定义
除非另外定义,否则本文使用的所有技术和科学术语具有本发明所属领域技术人员通常理解的含义。如本文使用的,除非另外规定,以下术语具有归因于其的含义。
“bont/a”指肉毒杆菌(clostridiumbotulinum)产生的血清型a肉毒杆菌毒素。
“肉毒杆菌毒素a”指商品名
“特异性结合”或“优先结合”(在本文可互换使用)抗体或多肽的表位是本领域公知的术语,并且确定这种特异性或优先结合的方法也是本领域公知的。如果分子与特定细胞或物质比其与另一细胞或物质更频繁、更快速、更持久和以更大亲和力反应或结合,则该分子被称为表现出“特异性结合”或“优先结合”。如果抗体以比其与其他物质更大亲和力、亲合力、更快速、更唯一和/或更持久地结合,则该抗体“特异性结合”或“优先结合”靶标。通过阅读该定义可以理解,例如,特异性或优先结合第一靶标的抗体(或部分或表位)可以或可以不特异性或优先结合第二靶标。因此,“特异性结合”或“优先结合”不一定要求(尽管其可以包括)唯一结合。一般但不一定,本文对结合的提及表示优先结合。
如本文使用的“snap25197”指25kda的突触体结合蛋白(snap25)的197个氨基酸的片段,其全长snap25蛋白由具有血清型a肉毒杆菌毒素的酶(轻链)活性的血清型a肉毒杆菌毒素或重组肉毒杆菌毒素裂解而产生。
如本文使用的“snap25206”指含有206个氨基酸的全长snap25蛋白。
应理解,本发明不限于特定示例的抗体、制剂或使用这种抗体的方法,因此其可以改变。还应理解,本文使用的技术术语仅是为了描述本发明的特定实施方案,而不意图限制。
ii.结合snap25197的抗体
包括突触小泡糖蛋白2c(sv2c)、成纤维细胞生长因子受体3(fgfr3)和snap25206在内的bont/a分子靶标在全身自主和感觉神经纤维中广泛表达和共定位,包括大鼠和灵长类动物膀胱和无毛皮肤。因此,神经末梢可能同样易受bont/a抑制作用的影响。尽管存在针对bont/a的各种分子靶标的抗体,但是还未鉴别用于在蛋白质印迹技术以及组织样品中(例如使用免疫荧光)检测裂解的(即,bont/a活性)snap25(例如,snap25197片段)的最佳抗体。
本公开内容的一些方面部分地包括可用于elisa测定、蛋白质印迹应用、细胞培养测定和组织样品中以检测bont/a活性的对bont/asnap25裂解产物特异的抗snap25抗体。本发明另一方面是具有针对snap25197片段的羧基末端的表位、基本上不结合全长snap25(snap25206)的抗snap25抗体。
制备抗体(单克隆或多克隆)的方法是本领域已知的。可使用的一种方法是kohler和milstein,nature256:495-497(1975)的方法或其修改形式。通常,单克隆抗体在例如小鼠的非人物种中产生。一般,使用小鼠或大鼠来免疫,但也可以使用其他动物。而且,一旦抗体被鉴别为具有所需的结合特征,包括其互补决定区(cdr)的抗原结合位点可以与其他物种(包括人)的抗体的恒定结构域或支持框架区融合。在一些情况下,产生这些重组单克隆抗体可以使在注射这些抗体的患者或宿主动物中不希望的免疫应答最小化。在其他情况下,产生这些重组单克隆抗体可以扩展具有某些结合特征的特异性抗体的效用,用于诊断或用于检测不同物种动物的组织中的bont/a活性。在一些情况下,重组单克隆抗体可以具有最小“脱靶”信号的优势,因为不仅cdr的选择性,而且因为重组igg骨架,其可以显著不同于目标组织的内源性igg。
美国专利公布号us2012/0225436a1中产生并描述了抗snap25197抗体,所述专利公布通过引用并入。对根据其在免疫组织化学(ihc)测定中的优良表现而选择的这些单克隆抗体之一的cdr进行测序并重组工程化至来自人(igg1)或鼠(igg2a)来源的免疫球蛋白骨架。对这些抗体连同可商购的抗体使用免疫荧光进一步表征在蛋白质印迹测定中对裂解的snap25(snap25197)的特异性和用于组织样品(大鼠组织或人类组织)。以下表1包括用于该比较的抗snap25的列表。
n/a=不可用;mab=小鼠单克隆抗体;rmab=重组单克隆抗体;pab=兔多克隆抗体
iii.抗snap25197抗体的表征
可以使用几种方法来表征抗snap25抗体。一种方法是鉴别其结合的表位。表位作图可商购自各种来源,例如pepscansystems(edelhertweg15,8219phlelystad,thenetherlands)。表位作图可用于确定抗snap25抗体结合的序列。表位可以是线性表位,即,单链氨基酸中含有的线性表位,或者是不一定在单链中含有的氨基酸的三维相互作用形成的构象表位。不同长度(例如,至少4-6个氨基酸长)的肽可以是分离的或合成的(例如重组地),并且用于抗snap25抗体的结合测定。抗snap25抗体结合的表位可以通过使用衍生自细胞外序列的重叠肽并测定抗snap25抗体的结合而在系统筛选中确定。
可用于表征抗snap25抗体的另一方法是使用与结合相同抗原或甚至相同抗原上的相同表位的其他抗体的竞争测定。竞争测定是本领域技术人员公知的。
表征抗snap25抗体的另一种方法是通过其结合的抗原。抗snap25抗体可用于蛋白质印迹。具体地,在一些实施方案中,抗snap25抗体用于蛋白质印迹测定以确定其对bont/a裂解的snap25(snap25197)的特异性。在蛋白质印迹测定中优先结合snap25197而非全长(即,未裂解的)snap25的抗snap25抗体是本公开内容的优选实施方案。蛋白质印迹测定中抗snap25抗体的表征在以下实施例中详述。
iv.诊断组织样品中bont/a活性的方法
结合bont/a裂解的snap25(snap25197)的抗体可用于鉴别多种组织中bont/a活性的存在或缺乏。此类组织可以包括皮肤(包括但不限于无毛或有毛的皮肤)、肌肉(包括但不限于骨骼肌和平滑肌)、膀胱、腺组织(包括但不限于前列腺、泪腺、内分泌腺和外分泌腺)、血管、脊髓和脑(包括任何或所有这些组织内的神经纤维)。这种组织可以作为活检或皮肤取样的一部分而获得。
适合用于检测组织中bont/a活性的抗体需要(1)特异性结合snap25197但不结合全长或未裂解的snap25(snap25206);和(2)能够优先检测组织样品中的snap25197(而不是全长或未裂解的snap25)。希望的另一特征是抗体会优先结合组织样品中bont/a裂解的snap25(snap25197)而基本不结合非特异性的抗原(即,低或无背景,非特异性结合)。
确定特定组织样品中bont/a、或bont/a样化合物活性的缺乏或存在可能对于许多临床和非临床诊断目的而言是重要的,包括但不限于:1)理解特定组织或临床适应症中bont/a的作用机制;2)评价bont/a活性和/或注射部位外的扩散;3)评价对bont/a的疑似免疫性;4)评价和理解bont/a的局部扩散(例如,从注射肌肉到邻近肌肉或组织);5)通过活检进行人肉毒杆菌中毒环境下对bont/a的潜在暴露的评价;和6)临床药理学研究。
