一种调控植物体内生长素平衡的EHD1蛋白及其应用的制作方法
本发明属于生物
技术领域:
,具体涉及一种调控植物体内生长素平衡的EHD1蛋白及其应用。
背景技术:
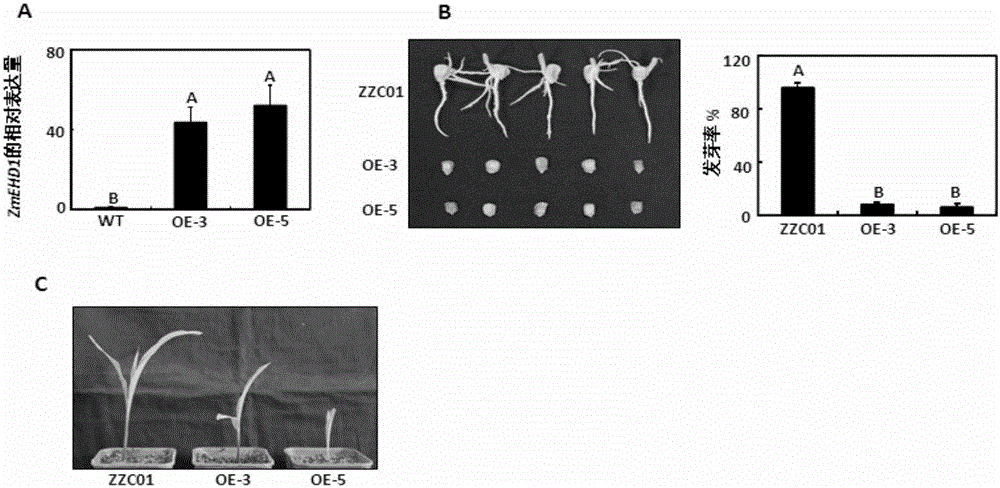
:内吞作用指质膜蛋白的内在转化或者吸收胞外物质转运到胞内的过程,分为依靠网络蛋白介导的内吞作用(CME,clathrin-mediatedendocytosis)和不依靠网络蛋白介导的内吞作用(Murphyetal.,2005)。在植物中,网络蛋白介导的内吞作用是外界物质进入植物细胞的主要途径(Fanetal.,2015)。与动物内吞作用的深入研究相比,最近几年才证明网络蛋白介导的内吞作用对植物发育的各个阶段非常关键,包括决定细胞的极性(Vanetal.,2011;Wangetal.,2013),雄性生殖器官的发育(BlackbournandJackson,1996;Kimetal.,2013),配子的发育(Backuesetal.,2010)和胚胎的形成(Fanetal.,2013;Kitakuraetal.,2011)。与此同时,网络蛋白介导的内吞作用在非生物和生物胁迫的植物响应方面也起着重要作用,这些胁迫是通过膜上的转运体或者受体转化成胁迫信号,例如硼转运子(Takanoetal.,2010),铁介导的转运子(Barberonetal.,2011),生长素输出载体(Dhonuksheetal.,2007),油菜素内酯受体(Dietal.,2013),细菌型鞭毛蛋白受体(Robatzeketal.,2006)和乙烯诱导木聚糖酶受体(BarandAvni,2009)。CME是一个复杂的过程,至少可以分为4个步骤:1,形成一个囊泡;2,将物质包裹在囊泡中;3,从质膜上释放成熟的囊泡;4,与内体融合(Sigismundetal.,2012)。因为膜蛋白不能直接结合到质膜或者物质上,所以含有网络蛋白内吞作用的囊泡的形成必须激活衔接蛋白复合体2(AP2)和质膜的联系,因此AP2在网格蛋白介导的内吞作用方面起着重要作用。AP2复合体形成异源四聚体的复合物,包括两个大亚基(β1-5和γ/α/δ/ε/ζ),一个中等亚基(σ1-5)和一个小亚基(σ1-5)(McMahonandBoucrot,2011)。除了典型的AP2复合体之外,其他的辅助蛋白也能把外界物质与质膜联系起来,形成含有网格蛋白内吞作用的囊泡,例如C端Eps15同源结构域。玉米在主要的粮食作物中,总产量处于首位,并且也是生物能源和其他工业材料重要的原材料(Mclaren,2005)。作为生物学研究重要的模式系统,玉米在对植物发育和进化的认识中有着突出的贡献,并且这些研究已经被用于阐释玉米籽粒形态建成的发育机理。缺陷型籽粒(dek)突变体是最早在玉米中进行遗传研究的材料之一。尽管几百个缺陷型籽粒突变体基因已被分离,但是由于致死等位基因研究的难度,只有几个相关的基因被分子克隆(ScanlonandTakacs,2009)。拟南芥ARF2通过抑制细胞分裂控制种子大小(Schruffetal.,2006),而水稻BIGGRAIN1(BG1)的活化通过调节生长素运输显著提高籽粒大小(Liuetal.,2015),同时,IAA-葡萄糖水解酶负调控水稻的胚乳发育(Ishimaruetal.,2013)。前人通过分析种子特异且可生长的玉米胚乳缺陷型突变体de18,阐明了玉米IAA合成关键基因ZmYUC1对于玉米正常的胚乳发育是必须的(Bernardietal.,2012)。因而,生长素的合成、转运和信号可能协作调控玉米的营养发育和生殖发育(Gallavottietal.,2008;LiandLi,2016)。技术实现要素:本发明的一个目的是提供一种蛋白质。本发明提供的蛋白质是如下a)或b)或c)或d)的蛋白质:a)氨基酸序列是序列4所示的蛋白质;b)氨基酸序列是序列2所示的蛋白质;c)在序列4或序列2所示的蛋白质的N端和/或C端连接标签得到的融合蛋白质;d)将序列4或序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的具有相同功能的蛋白质。其中,序列2和序列4均由547个氨基酸残基组成。为了使a)中的蛋白质便于纯化,可在序列表中序列4或序列2所示的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。表1、标签的序列标签残基序列Poly-Arg5-6(通常为5个)RRRRRPoly-His2-10(通常为6个)HHHHHHFLAG8DYKDDDDKStrep-tagII8WSHPQFEKc-myc10EQKLISEEDL上述c)中的蛋白质,所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。上述c)中的蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。上述c)中的蛋白质的编码基因可通过将序列1或序列3所示的DNA序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表1所示的标签的编码序列得到。本发明的另一个目的是提供与上述蛋白质相关的生物材料。本发明提供的与上述蛋白质相关的生物材料为下述A1)至A12)中的任一种:A1)编码上述蛋白质的核酸分子;A2)含有A1)所述核酸分子的表达盒;A3)含有A1)所述核酸分子的重组载体;A4)含有A2)所述表达盒的重组载体;A5)含有A1)所述核酸分子的重组微生物;A6)含有A2)所述表达盒的重组微生物;A7)含有A3)所述重组载体的重组微生物;A8)含有A4)所述重组载体的重组微生物;A9)含有A1)所述核酸分子的转基因植物细胞系;A10)含有A2)所述表达盒的转基因植物细胞系;A11)含有A3)所述重组载体的转基因植物细胞系;A12)含有A4)所述重组载体的转基因植物细胞系。上述相关生物材料中,A1)所述核酸分子为如下1)或2)或3)或4)所示的基因:1)其编码序列是序列3所示的cDNA分子或DNA分子;2)其编码序列是序列1所示的cDNA分子或DNA分子;3)与1)或2)限定的核苷酸序列具有75%或75%以上同一性,且编码上述蛋白质的cDNA分子或基因组DNA分子;4)在严格条件下与1)或2)或3)限定的核苷酸序列杂交,且编码上述蛋白质的cDNA分子或基因组DNA分子。其中,所述核酸分子可以是DNA,如cDNA、基因组DNA或重组DNA;所述核酸分子也可以是RNA,如mRNA或hnRNA等。本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码上述蛋白质的核苷酸序列进行突变。那些经过人工修饰的,具有编码上述蛋白质的核苷酸序列75%或者更高同一性的核苷酸,只要编码上述蛋白质且具有相同功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列2或序列4所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。本发明还有一个目的是提供上述蛋白质或上述相关生物材料的新用途。本发明提供了上述蛋白质或上述相关生物材料在调控植物体内生长素含量中的应用。上述应用中,所述调控为提高或降低。上述应用中,上述1)所述蛋白质或其相关生物材料是降低植物体内生长素含量。上述应用中,上述2)所述蛋白质或其相关生物材料是提高植物体内生长素含量。本发明还提供了上述蛋白质或上述相关生物材料在调控植物籽粒发育和/或营养生长中的应用。上述应用中,所述调控为促进或抑制。上述应用中,上述1)所述蛋白质或其相关生物材料是抑制植物籽粒发育和/或营养生长。上述应用中,上述2)所述蛋白质或其相关生物材料是促进植物籽粒发育和/或营养生长。本发明还提供了上述蛋白质或上述相关生物材料在调控植物内吞作用中的应用。上述应用中,所述调控植物内吞作用体现在与ZmAP2σ亚基互作影响植物网络蛋白介导的内吞作用。