猪流行性腹泻病毒M蛋白特异性重链抗体的制作方法
本发明属于生物技术领域,具体涉及以一种猪流行性腹泻病毒m蛋白单域抗体。
背景技术:
猪流行性腹泻(porcineepidemicdiarrhea,ped)是由猪流行性腹泻病毒(pedv)感染引起的猪的一种高度接触性肠道传染病。感染pedv的病猪主要通过粪便传播病毒,发病猪主要表现腹泻、呕吐和脱水等临床特征,是导致哺乳仔猪死亡的主要传染病之一。1971年,ped首次在英国发生,随后扩散到世界各地。我国于1980年首次分离到pedv,2010年国内大部分养猪地区发生ped疫情,从哺乳仔猪到繁殖母猪均有发病,给养猪户造成了严重地经济损失。pedv属于尼多病毒目(nidovirales)冠状病毒科(coronaviridae)冠状病毒属(coronavirus),基因组是单股正链具有感染性的rna,与其他冠状病毒相似,基因组5′端有一个帽子结构(cap),3′端有1个poly(a)尾,基因组全长为2,8033nt,分别编码s、e、m和n等4个病毒结构蛋白。s担保存在的中和表位能够诱导机体产生中和抗体,对仔猪具有良好的保护效果。pedv感染早期,机体能够产生抗n和n蛋白的高水平抗体,是研发病毒血清抗体诊断方法的候选抗原。e蛋白在抗病毒研究方面具有应用前景。
骆驼属外周血中存在的天然缺失轻链的重链抗体(variabledomainoftheheavychainofhcabs,vhh)具有其他陆生生物所不具备的生物学特征,它的抗原结合位点仅由重链可变区单一结构域组成,却同普通igg抗体一样保持着良好和特异的抗原结合能力。vhh是目前可以得到的、具有完整功能的、最小igg抗体分子片段,分子量为15kda,是常规igg的1/10,单克隆抗体的1/3,分子高度4.8nm,直径2.2纳米,也称为纳米抗体(nanobody)。vhh抗体抗原结合位点具有伸展的抗原互补结合区和组织穿透性,可结合普通igg抗体无法接触的抗原表位。另外,体外重组vhh具有易表达和水溶性好等独特性质,在作为小型化基因工程抗体、小分子抗体药物和高灵敏度诊断试剂研发领域具有广阔应用前景。
技术实现要素:
本发明的目的是提供一种猪流行性腹泻病毒m蛋白特异性重链抗体;
本发明的另一个目的是提供一种编码所述猪流行性腹泻病毒m蛋白特异性重链抗体的核苷酸序列;
本发明的再一个目的是提供所述的猪流行性腹泻病毒m蛋白特异性重链抗体或其编码序列在e.coli中的表达。
本发明采用以下技术方案,一种猪流行性腹泻病毒m蛋白特异性重链抗体,编码所述抗体的核苷酸序列为:seqidno.1,seqidno.2,seqidno.3,seqidno.4,seqidno.5,seqidno.6,seqidno.7或seqidno.8。
一种重组表达载体,含有上述的核苷酸序列。
一种宿主细胞,所述的宿主细胞为含有上述的核苷酸序列的原核细胞。
优选的,所述宿主细胞为含有上述表达载体的原核细胞。
所述猪流行性腹泻病毒m蛋白特异性重链抗体或其编码序列在e.coli中的表达。
本发明的效果在于:利用噬菌体展示技术构建pedvm蛋白双峰驼源免疫文库,筛选获得8株来自双峰驼igg重链抗体高变区抗m蛋白vhh抗体基因编码序列,对其进行可溶性表达和elisa实验证明其均可以同m蛋白相结合,吸光度od405值的不同表明其同m蛋白结合能力各有差异。8株vhh抗体可通过e.coli的表达,而制备体外重组的基因工程抗体。
附图说明
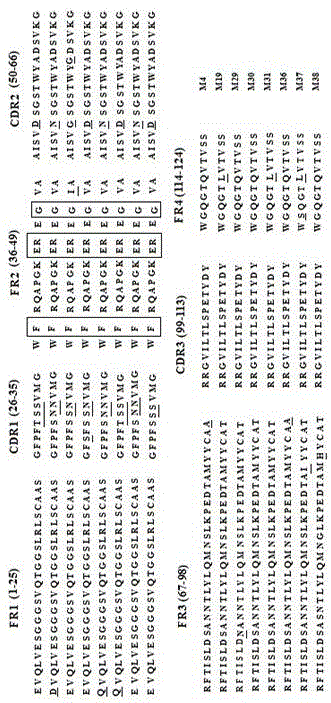
图1是是抗pedvm蛋白vhh克隆氨基酸序列比对结果图;
图2是e.coli中抗pedvm蛋白vhh抗体sds-page分析图。
具体实施方式
下面的实施例可以进一步说明本发明,但不以任何方式限制本发明。
实施例1、猪流行性腹泻病毒m蛋白特异性重链抗体的制备
1.材料和方法
1.1实验动物和试剂
2峰10月龄雌性双峰驼作为实验动物,牛淋巴细胞分离液,96孔酶标板;限制性内切酶,dna片段回收试剂盒、质粒提取试剂盒为omega产品,镍亲和层析树脂(qiagen);e.coli重组表达pedvm抗原(his标签和gst标签);菌株、辅助噬菌体和试剂大肠杆菌tg1和bl21star(de3)、辅助噬菌体m13k07、载体phen2购于neb公司,其它试剂均为国产分析纯。
1.2方法
1.2.1骆驼免疫和抗体监测
首次免疫,用206佐剂(购于法国seepic公司),乳化500μg重组m蛋白皮下多点(背部,后腿和颈部两侧肌肉)接种骆驼。首免21天后加强免疫,剂量为206乳化1000μgm蛋白,与首免间隔35天、42天和63天加强免疫,抗原接种剂量均为1000μgm蛋白。在首次免疫前和每次加强免疫前均采集静脉血并分离到血清,用m蛋白(gst标签)作为抗原的间接elisa检测pedv抗体。本次免疫实验过程中,骆驼饲养管理,免疫,血液样品的采集均在专业兽医工作人员监督指导下完成。