在一个实施方案中,用途可以包括在snap25197与特异性结合snap25197的抗体之间形成复合物。在本发明的诊断方法的一个实施方案中,抗snap25197抗体可以携带可检测标记。可以使用的标记实例包括放射性物质、荧光团、化学标记、生物剂(例如但不限于生物素/链霉亲和素检测)或酶底物标记。在其他实施方案中,可以携带可检测标记的另一物种的第二抗体可用于检测抗snap25197抗体。第二抗体的使用在一些情况下可以加强第一抗snap25197抗体的信号,从而检测组织样品中低/更低水平的bont/a活性。
v.本发明组合物
本发明还涵盖包含在elisa、蛋白质印迹测定和组织样品中可优先检测bont/a裂解的snap25而不检测未裂解的snap25(snap25206)的抗snap25197抗体的组合物。在一些实施方案中,抗snap25197抗体是其中cdr已经与不同物种的抗体的支持框架区融合的重组单克隆抗体。在其他实施方案中,抗snap25197抗体是其中cdr已经与相同物种的抗体的支持框架区融合的重组单克隆抗体。在一些实施方案中,抗snap25197抗体是其中cdr已经与人抗体的支持框架区融合的重组单克隆抗体。在一些实施方案中,抗snap25197抗体是其中cdr已经与小鼠抗体的支持框架区融合的重组单克隆抗体。
发现之前初步描述于us2012/0225436的一种抗体3c1a5因为其检测bont/a处理的组织中snap25197的能力而特别用于本发明。已知3c1a5在基于免疫的(例如elisa)测定和/或基于细胞的测定中优先结合snap25197。图1显示了snap25197上3c1a5抗体的推定表位结合位点的描述。如实施例中详细描述的,该抗体(及其重组人和鼠形式)在蛋白质印迹测定中也优先检测(或结合)bont/a裂解的snap25(snap25197)。令人惊讶地,该抗体(及其重组人和鼠形式)还能够优先检测(或结合)大鼠和人组织样品中bont/a裂解的snap25(snap25197)。尽管存在能够在蛋白质印迹和组织样品中优先检测(或结合)snap25197的抗裂解snap25抗体的报道,如实施例详细显示的,但3c1a5(或其重组人和鼠形式)是在所有测定和不同组织中始终检测snap25197的唯一抗体,并且不表现出对其他表位的非特异性结合。
在一些情况下,本发明的抗体是3c1a5。在其他情况下,本发明的抗体是包含3c1a5的抗原结合位点的重组抗体,但已经与来自相同或不同物种的另一抗体的框架区融合。
在一个实施方案中,抗snap25197抗体的轻链/重链包含以下表2所列序列中的一个或多个。
在一些情况下,本发明的抗体包含seqidno:1或seidno:2的轻链序列,并且是重组鼠抗体。在其他情况下,本发明的抗体包含seqidno:3或seqidno:4的重链序列,并且是重组鼠抗体。在其他情况下,本发明的抗体包含seqidno:5或seqidno:6的轻链序列,并且是重组人抗体。在其他情况下,本发明的抗体包含seqidno:7或seqidno:8的重链序列,并且是重组人抗体。
在其他实施方案中,本发明的抗体可以是结合具有包含seqidno:1-8的一个或多个序列的重链和/或轻链的抗体的相同表位的抗体。在一些情况下,本发明的抗体可以是竞争结合具有包含seqidno:1-8的一个或多个序列的重链和/或轻链的抗体的相同表位的抗体。
提供以下实施例以示例说明,但不限制本发明。
iv.实施例
实施例1.蛋白质印迹比较
首先通过蛋白质印迹分析比较了snap25抗体(表1中所列)识别来自用和未用bont/a处理的大鼠胚胎皮层细胞裂解物和sima细胞裂解物的全长(206)或bont/a裂解的(197)形式的snap25的能力(参见图2和3)。
从胚胎幼崽(e18)收获大鼠皮层神经元并在木瓜蛋白酶解离系统(worthingtonbiochemicalcorp.,lakewood,nj)中于37℃下消化15分钟以获得单个细胞。然后将皮层细胞转移至含有b-27补充物、0.5mml-谷氨酰胺和青霉素/链霉素的neurobasal培养基(lifetechnologies,carlsbad,ca)。汇集从新生幼崽(p7-p14)收获的大鼠背根神经节(drg)并在含有木瓜蛋白酶的hbss(1mml-半胱氨酸中每ml20单位木瓜蛋白酶的终浓度)中于37℃下消化15分钟。洗涤神经节经并随后在不含ca2+/mg2+、含有1型胶原酶(1.7mg/ml,sigma,stlouis,mo)的hbss中消化,并在37℃下孵育另外15分钟。然后在含有b-27补充物、0.5mml-谷氨酰胺、青霉素/链霉素和20ng/ml2.5s神经生长因子(ngf)的neurobasal-a培养基(lifetechnologies,carlsbad,ca)中洗涤神经节,并通过pasteur移液管温和磨碎。均匀分散皮层和drg细胞,铺板至放置在100-mm培养皿中的聚d-赖氨酸/层粘连蛋白涂布的12-mm盖玻片(bdbiosciences,sanjose,ca)并使其生长6至7div,然后处理。所有动物方案和程序经allergan动物管理和使用委员会(allerganinstitutionalanimalcareandusecommittee)批准并根据nih指南操作。
在选择日,培养物用或不用3nmbont/a(150kda;metabiologics,madison,wi)于37℃下处理3小时。处理之后,冲洗细胞然后在新鲜培养基中孵育过夜。然后用pbs洗涤皮层细胞,在新鲜制备的裂解缓冲液(20mmtrisph7.5,0.15m氯化钠,1nmedta,1mmegta,10%tritonx-100和一片无edta的蛋白酶抑制剂)中在冰上裂解20分钟,然后以4000rpm离心20分钟以在蛋白质印迹(wb)分析之前消除碎片。drg细胞经洗涤,用4%低聚甲醛固定10-15分钟并根据以下方案针对免疫细胞化学进行处理。
sima细胞(dsmz,germany)在具有通气帽的bdbiosciences品牌collageniv烧瓶(vwr,radnor,pa)中培养(marini,p.,macleod,r.a.,treuner,c.,bruchelt,g.,bohm,w.,wolburg,h.,schweizer,p.,和girgert,r.,1999.sima,anewneuroblastomacelllinecombiningpoorprognosticcytogeneticmarkerswithhighadrenergicdifferentiation.cancergenet.cytogenet.112,161-164.)。生长培养基由rpmi1640、0.1mm非必需氨基酸、10mmhepes、1mm丙酮酸钠、100u/ml青霉素、100μg/ml链霉素、和10%胎牛血清组成。细胞用或不用bont/a(0.01nm)于37℃下处理24小时。处理之后,sima细胞用pbs洗涤,在新鲜制备的裂解缓冲液中在冰上裂解20分钟,然后以4000rpm离心20分钟以在wb分析之前消除碎片。