上述应用中,所述影响为与ZmAP2σ亚基互作维持植物网络蛋白介导的内吞作用,使植物网络蛋白介导的内吞作用顺利进行。本发明还有一个目的是提供上述1)所述的蛋白质或与1)所述的蛋白质相关的生物材料的新用途。本发明提供了上述1)所述的蛋白质或与1)所述的蛋白质相关的生物材料在培育生长素含量降低和/或籽粒发育延缓的转基因植物中的应用。本发明的最后一个目的是提供一种培育生长素含量降低和/或籽粒发育延缓的转基因植物的方法。本发明提供的培育生长素含量降低和/或籽粒发育延缓的转基因植物的方法包括提高上述1)所述的蛋白质活性,得到转基因植物的步骤;所述转基因植物的生长素含量低于所述受体植物;所述转基因植物的籽粒发育延缓于所述受体植物。上述方法中,所述转基因植物的籽粒发育延缓于所述受体植物体现为如下a1)-a5)中的任一种:a1)所述转基因植物的百粒重低于所述受体植物;a2)所述转基因植物的发芽率低于所述受体植物;a3)所述转基因植物的初生根短于所述受体植物;a4)所述转基因植物的侧根数少于所述受体植物;a5)所述转基因植物的植株小于所述受体植物。上述方法中,所述提高上述1)所述的蛋白质活性的方法为将上述1)所述的蛋白质活性的编码基因导入受体植物。上述方法中,上述1)所述的蛋白质活性的编码基因的核苷酸序列为序列3。上述方法中,所述受体植物为单子叶植物或双子叶植物,所述单子叶植物具体为玉米。本发明从玉米中克隆了ZmEHD1基因,并构建得到籽粒发育和营养生长受损的转ZmEHD1mut玉米。通过实验研究表明:和野生型玉米植株相比,转ZmEHD1mut玉米的生长素含量降低,且外源施加有活性的NAA能够提高转ZmEHD1mut玉米的种子发芽率。ZmEHD1通过与ZmAP2σ亚基互作影响CME过程,且ZmEHD1介导的CME过程通过调节植物体内生长素含量而对玉米籽粒发育和营养生长起着至关重要的作用。附图说明图1为转ZmEHD1mut玉米纯合株系的表达量及发芽率检测。其中,OE3和OE5均为转ZmEHD1mut玉米纯合株系;ZZC01为转空载体植株;WT为野生型玉米植株。图2为转ZmEHD1mut玉米纯合株系的籽粒表型。其中,WT为野生型玉米植株,OE为转ZmEHD1mut玉米纯合株系。图3为转ZmEHD1mut玉米纯合株系和野生型玉米在授粉后15天、20天、25天和35天籽粒胚乳中ZmEHD1的表达情况。图4为转ZmEHD1mut玉米纯合株系和野生型玉米的表型检测。其中,OE为转ZmEHD1mut玉米纯合株系;WT为野生型玉米植株。图5为转ZmEHD1mut玉米纯合株系的内吞作用受到抑制。其中,OE3和OE5均为转ZmEHD1mut玉米纯合株系;ZZC01为转空载体植株;WT为野生型玉米植株。图6为玉米ZmEHD1蛋白的亚细胞定位。图7为玉米AP2σ能够回补拟南芥ap2σ突变体的表型。图8为玉米ZmEHD1蛋白与玉米AP2σ互作。图9为转ZmEHD1mut玉米纯合株系和野生型玉米的皮尔逊相关系数。其中,WT为野生型玉米植株,OE为转ZmEHD1mut玉米纯合株系。图10为通过RT-PCR验证RNA-seq结果。其中,WT为野生型玉米植株,OE为转ZmEHD1mut玉米纯合株系。图11为不同浓度1-NAA处理转ZmEHD1mut玉米纯合株系种子。其中,WT为野生型玉米植株,OE为转ZmEHD1mut玉米纯合株系。图12为用不同浓度赤霉素处理转ZmEHD1mut玉米纯合株系和野生型玉米种子。其中,WT为野生型玉米植株,OE为转ZmEHD1mut玉米纯合株系。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。下述实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中的pSPYNE载体和pSPYCE载体均在文献“Walteretal.,2004Visualizationofproteininteractionsinlivingplantcellsusingbimolecularfluorescencecomplementation.PlantJ.2004Nov;40(3):428-38.”中公开过,公众可从河南农业大学获得。下述实施例中涉及的试剂及培养基的配方:1)20%葡萄糖:称取20g葡萄糖,加入100ml蒸馏水,溶解充分后过滤除菌。2)0.2%腺嘌呤盐酸盐:称取0.4g固体粉末,加入200ml蒸馏水,溶解充分后过滤除菌。3)SD-Trp缺陷培养基:称取0.67gYNB(无氨基酸)培养基、0.074gDO-Trp、2g琼脂粉,加入90ml蒸馏水,121℃灭菌15min,超净台中加入10ml20%的无菌葡萄糖,铺平板。4)SD-Leu缺陷培养基:称取0.67gYNB(无氨基酸)培养基、0.069gDO-Leu、2g琼脂粉,加入90ml蒸馏水,121℃灭菌15min,超净台中加入10ml20%的无菌葡萄糖,铺平板。5)SD-Leu-Trp缺陷培养基(三缺培养基):称取0.67gYNB(无氨基酸)培养基、0.064gDO-Leu-Trp、2g琼脂粉,加入90ml蒸馏水,121℃灭菌15min,超净台中加入10ml20%的无菌葡萄糖,铺平板。6)SD-Leu-Trp-His缺陷培养基(四缺培养基):称取0.67gYNB(无氨基酸)培养基、0.062gDO-Leu-Trp-His、2g琼脂粉,加入90ml蒸馏水,121℃灭菌15min,超净台中加入10ml20%的无菌葡萄糖,铺平板。7)SD-Leu-Trp-His-Ade缺陷培养基:称取0.67gYNB(无氨基酸)培养基、0.060gDO-Leu-Trp-His-Ade、2g琼脂粉,加入90ml蒸馏水,121℃灭菌15min,超净台中加入10ml20%的无菌葡萄糖,铺平板。8)10×TE缓冲液(pH7.5):10ml1MTris-HCl(pH7.5)、2ml0.5MEDTA(pH8.0),定容至100ml后,121℃灭菌15min。9)50%的PEG-4000:称取50gPEG-4000溶于100ml蒸馏水后,121℃灭菌15min。下述实施例中的CPB载体在文献“WenwenLiu,QingSun,KaiWang,QingguoDuandWen-XueLiNitrogenLimitationAdaptation(NLA)isinvolvedinsource-to-sinkremobilizationofnitratebymediatingthedegradationofNRT1.7inArabidopsisNewPhytologist(2016)doi:10.1111/nph.14396”中公开过,公众可从河南农业大学获得。下述实施例中的抑制剂E-64D是CaymanChemical的产品,产品目录号为13533。下述实施例中的拟南芥ap2σ亚基功能缺失的突变体在文献“LushengFan1,2,*,HuaiqingHao1,*,YiqunXue1,2,LiangZhang3,KaiSong1,2,ZhaojunDing4,MiguelA.Botella5,HaiyangWang6andJinxingLin1,DynamicanalysisofArabidopsisAP2σsubunitrevealsakeyroleinclathrin-mediatedendocytosisandplantdevelopment.Development.2013Sep;140(18):3826-37.doi:10.1242/dev.095711.Epub2013Aug7.”中公开过,公众可从河南农业大学获得。下述实施例中的pCAMBIA3301载体在文献“LiJ,FuJ,ChenY,FanK,HeC,ZhangZ,LiL,LiuY,ZhengJ,RenD,WangG.TheU6Biogenesis-Like1PlaysanImportantRoleinMaizeKernelandSeedlingDevelopmentbyAffectingthe3'EndProcessingofU6snRNA.MolPlant.2016Nov4.pii:S1674-2052(16)30263-5.doi:10.1016/j.molp.2016.10.016.