一种用于微拟球藻基因沉默的RNAi载体及其应用的制作方法
本发明属于微生物基因工程技术领域,具体涉及公开一种用于微拟球藻基因沉默的rnai载体及其应用。
背景技术:
微藻是一个具有高度多样性的生物群体,广泛分布于海洋、胡泊、陆地等各种栖息环境,也是地球上最古老的生命类群之一。它们不仅在生态系统中扮演着重要角色,而且在进化关系上也跨越原核和真核生物两界。因此,它们拥有相对独特的进化地位,微藻进化研究一直是科学研究的热点问题。由于微藻进化历史和栖息环境的多样性和复杂性,不同的藻类在代谢途径、细胞发育调控以及生活史等发面都有差异及其独特性,而我们对其认识是非常有限的。另外,随着全球化石能源日益短缺,以及现代工农业的发展带来的全球气候变暖等问题,发展清洁、可再生型新能源已成为必然趋势,微藻生物能源就是其中被寄予厚望的救世主,同时微藻也能够用于生产多种高附加值化学品(如多糖、多不饱和脂肪酸和虾青素),广泛用于食品、保健品和化妆品等方面工业应用。微藻生物学功能的研究将有助于我们理解微藻遗传资源和加以利用,这些都迫切需要人类去认识微藻及改造微藻。
随着高通量基因测序的发展,许多藻类的基因组已经被测序,包括莱茵衣藻、硅藻、微拟球藻、小球藻、团藻、褐藻和红藻等。尽管对其开展全基因组测序,但是它们基因绝大多数(占30%以上)都是未知功能的,对于代谢过程的理解仍然是一个“黑箱”。这就需要我们发展遗传转化方法和基因操作工具去揭开这个“黑箱”之谜。反向遗传学工具如插入突变、靶基因的敲除只有在少数几个微藻(莱茵衣藻等)中建立。另外,据报道在微藻中建立同源重组方法是相对较困难的。因此,为实现基因的功能研究,rnai技术是一种比较好的选择,而且通过基因组测序发现,在绝大多数微藻中都存在有rnai元件如dicer酶、rna依赖的rna聚合酶和arogonute蛋白等。rnai指通过反义rna与正链rna形成双链rna特异性地抑制靶基因的现象,它通过人为地引入与内源靶基因具有相同序列的双链rna(有义rna和反义rna),从而诱导内源靶基因的mrna降解,达到阻止基因表达的目的。rnai发生于除原核生物以外的所有真核生物细胞内,并且在莱茵衣藻、三角褐指藻和杜氏盐藻中建立了rnai反向遗传学工具用于基因功能研究。
微拟球藻是一种单细胞光合微藻,广泛分布于海洋、淡水等环境水域。由于它的油脂含量高、较好的环境适应性及适于烟道气培养等特性,在微藻生物能源领域备受青睐。截至目前,已有多株微拟球藻的全基因组测序,基本涵盖了本属内的已知代表物种。同时,遗传转化方法也基本已经建立,反向遗传学工具如插入突变、过表达和同源重组的方法在不同种里面也有报道,唯独rnai这一强有力的遗传工具尚未建立,而本申请专利建立的方法将填补这一空白,将与其他遗传工具形成互补,能够更好地用于微拟球藻基因功能及未来的大规模遗传筛选研究。
技术实现要素:
本发明的目的是提供一种用于微拟球藻基因沉默的rnai载体及其应用。
为实现上述目的,本发明采用技术方案为:
一种用于微拟球藻基因沉默的rnai载体,载体以硅藻用rnai表达载体作为骨架,微拟球藻内源基因微管蛋白(tublin)启动子为启动子、终止子、抗性选择标记基因和靶基因。
所述微拟球藻内源基因微管蛋白(tubulin)启动子为
(1)具有序列表seqidno.1中的核苷酸序列;
或,(2)具有与序列表中seqidno.1的dna序列有较高的序列同源性,且编码相同功能蛋白质的dna序列。
上述微拟球藻微管蛋白tubulin基因,命名为tub,来源于工业微藻(nannochloropsisoceanicaimet1),其中,序列表中的seqidno.1由1218个碱基组成,编码一个完整的开放阅读框,它编码具有序列表seqidno.2的氨基酸残基序列的蛋白质。
所述靶基因为沉默靶基因及微拟球藻本身rnai元件识别的发夹结构。
所述发夹结构为针对靶基因形成发夹结构的反向重复序列;即,针对沉默靶基因序列设计约400bp的正向序列和约200bp的反向序列,其中这两个序列用于共同的200bp序列,在反向连接后能够互补形成颈环结构。
利用所述rnai技术的rnai表达载体为在微拟球藻中实现靶基因的沉默,需要设计靶基因的正反向重复序列形成颈环结构。
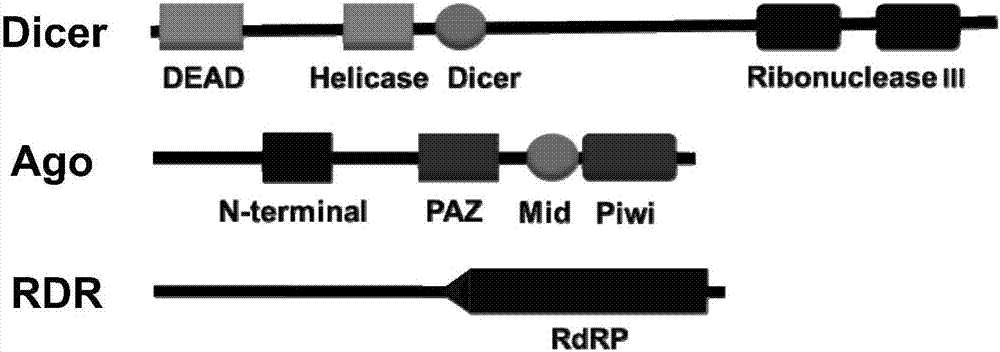