对于wb测定,我们采用了大鼠胚胎皮层神经元以及人成神经细胞瘤细胞系(sima),其因为其对bont/a介导的snap25裂解的敏感性而被周知(fernandez-salas,e.,wang,j.,molina,y.,nelson,j.b.,jacky,b.p.,和aoki,k.r.,2012.botulinumneurotoxinserotypeaspecificcell-basedpotencyassaytoreplacethemousebioassay.plos.one.7,e49516.)。通过电泳(bioradtgxanykd凝胶)从这些培养物分离总细胞裂解物,并且将凝胶转移至pvdf膜上。将印迹在缓冲液(1xtbs-0.1%吐温-20中5%奶粉)中于室温下封闭1小时,然后在4℃下与一抗在封闭缓冲液中孵育过夜。洗涤之后,将印迹与hrp缀合的二抗(biorad,hercules,ca)孵育,并通过eclplus(gehealthcare,pittsburgh,pa)显色。将单独的对照印迹探测甘油醛3-磷酸脱氢酶(gapdh)以显示细胞裂解物样品的相等负载。使用可变模式getyphoon9410成像仪扫描蛋白质印迹并用imagequanttlv.2005软件(gehealthcare,pittsburgh,pa)分析。
首先,针对所有形式snap25蛋白的可商购且广泛使用的单克隆抗体(smi-81r)识别snap25206和snap25197两者(图2a)。相反,被描述为识别两种形式的snap25的第二种可商购的单克隆抗体(mc-6050)令人惊讶地特异性针对来自毒素处理的细胞的裂解物中的snap25197(图2b)。而且,来自相同公司的被描述为仅识别bont/a裂解的snap25的另一抗体(mc-6053)在毒素处理的泳道中唯一地显示了细而模糊的条带(图2c)。
针对bont/a裂解的snap25的人(ab632)和鼠(ab635)rmab非常特异性针对snap25197;在毒素处理的裂解物中仅检测到单一条带,而在未处理的对照泳道中没有检测到条带(图2d,e)。相似地,我们内部的针对snap25197的兔pab(rgt-1092)仅检测到bont/a处理样品中一个条带,而在对照泳道中没有检测到条带(图2f)。然而,应注意,pab识别两个额外的模糊的条带,一个刚好在snap25197条带之上,并且另一个在仅存在于毒素处理样品中的约20kda处,这不太好解释。然而,上面的条带不大可能是完整的snap25,因为在未处理泳道中没有观察到条带。使用来自bont/a处理和未处理的sima细胞培养物的裂解物获得了相似的wb结果(图3),除了在对照或毒素处理泳道中没有观察到mc-6053的条带(图3c)。对于皮层和sima细胞裂解物实验,针对gapdh探测的对照印迹显示了所有样品的相等负载(图4a,b)。
使用bont/c和bont/e处理的sima细胞裂解物进行单独的wb分析以检查我们rmab的表位特异性,与bont/a处理的裂解物比较。公知bont/c在氨基酸(aa)残基198处裂解snap25,而bont/e在aa残基180处裂解snap25[11]。虽然smi-81rmab识别全长和bont裂解形式的snap25(图4c),但如预期的,我们的人rmab仅检测到bont/a处理的裂解样品中的单一条带(图4d)。
图2显示了比较针对snap25的抗体的特异性的wb分析,使用了用(l3)或未用(l2)bont/a处理的大鼠胚胎皮层神经元细胞裂解物。(a)用识别全长(206)和裂解(197)形式的snap25的可商购的抗snap25mab(smi-81r)探测的印迹。在泳道2中,仅检测到snap25206,而在泳道3中,检测到snap25206(箭)和snap25197(箭头)两者。(b)用据报道识别snap25206和snap25197两者的可商购的抗snap25mab(mc-6050)探测的印迹。仅snap25197作为泳道3中的单一条带出现(箭头)。(c)用据报道对snap25197特异的可商购的抗snap25mab(mc-6053)探测的印迹。该抗体识别泳道3中细的模糊snap25197条带。(d)用ab632抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(e)用ab635抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(f)用rgt-1092抗snap25197pab探测的印迹。该抗体主要识别泳道3中的snap25197(箭头),但是仅在snap25197条带上下可见两个模糊条带。泳道1,蛋白质分子量标准;泳道2,未处理的皮层细胞裂解物;泳道3,bont/a处理的(3nm)皮层细胞裂解物。
图3显示了比较针对snap25的抗体的特异性的wb分析,使用了用(l3)或未用(l2)bont/a处理的sima细胞裂解物。(a)用识别全长(206)和裂解(197)形式的snap25的可商购的抗snap25mab(smi-81r)探测的印迹。在泳道2中,仅检测到snap25206,而在泳道3,检测到snap25206(箭)和snap25197(箭头)两者。(b)用据报道识别snap25206和snap25197两者的可商购的抗snap25mab(mc-6050)探测的印迹。仅snap25197作为泳道3中的单一条带出现(箭头)。(c)用据报道对snap25197特异的可商购的抗snap25mab(mc-6053)探测的印迹。该抗体没有显示识别任何条带。(d)用ab632抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(e)用ab635抗snap25197rmab探测的印迹。在泳道2中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带(箭头)。(f)用rgt-1092抗snap25197pab探测的印迹。该抗体主要识别泳道3中的snap25197(箭头),但是仅在snap25197条带上下可见两个模糊条带。泳道1,蛋白质分子量标准;泳道2,未处理的sima细胞裂解物;泳道3,bont/a处理的(0.01nm)sima细胞裂解物。