[Epubaheadofprint]”中公开过,公众可从河南农业大学获得。实施例1、ZmEHD1蛋白的获得以野生型玉米(浚9058)的cDNA为模板,采用F:ATGGATATCGCGCGGGCGGC和R:TCACTCGTACGGGTTCCGCA引物进行PCR扩增,得到PCR扩增产物,并对PCR扩增产物进行测序分析。测序结果表明:PCR扩增得到大小为1644bp的条带,其核苷酸序列如序列1所示,将序列1所示的基因命名为ZmEHD1基因,ZmEHD1基因编码的蛋白质的氨基酸序列如序列表中序列2所示,将序列2所示的蛋白质命名为ZmEHD1蛋白。ZmEHD1蛋白含有EHD家族的典型结构:一个有两个calcium-bindingEF-hands(15-39aa和49-84aa)组成的EHdomain,一个P-loop(GQYSTGKT),一个dynamin-typeguaninenucleotide-bindingdomain,一个coil-coildomain。实施例2、转ZmEHD1mut玉米的制备及其表型检测一、转ZmEHD1mut玉米的制备1、表达载体的构建以BamHI(NewEnglandBiolabs,R3136L)为限制性酶切位点,利用In-Fusion(takara,639648)方法将序列3所示的ZmEHD1mut基因序列克隆到pCAMBIA3301载体上,得到重组质粒pCAMBIA3301-ZmEHD1mut。该质粒包含了一个由CaMV35S启动子启动的链霉菌属吸水链霉菌草丁膦乙酰转移酶基因(bar)。重组质粒pCAMBIA3301-ZmEHD1mut表达序列4所示的ZmEHD1mut蛋白。ZmEHD1mut基因序列为将序列1所示的ZmEHD1基因的第366位由G突变为A,且第58位由T突变为C,且第633位由G突变为C,且第1172位由T突变为C,且第1299位由A突变为G,且第1302位由C突变为T,且第1396位由A突变为G,且第1443位由C突变为G,且第1473位由C突变为T,且保持序列1其他序列不变后得到的序列。ZmEHD1mut蛋白的氨基酸序列为将序列2第391位由V突变为A,且第466位由M突变为V,且第481位由D突变为E,且保持序列2其他序列不变后后得到的序列。2、重组菌的制备将步骤1制备的重组质粒pCAMBIA3301-ZmEHD1mut通过电击法转入农杆菌EHA105(北京华越洋生物科技有限公司,GX0133-100),得到重组菌pCAMBIA3301-ZmEHD1mut/EHA105。将步骤1制备的pCAMBIA3301载体通过电击法转入农杆菌EHA105,得到重组菌pCAMBIA3301/EHA105。3、转ZmEHD1mut玉米的获得及鉴定用重组菌pCAMBIA3301-ZmEHD1mut/EHA105侵染野生型玉米自交系Qi319的幼胚,得到转基因植株。转基因植株的培养和筛选的具体步骤参照文献“X.-Q.Huang·Z.-M.WeiHigh-frequencyplantregenerationthroughcallusinitiationfrommatureembryosofmaize(ZeaMaysL.)PlantCellRep(2004)22:793–800DOI10.1007/s00299-003-0748-9”中的方法。用重组菌pCAMBIA3301/EHA105侵染野生型玉米自交系Qi319的幼胚,得到转空载体玉米(ZZCO1)。用荧光定量PCR方法对转基因植株做进一步检测,得到阳性转ZmEHD1mut玉米纯合株系。荧光定量检测的具体步骤如下:提取转基因植株和野生型玉米植株叶片的RNA,反转录为cDNA,用F:GATGACCTCCTCTGTGACCTGAA,R:AAGTTGTCTATGAGTCGCTGCTG引物进行荧光定量PCR检测。在96孔板中配制如下反应体系:cDNA2.0μl、SYBRPremixExTaqTM(2×)10μl、50×DyeⅡ0.4μl、PrimerF/PrimerR0.5/0.5μl,ddH2O补足20μl。使用ABI7500RealTimePCRSystem荧光定量PCR仪进行荧光定量PCR反应。每个基因设置三个重复,每个基因设置一个阴性对照,按如下程序进行扩增反应:95℃预变性30s;95℃变性5s,60℃退火30s,72℃延伸34s,40个循环。每个循环在延伸阶段采集信号。反应结束后导出Ct值,利用比较Ct值分析基因的表达水平,同时琼脂糖凝胶电泳检测扩增的专一性及其准确性。结果如图1A所示。阳性转ZmEHD1mut玉米纯合株系OE3和OE5中的ZmEHD1基因表达量显著高于野生型。选取阳性转ZmEHD1mut玉米纯合株系OE3和OE5,用于下述实验。二、转ZmEHD1mut玉米的表型分析1、授粉后籽粒发育情况检测对转ZmEHD1mut玉米纯合株系OE3和OE5及野生型玉米植株(野生型玉米自交系Qi319)进行授粉,并检测授粉后的籽粒发育情况。授粉的具体步骤如下:前期,将要授粉的植株的雌穗在吐丝之前套上雌穗袋,在雄穗颖壳打开露出花药时,提早套上雄穗袋。授粉时,将植株弯曲,使雄穗处于身后防止串粉,最后,一只手取下雌穗上的纸袋,另一只手小心滴将花粉均匀地洒在花丝上,将纸袋套在雌穗上,用大头针将其固定在雌穗上,写好标记。授粉后14天之前阳性转ZmEHD1mut玉米纯合株系OE3和OE5与野生型籽粒相似,但授粉后16天野生型籽粒已明显变为淡黄色,而转ZmEHD1mut玉米纯合株系籽粒仍为乳白色(图2A)。成熟后,阳性转ZmEHD1mut玉米纯合株系OE3和OE5胚乳皱缩,胚小(图2B),导致其百粒重显著下降(图2C)。为了研究转ZmEHD1mut玉米纯合株系和野生型籽粒的发育情况,用石蜡切片观察授粉后15天的籽粒胚的纵切面。石蜡切片的具体制备步骤参照文献“Yu-JunLiu,Zhi-HuiXiu,RobertMeeley,andBao-CaiTan,EmptyPericarp5EncodesaPentatricopeptideRepeatProteinThatIsRequiredforMitochondrialRNAEditingandSeedDevelopmentinMaize。ThePlantCell,Vol.25:868–883,March2013,www.plantcell.org2013AmericanSocietyofPlantBiologists.Allrightsreserved.”中的方法。石蜡切片观察胚的纵切面可以看出:转ZmEHD1mut玉米纯合株系胚的发育比野生型粒胚的发育要延缓(图2D),胚乳细胞的排列也较为疏松(图2E)。2、授粉后表达量检测按照步骤一的3中的荧光定量PCR方法检测转ZmEHD1mut玉米纯合株系和野生型玉米在授粉后15天、20天、25天和35天籽粒胚乳中ZmEHD1的表达情况。结果如图3所示。结果表明:ZmEHD1在转ZmEHD1mut玉米纯合株系中的表达量均高于野生型,且随着籽粒发育时间的延长,表达量呈下降趋势。3、发芽率检测用水浸泡种子8小时催芽,浸泡过程中保持通气。浸泡后将种子移至灭过菌的滤纸上,保持滤纸湿润,在26℃黑暗条件下萌发。每天观察种子发芽情况,统计发芽率。用纸培的方法测定发芽率发现转ZmEHD1mut玉米纯合株系的发芽率明显低于野生型玉米植株,只有3%(图4A),而且与野生型对照相比,转ZmEHD1mut玉米纯合株系初生根短,侧根数少(图4B),并且98%的转ZmEHD1mut玉米纯合株系到第二完全展开叶前死去(图4C)。转ZmEHD1mut玉米纯合株系OE3和OE5的籽粒的发芽率也明显低于转空载体玉米(ZZCO1)(图1B)。上述结果表明ZmEHD1对玉米的营养生长和籽粒发育至关重要。4、植株大小检测当发芽的转ZmEHD1mut玉米纯合株系OE3和OE5、野生型玉米和转空载体玉米(ZZCO1)种子长到一叶一心时移栽到装好土的培养盆里,在玉米培养间培养,观察植株生长状态。