里氏木霉的突变菌株的制作方法
本发明涉及在丝状真菌细胞中生产蛋白质的方法,包括在所述细胞中过表达trazf1基因或其变体。
背景技术
由于大量原料的可利用性以及乙醇作为燃料的优势,从纤维素生产乙醇的可能性受到了极大的关注。用于这个过程的基于纤维素的天然原料称为“生物质”。很多类型的生物质,例如木材、农业残渣、草本作物和城市固体废物被认为是生产生物燃料的潜在原料。这些材料主要由纤维素、半纤维素和木质素组成。
纤维素是由通过β-1,4键连接的葡萄糖分子组成的聚合物,其非常耐降解或解聚。一旦纤维素被转化为葡萄糖,很容易用酵母将后者发酵为生物燃料,例如乙醇。
研究用于将纤维素转化为葡萄糖的最老的方法基于酸水解。该方法可以在浓酸或稀酸存在下进行。然而,一些缺点,例如当使用浓酸时,酸的回收差以及在使用稀酸的情况下,葡萄糖产量低,对于酸水解方法的成本效益而言是不利的。
为了克服酸水解方法的缺点,最近的纤维素转化方法更涉及使用纤维素酶类型的酶的酶促水解。包含水解纤维素的酶的微生物例如有真菌木霉属(trichoderma)、曲霉属(aspergillus)、腐质霉属(humicola)和镰刀菌属(fusarium)。然而,这种木质纤维素生物质(例如纤维素)的酶促水解在工业规模上具有昂贵的缺点。
为了降低酶促水解木质纤维素有关的成本,业界持续在寻找更有生产力或提供更佳产量的方法。
因此,需要优化工业方法中的酶生产。尤其是,存在未被满足的期待已久的需要来开发改进的经济上有利的生产酶的方法。
因此,发明人开发了用于在丝状真菌中生产目标蛋白的改进的方法,其中所述蛋白尤其是纤维分解酶。
技术实现要素:
因此,本发明涉及在丝状真菌细胞中生产蛋白质的方法,包括在所述细胞中过表达trazf1基因或其变体。
本发明还涉及过表达trazf1基因或其变体的丝状真菌菌株,优选里氏木霉(trichodermareesei)菌株。表述“过表达trazf1基因或其变体”意欲是指所述菌株具有至少trazf1基因或其变体,所述基因或其变体组成型表达。
根据一个实施方案,根据本发明的丝状真菌菌株,优选里氏木霉菌株,包含内源trazf1基因,所述基因在组成型启动子的控制下。在这种情况下,内源trazf1基因存在于菌株的天然基因组中,且启动子被改造、突变或替换,以成为组成型的。
根据另一个实施方案,除了存在于其基因组中的trazf1基因,根据本发明的丝状真菌菌株,优选里氏木霉菌株,包含额外拷贝的trazf1基因,所述拷贝组成型表达。因此,在这种情况下,它包含至少两个拷贝的trazf1基因,这些拷贝之一组成型表达。
根据本发明的该突变菌株的特征在于,与没有被改造的相同里氏木霉菌株相比,或与参考里氏木霉菌株相比,纤维素分解酶的产量提高。
里氏木霉是分解纤维素的丝状真菌。鉴于里氏木霉能够分泌大量纤维素酶和半纤维素酶,该菌株用于生产将植物生物质材料转化为工业用途的生物产品(例如生物乙醇)的酶是非常有利的。
表述“里氏木霉参考菌株”意欲是指选自菌株qm6a、ng14、rutc30和qm9414的里氏木霉菌株。这些菌株是公众可得的,并且尤其是分别以以下保藏号保藏的主题:
-atcc13631(菌株qm6a);
-atcc56767(菌株ng14);
-atcc56765(菌株rutc30);和
-atcc26921(菌株qm9414)。
在一个具体的实施方案中,所述菌株是cl847菌株。该菌株是高产菌株。
在可以根据本发明使用的丝状真菌中,可以提及以下门的真菌:子囊菌(子囊菌门(ascomycota))、担子菌(担子菌门(basidiomycota))和接合菌(接合菌门(zygomycota))。通常,真菌选自以下纲:鸟纲菌纲、盘菌纲、座囊菌纲、欧洲菌纲、蜡苔藓纲、利奥菌纲、粪壳菌纲和酵母菌。
