具有磷脂酶C活性的多肽及其用途的制作方法
本申请提供了具有磷脂酶c活性的多肽及其用途。
背景技术:
脱胶是油脂精炼的重要步骤,传统的水化脱胶法经济成本高,物料能耗大,环境污染重,所以近些年来,很多工作致力于将酶法脱胶用于油脂精炼中的脱胶环节。同传统方法相比,酶法脱胶能够提高经济效益,实现节能减排,对生态环境污染少,在环保、经济、质量等方面具有较大的优势。油脂脱胶中所用的一种酶为磷脂酶。同其他脱胶酶相比,磷脂酶c(plc)表现出更大的优势,例如,增加甘二酯(dag)的得率,以及减少得油量的损失。
蜡样芽胞杆菌(bacilluscereus)的磷脂酰胆碱特异性磷脂酶c(bc-pc-plc)是研究较早的一种磷脂酶c。bc-pc-plc全长为283个氨基酸,其中包含24个氨基酸的信号肽和14个氨基酸的前导肽,成熟肽为245个氨基酸(参见,例如johansen,t.,holm,t.,guddal,p.h.,sletten,k.,haugli,f.b.,little,c.(1988)."cloningandsequencingofthegeneencodingthephosphatidylcholine-preferringphospholipasecofbacilluscereus."gene65(2):293-304)。
先前有文献报道,野生型bc-pc-plc的活性位点为glu4、asp55、tyr56、glu146、ser64、thr65、phe66、phe70、ile80、thr133、asn134、leu135、ser143(参见,例如hough.,e.,hansen,l.k.,birknes,b.,jynge,k.,hansen,s.,hordvik,a.,little,c.,dodson,e.,derewenda,z.(1989)"high-resolution(1.5a)crystalstructureofphospholipasecfrombacilluscereus."nature.338:357-60)。
bc-pc-plc的晶体结构已有报道,其由多个螺旋结构域组成,催化位点为55位天冬氨酸,并且含有至少三个zn2+结合位点(参见,例如hough.,e.,hansen,l.k.,birknes,b.,jynge,k.,hansen,s.,hordvik,a.,little,c.,dodson,e.,derewenda,z.(1989)"high-resolution(1.5a)crystalstructureofphospholipasecfrombacilluscereus."nature.338:357-60)。bc-pc-plc的异源表达研究较少,已有报道涉及在枯草芽孢杆菌(bacillussubtilis)和毕赤酵母(pichiapastoris)中表达bc-pc-plc(参见,例如durban,m.a.,silbersack,j.,schweder,t.,schauer,f.,bornscheuer,u.t.(2007)highlevelexpressionofarecombinantphospholipasecfrombacilluscereusinbacillussubtilis.applmicrobiolbiotechnol74(3):634-639;以及seo,k.h,rheej.i.(2004)high-levelexpressionofrecombinantphospholipasecfrombacilluscereusinpichiapastorisanditscharacterization.biotechnollett26(19):1475-1479)。
但现有报道的磷脂酶c的热稳定性不高,不能耐受超过60℃,使其最适的脱胶温度需要控制在50℃,因而使其在工业应用中具有一定的局限性。如能有效提高磷脂酶c的热稳定性,有利于磷脂酶c的工业应用,具体包括:首先脱胶时将温度提高,有利于降低油的粘度,提高油脂和磷脂的分离程度,减少磷脂中夹带的油脂,进一步提高油脂的得率;其次目前有采用plc与pla1联合使用进行深度脱胶,发明专利cn201480017114.5中就提到了一种新的pla1其最适的脱胶温度在65℃,因此更耐热的plc有利于与pla1进行联合脱胶,另外,有时毛油在储藏过程中特别是在高温天气,毛油的温度会超过50℃,因此如果plc的最适反应温度低于50℃还需要大量冷水进行对毛油降温,产生大量能耗。因此,开发新的具有耐热性能的磷脂酶c活性的多肽对于本领域具有重要实用和经济价值。
发明概述
第一方面,本申请提供了具有磷脂酶c活性的多肽,所述多肽具有1)包含在一个或多个位置发生氨基酸取代的seqidno:2所示的氨基酸序列,其中所述一个或多个位置选自seqidno:2所示的氨基酸序列中第6位、第8位、第10位、第104位、第205位或其任意组合;或
2)与1)至少80%,优选至少85%,更优选至少90%,更优选至少95%,更优选至少97%,更优选至少98%,更优选至少99%的序列同一性,且第6位、第8位、第10位、第104位、第205位的至少1个位置与seqidno:2所示的氨基酸序列中第6位、第8位、第10位、第104位、第205位不同。