1.2.2外周血的分离
最后一次免疫后7天(即第63天)颈静脉采抗凝血10ml(抗凝剂为肝素钠,10iu/ml),加入等量的han`s液做1倍稀释,而后加入等体积淋巴细胞分离液,进行密度梯度离心分离外周血总淋巴细胞。分离的淋巴细胞用mem重悬中加入到六孔板中(37℃,5%co2)静止30min,吸附除去细胞碎片和巨噬细胞,离心收集淋巴细胞加入mem,保存在液氮中用于细胞rna提取。
1.2.3vhh抗体免疫文库构建
设计3对引物(参见表1-vhh基因扩增用引物)通过三轮扩增得到编码vhh蛋白目的基因。通过trizol试剂抽提淋巴细胞总rna,用g1作为反转录引物,合成第一链cdna,以cdna为模板,h1、g1为引物扩增得到600bp和900bp大小2条dna带,回收600bp条带作为模板,进行第2轮pcr扩增。
以第2轮pcr产物为模板,h2、h3为引物得到450bpdna带。以2轮pcr产物为模板,p1,tn为上下游引物进行第3轮扩增得到450bp带。将第3轮pcr产物经过sfii/noti双酶切克隆到phen2载体,电转化方法将连接产物转化到tg1中,37℃,180r/min培养1h,离心浓缩后涂在氨苄抗性的2×ytg平板(2%葡萄糖,100μg/mlampr)。同时取10μl稀释至1×104倍涂板,37℃过夜培养,计算库容。转化后平板37℃温箱中培养16小时,用10ml的2×yt培养基将培养板上长出的菌苔刮洗干净,加入终浓度25%的甘油,分装保存在-70℃备用,此为获得的抗pedvm蛋白vhh免疫抗体库。随机挑选电转化后的克隆进行测序,根据多样性和克隆数目来计算库容量。
1.2.4通过二次筛选过程获得特异的抗pedvm蛋白vhh抗体
筛选抗体操作步骤如下:
(1)抗原包被:将m抗原(gst标签)稀释至20ug/ml,50ul/孔,4℃过夜。pbs洗3次;
(2)封闭:加入300ul/孔的2%m-pbs,与37℃封闭1h,pbs洗3次;
(3)结合:取20ulphage+180ul0.1mpbs,混匀,50ul/孔,37℃静置1h;甩出多余的噬菌体溶液,并倒置平板在纸巾上拍甩除去残液,0.1%pbst洗3次;
(4)一抗:用0.1mpbs按1:1000稀释兔抗m1350ul/孔37℃静置1h;甩出多余的液体,倒置平板在纸巾上拍甩除去残液,0.1%pbst洗3次;
(5)二抗:用0.1mpbss按1:10000稀释hrp-羊抗兔50ul/孔37℃静置1h;甩出多余的液体倒置平板在纸巾上拍甩除去残液,0.1%pbst洗4次;
(6)显色:取0.2m/lna2hpo4+0.1m/l柠檬酸各0.5ml加入少量opd,10ulh2o2混匀50ul/孔;2m硫酸终止25ul/孔,波长490nm检测结果。
1.2.5vhh可溶表达和elisa检测
随机挑选40个克隆,转化到e.colibl21star(de3)表达感受态中,然后挑单克隆于1ml培养基中,快速诱导表达,tg1空菌体作为空白对照。收集诱导的1ml菌液,12000rpm离心2min,弃上清,沉淀用0.5mltbs重悬;400w(6s超声,4是间隔)超声10min;将加入超声上清50ul/孔,37℃静置1h,tbs洗3次;加入2%m-tbs,200ul/孔,37℃封闭1h,0.1%tbst洗4次;取1m二乙醇胺+0.5mmmgcl2加入4mgpnpp,25ul/孔,3mnaoh终止,25ul/孔。波长405nm条件检测结果。选择阳性反应克隆进行序列测定和比对。
2.结果
2.1骆驼血清中m抗体水平检测
以2峰10月龄雌性双峰驼作为实验对照动物。
四次m抗原(his标签)免疫骆驼血清抗体做1∶50稀释检测表明(参见表2-m蛋白抗体检测(od450nm)),第2次加强免疫后,2头实验对照动物抗m的抗体已经是阳性,最后一次免疫后7天测抗体效价时,2峰骆驼的血清抗体od450nm达到1.58和1.92,因此可以判断此时机体内体液和细胞免疫水平均处在持续上升阶段,符合采集外周血淋巴细胞的实验要求。
2.2抗m蛋白vhh免疫文库构建和筛选
经过四轮的筛选后,可以检测到较为明显的富集效应(参见表3-每一轮噬菌体筛选的富集指数)。单克隆噬菌体elisa实验结果显示了40个阳性克隆,每个克隆的od450nm值见表4,表4中m13ko7、1%m-pbs为阴性对照。对这些克隆进行dna测序和核苷酸序列比对得到8个vhh基因碱基序列,通过氨基酸比对后获得6个独特型抗m蛋白vhh抗体基因。
2.3vhh可溶表达和elisa检测
利用m蛋白作为抗原包被酶标板,在30℃和37℃条件下同获得的6个独特型抗m蛋白vhh抗体基因进行可溶性表达和elisa检测发现,他们均可以同m蛋白相结合,通过吸光度值(od405nm)的高低说明其与抗原的结合能力各有不同(参见表5-m蛋白elisa鉴定)。