更具体地,可以提及里氏木霉、少孢节丛孢菌(arthrobotrysoligospora)、黑孢块菌(tubermelanosporum)、甘蓝链格孢菌(alternariabrassicicola)、baudoiniacompniacensis、异旋孢腔菌(cochliobolusheterostrophus)、禾旋孢腔菌(cochliobolussativus)、hysteriumpulicare、油菜茎基溃疡病菌(leptosphaeriamaculans)、松霉(mycosphaerellapini)、杨球腔菌(mycosphaerellapopulorum)、颖枯壳针孢(phaeosphaerianodorum)、菲氏假尾孢菌(pseudocercosporafijiensis)、圆核腔菌(pyrenophorateres)、小麦黄斑病真菌(pyrenophoratritici-repentis)、rhytidhysteronrufulum,玉米大斑病菌(setosphaeriaturcica)、叶斑病菌(zymoseptoriatritici)、荚膜阿耶罗菌(ajellomycescapsulatus)、皮炎组织胞浆菌(ajellomycesdermatitidis,本海姆节皮菌(arthrodermabenhamiae)、石膏样节皮菌(arthrodermagypseum)、太田节皮菌(arthrodermaotae,棘孢曲霉(aspergillusaculeatus)、炭黑曲霉(aspergilluscarbonarius)、棒曲霉(aspergillusclavatus)、黄曲霉(aspergillusflavus)、烟曲霉(aspergillusfumigatus)、黑曲霉(aspergillusniger)、米曲霉(aspergillusoryzae)、土曲霉(aspergillusterreus)、粗球孢子菌(coccidioidesimmitis)、双相粗球孢子菌(coccidioidesposadasii)、构巢裸胞壳(emericellanidulans)、费氏新萨托菌(neosartoryafischeri)、巴西副球孢子菌(paracoccidioidesbrasiliensis)、paracoccidioidessp.lutzii、产黄青霉(penicilliumchrysogenum)、马尔尼菲青霉(penicilliummarneffei)、柄篮状菌(talaromycesstipitatus)、马发癣菌(trichophytonequinum)、红色毛癣菌(trichophytonrubrum)、断发毛癣菌(trichophytontonsurans)、疣发癣菌(trichophytonverrucosum)、uncinocarpusreesii、cladoniagrayi、灰葡萄孢霉(botryotiniafuckeliana)、geomycesdestructans、核盘菌(sclerotiniasclerotiorum)、嗜碱枝顶孢(acremoniumalcalophilum)、球毛壳(chaetomiumglobosum)、希金斯刺盘孢(colletotrichumhigginsianum)、寄生隐丛赤壳菌(cryphonectriaparasitica)、羊茅香柱霉(epichloefestucae)、尖孢镰刀菌(fusariumoxysporum)、禾顶囊壳(gaeumannomycesgraminis)、串珠状赤霉(gibberellamoniliformis)、玉米赤霉(gibberellazeae)、禾谷球孢菌(glomerellagraminicola)、稻瘟病菌(magnaporthegrisea)、夏季斑枯病菌(magnaporthepoae)、嗜热毁丝霉(myceliophthorathermophila)、茎基腐病(nectriahaematococca)、粗糙链孢