在一些实施方案中,seqidno:2所示的氨基酸序列中第6位、第8位、第10位、第104位和第205位全部发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位的氨基酸取代为脯氨酸、甘氨酸、羟脯氨酸、丝氨酸或苏氨酸取代赖氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位的赖氨酸被脯氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位的氨基酸取代为异亮氨酸、亮氨酸、缬氨酸、甲硫氨酸、丙氨酸、苯丙氨酸或正亮氨酸取代赖氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位的赖氨酸被异亮氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位的氨基酸取代为苏氨酸或丝氨酸取代甘氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位的甘氨酸被苏氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第104位的氨基酸取代为甘氨酸或脯氨酸取代赖氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第104位的赖氨酸被甘氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第205位的氨基酸取代为酪氨酸、色氨酸、苯丙氨酸、或苏氨酸取代丝氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第205位的丝氨酸被酪氨酸取代。
在一些实施方案中,所述多肽的氨基酸序列包含如seqidno:4所示的氨基酸序列。
在一些实施方案中,所述多肽的氨基酸序列由如seqidno:4所示的氨基酸序列组成。
第二方面,本申请提供了一种分离的多肽,其中所述多肽与seqidno:4所示的氨基酸序列具有80%,优选至少85%,更优选至少90%,更优选至少95%,更优选至少97%,更优选至少98%,更优选至少99%的序列同一性,并且所述分离的多肽在对应于seqidno:4的第6位、第8位、第10位、第104位和第205位上的氨基酸残基分别是脯氨酸、异亮氨酸、苏氨酸、甘氨酸和酪氨酸。优选的,所述多肽获得自枯草芽孢杆菌(bacillussubtilis)。
第三方面,本申请提供了编码第一方面或第二方面所述的多肽的核酸分子。
在一些实施方案中,所述核酸分子包含如seqidno:3所示的核酸序列。
在一些实施方案中,所述核酸分子的序列如seqidno:3所示。
第四方面,本申请提供了包含第三方面所述的核酸分子的载体。
在一些实施方案中,所述载体为表达载体。
在一些实施方案中,所述载体被设计用于真核细胞或原核细胞中表达。
在一些实施方案中,所述载体被设计用于细菌细胞、真菌细胞、酵母细胞、哺乳动物细胞、昆虫细胞或植物细胞中表达。
第五方面,本申请提供了包含第三方面所述的核酸分子或第四方面所述的载体的细胞。
在一些实施方案中,所述细胞为真核细胞或原核细胞。
在一些实施方案中,所述细胞为细菌细胞、真菌细胞、酵母细胞、哺乳动物细胞、昆虫细胞或植物细胞。
第六方面,本申请提供了使用第五方面所述的细胞产生的磷脂酶c。
第七方面,本申请提供了如第一方面或第二方面所述的多肽、或如第三方面所述的核酸分子编码的多肽、或如第四方面所述的载体编码的多肽、或如第五方面所述的细胞表达的发酵液、浓缩液或多肽、或如第六方面所述的磷脂酶c作为磷脂酶c的用途。
在一些实施方案中,所述用途为在油脂脱胶工艺中的用途。
第八方面,本申请提供了一种酶组合物,其包括如第一方面或第二方面所述的多肽、或如第三方面所述的核酸分子编码的多肽、或如第四方面所述的载体编码的多肽、或如第五方面所述的细胞表达的多肽、或如第六方面所述的磷脂酶c,以及至少一种脱胶酶。
在一些实施方案中,至少一种脱胶酶选自:磷脂酶a1、磷脂酶a2、磷脂酶b、磷脂酶d、果胶酶和甘露聚糖酶。
第九方面,本申请提供了如第一方面或第二方面所述的多肽、或如第三方面所述的核酸分子、或如第四方面所述的载体、或如第五方面所述的细胞、或如第八方面所述的酶组合物在制备脱胶酶中的用途。
附图说明
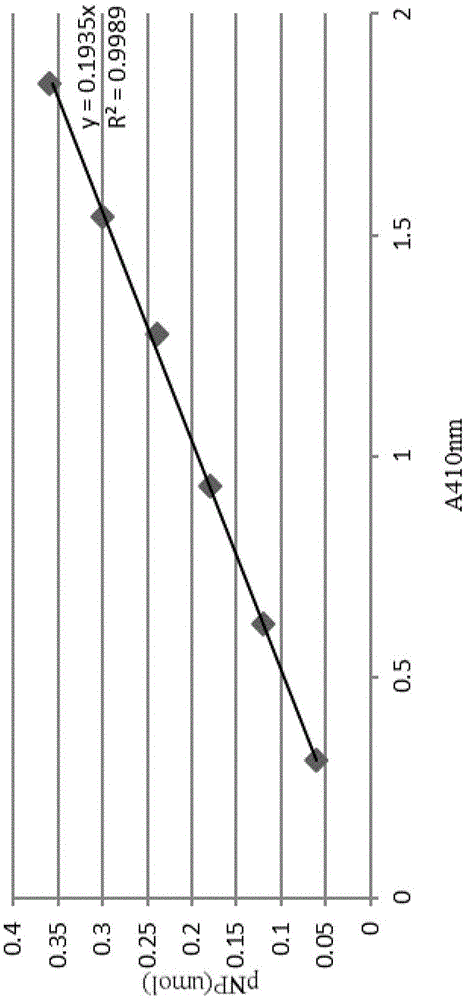
图1为磷脂酶活力测定标准曲线。
图2为plc-9-49和plc-n63dn131sn134d-y56h的热稳定性比较,其中圆圈表示plc-9-49,三角表示plc-n63dn131sn134d-y56h。
图3为plc-9-49和plc-n63dn131sn134d-y56h分别在55℃和60℃的条件下进行脱胶,相对于毛油的dag增量的比较。
序列说明
seqidno:1为plc-n63dn131sn134d-y56h的编码核酸序列。
seqidno:2为plc-n63dn131sn134d-y56h的氨基酸序列。
seqidno:3为plc-9-49的编码核酸序列。