本发明比较了微拟球藻rnai所要行驶功能的主要元件比较,发现在微拟球藻中存在三个基本元件:arogonuate蛋白、dicer酶及rna依赖的rna聚合酶,它们存在保守的遗传。
所述抗性选择标记基因为博来霉素抗性基因ble,所用启动子为微管蛋白(tubulin)基因启动子,终止子为硅藻fcp基因终止子。
本发明构建了用于反向遗传学研究rna干涉(rnai)技术的表达载体,其载体骨架主要包括启动子序列(微拟球藻内源tublin基因启动子),ble抗性基因序列以及针对靶基因形成发夹结构的反向重复序列,其原理主要是基于不同基因的反向重复序列形成发夹结构而被微拟球藻本身固有的rnai元件所识别,进而实现靶标基因的特异下调。
rnai表达载体构建,
rnai表达载体的构建主要包括如下步骤:采用硅藻用rnai表达载体作为骨架,将骨架的硅藻fcp基因启动子置换为微拟球藻内源启动子,并克隆微拟球藻中需要沉默的靶基因的正反向重复序列,再连接到载体得到rnai表达载体;而后通过电脉冲转化微拟球藻,转化后微拟球藻培养液于不同的培养环境下进行培养,筛选其特定表型的遗传转化株,通过荧光定量pcr测定靶基因是否被沉默及沉默效率,通过生理评估遗传转化株表型。。
一种用于微拟球藻基因沉默的rnai载体在基因转化中的应用。
一种宿主细胞,宿主细胞为包含权利要求1所述的载体。
所述宿主细胞为微拟球藻,包括微拟球藻属内所有种、亚种及不同株,如微拟球藻代表种nannochloropsisoceanica、nannochloropsissalina、nannochloropsisoculata、nannochloropsisgadatina等。
本发明所具有的优点:
本发明构建的rnai表达载体能够用于微拟球藻相关藻株的基因功能鉴定,所构建rnai载体具有在表达水平实现靶基因基因敲低(geneknockdown),rnai这一强有力的反向遗传工具在微拟球藻中的建立,将与其他遗传工具形成互补,能够更好地用于微拟球藻基因功能及未来的大规模遗传筛选研究,更好地服务于藻类生物学及工业应用研究。
附图说明
图1为本发明提供的微拟球藻中存在的rnai元件(dicer酶、rna依赖的rna聚合酶)结构示意图;
图2为本发明提供的微拟球藻靶基因rnai表达载体的构建示意图,主要包括启动子,抗性基因和目的基因的正反向重复序列及发夹结构;
图3为本发明提供的转化株的pcr验证的电泳图,利用pcr扩增抗性基因序列,发现转化株存在目的条带约400bp(第二、三、四、五、六、泳道有条带),其中wt为野生型(第八泳道无条带),第一泳道为阳性对照(质粒dna为pcr模板);
图4为本发明提供的突变株和野生型的ca(基因号:g2209)基因在空气水平co2浓度下相对表达丰度变化突变株的碳酸酐酶基因ca2在rna表达水平明显被抑制,相对表达丰度降低在60%-80%左右,而其他几个遗传转化株显示基因没有被明显抑制。
图5为本发明提供的ca(g2209)遗传转化株和野生型在空气水平co2培养条件下表型点滴实验和液体培养下的表型表征;
图6为本发明提供的突变株和野生型微拟球藻在空气水平co2浓度培养条件下的生长曲线;
图7为本发明提供的来自空气水平co2浓度培养条件下突变株和野生型微拟球藻细胞的光合放氧速率;
图8为本发明提供的来自空气水平co2浓度培养条件下突变株和野生型光合系统ii的光合活性。
图9为本发明提供的微拟球藻imet1基因bct-g1855敲低实施例结果
图10为本发明提供的微拟球藻ccmp1779基因ca2敲低实施例结果
具体实施方式
下面结合附图和实施例对本发明作进一步详细说明,但本发明保护范围不局限于所述内容。
本发明通过反向遗传学策略rna干涉(rnai)技术敲低工业微藻微拟球藻中的感兴趣靶基因基因进而获得了能够用于基因功能研究的转化株,通过对遗传转化株生理、生化表型等测量进一步鉴定沉默基因的功能,通过本发明建立的rnai方法可用于微拟球藻其他未知基因的功能研究,也可用于大规模遗传表型筛选。本发明所公开的方法为工业微藻固定工业co2废气及生物能源产业应用奠定了分子遗传基础及可行性方法。
本发明实施例中所用的方法如无特别说明均为常规实验方法。
实施例1微拟球藻(nannochloropsisoceanicaimet1)碳酸酐酶(基因号:g2209)全序列的获得
通过对微拟球藻全基因组测序,初步获得了核基因组编码的碳酸酐酶(g2209)的全序列,如seqidno.1和seqidno.2所示。