图4(a,b)显示用抗gapdhmab探测的图2中皮层细胞研究和图3中sima细胞研究的对照印迹,显示了泳道2和3中的相等的样品负载。泳道1,蛋白质分子量标准;泳道2,未处理的细胞裂解物;泳道3,bont/a处理的(3nm)皮层细胞裂解物和(0.01nm)sima细胞裂解物。图4(c,d)显示证明ab632-rmab的表位特异性的蛋白质印迹分析,使用了经bont/a(l3)、bont/c(l4)、bont/e(l5)或没有毒素(l2)处理的sima细胞裂解物。(c)用识别全长(206)和裂解(bont/a的197,bont/c的198,和bont/e的180)形式snap25的可商购的抗snap25mab(smi-81r)探测的印迹。在泳道2中,仅检测到snap25206,而在泳道3,检测到snap25206(箭)和snap25197两者。在泳道4中,检测到snap25206(箭)和snap25198两者,并且在泳道5中,检测到snap25206(箭)和snap25180两者;(d)用ab632抗snap25197rmab探测的印迹。在泳道2、4和5中,没有检测到条带,而在泳道3中,检测到snap25197的单一条带。泳道1,蛋白质分子量标准;泳道2,未处理的sima细胞裂解物;泳道3,bont/a处理的sima细胞裂解物;泳道4,bont/c处理的sima细胞裂解物;泳道5,bont/e处理的sima细胞裂解物。
实施例2.免疫组织化学比较—大鼠组织
然后,使用免疫组织化学(ihc)在已经肉毒杆菌毒素a或盐水处理并在注射后两天收获的大鼠膀胱和无毛皮肤中测试抗体的特异性。在这些ihc研究中,相邻的无一抗处理的切片仅显示背景染色(数据未显示)。
雄性sprague-dawley大鼠(charlesriverlaboratories,wilmington,ma)用于该研究。大鼠成对圈养在rd3饲养所,自由获取食物和水,12小时光-暗循环。所有程序经allergan动物管理和使用委员会批准并符合nih指南。
分别在0.9%盐水或0.5%bsa/0.9%盐水中制备肉毒杆菌毒素a(
大鼠在注射后两天被处死,并收获膀胱或后爪种跖表面(plantersurface)中心部分并在zamboni固定剂(americanmastertech,lodi,ca)中于4℃下下固定过夜。洗涤组织并在30%蔗糖/pbs溶液中于4℃下低温保存过夜。将膀胱和皮肤样品沿中线切成对半,包埋在o.c.t.(tissue-tek)中并在-80℃下冷冻储存直至切片。组织块经冷冻切片(14μm厚),载玻片固定并将载玻片保持在-20℃下直至使用。
首先在封闭缓冲液(1xpbs+0.1%tritonx-100+10%正常驴血清)中封闭载玻片固定的组织切片和细胞培养盖玻片的非特异性信号,然后与所需浓度的一抗在封闭缓冲液中于4℃下孵育过夜。几次洗涤之后,将切片与盖玻片与在封闭缓冲液中稀释的二抗(jacksonimmunoresearch,westgrove,pa)一起在4℃下孵育2小时,然后再次洗涤。将具有培养物的盖玻片倒置并使用含有1.5μg/mldapi的fluoromount-g(emsciences,hatfield,pa)固定至显微镜载玻片。使用相同的固定介质将载玻片固定的切片加盖玻片。没有用一抗处理的相邻切片用作阴性对照以显示背景信号。将交替切片用苏木精&伊红染色以求更好的解剖学鉴别。
使用具有zen软件的zeisslsm-710共聚焦显微镜(carlzeiss,thornwood,ny)或olympusfv1000共聚焦显微镜(olympus,centervalley,pa)捕获和分析图像。使用
在大鼠膀胱中,针对snap25的smi-81r抗体在贯穿逼尿肌的神经纤维中显示了ir信号。如期望的,snap25-ir模式在肉毒杆菌毒素a和盐水处理的膀胱中都是相同的(图5a,f)。据报道识别完整和bont/a裂解形式snap25的mc-6050单克隆抗体在毒素处理而非盐水处理的膀胱的逼尿肌的神经纤维中显示了的ir信号(图5b,g),与来自蛋白质印迹分析的结果相当。同样,mc-6053单克隆抗体仅在毒素处理而非盐水处理的膀胱中显示了ir信号(图5c,h)。与wb分析中检测的特异性相反,针对snap25197产生的rgt-1092pab在毒素处理和盐水处理的膀胱中都显示了ir信号(图5e,j)。更重要的是,ab632-rmab在毒素处理的膀胱的逼尿肌的神经纤维中显示了清晰的ir信号(图5d),而在盐水处理的对照膀胱中没有检测到信号(图5i)。而且,在单独的研究中,ab635-rmab在大鼠膀胱中显示了与ab632-rmab相似的对snap25197的特异性(图8m,r))。
在大鼠无毛皮肤中,smi-81r抗体表现出在血管周围(在其他皮肤区域中)神经纤维中的ir信号。如针对该抗体所预期的,snap25-ir模式在肉毒杆菌毒素a和盐水处理的皮肤中都是相同的(图6a,f)。mc-6050单克隆抗体主要在毒素处理的皮肤的神经纤维中显示了ir信号。然而,在盐水处理的皮肤的神经纤维中也可见微弱的ir信号(图6b,g)。mc-6050抗体还在血管管腔中显示了强ir信号(图6b,g)。但是因为该特定ir信号从未得到其他snap25抗体的证明,所以确定其是非特异性的。相似地,mc-6053单克隆抗体在毒素和盐水处理的皮肤的血管周围的神经纤维中都显示了ir特异性信号,以及在血管管腔中的非特异性信号(图6c,h)。再次,针对snap25197产生的rgt-1092pab在毒素处理和盐水处理的大鼠皮肤中都显示了ir信号(图6e,j),表明不论其在wb分析中的特异性,该抗体不适用ihc。形成明显对比,ab632-rmab仅在毒素处理而非盐水处理的皮肤的神经纤维中显示ir信号(图6d,i),支持其极好的特异性。
我们的大鼠无毛皮肤的样品经常含有下面的骨骼肌,从而提供了良好机会以验证我们的rmab识别运动神经末梢(mnt)内bont/a裂解的snap25的能力。在mnt中观察到与其他皮肤神经纤维类型相似的抗体特异性的结果(图7)。虽然smi-81rmab识别mnt和轴突中的全长和bont/a裂解形式snap25,但是商业mab,mc-6050和mc-6053显示了主要在毒素处理皮肤的神经纤维中的ir信号。然而,对于这些商业mab,在盐水处理组织中还观察到了非特异性的ir信号(图7f,g)。相反,ab632-rmab仅在毒素处理而非盐水处理的皮肤中表现出在mnt和轴突中的ir信号(图7d,h)。
为了进一步例示ab632-rmab对组织中snap25197的优良特异性,我们比较了ab632和ab635与初始批号人rmab(5/02/2011)、从腹水纯化的原始的天然3c1a5mab和用于allergan的基于细胞的
在大鼠无毛皮肤中,ab632和ab635显示了在
在
图5显示免疫组织化学分析,比较了针对用肉毒杆菌毒素a(10u/kg)或媒介物处理之后大鼠膀胱切片中的snap25的抗体的特异性。(a-e)用肉毒杆菌毒素a注射并用以下抗体探测的膀胱逼尿肌(dm)的共聚焦图像:(a)针对全长(206)和裂解的(197)snap25的商业mab(smi-81r),(b)针对snap25197和snap25206的第二商业mab(mc-6050),(c)针对snap25197的商业mab(mc-6053),(d)针对snap25197的ab632-rmab,和(e)针对snap25197的rgt-1092pab。(f-j)用媒介物注射和用相同的五种抗体探测的对照大鼠膀胱的共聚焦图像。箭(f和j)指示来自媒介物处理的膀胱的神经纤维内的ir信号。dm,逼尿肌;比例尺=50μm。
图6显示免疫组织化学分析,比较了针对用肉毒杆菌毒素a(30u/kg)或媒介物处理之后大鼠无毛皮肤切片中的snap25的抗体的特异性。(a-e)用肉毒杆菌毒素a注射并用以下抗体探测的大鼠皮肤的共聚焦图像:(a)针对全长(206)和裂解的(197)snap25的商业mab(smi-81r),(b)针对snap25197和snap25206的第二商业mab(mc-6050),(c)针对snap25197的商业mab(mc-6053),(d)针对snap25197的ab632-rmab,和(e)针对snap25197的rgt-1092pab。(f-j)用媒介物注射和用相同的五种抗体探测的对照大鼠皮肤的共聚焦图像。箭(f、g、h和j)指示来自媒介物处理大鼠皮肤的神经纤维内的ir信号。星号(b、c、g和h)指示血管腔内非特异性ir信号。bv,血管;ct,结缔组织;比例尺=50μm。
图7显示免疫组织化学分析,比较了针对用肉毒杆菌毒素a(30u/kg)或媒介物处理之后大鼠无毛皮肤下骨骼肌中的snap25的抗体的特异性。(a–d)显示用肉毒杆菌毒素a注射并用以下抗体探测的大鼠爪的下面肌肉内运动神经末梢(mnt,箭)的共聚焦图像:(a)针对全长(206)和裂解的(197)snap25的商业mab(smi-81r),(b)针对snap25197和snap25206的第二商业mab(mc-6050),(c)针对snap25197的商业mab(mc-6053),和(d)针对snap25197的ab632-rmab;(e–h)来自用媒介物注射并用相同四种抗体探测的对照大鼠爪的共聚焦图像。snap25-ir信号呈绿色(在黑白图中显示为灰色),并且使用dic照明来描绘下面的肌肉纤维(mf)。箭(e)指示来自媒介物处理的大鼠爪的mnt内的ir信号;星号(b、c、f和g)指示肌肉内非特异性ir信号。比例尺=50μm。
图8显示免疫组织化学分析,比较了用
实施例3.免疫组织化学比较—人组织
在针对snap25的可商购的抗体中,靶向snap25197的mc-6053单克隆抗体在抗体类型和物种方面与我们的鼠ab635-rmab最相似(表1)。因此,我们在肉毒杆菌毒素a和盐水处理的人背部皮肤的活检样品中进行了我们ab635-rmab与mc-6053mab之间的snap25-ir表达模式的头对头比较(head-to-headcomparison)。推测是因为这是使用小鼠抗体的人组织的探针,所以非特异性ir信号(不论其来源)将是最小的。
通过1期allergan临床研究获得了人皮肤活检样品。根据良好临床实践的指南和规定以及所有相关的地方和国家隐私指南进行研究。研究方案、知情同意和所有适当的研究相关文件由机构审查委员会/伦理委员会批准。
在成人背部皮肤id注射10u肉毒杆菌毒素a或媒介物。处理后14天从背部皮肤收获取样的活检样品并在相同固定剂中固定过夜。洗涤组织并在30%蔗糖/pbs溶液中于4℃下低温保存过夜。将皮肤样品沿中线切成对半,包埋在o.c.t.(tissue-tek)中并在-80℃下冷冻储存直至切片。组织块经冷冻切片(14μm厚),载玻片固定并将载玻片保持在-20℃下直至使用。如上所述进行ihc和数据分析。
在人背部皮肤中,在血管周围神经纤维以及皮肤内的汗腺中观察到了这两种抗体的ir信号(图9)。在来自肉毒杆菌毒素a和盐水处理的背部皮肤的神经纤维中观察到了mc-6053mab的ir信号。形成明显对比,仅在来自肉毒杆菌毒素a处理的而非盐水处理的人背部皮肤的神经纤维中观察到了我们的鼠ab635-rmab的ir信号(图9),证明了其优良的特异性,并且更重要的是其作为临床诊断工具的效用。
图9显示用肉毒杆菌毒素a(10u)或媒介物处理之后人背部皮肤切片中针对snap25197的可商购的(mc-6053)mab与ab635-rmab的免疫组织化学比较。用(a,e)mc-6053mab或(b,f)ab635-rmab探测的肉毒杆菌毒素a(a,b)和媒介物处理的(e,f)人皮肤中血管的共聚焦图像。用(c,g)mc-6053mab或(d,h)ab635-rmab探测的肉毒杆菌毒素a(c,d)和媒介物处理的(g,h)人皮肤中的汗腺。箭指示来自媒介物处理的人皮肤的神经纤维内的ir信号。bv,血管;ct,结缔组织;sg,汗腺;比例尺=50μm。
考虑到检测细胞内bont/a定位和移动的难度,专有的重组人源化α-snap25197和专有的重组鼠α-snap25197可用于交叉检测其他物种中的snap25197。虽然其他α-snap25抗体能够检测,使用重组鼠α-snap25197检测人组织中的snap25197或使用重组人源化α-snap25197检测鼠中的snap25197允许深度分析bont/a的作用机制,这是其他可用抗体所不能的。
实施例4.免疫细胞化学比较
一些抗体针对一个测定/适应症可以比另一抗体作用更好。因此,为了完成我们的分析,我们比较了经bont/a(3nm)或盐水处理的drg细胞培养物中几个抗体的ir信号。
如上所述准备和处理drg细胞培养物。如上详细描述地进行免疫细胞化学和数据分析。
如在组织中的,smi-81r抗体在bont/a和盐水处理的培养物中都显示了强的ir信号(图10a,d)。mc-6053可商购的mab和我们的人ab632-rmab显示了来自bont/a处理的培养物的神经元细胞中的特异性snap25197-ir信号(图10b,c)。在盐水处理的培养物中没有检测到信号(图10e,f)。然而,mc-6053mab表现出盐水处理的培养物中神经元胞体的微弱背景信号(图10e)。
图10显示用针对不同形式snap25的抗体探测的bont/a(3nm)或媒介物处理的背根神经节(drg)培养物的共聚焦图像。(a-c)暴露于bont/a3小时并之后用以下抗体染色的drg培养物:(a)针对全长(206)和裂解的(197)snap25的商业(smi-81r)mab,(b)针对snap25197的商业(mc-6053)mab,和(c)针对snap25197的ab632-rmab。(d-f)暴露于媒介物并用相同三种抗体探测的对照drg培养物。e中箭头指示表现出背景标记的神经元胞体。