seqidno:4为plc-9-49的氨基酸序列。
发明详细描述
定义
本文中所述的磷脂酰胆碱特异性磷脂酶c(specificphosphatidylcholinephospholipasec)与磷脂酰胆碱偏好型磷脂酶c(phosphatidylcholine-preferringphospholipasec)为同义术语,并且也是本领域技术人员能够容易理解的。本文中使用简写pc-plc来表示磷脂酰胆碱特异性磷脂酶c或磷脂酰胆碱偏好型磷脂酶c。
本文中使用的磷脂酰胆碱特异性磷脂酶c的一个实例为蜡样芽胞杆菌(bacilluscereus)的磷脂酰胆碱特异性磷脂酶c,在本文中以简写bc-pc-plc表示。应当理解,在本文中,bc-pc-plc可表示蜡样芽胞杆菌的野生型磷脂酰胆碱特异性磷脂酶c,可也表示本申请中基于该野生型磷脂酰胆碱特异性磷脂酶c获得的突变体。
在本文中涉及用数字表示氨基酸位置的情况中,所述数字是参照seqidno:2中的氨基酸位置,seqidno:2为蜡样芽胞杆菌的磷脂酰胆碱特异性磷脂酶c突变体plc-n63dn131sn134d-y56h的氨基酸序列。
本文中使用了国际通用的氨基酸的单字母或三字母缩写。
本文所用的术语“多肽”、“肽”和“蛋白”可互换使用,表示多个氨基酸通过肽键连接形成的聚合物。氨基酸可以是天然存在的或人工合成的类似物。
本文所用的术语“核酸”和“多核苷酸”可互换使用,包括但不限于dna、rna等。核苷酸可以是天然存在的或人工合成的类似物。
本文中的细胞可以是真核细胞或原核细胞,例如,但不限于细菌细胞、真菌细胞、酵母细胞、哺乳动物细胞、昆虫细胞或植物细胞。
本文所用的术语“保守型取代”指蛋白的氨基酸组成的变化,所述变化不会显著改变蛋白的活性。因此,具体氨基酸序列的“保守型取代”指对蛋白活性并非关键的那些氨基酸的取代,或用具有相似性质(例如酸性、碱性、带正电荷或带负电荷、极性或非极性等)的其它氨基酸取代氨基酸,使得即便是关键氨基酸的取代也不会显著改变活性。
本文所用的术语“随机饱和突变”指将需要突变的位点在pcr引物中以设计为nnk兼并密码子,从而能够覆盖所有20种氨基酸达到饱和突变的效果,又由于选择了多个位点进行突变,其组合方式具有随机性,因此称之为随机饱和突变。
提供功能上相似的氨基酸的保守型取代表在本领域中是熟知的。例如,以下列出了各个氨基酸的可选的保守氨基酸取代:
丙氨酸(ala)的保守取代为缬氨酸(val)*、亮氨酸(leu)、异亮氨酸(ile);
精氨酸(arg)的保守取代为赖氨酸(lys)*、谷氨酰胺(gln)、天冬酰胺(asn);
天冬酰胺(asn)的保守取代为谷氨酰胺(gln)*、组氨酸(his)、赖氨酸(lys)、精氨酸(arg)、天冬氨酸(asp);
天冬氨酸(asp)的保守取代为谷氨酸(g1u)*、天冬酰胺(asn);
半胱氨酸(cys)的保守取代为丝氨酸(ser);
谷氨酰胺(gln)的保守取代为天冬酰胺(asn)*、组氨酸(his)、赖氨酸(lys);
谷氨酸(g1u)的保守取代为天冬氨酸(asp)*、γ-羟基谷氨酸(gla);
甘氨酸(gly)的保守取代为脯氨酸(pro);
组氨酸(his)的保守取代为天冬酰胺(asn)、谷氨酰胺(gln)、赖氨酸(lys)、精氨酸(arg)*;
异亮氨酸(ile)的保守取代为亮氨酸(leu)*、缬氨酸(val)、甲硫氨酸(met)、丙氨酸(ala)、苯丙氨酸(phe)、正亮氨酸(nle);
亮氨酸(leu)的保守取代为正亮氨酸(nle)、异亮氨酸(ile)*、缬氨酸(val)、甲硫氨酸(met)、丙氨酸(ala)、苯丙氨酸(phe);
赖氨酸(lys)的保守取代为精氨酸(arg)*、谷氨酰胺(gln)、天冬酰胺(asn)、鸟氨酸(orn);
甲硫氨酸(met)的保守取代为亮氨酸(leu)*、异亮氨酸(ile)、苯丙氨酸(phe)、正亮氨酸(nle);
苯丙氨酸(phe)的保守取代为亮氨酸(leu)*、缬氨酸(val)、异亮氨酸(ile)、丙氨酸(ala);
脯氨酸(pro)的保守取代为甘氨酸(gly)*、羟脯氨酸(hyp)、丝氨酸(ser)、苏氨酸(thr);
丝氨酸(ser)的保守取代为苏氨酸(thr);
苏氨酸(thr)的保守取代为丝氨酸(ser);
色氨酸(trp)的保守取代为酪氨酸(tyr);
酪氨酸(tyr)的保守取代为色氨酸(trp)、苯丙氨酸(phe)*、苏氨酸(thr)、丝氨酸(ser);
缬氨酸(val)的保守取代为异亮氨酸(ile)、亮氨酸(leu)*、甲硫氨酸(met)、苯丙氨酸(phe)、丙氨酸(ala)、正亮氨酸(nle)。
其中,*表示优选的保守取代。
此外,还可参阅creighton,proteins:structuresandmolecularproperties(蛋白:结构和分子特性),w.h.freemanandcompany,newyork(2nded.,1992)。
技术方案详细描述