进一步通过聚合酶链式反应pcr验证获得其碳酸酐酶的完整阅读框(orf),pcr引物序列为:f1为5’atgagcgtacaggcagtgcgag3’;r1为5’ctactcagacagcttcgccttc3’。pcr反应体系包括5xpcr反应缓冲液(5xdnabuffer)5ul、脱氧核糖核苷三磷酸(dntp)4ul、硫酸镁(mgso4)2ul、正向引物(forwardprimer)2ul、反向引物(reverseprimer)2ul、dna3ul、dna聚合酶(koddnapoleymase)0.5ul、ddh2o为32.5ul,总反应体系为50ul;
pcr反应程序均为第一步95℃5min,第二步95℃反应1min、58℃30s、72℃反应30s(30个循环),第三步72℃延伸7min,第四步4℃反应5min。经过pcr验证阅读框的结果是seqidno.1,与基因组注释预测序列相符。
实施例2
1)rnai表达载体的构建
rnai表达载体的构建以phir-ptgus为载体骨架,首先通过pcr扩增反应以微拟球藻基因组为模板,扩增获得208bp的片段(ca-2209基因序列195bp到413bp位置)和404bp片段(ca2基因序列195bp到599bp位置)两个pcr片段;
其中,扩增208bp的片段的引物为:
ca2209_fw(5’cggaattcatttttcgtgcctgcgttt3’;包含ecori位点)/ca2209_rv1(5’gctctagatggtctaaatgtcgattgttcg3’;包含xbai位点);
扩增404bp的片段的引物为:
ca2209_fw(5’cggaattcatttttcgtgcctgcgttt3’;包含ecori位点)/ca2209_rv2(5’gctctagagctccttgtgcttctccgta3’;包含xbai位点)。
上述进行pcr扩增的反应体系均为50ul:包括5xpcr反应缓冲液(5xdnabuffer)5ul、脱氧核糖核苷三磷酸(dntp)4ul、硫酸镁(mgso4)2ul、正向引物(forwardprimer)2ul、反向引物(reverseprimer)2ul、dna3ul、重组dna聚合酶(koddnapoleymase)0.5ul、ddh2o为32.5ul,总反应体系为50ul;
pcr反应程序均为第一步95℃5min,第二步95℃反应1min、60℃30s、72℃反应30s(30个循环),第三步72℃延伸7min,第四步4℃反应5min。
由上述获得两个片段含有一个共同的片段为209bp,目的形成反向互补序列(颈环发夹结构)。然后用ecori和xbai限制内切酶消化以上两个pcr片段并用pcr产物纯化试剂盒进行纯化,同时用ecori消化phir-ptgus载体并用牛小肠碱性磷酸酶进行片段去磷酸化处理后纯化。再将ecori和xbai消化的两个pcr片段与线性化载体(phir-ptgus的ecori消化后线性片段)用t4连接酶进行连接得到质粒phir-ptca2。微拟球藻beta-tubulin基因的启动子使用引物notub_fw(5’gcgagctcgccagctgcctcat3’;包含saci位点)和notub_rv(5’ggccatgg3tgtgtccgccgcc’;包含ncoi位点)被扩增,然后用saci和ncoi限制性内切酶酶切,同时载体phir-ptca2也用用saci和ncoi限制性内切酶酶切并纯化,随后再将beta-tubulin基因的启动子片段与线性化载体(beta-tubulin基因的启动子)用t4dna连接酶进行连接得到载体phir-ngca2,其载体包含beta-tubulin基因的启动子区、抗性基因ble、ca2的正反向重复序列以及fcp基因的终止子,组成表达盒(如图1所示)。
2)rnai表达盒的pcr扩增
用pcr引物(f1:5’cccagtcacgacgttgtaaaacg3’;r1:5’ggaaacagctatgaccatg3’)扩增以上实施例2的表达载体得到包含有tub基因启动子、抗性基因ble、能形成发夹结构的ca2序列以及转录终止子序列,pcr反应体系为5xpcr反应缓冲液(5xdnabuffer)5ul,脱氧核糖核苷三磷酸(dntp)4ul,硫酸镁(mgso4)2ul,正向引物(primerf1)2ul,反向引物(primerr1)2ul,模板dna(dna)3ul,dna聚合酶(koddnapoleymase)0.5ul,ddh2o32.5ul,总反应体系为50ul;pcr反应程序为第一步95℃5min,第二步95℃1min56℃30s72℃45s(30个循环),第三步72℃延伸7min,第四步4℃5min。待pcr扩增反应完成后,将得到的pcr产物电泳检测和进一步pcr产物纯化,使其浓度达到3-5ug/ul,放于-20℃保存,待下一步电脉冲转化时备用。