经常通过使用针对裂解底物(snap25197)的选择性抗体来确定表达snap25的细胞中活性bont/a的存在。在本研究中,我们引入了内部开发的针对snap25197的几种rmab,并且使用多种不同方法比较了其与商业抗体的免疫反应性信号(表3)。我们的人和鼠rmab在所有测定和不同组织中始终检测到snap25197,并且如预期的,没有检测到全长snap25(snap25206)。使用表现出可变的测定依赖性特异性的其他宣称的snap25197选择性抗体则不是这样。这些结果证实bont/a裂解的snap25表位难以被抗体特异性靶向,也不识别完整snap25蛋白,如果没有适当对照,其可以导致结果的潜在误解。因此,任何给定snap25197抗体应该在多种条件和组织类型下测试以确保其在检测bont/a裂解的snap25存在时的保真性。
位点特异性抗体日益用于体外和体内分析。这些抗体可以检测磷酸化位点或酶促裂解位点,并且是我们理解蛋白质成熟、活性和降解的有价值的工具(mort,j.s.和buttle,d.j.,1999.theuseofcleavagesitespecificantibodiestodelineateproteinprocessingandbreakdownpathways.mol.pathol.52,11-18.;mort,j.s.,flannery,c.r.,makkerh,j.,krupa,j.c.,和lee,e.r.,2003.useofanti-neoepitopeantibodiesfortheanalysisofdegradativeeventsincartilageandthemolecularbasisforneoepitopespecificity.biochem.soc.symp.107-114.;nagata,k.,izawa,i.,和inagaki,m.,2001.adecadeofsite-andphosphorylationstate-specificantibodies:recentadvancesinstudiesofspatiotemporalproteinphosphorylation.genescells6,653-664.)。在肉毒杆菌神经毒素领域,裂解位点特异性抗体可以帮助检测小量bont轻链的活性,否则其可能极难察觉。为此,多克隆抗体的使用可能具有有限的价值,因为可能生成混合的免疫球蛋白群,不是所有都具有裂解表位所需的特异性(mort,j.s.和buttle,d.j.,1999.theuseofcleavagesitespecificantibodiestodelineateproteinprocessingandbreakdownpathways.mol.pathol.52,11-18.)。而且,肽抗原应该相对短,以减少产生结合远离靶表位的序列部分的抗体的可能性。
目前研究中提供的抗snap25197mab经初始筛选并根据其在ihc测定中的优良性能来选择。随后从该抗体产生的rmab(ab632和ab635)在几种不同测定(包括ihc)中证明了对bont/a裂解的snap25的极好的特异性。而且,这些rmab与其他测试抗体相比显示了更好的snap25197特异性(表3)。因此,我们的rmab代表了用于检测细胞内和临床样品中bont/a活性的有效的新工具,并且将在未来研究中用于表征感兴趣组织的bont/a的功效。这些抗体的特定序列提供于上表2。
总而言之,在蛋白质印迹测定、对组织进行的免疫组织化学和对细胞进行的免疫细胞化学中测试的抗体显示了各种针对bont/a裂解的snap25(snap25197)和未裂解的snap25(snap25206)的特异性。结果概括显示于下表3。
n/t=未测试;b=背景
要理解,本文描述的实施例和实施方案仅用于示例说明目的,并且根据其的各种修改或变化将暗示给本领域技术人员并且包括在本申请的精神和范围内。本文引用的所有出版物、专利和专利申请通过引用以其整体出于所有目的并入本文,其程度如同每一个单独出版物、专利或专利申请明确且单独地指示如此通过引用并入一样。
序列表
<110>broide,rons.
cai,brian
fernandez-salas,ester
francis,joseph
rheaume,catherine
<120>检测组织样品中裂解的snap25的方法
<130>19438(ntb)
<140>14/
<141>2015-07-07
<160>9
<170>patentin3.5版
<210>1
<211>219
<212>prt
<213>小鼠属种
<400>1
aspvalvalmetthrglnthrproleuthrleuservalthrilegly
151015
glnproalaserilesercyslysserserglnserleuleuasnthr
202530
asnglylysthrtyrleuthrtrpleuileglnargproglyglnser
354045
proglnargleuiletyrleuvalserlysleuaspserglyvalpro
505560
aspargpheserglyserglyserglythraspphethrleulysile
65707580
serargvalglualagluaspleuglyvaltyrtyrcysleuglnser
859095
serhispheprophethrpheglyserglythrlysleugluilelys
100105110
argalaaspalaalaprothrvalserilepheproproserserglu
115120125
glnleuthrserglyglyalaservalvalcyspheleuasnasnphe
130135140
tyrprolysaspileasnvallystrplysileaspglysergluarg
145150155160
glnasnglyvalleuasnsertrpthraspglnaspserlysaspser
165170175
thrtyrsermetserserthrleuthrleuthrlysaspglutyrglu
180185190
arghisasnsertyrthrcysglualathrhislysthrserthrser
195200205
proilevallysserpheasnargasnglucys
210215
<210>2
<211>112
<212>prt
<213>小鼠属种
<400>2
aspvalvalmetthrglnthrproleuthrleuservalthrilegly
151015
glnproalaserilesercyslysserserglnserleuleuasnthr
202530
asnglylysthrtyrleuthrtrpleuileglnargproglyglnser