ZmEHD1mut玉米纯合株系OE3和OE5的种子可以生长到两叶期,但明显比转空载体玉米(ZZCO1)小(图1C)。野生型玉米种子的生长情况和转空载体玉米(ZZCO1)种子的生长情况无显著性差异。5、内吞作用FM4-64在水相中没有荧光,只有插入脂质生物膜后才显示较强的荧光,胞吞回收的囊泡因吞进细胞内部而被荧光标记,利用这种荧光探针能够对细胞的胞吞过程进行有效标记。采用FM4-64标记实验来验证玉米ZmEHD1蛋白在玉米内吞作用中发挥作用。具体步骤如下:将野生型玉米植株、转ZmEHD1mut玉米纯合株系OE3和OE5和转空载体玉米(ZZCO1)的根用5μMFM4-64(LifeTechnologies,F34653)标记10分钟,然后观察随着时间的延伸,植株内吞作用的变化。结果如图5所示:野生型玉米植株的根标记20分钟后可观察到约40%细胞(40/105)出现荧光斑点,而此时转ZmEHD1mut玉米纯合株系仍然观察不到荧光斑点(0/120)。标记60分钟后,野生型玉米植株有73%细胞(62/85)出现荧光斑点,转ZmEHD1mut玉米纯合株系却只有37%的细胞(26/70)出现(图5A)。转空载体玉米(ZZCO1)的根系与野生型玉米植株出现的同样的现象(图5B),说明转ZmEHD1mut玉米纯合株系中内吞作用受到抑制,玉米ZmEHD1蛋白在内吞作用中起着重要作用。实施例3、ZmEHD1蛋白的亚细胞定位一、野生型玉米中ZmEHD1蛋白的亚细胞定位1、以BamHI(NewEnglandBiolabs,R3136L)为限制性酶切位点,利用In-Fusion(takara,639648)方法将序列1所示ZmEHD1基因的CDS全长克隆到CPB载体上,得到表达载体,该表达载体表达ZmEHD1蛋白和黄色荧光蛋白(YFP)的融合蛋白(YFP-ZmEHD1融合蛋白)。2、将步骤1的表达载体导入烟草表皮细胞中,在烟草表皮细胞中表达YFP-ZmEHD1融合蛋白,YFP信号主要在膜上(图6A),与红色荧光染料FM4-64结果一致。3、用抑制剂E-64D(影响核内体和液泡的融合)处理已经注射农杆菌的烟草,检测ZmEHD1蛋白的运输。结果如图6所示。当用E-64D处理时,YFP-ZmEHD1在烟草表皮细胞聚集,而且与FM4-64信号共存,这个结果说明质膜的内在化(图6A)。二、转ZmEHD1mut玉米纯合株系中ZmEHD1mut蛋白的亚细胞定位1、以BamHI(NewEnglandBiolabs,R3136L)为限制性酶切位点,利用In-Fusion(takara,639648)方法将ZmEHD1mut基因序列克隆到CPB载体上,得到表达载体,该表达载体表达ZmEHD1mut蛋白和黄色荧光蛋白(YFP)的融合蛋白(YFP-ZmEHD1mut融合蛋白)。2、将步骤1的表达载体导入烟草表皮细胞中,在烟草表皮细胞中表达YFP-ZmEHD1mut融合蛋白,YFP信号主要在膜上(图6A),与红色荧光染料FM4-64结果一致。3、用抑制剂E-64D(影响核内体和液泡的融合)处理已经注射农杆菌的烟草,检测ZmEHD1mut蛋白的运输。结果表明:当没有用E-64D处理时,YFP-ZmEHD1mut蛋白信号在烟草表皮细胞聚集并与FM4-64信号共存(图6B)。此现象表明在转ZmEHD1mut玉米纯合株系中核内体和液泡的融合受到损坏。实施例4、ZmEHD1与ZmAP2σ亚基互作为了验证ZmEHD1是否通过与ZmAP2σ亚基互作影响网络蛋白介导的内吞作用进行了如下实验。一、ZmAP2σ影响网络蛋白介导的内吞作用1、表达载体的构建将玉米AP2σ亚基(ZmAP2σ,GRMZM2G052713)的cDNA序列(序列5)通过LR反应(invitrogen,11791020)连接到pMDC32(NTCC典型培养物保藏中心BioVector质粒载体菌种细胞基因保藏中心,pMDC32),得到重组载体。2、重组菌的制备将重组载体导入GV3101菌(普如汀生物技术(北京)有限公司,Biovector610135)中,得到重组菌。3、转ZmAP2σ拟南芥的获得将含有重组载体的农杆菌通过Floral-dip法侵染拟南芥ap2σ亚基功能缺失的突变体中,将侵染过的拟南芥进行培养,得到T0代转ZmAP2σ拟南芥种子。将T0代转ZmAP2σ拟南芥种子表面消毒后,播种于1/2MS+hyg培养基上筛选,得到T1代转ZmAP2σ拟南芥株系#1和#3,并采用引物AP2RT-1F:GAGTTCCGCACGCACAAAGT和AP2RT-1R:TTCTCAAGCTCGCCCATCCT对T1代转ZmAP2σ拟南芥株系#1和#3进行RT-PCR检测。RT-PCR检测结果表明:ZmAP2σ在拟南芥突变体中表达(图7A、图7B),ZmAP2σ基因的转入对拟南芥ap2σ亚基功能缺失突变体的表型进行了恢复(图7C)。表明ZmAP2σ与AtAP2σ在网络蛋白介导的内吞作用方面具有相似的功能,ZmAP2σ可影响网络蛋白介导的内吞作用。二、荧光双分子互补实验1、表达载体的制备(1)nYFP-ZmAP2σ载体的构建以BamHI(NewEnglandBiolabs,R3136L)为限制性酶切位点,利用In-Fusion(takara,639648)方法将序列5所示的DNA片段克隆到pSPYNE载体上,且保持pSPYNE载体的其他序列不变,得到nYFP-ZmAP2σ载体;(2)cYFP-ZmEHD1载体的构建以BamHI为限制性酶切位点,利用In-Fusion方法将序列1所示的ZmEHD1基因序列克隆到pSPYCE载体上,且保持pSPYCE载体的其他序列不变,得到cYFP-ZmEHD1载体;(3)cYFP-ZmEHD1mut载体的构建以BamHI为限制性酶切位点,利用In-Fusion方法将序列3所示的ZmEHD1mut基因序列克隆到pSPYCE载体上,且保持pSPYCE载体的其他序列不变,得到cYFP-ZmEHD1mut载体。2、重组菌的制备分别将步骤1制备的表达载体转入农杆菌EHA105(北京华越洋生物科技有限公司,GX0133-100)中,分别得到含有nYFP-ZmAP2σ载体的重组菌、含有cYFP-ZmEHD1载体的重组菌和含有cYFP-ZmEHD1mut载体的重组菌。将pSPYNE载体转入农杆菌EHA105中,得到含有pSPYNE载体的重组菌。将pSPYCE载体转入农杆菌EHA105中,得到含有pSPYCE载体的重组菌。3、烟草的注射及YFP信号检测通过农杆菌转化的方法,分别将目的基因注射到本氏烟草表皮细胞。具体步骤如下:(1)重组菌的重悬分别将含有pSPYNE载体的重组菌、含有pSPYCE载体的重组菌、含有nYFP-ZmAP2σ载体的重组菌、含有cYFP-ZmEHD1载体的重组菌和含有cYFP-ZmEHD1mut载体的重组菌、含有p19质粒的重组菌(addgene,MLCC13519)2000g离心10min后收集菌体,之后用重悬液(MgCl20.406g,MES0.426g,按照2/1000量加入AS(100mM母液),使得AS终浓度为0.2mM后,定容到200毫升,调节pH值至6.0左右)进行重悬,调节菌液OD600值至1.0,同时将含有p19质粒的农杆菌用重悬液调节OD600值至0.6。(2)重组菌的侵染将重悬后的含有nYFP-ZmAP2σ载体的重组菌、含有cYFP-ZmEHD1载体的重组菌和含有p19质粒的重组菌等体积混合,室温下黑暗孵育2h后注射本氏烟草表皮细胞。将重悬后的含有nYFP-ZmAP2σ载体的重组菌、含有cYFP-ZmEHD1mut载体的重组菌和含有p19质粒的重组菌等体积混合,室温下黑暗孵育2h后注射本氏烟草表皮细胞。将重悬后的含有pSPYNE载体的重组菌、含有cYFP-ZmEHD1载体的重组菌和含有p19质粒的重组菌等体积混合,室温下黑暗孵育2h后注射本氏烟草表皮细胞。将重悬后的含有pSPYCE载体的重组菌、含有cYFP-ZmEHD1mut载体的重组菌和含有p19质粒的重组菌等体积混合,室温下黑暗孵育2h后注射本氏烟草表皮细胞。(3)荧光信号检测将步骤(2)获得的注射重组菌的烟草正常培养3天后用LSM700激光共聚焦显微镜观察荧光信号。