3)微拟球藻突变株的获得
采用电脉冲转化方法将上述得到rnai表达盒片段导入野生型微拟球藻。具体实施方法为:将野生型微拟球藻在f/2培养基中培养至浓度达到3×107cells/ml(培养条件为25℃,连续光照培养,光强为50umol/m-2s-1,通气量为100ml/min),离心收集微藻细胞倒掉上清液,然后用4℃预冷的375mm的山梨醇溶液洗涤藻细胞三次并用其重悬藻细胞,使藻细胞浓度为8×108cells/ml;将浓缩后的藻细胞溶液分成200ul一份,然后向每份中加入3ug上述获得线性化载体和1ul鲑鱼精dna(15ug/ml),混匀后冰上放置10min;再将混合物转移入2mm电击杯,以2200v(hv),600欧姆,50微法(uf)进行电击,电击后立即将藻体以5ml新鲜的f/2培养基转移入无菌的白头玻璃管,遮光后在光照摇床中培养(用弱光进行复苏),转速设置为100rpm,25℃复苏48h;复苏后将其在5000g离心5min,弃尽上清,而后用1ml20%玉米淀粉重悬藻体后,倒在琼脂含量0.8%的f/2的平板上,均匀铺开并将平板在超净工作台吹干后密封,放于25℃光照培养,待3-4周后挑选转化子。
所述20%玉米淀粉为取2g玉米淀粉,用无水乙醇和超纯水交替洗3遍,再用10ml70%乙醇重悬即成,以f/2培养基洗4次,再以10ml含0.8%peg8000的f/2培养液进行重悬。
所述琼脂含量0.8%的f/2的平板上为预先制备琼脂平板,再添加3-5ug/ml的zeocin抗生素)
实施例3转化子的pcr验证
用牙签挑选转化子于新鲜的f/2培养基(包含3-5ug/ml的zeocin抗生素),放于光照培养箱培养,待其生长2-3周,离心收集藻细胞,用基因组dna提取试剂盒(omegahpdnaextrationkit)提取转化株的基因组dna,同时也提取野生型微拟球藻的基因组dna,作为后续实验的对照。提取基因组并用nannodrop定量后,用正向引物f1(5’ttatcaacggcataccggcactg3’)和反向引物r1(5’ctgatgaacagggtcacgtcgt3’)进行pcr扩增反应,反应体系为50ul:5xpcr反应缓冲液(5xdnabuffer)5ul,脱氧核糖核苷三磷酸(dntp)4ul,硫酸镁(mgso4)2ul,正向引物(primerf1)2ul,反向引物(primerr1)2ul,模版dna(dna)3ul,dna聚合酶(kodplusdnapoleymase)为0.5ul,ddh2o32.5ul,总反应体系为50ul。反应程序为:第一步95℃5min,第二步95℃1min56℃30s72℃45s(30个循环),第三步72℃延伸7min,第四步4℃5min。pcr扩增反应完成后,琼脂糖凝胶电泳检测,看是否使用转化子dna为模板有400bp条带,而且野生型没有条带,如图所示2,证实外源片段已经整合到微拟球藻基因组,电脉冲转化成功,得到突变株,突变株可分别标记为m1、m2、m3、m4和m5。
实施例4微拟球藻突变株ca(g2209)基因相对表达水平测量
将上述获得突变株和野生型分别在空气水平co2浓度下培养,光强控制一致在50umol/m2/s,通气速率在100ml/min。待微藻培养至对数生长期时收集微藻,5000g离心5min,然后用液氮迅速冷冻,保存在-80℃备用。
突变株表达丰度实验测量分为以下三步:
第一步rna的提取,将冷冻保存的藻细胞进行rna提取,首先采用液氮研磨的方式破碎藻细胞,一般液氮研磨5-10min,然后加入1mltrizol(invitrogen公司)迅速充分混匀并加入200ul氯仿在12000rpm下离心10min,吸取上清液转入新的eppendorf管中再加入等体积氯仿,充分振荡混匀,静置2-3min后以12000rpm下离心10min,将上清液转入新的离心管并加入等体积异丙醇室温沉淀15min,离心15min后,再用75%乙醇洗涤沉淀后,吹干加入50uldepc处理的ddh2o,测量rna浓度后,保存于-80℃备用;