菌(neurosporacrassa)、离散脉孢菌(neurosporadiscreta)、四孢脉孢菌(neurosporatetrasperma)、柄孢霉(podosporaanserina)、大孢粪壳(sordariamacrospora)、太瑞斯梭孢壳霉(thielaviaterrestris)、深绿木霉(trichodermaatroviride)、绿木霉菌(trichodermavirens)、黑白轮枝孢(verticilliumalbo-atrum)、大丽轮枝菌(verticilliumdahliae)、卢西坦棒孢(clavisporalusitaniae)、膜醭毕赤酵母(pichiamembranifaciens)、树干毕赤酵母(scheffersomycesstipitis)、异常威克汉姆酵母(wickerhamomycesanomalus)、虫拟蜡菌(ceriporiopsissubvermispora)、红缘拟层孔菌(fomitopsispinicola)、大伏革菌(phlebiopsisgigantea,wallemiasebi)、松杨栅锈菌(melampsoralarici-populina)或米根霉(rhizopusoryzae)。
优选地,可以根据本发明使用的丝状真菌是里氏木霉。
在本发明的上下文中,术语“trazf1基因”意欲是指序列seqidno:1或seqidno:3的基因。该基因编码的转录因子的天然蛋白序列如seqidno:2所示。以genbank登录号xm_006961831.1(里氏木霉菌株qm6a),在里氏木霉基因组专门的位置,以号码id103275可以在以下地址在线获得该序列:http://genome.jgi.doe.gov/cgi-bin/dispgenemodel?db=trire2&id=103275。
序列seqidno:1是没有内含子的trazf1基因的核苷酸序列。
序列seqidno:3是trazf1基因的基因组核苷酸序列。它由4个外显子和3个内含子组成。它如下所示,其中示出了3个内含子。atggccctcgcagctcaacagcatactcaggccgattggggccgctggtctcaccagattccacagagttttcccatgatgggctctccaggattcatgtcatacgatcccagagctcaggacggcagtcagatgcagcgtcaggtgtctgctcagtacctggtgaactcgaactacaaccagcccccgatgcccactgcttcgtctccccagtatcaacacgcagggccattttcctatgtgccttaccacagcccgccgccgtccactcctcttggttccccattcaagagcgaatttcccgagcaccctcttacgcgcatgacacactctacggtcgatcgacaccattctcaggccatgagggactaccaaccttattctcctgtatcgaggaggggatcgatttcttcagtcgccaccaagccctcagcagctcccgtcacaccaggtccaactactcctggctctttcacttcaagttccgacgcccagagccccagcactccaaacccccagactgcgtctcagcctgtcagctccaagactctcacttacaatgagaccgttcatccgggcgataggatcagcttcagaaccgatgttgatgaactcatgaaggccatccagaagacacagacgaccgacgagtgtcagcaaacactcacacctgcgcgaacaccaaagaactgtaccacaagtactcccgtacttcgtacacaaagcgggaagccgagaaaacagtgggtttgcgatggccccaactgcggcaaggcctttgtccagaagacgcatcgcgacattcaccgacgcactcacaccggccatcgaccatacgtacgcgcccagctcctcttcactgcaacgccggctaattaaatgttgataggtctgcaccatggaaaattgcggtcttacgttctcgcagcgaggaaacctcaaggtaagcttcagctgctaagaatctcctttgagaatgcgtatactgaccagatggtgtgtggacagactcacatacgacgccacacaggtgaaaagccgttctcttgcgctgcttgtggcaagtgcttcgctcagcgtgggaatcttcgatcccacgaggagacacacaaaggcctgaagcccttcgtctgccggctcgatgattgcaacaagtcgttttctcagctgggcaatatgaaggtatgcaacatctagcacatgaaagcagtatgagaacgctctaacgctgagggaactgcagactcatcagaacaactttcacaaagaaacgctccagaaactcacacacatgtttgtgcaattctcggagaacggcgaggtgcccagagactatcaggatcttttcgaatacttccagaagcactacaagaatagcaacaagggagtcaagggccgaggaaagactcgcgctgtggcagctcgtgggcctcaagattccgcgtttcggcaggctgcctccccagtgcccgcgttactgaagacgccggctacgactcatttgccccagatgacaatgccagcccatgatccccatggcagaatctcaccatacgccatgacccagggagctgcgaacactctgagcaatgtcctgcgcaaccccaacccctcttacggcctttatggacccacgtttgccccgggccctgtacgagatggcgtctttcacatgggcattgcgagccacctatcctga
(seqidno:3)
因此,序列seqidno:1对应于没有内含子的序列seqidno:3。
因此,根据本发明的菌株优选包含至少两个拷贝的序列seqidno:3或至少一个拷贝的序列seqidno:1。
术语trazf1基因的“变体”意欲是指编码的蛋白与序列seqidno:2具有相同功能(即,转录因子)的基因。优选地,trazf1基因的变体是同源基因。
优选地,trazf1基因的变体编码的蛋白与序列seqidno:2具有相同功能。优选地,trazf1基因的变体编码的蛋白选自序列seqidno:6-seqidno:140。
发明人首次证明了过表达trazf1基因造成纤维分解蛋白的产量显著增加。
尤其是,发明人证明了根据本发明突变的里氏木霉菌株rutc30和cl847(即,包含两个拷贝的trazf1基因,其中之一组成型表达)显示胞外蛋白质,尤其是纤维分解酶的产量提高。
根据本发明产生的蛋白优选纤维分解酶,更优选纤维素酶或半纤维素酶,甚至更优选它们是纤维素酶。
根据本发明的方法涉及在丝状真菌细胞中生产蛋白质,包含在所述细胞中过表达trazf1基因或其变体。
过表达trazf1基因优选通过在细胞基因组中引入包含所述基因的盒来进行。这种方法的知识是本领域技术人员的知识的一部分。
优选地,所述盒包含:
a)至少一个组成型启动子;
b)序列seqidno:1或seqidno:3的基因或其变体;和
c)任选地,终止子。
组成型启动子a)是强启动子。术语“组成型启动子”意欲是指非诱导性的启动子。该组成型启动子总是表达基因;因此,产生持续且强烈的蛋白质生产。