在本申请发明人先前的专利申请cn201510946696.1中,将bc-pc-plc的四个氨基酸(即第56位、第63位、第131位和第134位的氨基酸)分别突变为组氨酸(y56h)、天冬氨酸(n63d)、丝氨酸(n131s)和天冬氨酸(n134d),突变后的氨基酸序列如seqidno:2所示,编码序列如seqidno:1所示,本文也称为“plc-n63dn131sn134d-y56h”。plc-n63dn131sn134d-y56h的比酶活相比于野生型bc-pc-plc有提高,但此突变体的稳定性不高,不能耐受超过60℃的温度,最合适的脱胶温度为50℃,这使得此突变体在工业应用中具有一定的局限性。首先,在脱胶时,一般会提高温度,这样有利于降低油的粘度,提高油脂和磷脂的分离程度,减少磷脂中夹带的油脂,而进一步提高油脂的得率。其次,目前的技术中可能联合使用plc与pla1以进行深度脱胶。例如,在中国专利申请第201480017114.5号中涉及一种新的具有磷脂酶a活性的多肽,其最适的脱胶温度约65℃(参见专利申请cn105073985a的第[0173]段),而与这样的pla1进行联合脱胶最好希望plc也能配合这样的脱胶温度。此外,有时毛油在储藏过程中尤其是在高温天气,其温度会超过50℃,因此,如果plc的最适反应温度低于50℃,则还需要冷水对毛油进行降温,这将产生能耗。
因此,为了提高磷脂酶c的热稳定性和/或提高磷脂酶c的适用性,本申请的发明人对plc-n63dn131sn134d-y56h进行了改进,并建立了本申请的各项发明。
第一方面,本申请提供了具有磷脂酶c活性的多肽,所述多肽具有1)包含在一个或多个位置发生氨基酸取代的seqidno:2所示的氨基酸序列,其中所述一个或多个位置选自seqidno:2所示的氨基酸序列中第6位、第8位、第10位、第104位、第205位或其任意组合;2)与1)至少80%,优选至少85%,更优选至少90%,更优选至少95%,更优选至少97%,更优选至少98%,更优选至少99%的序列同一性,且第6位、第8位、第10位、第104位、第205位的至少1个位置与seqidno:2所示的氨基酸序列中第6位、第8位、第10位、第104位、第205位不同。
在一些实施方案中,seqidno:2所示的氨基酸序列中第6位、第8位、第10位、第104位和第205位全部发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位和第8位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位和第10位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位和第10位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第8位和第10位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第8位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第8位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第10位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第10位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位、第10位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位、第10位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位、第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位、第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第8位、第10位和第104位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第8位、第10位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第8位、第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位、第10位、第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位、第10位、第104位和第205位的氨基酸发生氨基酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位的氨基酸取代为脯氨酸、甘氨酸、羟脯氨酸、丝氨酸或苏氨酸取代赖氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第6位的赖氨酸被脯氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位的氨基酸取代为异亮氨酸、亮氨酸、缬氨酸、甲硫氨酸、丙氨酸、苯丙氨酸或正亮氨酸取代赖氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第8位的赖氨酸被异亮氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位的氨基酸取代为苏氨酸或丝氨酸取代甘氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第10位的甘氨酸被苏氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第104位的氨基酸取代为甘氨酸或脯氨酸取代赖氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第104位的赖氨酸被甘氨酸取代。