第二步,cdna的制备,rna样本按照takara试剂
第三步,荧光定量pcr反应,通过
通过以上方法计算ca-2209基因在突变株中的相对野生型的基因表达丰度,结果证实突变株(m4或m5)的ca-2209基因表达丰度无论是在空气水平co2浓度培养条件下显著低于野生型ca-2209的表达,证实ca-2209基因在转录水平被抑制,大约下调60%-80%,如图3所示,进一步证实ca-2209基因在突变株m4、m5中被沉默,而突变株m1、m2和m3中ca-2209基因没有被沉默。
实施例5微拟球藻ca-2209基因沉默转化株的生长表型测量
将微拟球藻ca-2209基因沉默转化株和野生型微拟球藻分别用牙签挑取单藻落置于f/2培养基中在50ml三角瓶中培养2-3周,然后测量od750下的细胞密度,而后重新接种至新鲜的f/2培养基,使接种浓度保持一致,即相同的od750值=0.5-1.0,然后连续传代培养2-3次,将微藻接种至200ml的柱式反应器培养(反应器内培养基为f/2培养基),当生长到对数生长期后,将突变株和野生型微拟球藻分别稀释至相同的浓度(上述测定od750值相同),分别吸取10ul稀释液滴到预先制备好的f/2固体琼脂平板上(琼脂浓度为0.8%并且不添加抗生素),每个藻样分别滴到两块平板上(在空气水平co2浓度下培养,两个重复),在超净工作台上吹干后,在室温、无光或低光环境下放置2-3天,然后将平板放置于简易培养箱(专利申请号:201410339234.9),并将向培养箱通入空气浓度co2(0.04%),光强50umol/m2/s,用此培养箱可以进行相对封闭式培养筛选,通过调节培养箱进气口和出气口阀门可以控制培养箱内部稳定的co2环境,不定期观察微藻(突变株和野生型)的生长情况(图4)。同时,也将从200ml柱式反应器中接种微藻到新鲜液体培养基中测量生长曲线和干重等生理指标。测量结果如图5,6所示,发现突变株在空气水平co2浓度下生长明显慢于野生型,经10天培养后干重增加30%左右(图6)。因此,通过基因敲低的方法验证了碳酸酐酶ca-2209的基因功能,这个基因在微拟球藻低co2浓度(如空气水平co2浓度)生长中扮演重要角色,可能是参与co2浓缩机制的重要酶,此基因的功能鉴定为工业化烟道气下培养工业微藻打下了坚实的产业化基础。
实施例6微拟球藻ca-2209基因沉默转化株的生理表型测量
将上述微拟球藻ca-2209基因沉默转化株和野生型细胞的光合放氧速率测定采用clark-ii型氧电极(英国hansatech公司)进行,测定时反应槽的温度为4℃(水浴调整),光强用遮光片调整为300μmolm-2s-1。将微拟球藻突变株和野生型细胞培养至对数生长期,以5000g离心5min收集藻细胞与15ml离心管,每个样品收集两份,一份用于测定光合放氧速率,另外一份用于叶绿素含量的测量。收集的微藻细胞先用无碳水(预先通入n2排除掉溶液中的co2)洗涤,然后又悬浮在不同ph值的缓冲液中,依次为20mmoll-1的mes缓冲液,20mmoll-1tris缓冲液和20mmoll-1的apes缓冲液的无碳水中(ph分别是6.0、7.8或9.0),缓冲液的配置用海水然后分别测定突变株和野生型细胞在不同ph条件下的光合放氧速率。同时每个样品留取一份用来测量叶绿素a的含量,叶绿素a的测量采用甲醇抽提的方法,采用以下这个公式计算叶绿素a(chla)的含量:
chla的含量(ug/ml)=16.5169xa665-8.0962xa652
光合放氧速率用叶绿素a的含量进行换算。测量结果显示,突变株细胞的光合放氧速率在ph6.0条件下,微拟球藻ca-2209基因沉默转化株株的光合放氧速率有显著提高,而在ph9.0条件下,突变株的光合放氧速率又显著降低,因此突变株能够抵抗由于co2浓度升高导致的ph降低,在一定程度上显示出耐酸性,可能叶绿体内部的微环境得到改善,即ph得到提升,但在ph碱性条件下基因被抑制,所以显现出慢速生长,如图7所示。此外,通过测定光合系统ii的光合光能转换效率,也显示微拟球藻ca-2209基因沉默转化株有较低的光合活性值,如图8所示,也进一步证实微拟球藻ca-2209基因在光合作用中起着非常关键作用。