该启动子尤其可以来源于里氏木霉,但也可以来源于构巢曲霉(aspergillusnidulans)。
优选地,组成型启动子a)选自:
-里氏木霉的gpd启动子(lij.等(2012),achievingefficientproteinexpressionintrichodermareeseibyusingstrongconstitutivepromoters,microbialcellfactories,11(1),84.doi:10.1186/1475-2859-11-84)。该启动子的序列是seqidno:4;
-构巢曲霉的gpd启动子(
-里氏木霉的tef1启动子(nakari-
优选地,组成型启动子a)是序列seqidno:4的gpd启动子。
用于本发明的方法的盒还包含元件b),即序列seqidno:1或seqidno:3的基因或其变体。变体如上所述。
最后,用于本发明的方法的盒可以任选地包含终止子c)。
终止子优选是序列seqidno:5的里氏木霉的gpd终止子。
本发明还涉及如上所述的盒的用于生产蛋白质,尤其是纤维分解酶的用途。
过表达trazf1基因优选通过引入以下来进行:包含trazf1基因的dna片段和组成型启动子、终止子和可选择标志物。这个盒还可以通过包含该盒的载体,例如质粒引入细胞。根据本发明,术语“载体”意欲是指可能在其中插入外源核酸片段的任何dna序列,载体能够将外源dna引入宿主细胞。载体的实例有质粒、粘粒、酵母人工染色体(yac)、细菌人工染色体(bac)和细菌噬菌体p1衍生的人工染色体(pac)和病毒衍生的载体。
根据本发明的载体还可以携带可选择标志物。术语“可选择标志物”意欲是指其表达赋予包含它的细胞使得能够选择它们的特征的基因。它例如是耐抗生素基因。
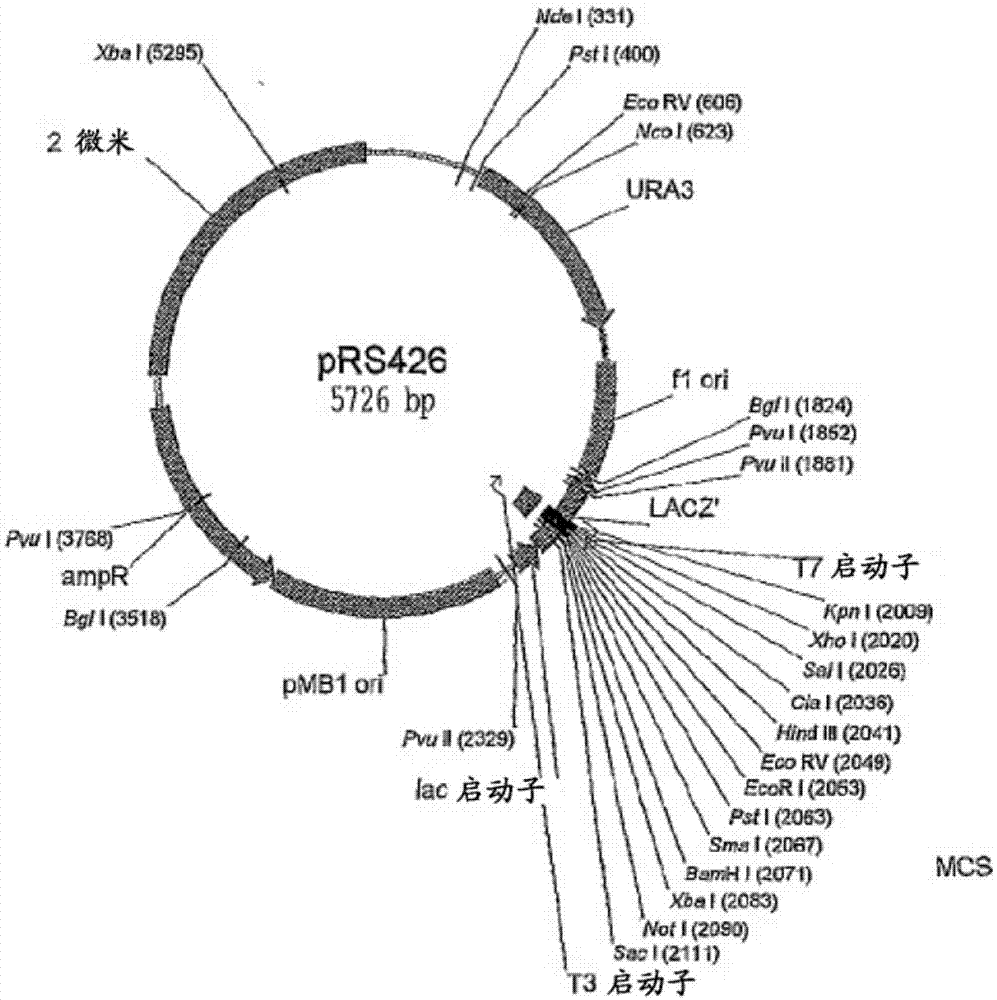
优选地,所述载体是质粒。更优选地,质粒是prs426类型的质粒,如实施例和图1所述。
一旦插入基因组,根据本发明的盒允许目标trazf1基因的翻译,其提供相应的蛋白。
本发明还涉及根据本发明的菌株用于生产纤维分解酶的用途。因此,在一个具体的实施方案中,本发明涉及里氏木霉菌株用于生产纤维分解酶的用途,所述菌株过表达trazf1基因或其变体。
本发明还涉及根据本发明的菌株用于将纤维素及其降解产物,包括纤维二糖水解为葡萄糖的用途。
本发明的一个主题还是根据本发明的菌株用于生产生物燃料的用途。根据本发明,术语“生物燃料”可以定义为从生物质转化产生的可以用于能源目的的任何产物。此外,不希望受限与此,可以提及的实例有生物气,可以加入燃料(任选在随后的转化之后)或其本身可以称为燃料的产物,例如醇类(乙醇、丁醇和/或异丙醇,取决于所用的发酵生物的类型)、溶剂(丙酮)、酸(丁酸)、脂质及其衍生物(短链或长链脂肪酸、脂肪酸酯)和氢气。
优选地,根据本发明的生物燃料是醇类,例如乙醇、丁醇和/或异丙醇。更优选地,根据本发明的生物燃料是乙醇。在另一个实施方案中,生物燃料是生物气。
本发明还涉及根据本发明的菌株用于水解β-寡糖的用途。
以下实施例说明本发明而不限制其范围。
附图说明
图1:prs426质粒的示意图。
图2:用于验证盒的存在的引物的位置,所述盒用于转化菌株的过表达。
图3:培养7天后,根据本发明转化的rutc30菌株和没有被改造的里氏木霉的rutc30菌株的胞外蛋白的产量。
图4:培养7天后,根据本发明转化的cl847菌株和没有被改造的里氏木霉的cl847菌株的胞外蛋白的产量。
图5:在补料瓶培养条件下,cl847菌株和两个转化体的比生产率。
具体实施方式
实施例1:里氏木霉菌株atcc56767和atcc56765的乳糖诱导的转录组学研究
在该研究中,发明人使用以下里氏木霉菌株:
-atcc56767(ng14),和
-atcc56765(rutc30)。
rutc30菌株通过突变ng14菌株获得,具有增加的纤维素酶产量。