在一些实施方案中,seqidno:2所示的氨基酸序列的第205位的氨基酸取代为酪氨酸、色氨酸、苯丙氨酸、或苏氨酸取代丝氨酸。
在一些实施方案中,seqidno:2所示的氨基酸序列的第205位的丝氨酸被酪氨酸取代。
在一些实施方案中,所述多肽的氨基酸序列包含如seqidno:4所示的氨基酸序列。
在一些实施方案中,所述多肽的氨基酸序列由如seqidno:4所示的氨基酸序列组成。
本申请还考虑了第一方面所述的多肽的功能性变体。在一些实施方案中,所述功能性变体为保守型取代变体。
第二方面,本申请提供了一种分离的多肽,其中所述多肽与seqidno:4所示的氨基酸序列具有至少80%,优选至少85%,更优选至少90%,更优选至少95%,更优选至少97%,更优选至少98%,更优选至少99%的序列同一性,并且所述分离的多肽在对应于seqidno:4的第6位、第8位、第10位、第104位和第205位上的氨基酸残基分别是脯氨酸、异亮氨酸、苏氨酸、甘氨酸和酪氨酸。
在一些实施方案中,所述多肽获得自枯草芽孢杆菌(bacillussubtilis)。
在一些实施方案中,所述多肽与seqidno:4所示的氨基酸序列具有至少97%、至少98%、至少99%、至少99.5%或更高的序列同一性。
在一些实施方案中,所述多肽与seqidno:4所示的氨基酸序列具有至少97.2%、至少97.6%、至少98%、至少98.4%、至少98.8%、至少99.2%、至少99.6%或更高的序列同一性。
在一些实施方案中,所述分离的多肽在对应于seqidno:4的第6位上的氨基酸残基是脯氨酸。
在一些实施方案中,所述分离的多肽在对应于seqidno:4的第8位上的氨基酸残基是异亮氨酸。
在一些实施方案中,所述分离的多肽在对应于seqidno:4的第10位上的氨基酸残基是苏氨酸。
在一些实施方案中,所述分离的多肽在对应于seqidno:4的第104位上的氨基酸残基是甘氨酸。
在一些实施方案中,所述分离的多肽在对应于seqidno:4的第205位上的氨基酸残基是酪氨酸。
在一些实施方案中,所述分离的多肽在对应于seqidno:4的第6位上的氨基酸残基是脯氨酸,和/或所述分离的多肽在对应于seqidno:4的第8位上的氨基酸残基是异亮氨酸,和/或所述分离的多肽在对应于seqidno:4的第10位上的氨基酸残基是苏氨酸,和/或所述分离的多肽在对应于seqidno:4的第104位上的氨基酸残基是甘氨酸,和/或所述分离的多肽在对应于seqidno:4的第205位上的氨基酸残基是酪氨酸。
第三方面,本申请提供了编码第一方面或第二方面所述的多肽的核酸分子。本申请考虑了由于遗传密码子的简并性或不同物种对于密码子的偏好性所能获得的不同核酸分子。
在一些实施方案中,所述核酸分子包含如seqidno:3所示的核酸序列。
在一些实施方案中,所述核酸分子的序列如seqidno:3所示。
第四方面,本申请提供了包含第三方面所述的核酸分子的载体。
在一些实施方案中,所述载体为表达载体。
在一些实施方案中,所述载体被设计用于真核细胞或原核细胞中表达。
在一些实施方案中,所述载体被设计用于细菌细胞、真菌细胞、酵母细胞、哺乳动物细胞、昆虫细胞或植物细胞中表达。
在一些实施方案中,所述载体为质粒。
合适的真核细胞或原核细胞载体是本领域技术人员公知的,并且多种母体载体都是可以商业购买的。载体的实例包括但不限于本申请的实施例中使用的多种载体。
第五方面,本申请提供了包含第三方面所述的核酸分子或第四方面所述的载体的细胞。
在一些实施方案中,所述细胞为真核细胞或原核细胞。
在一些实施方案中,所述细胞为细菌细胞、真菌细胞、酵母细胞、哺乳动物细胞、昆虫细胞或植物细胞。
在一些实施方案中,所述细胞为毕赤酵母(pichiapastoris)细胞。
在一些实施方案中,所述细胞为枯草芽孢杆菌(bacillussubtilis)细胞。
在一些实施方案中,所述细胞为大肠杆菌细胞。
关于包含本申请的核酸分子的细胞,所述核酸分子可位于染色体外(例如位于载体中),也可以被整合到宿主细胞的染色体中。将核酸分子整合到宿主细胞的染色体中以及将载体通过转化或转染引入宿主细胞中的技术均是本领域技术人员所公知的。
第六方面,本申请提供了使用第五方面所述的细胞产生的磷脂酶c。利用基因工程化宿主细胞产生目标多肽或蛋白的技术是本领域技术人员所公知的。
第七方面,本申请提供了如第一方面或第二方面所述的多肽、或如第三方面所述的核酸分子编码的多肽、或如第四方面所述的载体编码的多肽、或如第五方面所述的细胞表达的发酵液、浓缩液或多肽、或如第六方面所述的磷脂酶c,其作为磷脂酶c的用途。