354045
proglnargleuiletyrleuvalserlysleuaspserglyvalpro
505560
aspargpheserglyserglyserglythraspphethrleulysile
65707580
serargvalglualagluaspleuglyvaltyrtyrcysleuglnser
859095
serhispheprophethrpheglyserglythrlysleugluilelys
100105110
<210>3
<211>436
<212>prt
<213>小鼠属种
<400>3
glnvallysleuglngluserglyalagluleuvallysproglyala
151015
servallysilesercyslysalaserglytyrthrphethrasphis
202530
serilehistrpvallysglnlysproglyglnglyleuglutrpile
354045
glytyrleupheproglyasnglyasnpheglutyrasnglulysphe
505560
lysglylysalathrleuthralaasplysserserserthrvaltyr
65707580
mettyrleuasnserleuthrsergluaspseralavaltyrphecys
859095
lysargmetglytyrtrpglyglnglythrthrvalthrvalserser
100105110
alalysthrthrproproservaltyrproleualaproglyserala
115120125
alaglnthrasnsermetvalthrleuglycysleuvallysglytyr
130135140
pheprogluprovalthrvalthrtrpasnserglyserleuserser
145150155160
glyvalhisthrpheproalavalleugluseraspleutyrthrleu
165170175
serserservalthrvalproserserproargprosergluthrval
180185190
thrcysasnvalalahisproalaserserthrlysvalasplyslys
195200205
ilevalproargaspcysglycyslysprocysilecysthrvalpro
210215220
gluvalserservalpheilepheproprolysprolysaspvalleu
225230235240
thrilethrleuthrprolysvalthrcysvalvalvalaspileser
245250255
lysaspaspprogluvalglnphesertrpphevalaspaspvalglu
260265270
valhisthralaglnthrglnproargglugluglnpheasnserthr
275280285
pheargservalsergluleuproilemethisglnasptrpleuasn
290295300
glylysgluphelyscysargvalasnseralaalapheproalapro
305310315320
ileglulysthrileserlysthrlysglyargprolysalaprogln
325330335
valtyrthrileproproprolysgluglnmetalalysasplysval
340345350
serleuthrcysmetilethraspphepheprogluaspilethrval
355360365
glutrpglntrpasnglyglnproalagluasntyrlysasnthrgln
370375380
proilemetasnthrasnglysertyrphevaltyrserlysleuasn
385390395400
valglnlysserasntrpglualaglyasnthrphethrcysserval
405410415
leuhisgluglyleuhisasnhishisthrglulysserleuserhis
420425430
serproglylys
435
<210>4
<211>112
<212>prt
<213>小鼠属种
<400>4
glnvallysleuglngluserglyalagluleuvallysproglyala
151015
servallysilesercyslysalaserglytyrthrphethrasphis
202530
serilehistrpvallysglnlysproglyglnglyleuglutrpile
354045
glytyrleupheproglyasnglyasnpheglutyrasnglulysphe
505560
lysglylysalathrleuthralaasplysserserserthrvaltyr
65707580
mettyrleuasnserleuthrsergluaspseralavaltyrphecys
859095
lysargmetglytyrtrpglyglnglythrthrvalthrvalserser
100105110
<210>5
<211>219
<212>prt
<213>智人
<400>5
aspvalvalmetthrglnthrproleuthrleuservalthrilegly
151015
glnproalaserilesercyslysserserglnserleuleuasnthr
202530
asnglylysthrtyrleuthrtrpleuileglnargproglyglnser
354045
proglnargleuiletyrleuvalserlysleuaspserglyvalpro
505560
aspargpheserglyserglyserglythraspphethrleulysile
65707580
serargvalglualagluaspleuglyvaltyrtyrcysleuglnser
859095
serhispheprophethrpheglyserglythrlysleugluilelys
100105110
argthrvalalaalaproservalpheilepheproproseraspglu
115120125
glnleulysserglythralaservalvalcysleuleuasnasnphe
130135140
tyrproargglualalysvalglntrplysvalaspasnalaleugln