trazf1基因(id103275)是转录因子(参与其他基因调控的蛋白质),其被鉴定为在乳糖诱导的动力学期间差异性表达(poggi-parodi等2014)。更具体地,转录组学研究表明,trazf1的表达在ng14菌株的诱导期间降低,而在rutc30中,它的表达没有变化。
实施例2:构建trazf1基因的过表达盒
过表达盒由三个dna片段组成:
-里氏木霉甘油醛-3-磷酸脱氢酶基因的启动子区(gpdp)。启动子区的大小在li等2012的文章中定义(seqidno:141);
-目标基因(trazf1)的编码区(seqidno:3);
-里氏木霉甘油醛-3-磷酸脱氢酶基因的终止子区(gpdt)。终止子区的大小在li等2012的文章中定义(seqidno:142)。
为了能够选择转化体,将包含hph基因的称为hygror的盒(seqidno:143)与过表达盒融合。hph基因编码负责对潮霉素b的抗性的潮霉素b磷酸转移酶。它从大肠杆菌(escherichiacoli)分离。为了在里氏木霉中表达基因,将hph基因置于粗糙脉孢霉(neurosporacrassa)的cpc-1基因的启动子(ncu04050)和构巢曲霉的trpc基因的终止子(anid_00648)的控制下。
根据基于文献(schuster等2012)的方法,通过在酿酒酵母(s.cerevisiae)中重组,用prs426质粒(atcc77107)作为载体来构建缺失盒。为此,用ecor和xhoi(newenglandbiolabs)消化质粒,并用qiaquickgelextraction试剂盒(qiagen)通过凝胶电泳纯化。prs426质粒如图1所示。
基于v2.0版本的基因组(http://genome.jgi-psf.org/trire2/trire2.home.html)中的orf预测来设计对于目标基因trazf1特异性的引物部分。
扩增里氏木霉甘油醛-3-磷酸脱氢酶基因的启动子区gpdp
用于从rutc30菌株的dna扩增启动子区gpdp的引物如下所述:
·正向引物由两部分组成:一部分是在xhoi限制性位点附近的20个核苷酸(nt)且与prs426载体序列同源(图1),一部分是20nt且与gpdp(seqidno:141)的5’端同源,整体形成序列seqidno:144
(attgggtaccgggcccccccgacgcagaagaaggaaatcg)。
·反向引物由两部分组成:一部分是10nt且与trazf1基因(seqidno:3)的编码区的5’端同源;一部分是20nt且与gpdp(seqidno:141)的3’端同源,整体形成序列seqidno:145
(cgagggccattttgtatctgcgaattgagc)。
扩增目标trazf1基因的编码区
用于从rutc30菌株的dna扩增目标trazf1基因的引物如下所述:
·正向引物由两部分组成:一部分是10nt且与gpdp(seqidno:141)的3’端同源,一部分是20nt且与trazf1基因(seqidno:3)的5’编码区同源;整体形成序列seqidno:146
(cagatacaaaatggccctcgcagctcaaca)。
·反向引物由两部分组成:一部分是10nt且与gpdt区(seqidno:142)的5’端同源,一部分是20nt且与trazf1基因(seqidno:3)的3’编码区同源,整体形成序列seqidno:147
(aacacagcactcaggataggtggctcgcaatg)。
扩增里氏木霉甘油醛-3-磷酸脱氢酶基因的终止子区gpdt
用于从rutc30菌株的dna扩增终止子区gpdt的引物如下所述:
·正向引物由两部分组成:一部分是10nt且与trazf1基因(seqidno:3)的编码区的3’端同源,一部分是20nt且与gpdt(seqidno:142)的5’端同源;整体形成序列seqidno:148