在一些实施方案中,所述用途为在油脂脱胶工艺中的用途。
磷脂酰胆碱特异性磷脂酶c在油脂脱胶工艺中的应用是本领已知的。磷脂酶c能够水解油中的胶质成分磷脂,生成亲水的磷酸部分和亲油的dag,亲水部分被水带走从而去除胶质部分,dag则增加了油的得率。例如,酶法脱胶过程为将原料油加热至60℃,加入磷脂酶c溶液,高速剪切混合后,在反应器中搅拌反应2h,随后离心分离水相及油相。
第八方面,本申请提供了一种酶组合物,其包括如第一方面或第二方面所述的多肽、或如第三方面所述的核酸分子编码的多肽、或如第四方面所述的载体编码的多肽、或如第五方面所述的细胞表达的多肽、或如第六方面所述的磷脂酶c,以及至少一种脱胶酶。
在一些实施方案中,所述至少一种脱胶酶选自:磷脂酶a1、磷脂酶a2、磷脂酶b、磷脂酶d、果胶酶和甘露聚糖酶。
在一些实施方案中,酶组合物包括如第一方面或第二方面所述的多肽以及至少一种脱胶酶。
在一些实施方案中,酶组合物包括如第三方面所述的核酸分子编码的多肽以及至少一种脱胶酶。
在一些实施方案中,酶组合物包括如第四方面所述的载体编码的多肽以及至少一种脱胶酶。
在一些实施方案中,酶组合物包括如第五方面所述的细胞表达的多肽以及至少一种脱胶酶。
在一些实施方案中,酶组合物包括如第六方面所述的磷脂酶c以及至少一种脱胶酶。
在一些实施方案中,酶组合物包括如第一方面或第二方面所述的多肽,和/或如第三方面所述的核酸分子编码的多肽,和/或如第四方面所述的载体编码的多肽,和/或如第五方面所述的细胞表达的多肽,和/或如第六方面所述的磷脂酶c,以及至少一种脱胶酶。
第九方面,本申请提供了如第一方面或第二方面所述的多肽、或如第三方面所述的核酸分子、或如第四方面所述的载体、或如第五方面所述的细胞、或如第八方面所述的酶组合物在制备脱胶酶中的用途。
应当理解,以上详细描述仅为了使本领域技术人员更清楚地了解本申请的内容,而并非意图在任何方面加以限制。本领域技术人员能够对所述实施方案进行各种改动和变化。
实施例
提供以下实施例进一步描述本申请,而并非加以任何限制。
实验材料
本申请的实施例中所用的主要材料如下:
1.菌株
毕赤酵母smd1168(invitrogen,货号c17500),大肠杆菌dh5α(takara,货号d9057a)。
2.培养基和溶液
lb液体培养基:0.5%酵母提取物,1%胰化蛋白胨,1%nacl,ph7.0。
lb固体培养基:在lb液体培养基中加入浓度1.5%的琼脂。
ypd液体培养基:1%酵母提取物,2%蛋白胨,2%葡萄糖。
ypd固体培养基:在lb液体培养基中加入浓度2%的琼脂。
mgys固体培养基:1.34%酵母氮源碱(ynb)(含硫酸铵、不含氨基酸),1%甘油,1m山梨醇,4×10-5%d-生物素,2%琼脂。
bmm-大豆磷脂筛选培养基:1.34%酵母氮源碱(ynb)(含硫酸铵、不含氨基酸),4×10-5%的d-生物素,0.5%甲醇(灭菌后加入),2%大豆磷脂乳化液,0.1m柠檬酸-柠檬酸钠缓冲液(ph=6.6),2%琼脂,并添加10um的znso4·7h2o。
2%大豆磷脂乳化液的制备:称取2g大豆磷脂、100ml的h2o,用高速匀浆机于8000rpm匀浆1min。
bmgy液体培养基:1%酵母提取物,2%蛋白胨,1.34%酵母氮源碱(ynb)(含硫酸铵、不含氨基酸),1%甘油,4×10-5%的d-生物素,0.1m磷酸二氢钾-磷酸氢二钾缓冲液(ph=6.0)。
bmmy液体培养基:1%酵母提取物,2%蛋白胨,1.34%酵母氮源碱(ynb)(含硫酸铵、不含氨基酸),0.3%的znso4·7h2o,0.5%甲醇(灭菌后加入),4×10-5%的d-生物素(灭菌后加入),0.1m的柠檬酸-柠檬酸钠缓冲液(ph=6.6)。
3.酶活测定:pnppc法
3.1绘制磷脂酶活力测定标准曲线
称取0.01391g对硝基苯酚,溶于50ml无菌水中,制备成2mmol/l工作液。参见表1,添加各种试剂并制作标准曲线,获得的标准曲线如图1所示。试验中测定样品酶活的条件与制作标准曲线的条件相一致。
表1:绘制磷脂酶活力测定标准曲线时的试剂添加量。
将以上各种溶液混合后,在37℃处理15分钟,再加入500μl的0.5n的naoh,于410nm测定吸光度。
3.2反应缓冲液配制
0.1m的硼酸-硼酸钠缓冲液(ph=7.6)中含有20mmpnppc。
取600μl上述缓冲液,加入25μl的待测酶液,于37℃反应15min,再加入500μl的0.5nnaoh,终止反应,于410nm测定吸光度。
4.酶活计算
按以下公式计算酶活:
样品酶活(u/ml)=a(410nm吸光度)×0.1935×稀释倍数×10/15
5.蛋白浓度检测试剂:
改良型bradford法蛋白浓度测定试剂盒(购自上海生工生物工程有限公司)。
6.实验所用的酶:
限制性内切酶sali(购自纽英伦生物技术(北京)有限公司);
dna聚合酶:
t4dna连接酶(购自富酶泰斯有限公司)。
实施例1:磷脂酶c突变体文库构建及筛选
按中国发明申请cn201510946696.1中所述的方法制备pmao-plc-n63dn131sn134d-y56h载体。