实施例7微拟球藻imet1中敲低碳酸氢盐转运蛋白(bct-g1855)参照实施例2、3的载体构建方法,将表达载体中imet1中基因ca-2209正反向重复序列替换为bct-1855基因的正反向重复序列(基因序列为seqidno.3),实现敲低一个碳酸氢盐转运蛋白基因,通过定量pcr验证转化株的表达水平被抑制60-80%,然而,通过表型测量未发现遗传转化株有明显表型变化(如图9),推测与此基因的组成型表达有关系,因为此基因在不同营养盐条件下(二氧化碳限制或氮元素缺乏)的表达均未变化。
实施例8微拟球藻ccmp1779体系建立rnai技术平台以上实施例是在微拟球藻imet1中建立的rnai基因敲低方法,为了验证本发明构建的rnai表达载体更有普遍适用性,在微拟球藻ccmp1779中参照实施例2、3的载体构建方法,将实施例2中rnai表达载体中imet1中基因ca-2209正反向重复序列替换为ccmp1779的ca2基因(基因序列为seqidno.4)的正反向重复序列,实现敲低一个碳酸酐酶ca2基因,通过定量pcr验证转化株的表达水平被抑制,通过表型测量发现遗传转化株在空气水平co2条件下生长速率未有明显变化,而在高co2(5%)培养条件下敲低了ca2基因的遗传转化株较野生型微拟球藻ccmp1779有较高的生长速率,提高约30-40%,如图10。
seqidno.1
atgagcgtacaggcagtgcgaggaacggcatggttgcatttttcgtgcctgcgtttcgggctttgggtgaggaggcgagcttggcgaagggactctcggaggaggcagaccaaacatggcagggtgcttattcttgataatccgtccatttcacgctttggcacctcattcatacactcttggccattctatttgagactgctgtgtccgggtggtgtatttggttctggggttggggtggttggcccgaacaatcgacatttagaccaagcatcatcatcctcatcctcgaagcaagcagggcaagcaaaatctagtcacccgcatcgcctttacacccgcacaggctactctacgacacgcagcagcatgtcgcgcatgattcggaaaggattggacacgctgggcaaggcgttgcgggagaccggtcaagctttggaccgagcaggcctcgaggctttgggtacggagaagcacaaggagctttggtcacgacaccgccaagtctccaaccttttcgaccaacgcccgcgcgtcgccagcaatgccttcgtcgccccatcggccacggtcgtgggtgaagtcgagctctgggacaaggtctccatctggtacggcgctgtcatccgcggcgatcgcaacagcgtcaagatcgggaattgtaccaatgtgcaagaccacgcggttatcaccacagtccccacactcgatacgggctttcccgccaaggttgacatcggacacatggtcactatcggccacggcgcgaccatcacctcagccaccatccaagacaaagcactcatcgggatcgggagcgtcgtctccgaaggcgcgcttgtcgaaaccggcgcccagctcgcggccggttccgtggtcccgcccgggcgccgcatccctgcaggagagctctggggcggcaaccccgtcaaatttatccgtaagctggaggatgaggaggtagtggcaaatgtgacggcagcagaggcctaccatgccttggctcgcacccatgagaatgagttccaaccctttggggctgcttacctggacgcggagaaggcgaagctgtctgagtag
(a)序列特征:orf序列
●长度:1080bp
●类型:dna序列
●链型:单链
●拓扑结构:线性
(b)分子类型:全部为外显子序列
(c)假设:否
(d)反义:否
(e)最初来源:微拟球藻imet1
seqidno.2