(cctatcctgagtgctgtgttcctcagaatg)。
·反向引物由两部分组成:一部分是10nt且与hygror盒(seqidno:143)的5’端同源,一部分是20nt且与gpdt(seqidno:142)的3’端同源,整体形成序列seqidno:149
(ggtacacttgttacggatctgatcactcgg)。
扩增hygror盒
用于扩增hygror盒的引物如下所述:
·正向引物由两部分组成:一部分是10nt且与gpdt(seqidno:142)的3’端同源,一部分是20nt且与hygror盒(seqidno:143)的5’端同源;整体形成序列seqidno:150
(agatccgtaacaagtgtacctgtgcattctg)。
·反向引物由两部分组成:一部分是在ecori限制性位点附近的20nt且与prs426载体序列同源,一部分是20nt且与hygror盒(seqidno:143)的3’端同源,整体形成序列seqidno:151
(tggatcccccgggctgcaggggcagtgctagtgtgtgtac)。
用以上引物进行的pcr产生具有同源末端的dna片段。用这些dna片段和消化的prs426质粒转化感受态的酿酒酵母菌株w303。
从酿酒酵母提取包含trazf1基因的过表达盒和hygror盒的prs426质粒,并将其作为模板用正向引物(seqidno:152)和反向引物(seqidno:153)来pcr扩增与hygror盒融合的过表达盒(seqidno:154)。
实施例3:用trazf1过表达盒转化里氏木霉菌株rutc30和cl847
用pcr产物(seqidno:154)转化里氏木霉rutc30(atcc56765)和cl847菌株(durand等,1988)。基于hph可选择标志物基因功能选择转化体。从由单个孢子产生的菌落纯化转化体。
通过三次pcr扩增确认盒的异位整合。所用引物以及扩增片段的位置如图2所示。用于pcr的引物如下所述:
·用于证实扩增1的引物:
-gpdp启动子中的正向引物(seqidno:155):
gtcagaaacgaccaagctaag。
-trazf1基因中的反向引物(seqidno:155):
gcctgagaatggtgtcgatc。
·用于证实扩增2的引物:
-trazf1基因中的正向引物(seqidno:157):
tcgtgggcctcaagattc。
-gpdt终止子中的反向引物(seqidno:158):
gacgcctgagaggtccta。
·用于证实扩增3的引物:
-gpdt终止子中的正向引物(seqidno:159):
ccttcttagagagctctcgg。
-hygror盒中的反向引物(seqidno:160):
cgggtttacctcttccagat。
从纯化转化体的基因组dna进行扩增:每个菌株选择4个转化体。
实施例4:培养具有trazf1过表达盒的里氏木霉菌株rutc30并分析培养物的蛋白生产
用源自rutc30菌株的四个转化体并且显示trazf1过表达盒异位整合的孢子接种每个孔含有2ml培养基的24孔培养板。
培养基由以下组成:k2hpo48.7g.l-1;(nh4)2so44.2g.l-1;mgso4.7h2o0.3g.l-1;玉米浆1.5g.l-1;乳糖10g.l-1;纤维素10g.l-1;马来酸11.6g.l-1;cacl20.3g.l-1;feso4.7h2o5.0mg.l-1;mnso4.h2o1.6mg.l-1;znso4.7h2o1.4mg.l-1;cocl2.6h2o2.0mg.l-1;ph6。
于30℃下以150rpm进行摇瓶培养,重复两次。
培养7天后,收集上清液以测量培养基中的蛋白浓度(folin法)。培养物的胞外蛋白产量如图3所示。该图表明,与未改造的里氏木霉菌株相比,转化体的蛋白产量增加。