以pmao-plc-n63dn131sn134d-y56h载体为模板,构建第6位、第8位、第10位、第104位、第205位氨基酸进行随机饱和突变的文库(委托苏州泓迅生物科技股份有限公司进行文库构建)。将质粒文库转化至大肠杆菌dh5α菌株中,将所有获得的大肠杆菌克隆洗至lb液体培养基中(含100μg/ml地氨苄青霉素),于37℃培养4h。抽提质粒,用sali进行线性化,回收约8.5kb的片段。取500ng载体,用电转化法将载体转化至毕赤酵母m314菌株的感受态细胞中。将转化物接种于mgys平板上,于30℃培养3天,得到plc-n63dn131sn134d-y56h的毕赤酵母突变体文库。挑取平板上的单克隆,接种于bmm-大豆磷脂筛选平板上。选取白色沉淀圈大的克隆。获得突变体菌株,将其指定为plc-9-49。
实施例2:磷脂酶c突变体序列分析
将plc-9-49菌株接种于3mlypd液体培养基中,30℃培养过夜,抽提基因组dna。以plc-9-49菌株的基因组dna为模板,使用
引物aox1-5的序列为:5’-gactggttccaattgacaacg-3’;
引物aox1-3的序列为:5’-ggcaaatggcattctgacatcctc-3’。
将获得的序列送往上海生工生物工程公司,用引物对aox1-5/aox1-3进行测序。plc-9-49的dna测序结果如seqidno:3所示。经过比对发现,与seqidno:1相比,seqidno:3中有数个碱基发生了突变,使得seqidno:2中的第6位的赖氨酸突变为脯氨酸、第8位的赖氨酸突变为异亮氨酸、第10位的甘氨酸突变为苏氨酸、第104位的赖氨酸突变为甘氨酸以及第205位的丝氨酸突变为酪氨酸。plc-9-49的氨基酸序列如seqidno:4所示。
实施例3:plc-9-49突变体热稳定性分析
取plc-9-49菌株及plc-n63dn131sn134d-y56h菌株,先在液体ypd中活化,然后接种于bmgy培养基中,于30℃以220rpm的转速振荡培养过夜。将培养物转至bmmy培养基中,初始od600为6。
首先,用2%甲醇进行诱导,在24h和32h后各补加1%甲醇,48h和56h后各补加1%甲醇,于72h取样。将获得的样品用截留分子量为10kda的超滤管进行超滤脱盐浓缩40倍。将处理后的样品加入缓冲液(20mm柠檬酸-柠檬酸钠缓冲液(ph=6.6),含有10um的znso4)中。
将超滤浓缩后的发酵液在60℃、65℃、70℃和75℃条件下保温2h,取0.5μl发酵液浓缩液添加至600μlpnppc反应缓冲液中,于37℃反应15min,再加入500μl0.5n的naoh终止反应,于410nm测定吸光度。根据标准曲线,计算得到各个发酵液样品的磷脂酶c活力。
plc-9-49和plc-n63dn131sn134d-y56h的热稳定性如图2所示,在60℃条件下保温2h,plc-9-49突变株的活力保留了91%,而plc-n63dn131sn134d-y56h的活力下降至44%。在70℃条件下保温2h,plc-9-49突变株的活力保留了83%,而plc-n63dn131sn134d-y56h的活力下降至13%。在75℃条件下保温2h,plc-9-49突变株的活力保留了61%,而plc-n63dn131sn134d-y56h的活力下降至3%。
由此可见,plc-9-49的热稳定性较plc-n63dn131sn134d-y56h有明显的提高。
实施例4:plc-9-49脱胶测试
取大豆毛油100g,分别加热至55℃和60℃进行脱胶,分别加入50ppm的plc-9-49和plc-n63dn131sn134d-y56h样品,使得体系中的水相为3%,用高速剪切机高速(10000r/min)剪切1min,在55℃和60℃搅拌(750r/min)反应2h,升温至85℃,维持5min。将样品以12000rpm离心10min,取约10g上层油样,用hplc检测其dag含量。plc-9-49样品和plc-n63dn131sn134d-y56h样品相对于毛油的dag增量如图3所示。plc-9-49在60℃脱胶时相比于在55℃脱胶时,dag的增量增加了约10%,而plc-n63dn131sn134d-y56h在60℃脱胶时相较于在55℃脱胶时,dag的增量减少了9.4%。
由此可见,plc-9-49的脱胶温度相比于plc-n63dn131sn134d-y56h上升约5℃,因此工业适用性更好。
序列表
<110>丰益(上海)生物技术研发中心有限公司
<120>具有磷脂酶c活性的多肽及其用途
<160>4
<170>siposequencelisting1.0
<210>1
<211>738
<212>dna
<213>人工序列(artificialsequence)
<400>1
tggtcagctgaggacaagcataaggaaggtgtgaatagtcacttatggatcgtgaaccgt60
gccattgatataatgtctaggaatacaactctggttaagcaagatagagttgctcaattg120
aatgaatggcgtacagagctagagaatggcatctacgctgctgatcatgaaaacccctat180