145150155160
serglyasnserglngluservalthrgluglnaspserlysaspser
165170175
thrtyrserleuserserthrleuthrleuserlysalaasptyrglu
180185190
lyshislysvaltyralacysgluvalthrhisglnglyleuserser
195200205
provalthrlysserpheasnargglyglucys
210215
aspvalvalmetthrglnthrproleuthrleuservalthrilegly
151015
glnproalaserilesercyslysserserglnserleuleuasnthr
202530
asnglylysthrtyrleuthrtrpleuileglnargproglyglnser
354045
proglnargleuiletyrleuvalserlysleuaspserglyvalpro
505560
aspargpheserglyserglyserglythraspphethrleulysile
65707580
serargvalglualagluaspleuglyvaltyrtyrcysleuglnser
859095
serhispheprophethrpheglyserglythrlysleugluilelys
100105110
<210>7
<211>442
<212>prt
<213>智人
<400>7
glnvallysleuglngluserglyalagluleuvallysproglyala
151015
servallysilesercyslysalaserglytyrthrphethrasphis
202530
serilehistrpvallysglnlysproglyglnglyleuglutrpile
354045
glytyrleupheproglyasnglyasnpheglutyrasnglulysphe
505560
lysglylysalathrleuthralaasplysserserserthrvaltyr
65707580
mettyrleuasnserleuthrsergluaspseralavaltyrphecys
859095
lysargmetglytyrtrpglyglnglythrthrvalthrvalserser
100105110
alaserthrlysglyproservalpheproleualaproserserlys
115120125
serthrserglyglythralaalaleuglycysleuvallysasptyr
130135140
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
145150155160
glyvalhisthrpheproalavalleuglnserserglyleutyrser
165170175
leuserservalvalthrvalproserserserleuglythrglnthr
180185190
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
195200205
lysvalgluprolyssercysasplysthrhisthrcysproprocys
210215220
proalaprogluleuleuglyglyproservalpheleuphepropro
225230235240
lysprolysaspthrleumetileserargthrprogluvalthrcys
245250255
valvalvalaspvalserhisgluaspprogluvallyspheasntrp
260265270
tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu
275280285
gluglntyrasnserthrtyrargvalvalservalleuthrvalleu
290295300
hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn
305310315320
lysalaleuproalaproileglulysthrileserlysalalysgly
325330335
glnproarggluproglnvaltyrthrleuproproserargaspglu
340345350
leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr
355360365
proseraspilealavalglutrpgluserasnglyglnprogluasn
370375380
asntyrlysthrthrproprovalleuaspseraspglyserphephe
385390395400
leutyrserlysleuthrvalasplysserargtrpglnglnglyasn
405410415
valphesercysservalmethisglualaleuhisasnhistyrthr
420425430
glnlysserleuserleuserproglylys
435440
<210>8
<211>112
<212>prt
<213>智人
<400>8
glnvallysleuglngluserglyalagluleuvallysproglyala
151015
servallysilesercyslysalaserglytyrthrphethrasphis
202530
serilehistrpvallysglnlysproglyglnglyleuglutrpile
354045
glytyrleupheproglyasnglyasnpheglutyrasnglulysphe
505560
lysglylysalathrleuthralaasplysserserserthrvaltyr
65707580
mettyrleuasnserleuthrsergluaspseralavaltyrphecys
859095
lysargmetglytyrtrpglyglnglythrthrvalthrvalserser
100105110
<210>9
<211>12
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>9
aspserasnlysthrargileaspglualaasngln
1510
- 还没有人留言评论。精彩留言会获得点赞!