当nYFP-ZmAP2σ和cYFP-ZmEHD1同时注射时,可以在膜上观察到YFP信号(图8A)。同样的,当nYFP-ZmAP2σ和cYFP-ZmEHD1mut同时注射时,也可以观察到YFP信号,但信号不仅在膜上,而是扩散到细胞质(图8A)。说明ZmEHD1蛋白和ZmEHD1mut蛋白均能与ZmAP2σ亚基互作。三、酵母双杂实验1、质粒构建以Sfi-HF内切酶为限制性酶切位点,利用In-Fusion方法将序列1所示的ZmEHD1基因序列克隆到pPR3-NubG质粒上,且保持pPR3-NubG质粒的其他序列不变,得到pPR3-ZmEHD1质粒;以Sfi-HF内切酶为限制性酶切位点,利用In-Fusion方法将序列3所示的ZmEHD1mut基因序列克隆到pPR3-NubG质粒上,且保持pPR3-NubG质粒的其他序列不变,得到pPR3-ZmEHD1mut质粒;以Sfi-HF内切酶为限制性酶切位点,利用In-Fusion方法将序列5所示的DNA片段克隆到pBT3-STE质粒上,且保持pBT3-STE质粒的其他序列不变,得到pBT3-ZmAP2σ质粒。2、Bait建立(1)在YPDA固体培养基上挑选新鲜的NMY51克隆(在文献“WenwenLiu,QingSun,KaiWang,QingguoDuandWen-XueLiNitrogenLimitationAdaptation(NLA)isinvolvedinsource-to-sinkremobilizationofnitratebymediatingthedegradationofNRT1.7inArabidopsisNewPhytologist(2016)doi:10.1111/nph.14396”中公开过),接菌至50mlYPDA液体培养基中,30℃,200rpm振荡过夜培养。(2)培养OD546值至0.6-0.8。如果OD546超过1.0,则用新鲜培养基稀释至0.2后继续培养至OD546为0.6。(3)2500g离心5min,收集菌体,用2.5ml无菌水重悬,得到酵母感受态细胞。(4)将鲑鱼精DNA(北京立虹东方生物科技中心,D1626)在99℃条件下变性5min,重复一次,得到变性后的鲑鱼精DNA。在1.5mlEP管中准备PEG/LiAcmix:50%PEG-40001.2ml、1MLiAc180μl、变性后的鲑鱼精DNA125μl。(5)准备1.5μgpBT3-ZmAP2σ质粒,加入300μl步骤(4)中的PEG/LiAcmix,涡旋混匀。(6)向(5)中加入100μl步骤(1)-(3)制备的酵母感受态细胞,充分涡旋混匀1min。(7)将上述混合物42℃孵育45min。(8)700g离心5min,收集菌体。(9)用100μl0.9%的无菌NaCl重悬菌体,取适量涂布在SD-Leu培养基上,30℃培养2-4天。(10)挑选阳性克隆(含有pBT3-ZmAP2σ质粒的酵母菌NMY51),PCR检测(F:atgatccggttcatcctgctgcaga和R:tcattccagcttctcaagctcgccca)是否含有目的质粒。阳性菌落可用终浓度为25%的甘油保菌。3、Bait的自激活验证(1)将步骤2中的阳性菌落接种到10mlSD-Leu液体培养基中,30℃振荡过夜培养。(2)将过夜培养的菌体2500g离心10min,收集菌体。用YPDA液体培养基重悬后接种到50mlYPDA液体培养基中,加入培养基调整起始OD546约0.15-0.2。(3)30℃,220rpm摇菌3-5h,至OD546值为0.6-0.7。(4)准备500μl的鲑鱼精DNA,99℃变性5min,重复一次,得到变性后的鲑鱼精DNA。(5)在10ml无菌EP管中准备LiAc/TEmix8ml:10×TEbuffer0.88ml、1MLiAc0.88ml、ddH2O6.24ml。(6)在50ml无菌EP管中准备PEG/LiAcmix12ml:10×TEbuffer1.2ml、1MLiAc1.2ml、50%PEG-40009.6ml。(7)将(3)中的菌液700g离心5min,得到菌体。用无菌水重悬沉淀后700g离心5min,得到菌体。(8)用1mlLiAc/TEmix重悬沉淀,700g离心5min,去上清,取沉淀。(9)将步骤(8)获得的沉淀用1.2mlLiAc/TEmix重悬得到感受态细胞。(10)将pPR3-NubG6μg、变性后的鲑鱼精DNA100μl、步骤(9)中制备的感受态细胞600μl、PEG/LiAcmix2.5ml混合,得到混合物,并将混合物涡旋振荡1min,混合均匀。(11)30℃孵育45min,每隔15min混匀一次。(12)42℃孵育20min。700g离心15min,去上清,收集沉淀。向沉淀中加入3mlYPDA重悬。30℃,150rpm离心90min,收集沉淀。(13)700g离心5min,收集沉淀,加入2ml0.9%NaCl,取600μl涂抹平板SD-Trp-Leu、SD-Trp-Leu-His及SD-Trp-Leu-His-Ade。30℃培养4-6天,观察每种培养基上转化细胞的生长状况。若在三缺和四缺培养基上有一定量菌斑的形成,说明存在自激活现象,需要用不同浓度的3-AT进行背景抑制,具体判断方法见该试剂盒说明书33页,最终将7.5mM的3-AT作为正确浓度。4、Prey质粒的导入重复步骤3,分别将质粒pPR3-ZmEHD1及pPR3-ZmEHD1mut导入到含有pBT3-ZmAP2σ质粒的酵母菌NMY51,分别得到重组酵母菌(AP2σ-Cub+NubG-ZmEHD1)和重组酵母菌(AP2σ-Cub+NubG-ZmEHD1mut),并将最终菌体涂布在含有7.5mM3-AT的SD-Trp-Leu、SD-Trp-Leu-His及SD-Trp-Leu-His-Ade上,30℃培养4-6天,观察每种培养基上转化细胞的生长状况。选取SD-Trp-Leu-His-Ade上正常生长的白斑,进行酵母菌的菌落PCR检测。同时设置pTSU2-APPbait库的建立和pNubG-Fe65的转化作为对照。5、检测β-半乳糖苷酶的活性将在SD-Trp-Leu-His-Ade培养基上正常生长的酵母菌接种到10mlSD-Trp-Leu液体培养基中,30℃振荡培养过夜。2500g离心5min,收集菌体,用5ml0.9%NaCl重悬菌体,重复操作一次。测定OD546值。取100μl菌体,用0.9%NaCl分别稀释10倍、100倍、1000倍。取相同体积的不同浓度的菌体分别滴在SD-Trp-Leu-His-Ade、SD-Trp-Leu-Ade、SD-Trp-Leu-Ade+X-gal培养基上,30℃培养2-4天,观察菌体生长状况。结果如图8B所示。ZmEHD1和ZmEHD1mut均能与AP2σ亚基互作且互作的结果在缺His和添加β半乳糖苷酶活性的培养基上进行了验证。说明ZmEHD1和ZmEHD1mut均可与ZmAP2σ亚基互作影响植物网络蛋白介导的内吞作用。实施例5、ZmEHD1在调控植物体内生长素平衡中的应用一、转ZmEHD1mut玉米纯合株系的转录组分析为了深入探究参与ZmEHD1介导信号途径的分子机制,利用RNA-seq技术比对分析转ZmEHD1mut玉米纯合株系(OE)的转录组分析和野生型玉米植株(WT)15天胚乳的完整转录组。每个样本进行两个生物学重复,通过Illumina高通量测序技术进行测序建库。4个RNA库总共获得了超过5亿个原始reads,通过过滤去除接头污染reads、高含N比reads和低质量reads后,大约97%的reads得到保留。其中,超过1.2亿的reads能够完全与玉米B73RefGen_V3.27参考序列(ftp://ftp.ensemblgenomes.org/pub/plants/release-27/fasta/zea_mays)匹配。把不能匹配到玉米基因组的序列删除,只有完全匹配的序列进行深入的分析。以每百万reads中map到外显子每Kb的reads个数(RPKM)来检测每个基因的丰度。计算了每个处理的两个生物学重复的相关系数(R值),考察重复之间的变异性。两个处理重复间的R值都达到0.99(图9),表明两个生物学重复高度相关。