2.4抗m蛋白vhh基因序列比对和在e.coli中的表达
通过对所获得vhh基因进行核苷酸测序和序列分析见图1,氨基酸序列比对结果见图2。通过thekabatetal.(kabat,1991)比对方法进行分析,图1方框中氨基酸表示双峰驼源vhh抗体特有的标志性氨基酸,带下划线的氨基酸表示不同vhh克隆间的差异氨基酸。为了稳定获得vhh抗体,分别将vhh基因序列克隆到pet-28a载体,转入e.colibl21(de3)感受态细胞中,1.0mmiptg进行诱导表达,sds-page蛋白电泳分析发现目的条带,分子量大小在25kda附近(vhh约13.6kda,组氨酸标签约8~10kda),目的条带如图中箭头所示,证实所有vhh抗体均以成功表达。
通过图1和表4说明:利用噬菌体展示技术筛选得到8株编码pedvm蛋白vhh抗体序列,氨基酸序列比对结果表明,抗体fr2区序列具有典型的重链抗体氨基序列特征,即序列中的氨基酸残基分别为f37/e44/r45/g47(图1),证明这些抗体序列来自于重链抗体的高变区。抗体的cdr1区氨基酸残基31s和或n、32s和或n位,cdr2氨基酸残基54d或n存在变异,说明编码8株抗体的氨基酸序列的多样性存在显著差异。
序列表
<110>中国农业科学院兰州兽医研究所
<120>一种猪流行性腹泻病毒m蛋白特异性重链抗体
<130>2015
<160>8
<170>patentinversion3.3
<210>1
<211>372
<212>dna
<213>camelusbactrianus
<400>1
gaggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctgggttcccctttactagctccgtcatgggatggttccgccaggct120
ccagggaaggaacgcgagggggtcgcagctatttcggttgatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccaacaacacgctgtat240
ctgcaaatgaacagcctgaaacctgaggacactgccatgtactactgtgcggctagacgt300
ggagttattcttacactaagcccagagacctatgactactggggccaggggacccaggtc360
accgtctcctca372
<210>2
<211>372
<212>dna
<213>camelusbactrianus
<400>2
gatgtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctggattcccctttagtaacaacgtcatgggctggttccgccaggct120
ccagggaaggaacgcgagggggtcgcagctatttcggttaatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccaacaacacgctgtat240
ctgcaaatgaacagcctgaaacctgaggacactgccatgtactactgtgcgactagacgt300
ggagttattcttacactaagcccagagacctatgactactggggccaggggaccctggtc360
accgtctcctca372
<210>3
<211>372
<212>dna
<213>camelusbactrianus
<400>3
gaggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctggattcccctttagtagcaacgtcatgggctggttccgccaggct120
ccagggaaggaacgcgaggggatcgcagctatttcggttggtagtggtagcacatggtat180
ggcgactccgtgaagggccgattcaccatctccctggacaacgccaacaacacgctgtat240
ctccaaatgaacagcctgaaacctgaggacactgccatgtactactgtgcgactagacgt300
ggagttattcttacactaagcccagagacctatgactactggggccaggggacccaggtc360
accgtctcctca372
<210>4
<211>372
<212>dna
<213>camelusbactrianus
<400>4
gaggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctgggttctcctttagtagcaacgtcatgggctggttccgccaggct120