实施例5:培养具有trazf1过表达盒的里氏木霉菌株cl847并分析培养物的蛋白生产
用源自cl847菌株的四个转化体并且显示trazf1过表达盒异位整合的孢子接种每个孔含有2ml培养基的24孔培养板。
培养基由以下组成:k2hpo48.7g.l-1;(nh4)2so44.2g.l-1;mgso4.7h2o0.3g.l-1;玉米浆1.5g.l-1;乳糖10g.l-1;纤维素10g.l-1;马来酸11.6g.l-1;cacl20.3g.l-1;feso4.7h2o5.0mg.l-1;mnso4.h2o1.6mg.l-1;znso4.7h2o1.4mg.l-1;cocl2.6h2o2.0mg.l-1;ph6。
于30℃下以150rpm进行摇瓶培养,重复两次。
培养7天后,收集上清液以测量培养基中的蛋白浓度(folin法)。培养物的胞外蛋白产量如图4所示。该图表明,与未改造的里氏木霉菌株相比,转化体的蛋白产量增加。
实施例6:在补料瓶中培养具有过表达的azf1的里氏木霉菌株cl847并分析培养物的蛋白生产和生长
使用专利wo2013/026964a1中所述的纤维素酶生产方法,培养里氏木霉菌株cl847的两个转化体和未转化菌株。随时间测量胞外蛋白的生产和生物质,以获得培养结束时蛋白的比生产率(每单位时间内每mg真菌生物质产生的蛋白或qp)。这些数值如图5所示。
数据表明,与未改造的里氏木霉菌株相比,突变体的qp值升高。
参考文献
durand,h.,clanet,m.,&tiraby,g.(1988).geneticimprovementoftrichodermareeseiforlargescalecellulaseproduction.enzymeandmicrobialtechnology,10,341–346.
li,junxin;wang,juan;wang,shaowen;xing,miao;yu,shaowen;liu,gang(2012)achievingefficientproteinexpressionintrichodermareeseibyusingstrongconstitutivepromoters.in:microbialcellfactories,vol.11,p.84.doi:10.1186/1475-2859-11-84.
poggi-parodi,dante;bidard,frédérique;pirayre,aurélie;portnoy,thomas;blugeon,corinne;seiboth,bernhardetal.(2014)kinetictranscriptomeanalysisrevealsanessentiallyintactinductionsysteminacellulasehyper-producertrichodermareeseistrain.in:biotechnologyforbiofuels,vol.7,n°1,p.173.doi:10.1186/s13068-014-0173-z.
schuster,andré;bruno,kenneths.;collett,jamesr.;baker,scotte.;seiboth,bernhard;kubicek,christianp.;schmoll,monika(2012)aversatiletoolkitforhighthroughputfunctionalgenomicswithtrichodermareesei.in:biotechnologyforbiofuels,vol.5,n°1,p.1.doi:10.1186/1754-6834-5-1.
wo09026716(a1)-methodforcellulaseproduction。
- 还没有人留言评论。精彩留言会获得点赞!