依据Log2≥1并且显著值≤0.05的条件,利用DEGseq软件鉴定了4760个基因为差异表达基因(DEGs),其中包括ZmEHD1。对转ZmEHD1mut玉米纯合株系而言,这些差异基因中有2208个基因上调表达及2551个基因下调表达。通过实时荧光定量PCR来确定RNA-seq的结果。GRMZM2G088273、GRMZM2G346897、GRMZM2G067929和GRMZM2G418119表达水平与RNA-seq数据一致,在转ZmEHD1mut玉米纯合株系中表达量低于野生型(图10A)。如预期的一样,转ZmEHD1mut玉米纯合株系中GRMZM2G156877和GRMZM2G420988的表达量高于野生型(图10A),表明了RNA-seq数据真实可靠。然后进行了基因本体分析(GO;http://bioinfo.cau.edu.cn/agriGO/),确定与差异表达基因相关的分子机制。GO分析结果表明4760个DEGs高度富集于几个生物学过程,包括应激反应(GO:0006950,P=1.2e-12)、integraltomembrane(GO:0016021,P=1e-5)、intracellularmembrane-boundedorganelle(GO:0043231,P=0.0002)、液泡(GO:0005773,P=0.0007)、淀粉代谢过程(GO:0005982,P=1.6e-5)和糖类代谢过程(GO:0006950,P=1.2e-12)。一些DEGs具有已知或被推测的参与生长素介导信号途径(GO:0009734,P=1.2e-25)和生长素刺激响应(GO:0009733,P=2.1e-25)的功能,其中包括生长素响应因子、AUX/IAA转录因子、生长素上调小RNA(SAUR)、吲哚乙醛氧化酶、生长素运输蛋白和输出载体(表1)。随机选择表1中的四个DEGs(GRMZM2G082943、GRMZM2G365188、GRMZM5G809195和GRMZM2G019799),通过实时荧光定量验证了他们在转ZmEHD1mut玉米纯合株系和野生型中的不同表达水平(图10B)。这些结果表明ZmEHD1可能影响玉米中生长素平衡。表1涉及植物生长素过程的差异表达基因二、ZmEHD1在调控植物体内生长素平衡中的应用1、野生型和转ZmEHD1mut玉米纯合株系的IAA含量检测为了验证ZmEHD1是否影响玉米生长素体内平衡,测定了野生型玉米植株(野生型玉米自交系Qi319)(WT)和转ZmEHD1mut玉米纯合株系OE3(OE)授粉后15天籽粒的IAA含量。IAA含量测定的具体步骤参照文献“Wang,B.,Chu,J.,Yu,T.,Xu,Q.,Sun,X.,Yuan,J.,Xiong,G.,Wang,G.,Wang,Y.,andLi,J.(2015)Tryptophan-independentauxinbiosynthesiscontributestoearlyembryogenesisinArabidopsis.ProcNatlAcadSciUSA112:4821-4826.”中的方法。结果表明:转ZmEHD1mut玉米纯合株系(OE)籽粒中的IAA含量为2,558pgmg-1FW,和野生型玉米植株(WT)相比,转ZmEHD1mut玉米纯合株系OE3籽粒中的IAA含量降低了约30%(图11A)。说明ZmEHD1蛋白可以调控植物体内生长素的含量。2、外源施加1-NAA可以提高转ZmEHD1mut玉米纯合株系的发芽率通过外源施加有活性的生长素化合物(1-NAA,1-naphthaleneaceticacid)来进一步验证转ZmEHD1mut玉米纯合株系发芽率低是由生长素体内平衡失调造成。具体步骤如下:分别用如下浓度:5mg/L、10mg/L、20mg/L、30mg/L的1-NAA(PhytoTechnologyLaboratories,N600)水溶液和如下浓度:10mg/L、20mg/L、30mg/L、50mg/L的GA3(SIGMA-ALORICH,G7645)水溶液浸泡转ZmEHD1mut玉米纯合株系OE3种子12个小时,然后纸培(参照实施例2中的纸培方法),培养4天后,检测种子发芽率。同时以水作为对照。30mgL-11-NAA水溶液和水均没有提高转ZmEHD1mut玉米纯合株系种子的发芽率(图11B,C)。但是当施加低浓度的1-NAA时,转ZmEHD1mut玉米纯合株系OE发芽率明显提高,其中,施加10mgL-11-NAA最为明显(图11B)。同时,外源施加低浓度的1-NAA能使转ZmEHD1mut玉米纯合株系长到两片完全展开叶时期。而外源施加GA3并不能提高转ZmEHD1mut玉米纯合株系的发芽率(图12)。上述结果表明:外源施加1-NAA可以提高转ZmEHD1mut玉米纯合株系的发芽率。转ZmEHD1mut玉米纯合株系生长缺陷表型是由生长素体内平衡丧失引起的,说明ZmEHD1蛋白具有调控植物体内生长素平衡的功能。序列表<110>河南农业大学<120>一种调控植物体内生长素平衡的EHD1蛋白及其应用<160>5<210>1<211>1644bp<212>DNA<213>人工序列<220><223><400>1atggatatcgcgcgggcggctgctggcgtggcctgctgcaaggagcaccagaggatctac60gccgagtggttcgccctggccgacccagatggcgacggccgcgtcacgggcgccgatgcc120accagcttcttcgccatgtccgcgctttcacgcgccgatctcaagcaggtgtgggcgatc180gcggattccaggcggcaggggtacctcggattcgccgagttcgtcacagcaatgcagctc240gtgtctctggcccaagcagggaacgagatcacacaggacagtctgaaacgtgaagacctg300agcaccttggatccgcctgtgatggaaggtgttgatgagctactggctagatccaaggct360gttgtgaagagagttcacccagatgacaatggcaccactcaggcccaagcaccttccata420tatcactggtttggttcaaaatcagcacagaaggtgcagatgcctctgactgctgttacc480tctgtaattgatggcttgaaaagactctatgttgaaaaactgaagcctttggaagtcgcc540tacagatacaatgatttttctcacccattattgacaaacagtgattttgaagcaaagcca600atggttatgctcttgggtcagtattcaacagggaaaacaacattcattaagcatctgcta660aagacaagctacccaggagctcacgtcggaccagagccaacaactgatagattcgtagtt720gtcatgtcaggatctgatggaaggaccattcctggcaatactattgccgttcaagccgac780atgccattcactggtcttacaacctttgggggagcgtttctgtcgaagtttgaatgctct840cagatgccacatcccctgctagagcatatcacctttgtggacactcctggtgttctctct900ggggaaaagcaacggacccaacgtagctatgatttcactggtgtgacctcatggtttgct960gccaagtgtgaccttatccttcttctgtttgaccctcacaagcttgatataagtgatgag1020ttcaaacgtgtcatttcatctttacgtgggcatgatgacaaaatacgcgtagtgttgaac1080aaggccgaccaagttgacacacaacagcttatgagagtgtatggtgcattgatgtggtct1140cttgggaaagtgctaaatactcctgaggttgtgcgtgtgtatatcggatcatttaatgat1200aagccagtgaatgattcagctgttgggccaattggaaaagacttgtttgagagggagcaa1260gatgacctcctctgtgacctgaaggacattcctaagaaagcctgtgatcgacgggtcaat1320gagtttgttaagcgtgcccgagctgccaagatccatgcttacataattggccatctgaag1380aaggagatgcctgcaatgatgggaaaagccaaggctcagcagcgactcatagacaactta1440gacgaggagtttgcaaaggtgcagagggaataccatctccccgccggagactttcccgac1500gtcgagcacttcaagcaggtgctgggcgggtacagcatcgacaagttcgagaagctgaag1560ccgaagatggtgcaggcggtggacgacatgctcgctcacgacatcccggagctcctcaag1620aacttgcggaacccgtacgagtga1644<210>2<211>547<212>PRT<213>人工序列<220><223><400>2MetAspIleAlaArgAlaAlaAlaGlyValAlaCysCysLysGluHis151015GlnArgIleTyrAlaGluTrpPheAlaLeuAlaAspProAspGlyAsp202530GlyArgValThrGlyAlaAspAlaThrSerPhePheAlaMetSerAla354045LeuSerArgAlaAspLeuLysGlnValTrpAlaIleAlaAspSerArg505560ArgGlnGlyTyrLeuGlyPheAlaGluPheValThrAlaMetGlnLeu65707580ValSerLeuAlaGlnAlaGlyAsnGluIleThrGlnAspSerLeuLys859095ArgGluAspLeuSerThrLeuAspProProValMetGluGlyValAsp100105110GluLeuLeuAlaArgSerLysAlaValValLysArgValHisProAsp115120125AspAsnGlyThrThrGlnAlaGlnAlaProSerIleTyrHisTrpPhe130135140GlySerLysSerAlaGlnLysValGlnMetProLeuThrAlaValThr145150155160SerValIleAspGlyLeuLysArgLeuTyrValGluLysLeuLysPro165170175LeuGluValAlaTyrArgTyrAsnAspPheSerHisProLeuLeuThr180185190AsnSerAspPheGluAlaLysProMetValMetLeuLeuGlyGlnTyr195200205SerThrGlyLysThrThrPheIleLysHisLeuLeuLysThrSerTyr210215220ProGlyAlaHisValGlyProGluProThrThrAspArgPheValVal225230235240ValMetSerGlySerAspGlyArgThrIleProGlyAsnThrIleAla245250255ValGlnAlaAspMetProPheThrGlyLeuThrThrPheGlyGlyAla260265270PheLeuSerLysPheGluCysSerGlnMetProHisProLeuLeuGlu275280285HisIleThrPheValAspThrProGlyValLeuSerGlyGluLysGln290295300ArgThrGlnArgSerTyrAspPheThrGlyValThrSerTrpPheAla305310315320AlaLysCysAspLeuIleLeuLeuLeuPheAspProHisLysLeuAsp325330335IleSerAspGluPheLysArgValIleSerSerLeuArgGlyHisAsp340345350AspLysIleArgValValLeuAsnLysAlaAspGlnValAspThrGln355360365GlnLeuMetArgValTyrGlyAlaLeuMetTrpSerLeuGlyLysVal370375380LeuAsnThrProGluValValArgValTyrIleGlySerPheAsnAsp385390395400LysProValAsnAspSerAlaValGlyProIleGlyLysAspLeuPhe405410415GluArgGluGlnAspAspLeuLeuCysAspLeuLysAspIleProLys420425430LysAlaCysAspArgArgValAsnGluPheValLysArgAlaArgAla435440445AlaLysIleHisAlaTyrIleIleGlyHisLeuLysLysGluMetPro450455460AlaMetMetGlyLysAlaLysAlaGlnGlnArgLeuIleAspAsnLeu465470475480AspGluGluPheAlaLysValGlnArgGluTyrHisLeuProAlaGly485490495AspPheProAspValGluHisPheLysGlnValLeuGlyGlyTyrSer500505510IleAspLysPheGluLysLeuLysProLysMetValGlnAlaValAsp515520525AspMetLeuAlaHisAspIleProGluLeuLeuLysAsnLeuArgAsn530535540ProTyrGlu545<210>3<211>1644bp<212>DNA<213>人工序列<220><223><400>3atggatatcgcgcgggcggctgctggcgtggcctgctgcaaggagcaccagaggatctac60gccgagtggttcgccctggccgacccagatggcgacggccgcgtcacgggcgccgatgcc120accagcttcttcgccatgtccgcgctttcacgcgccgatctcaagcaggtgtgggcgatc180gcggattccaggcggcaggggtacctcggattcgccgagttcgtcacagcaatgcagctc240gtgtctctggcccaagcagggaacgagatcacacaggacagtctgaaacgtgaagacctg300agcaccttggatccgcctgtgatggaaggtgttgatgagctactggctagatccaaggct360gttgtaaagagagttcacccagatgacaatggcaccactcaggcccaagcaccttccata420tatcactggtttggttcaaaatcagcacagaaggtgcagatgcctctgactgctgttacc480tctgtaattgatggcttgaaaagactctatgttgaaaaactgaagcctttggaagtcgcc540tacagatacaatgatttttctcacccattattgacaaacagtgactttgaagcaaagcca600atggttatgctcttgggtcagtattcaacaggcaaaacaacattcattaagcatctgcta660aagacaagctacccaggagctcacgtcggaccagagccaacaactgatagattcgtagtt720gtcatgtcaggatctgatggaaggaccattcctggcaatactattgccgttcaagccgac780atgccattcactggtcttacaacctttgggggagcgtttctgtcgaagtttgaatgctct840cagatgccacatcccctgctagagcatatcacctttgtggacactcctggtgttctctct900ggggaaaagcaacggacccaacgtagctatgatttcactggtgtgacctcatggtttgct960gccaagtgtgaccttatccttcttctgtttgaccctcacaagcttgatataagtgatgag1020ttcaaacgtgtcatttcatctttacgtgggcatgatgacaaaatacgcgtagtgttgaac1080aaggccgaccaagttgacacacaacagcttatgagagtgtatggtgcattgatgtggtct1140cttgggaaagtgctaaatactcctgaggttgcgcgtgtgtatatcggatcatttaatgat1200aagccagtgaatgattcagctgttgggccaattggaaaagacttgtttgagagggagcaa1260gatgacctcctctgtgacctgaaggacattcctaagaaggcttgtgatcgacgggtcaat1320gagtttgttaagcgtgcccgagctgccaagatccatgcttacataattggccatctgaag1380aaggagatgcctgcagtgatgggaaaagccaaggctcagcagcgactcatagacaactta1440gaggaggagtttgcaaaggtgcagagggaatatcatctccccgccggagacttccccgac1500gtcgagcacttcaagcaggtgctgggcgggtacagcatcgacaagttcgagaagctgaag1560ccgaagatggtgcaggcggtggacgacatgctcgctcacgacatcccggagctcctcaag1620aacttgcggaacccgtacgagtga1644<210>4<211>547<212>PRT<213>人工序列<220><223><400>4MetAspIleAlaArgAlaAlaAlaGlyValAlaCysCysLysGluHis151015GlnArgIleTyrAlaGluTrpPheAlaLeuAlaAspProAspGlyAsp202530GlyArgValThrGlyAlaAspAlaThrSerPhePheAlaMetSerAla354045LeuSerArgAlaAspLeuLysGlnValTrpAlaIleAlaAspSerArg505560ArgGlnGlyTyrLeuGlyPheAlaGluPheValThrAlaMetGlnLeu65707580ValSerLeuAlaGlnAlaGlyAsnGluIleThrGlnAspSerLeuLys859095ArgGluAspLeuSerThrLeuAspProProValMetGluGlyValAsp100105110GluLeuLeuAlaArgSerLysAlaValValLysArgValHisProAsp115120125AspAsnGlyThrThrGlnAlaGlnAlaProSerIleTyrHisTrpPhe130135140GlySerLysSerAlaGlnLysValGlnMetProLeuThrAlaValThr145150155160SerValIleAspGlyLeuLysArgLeuTyrValGluLysLeuLysPro165170175LeuGluValAlaTyrArgTyrAsnAspPheSerHisProLeuLeuThr180185190AsnSerAspPheGluAlaLysProMetValMetLeuLeuGlyGlnTyr195200205SerThrGlyLysThrThrPheIleLysHisLeuLeuLysThrSerTyr210215220ProGlyAlaHisValGlyProGluProThrThrAspArgPheValVal225230235240ValMetSerGlySerAspGlyArgThrIleProGlyAsnThrIleAla245250255ValGlnAlaAspMetProPheThrGlyLeuThrThrPheGlyGlyAla260265270PheLeuSerLysPheGluCysSerGlnMetProHisProLeuLeuGlu275280285HisIleThrPheValAspThrProGlyValLeuSerGlyGluLysGln290295300ArgThrGlnArgSerTyrAspPheThrGlyValThrSerTrpPheAla305310315320AlaLysCysAspLeuIleLeuLeuLeuPheAspProHisLysLeuAsp325330335IleSerAspGluPheLysArgValIleSerSerLeuArgGlyHisAsp340345350AspLysIleArgValValLeuAsnLysAlaAspGlnValAspThrGln355360365GlnLeuMetArgValTyrGlyAlaLeuMetTrpSerLeuGlyLysVal370375380LeuAsnThrProGluValAlaArgValTyrIleGlySerPheAsnAsp385390395400LysProValAsnAspSerAlaValGlyProIleGlyLysAspLeuPhe405410415GluArgGluGlnAspAspLeuLeuCysAspLeuLysAspIleProLys420425430LysAlaCysAspArgArgValAsnGluPheValLysArgAlaArgAla435440445AlaLysIleHisAlaTyrIleIleGlyHisLeuLysLysGluMetPro450455460AlaValMetGlyLysAlaLysAlaGlnGlnArgLeuIleAspAsnLeu465470475480GluGluGluPheAlaLysValGlnArgGluTyrHisLeuProAlaGly485490495AspPheProAspValGluHisPheLysGlnValLeuGlyGlyTyrSer500505510IleAspLysPheGluLysLeuLysProLysMetValGlnAlaValAsp515520525AspMetLeuAlaHisAspIleProGluLeuLeuLysAsnLeuArgAsn530535540ProTyrGlu545<210>5<211>429bp<212>DNA<213>人工序列<220><223><400>5atgatccggttcatcctgctgcagaaccggcaggggaaaacacggctggccaagtactat60gtcgctctcgaggactccgagaagcacaaggttgagtacgaggtgcatcggctcgtggtc120aaccgggaccccaagttcaccaacttcgtcgagttccgcacgcacaaagttatatatagg180agatatgcaggactttttttctctatatgtgtggacatcaccgacaacgaattggcatac240ttggaatgtatccatctgtttgttgagatattggaccatttcttcagcaatgtgtgtgaa300cttgatttagtatttaatttccacaaggtttacttgatattggatgagttcattcttgct360ggagagcttcaagaaacaagcaaaagggcaattatagagaggatgggcgagcttgagaag420ctggaatga429当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1