msvqavrgtawlhfsclrfglwvrrrawrrdsrrrqtkhgrvlildnpsisrfgtsfihswpfylrllcpggvfgsgvgvvgpnnrhldqasssssskqagqaksshphrlytrtgysttrssmsrmirkgldtlgkalretgqaldraglealgtekhkelwsrhrqvsnlfdqrprvasnafvapsatvvgevelwdkvsiwygavirgdrnsvkignctnvqdhavittvptldtgfpakvdighmvtighgatitsatiqdkaligigsvvsegalvetgaqlaagsvvppgrripagelwggnpvkfirkledeevvanvtaaeayhalarthenefqpfgaayldaekaklse
(a)序列特征:氨基酸序列
●长度:359aa
●类型:蛋白质编码序列
●链型:单链
●拓扑结构:线性
(b)分子类型:氨基酸
(c)假设:否
(d)反义:否
(e)最初来源:微拟球藻imet1
seqidno.3
atgctatccaaacgccagcttacgggaggcgtagcgctagcactattcagcgcgtatccggcacactccttctgttactcccgtttcagtttgccagttgtggatagaacgactctctcctccgcctccccctcctcctccatttcctcctcctcccctttctctggcagatcgctaccaagcagtcgcggcggcaacgcacgcacgtatggagagcctctacgtgtctcgatgccctttcacataaggtttagtaccggtgtagccatgccaccggacgactctccctcctcctattcctcctcatcccggtcatcatccaccgcttccctctactcagcccccacctcctcctcttccccctcctctttccccgccttgcccagccgaggatggagaatagaatggtttaagggcctcagggaggacattgccaatcgtaggccgctctataaatcggattggaccgatggaatgaaagtaaagacgattccgaccatcctctttttatactttgcctgtttggctcctgcggttgcatttggagggatcagctatgggttgacacagggggctatgggcgtcattgagttcttgctgtcttgtggggtgtcgggcatgttgtatgcaattttttcggggcagccaatgaccttcatcggccccacgggactgacattgtgttttatggctgcgctcttcaatttttgcaattcttttgggttggcgttcatgcccatgtacacctgggtagggatctggacgagcctgatgcttgtgacgctgtcatccgcgaatgcctgtttgttgaccaagcactgtacgcgctttacggacgaggtgttcaacgcgctcttggcgctgaattttacctacgaagctctcaaaaatatcgtcgtccgcttcttctccaccggtaccgacaaaacccaacccttcctcgccctcaacatggccttggcaacatttgccctgtcccaacgcctggtctcgtaccgttcttcgatcctcttcacgcgtcgcatccgaggcttcttgtcggatttcggccccgccctcaccatcatcatcatgacgttgctcggctccctacccatggtccgtggcctgggcatcgagtatttgaaagccccgaaacggttccagttcggtaacgggcggcgtttattcgtccctttgtgggatttaccggtctgggcgcgtggattggcgttggtaccggcggcggccttgacagtgttgttctttttagatcacaatattagtgtgcgggtggtaaattcacccgacaacaagatgaagaaaagcccggcgtatcacctggatttgtttgtgctagggagcattgtctttttgtgctcgattgtggggttgccgtggatgtgtgcggcgacgattcagagcctgaatcatgtgaggagtgtcactgagtatgaacctgtcggagtaggagaaggaggaaagggggggggaacagcagcagtagttgtaagcggaagcggaggaggtaaaggagatgtaaaacaggaagaggaagagacggcgactgtgatggagacgcggttgacgggcttggggatccacgccctggtgttgtcgagcattggccttttgcctctattgagacgtatcccgattcccgtgatcagtggcatctttctttatcttggccggacggtcatgacgggcaatacgttcttacaacgggtcaagggcgcgttttttgacaagagtatgctacccgaagactcgcccttcaaacggttggggagaacgactgtgaacaagtttacggcgattcaggtgggctgcttggcggtgttgtggaccttgaagagtaatccggcaaccgggatgtttttcccgagcgtgattgggatgttgattctgacgaggatcttggtgatcccgcggttgttttccaaggaggagatttcggtgttggatgtggattag
(a)序列特征:orf序列
●长度:1956bp
●类型:dna序列
●链型:单链
●拓扑结构:线性
(b)分子类型:全部为外显子序列
(c)假设:否
(d)反义:否
(e)最初来源:微拟球藻imet1
seqidno.4
atgtggcggcgtgtgctcgcatattcctcgagattcgatgtaacacatctcaatagcgaggtaacgctatccacgtatctcaccattgtctccaccctctctattaaccccatctttctatgcagttcgcaaaaatctccgatacgcggccctgcggttaatccaacaagtcaagaccttgaaacgtcggaaatgagcaacccgccgtctcctgtcgtgccgtctcccagccttacacaatcaaatcatgacatgctcaccagcatccgcagccacctagacgaccaagaggtcaagatccaggaacaattatttcgcttccagggcattaagagggccattgaccgcatttgcaagaatgacgaggcctacatggactcccccgatattgtcaaggcacagaccacgctcgaagatcgcccggagctggaaaggaaaggagagactgctgctgaagccttgcgcctgctcaaagtgggcaacgctgcctttttgcgcgatgaggttgtgcctgtccaccgctctgacagacatattggcctacattttgcccagaaaccccacgccataattattgcttgttccgactctcgggtcccgccggagatgatctttaattgcgcgtttggtgaactcttcgtcatccgccttgccggcaacaccgtcgatgcactcgctcgcgccagcctcctttacgcggtgcaacacctccaatctcccctggtcgttgttctgggccacgaaaagtgcggcgccgtcacagccgccttgcagccggaagaggagctggcggatgctcccggggacattaagaccttggtgcgaaacatcaaaaggggcatagagtgcgaattgattgacccgagcgcccacattttcggcgatgaaagactgttgtgcgcgattgtctgcaacgtgcattacgtggcccgcaccctgtcggaacaccccgatatccgcccatttattgaccgaaggaaactgtccgtcgtgggcgcctactacgcttttagtggggtggtgtcgttctttgacgaccaggatgagggaggaaggatggagagagggcgggtgtttgggggtgtggagggaaacggatccttgtgcttaagttgttccggaactttgtccggaaactcgagtccgatggcggggggagggagcaagggatccagtccgcctccgagcgtgcaggatggggttaataaaagcgtctag
(a)序列特征:orf序列
●长度:1218bp
●类型:dna序列
●链型:单链
●拓扑结构:线性
(b)分子类型:全部为外显子序列
(c)假设:否
(d)反义:否
(e)最初来源:微拟球藻ccmp1779。
- 还没有人留言评论。精彩留言会获得点赞!