ccagggaaggaacgcgagggggtcgcagctatttcggttgatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccaacaacacgctgtat240
ctgcaaatgaacagcctgaaacctgaggacactgccatgtactactgtgcgactagacgt300
ggagttattcttacactgagcccagagacctatgactactggggccaggggacccaggtc360
accgtctcctca372
<210>5
<211>372
<212>dna
<213>camelusbactrianus
<400>5
caggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctggattcccctttagtaacaacgtcatgggctggttccgccaggct120
ccagggaaggaacgcgagggggtcgcagctatttcggttaatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccaacaacacgctgtat240
ctgcaaatgaacagcctgaaacctgaggacactgccatgtactactgtgcgactagacgt300
ggagttattcttacactaagcccagagacctatgactactggggccaggggaccctggtc360
accgtctcctca372
<210>6
<211>372
<212>dna
<213>camelusbactrianus
<400>6
caggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctgggttcccctttactagctccgtcatgggatggttccgccaggct120
ccagggaaggaacgcgagggggtcgcagctatttcggttgatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccaacaacacgctgtat240
ctgcaaatgaacagcctgaaacctgaggacactgccatgtactactgtgcggctagacgt300
ggagttattcttacactaagcccagagacctatgactactggggccaggggacccaggtc360
accgtctcctca372
<210>7
<211>372
<212>dna
<213>camelusbactrianus
<400>7
gaggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctggattcccctttagtaacaacgtcatgggctggttccgccaggct120
ccagggaaggaacgcgagggggtcgcagctatttcggttaatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccaacaacacgctgtat240
ctgcaaatgaacagcctgaaacctgaggacactgccatttactactgtgcgactagacgt300
ggagttattcttacactaagcccagagacctatgactactggagccaggggaccctggtc360
accgtctcctca372
<210>8
<211>372
<212>dna
<213>camelusbactrianus
<400>8
gaggtgcagctggtggagtctgggggaggctcggtgcagactggagggtctctgagactc60
tcctgcgcagcctctggattcccctttagtagcagcgtcatgggctggttccgccaggct120
ccaggaaaggaacgcgagggggtcgcagctatttcggtagatagtggtagcacatggtat180
gccgactccgtgaagggccgattcaccatctccctggacagcgccagcaacacactgtat240
ctgcaaatgaacggcctgaaacctgaggacactgccatgcactactgtgcgactagacgt300
ggagttattcttacactaagcccagagacctatgactactggggccaggggacccaggtc360
accgtctcctca372
- 还没有人留言评论。精彩留言会获得点赞!