tacgatgacagtaccttcgcttctcacttttacgatccagacaacggaaagacatatatc240
ccattcgccaagcaagctaaggagactggagctaagtacttcaagttggctggagagtca300
tacaagaataaagacatgaagcaggccttcttttatcttgggttgtcattgcattatttg360
ggcgatgtcaaccaacctatgcatgccgcatcctttacggacctgtcctatccacagggt420
tttcactccaagtacgagaactttgtcgatactattaaagacaactacaaagttaccgat480
gggaacggatattggaattggaaaggcaccaaccctgaagaatggattcacggtgcagca540
gtagttgcaaaacaggactactctggaattgtcaatgacaataccaaagattggtttgtg600
aaagccgcagtctcccaggaatatgcagataaatggagagctgaagttacacctatgact660
ggtaaacgactaatggatgcccaaagagttactgctggttacattcaattatggttcgac720
acttacggtgacaggtaa738
<210>2
<211>245
<212>prt
<213>人工序列(artificialsequence)
<400>2
trpseralagluasplyshislysgluglyvalasnserhisleutrp
151015
ilevalasnargalaileaspilemetserargasnthrthrleuval
202530
lysglnaspargvalalaglnleuasnglutrpargthrgluleuglu
354045
asnglyiletyralaalaasphisgluasnprotyrtyraspaspser
505560
thrphealaserhisphetyraspproaspasnglylysthrtyrile
65707580
prophealalysglnalalysgluthrglyalalystyrphelysleu
859095
alaglyglusertyrlysasnlysaspmetlysglnalaphephetyr
100105110
leuglyleuserleuhistyrleuglyaspvalasnglnpromethis
115120125
alaalaserphethraspleusertyrproglnglyphehisserlys
130135140
tyrgluasnphevalaspthrilelysaspasntyrlysvalthrasp
145150155160
glyasnglytyrtrpasntrplysglythrasnprogluglutrpile
165170175
hisglyalaalavalvalalalysglnasptyrserglyilevalasn
180185190
aspasnthrlysasptrpphevallysalaalavalserglnglutyr
195200205
alaasplystrpargalagluvalthrprometthrglylysargleu
210215220
metaspalaglnargvalthralaglytyrileglnleutrppheasp
225230235240
thrtyrglyasparg
245
<210>3
<211>738
<212>dna
<213>人工序列(artificialsequence)
<400>3
tggtcagctgaggaccctcatattgaaactgtgaatagtcacttatggatcgtgaaccgt60
gccattgatataatgtctaggaatacaactctggttaagcaagatagagttgctcaattg120
aatgaatggcgtacagagctagagaatggcatctacgctgctgatcatgaaaacccctat180
tacgatgacagtaccttcgcttctcacttttacgatccagacaacggaaagacatatatc240
ccattcgccaagcaagctaaggagactggagctaagtacttcaagttggctggagagtca300
tacaagaatggggacatgaagcaggccttcttttatcttgggttgtcattgcattatttg360
ggcgatgtcaaccaacctatgcatgccgcatcctttacggacctgtcctatccacagggt420
tttcactccaagtacgagaactttgtcgatactattaaagacaactacaaagttaccgat480
gggaacggatattggaattggaaaggcaccaaccctgaagaatggattcacggtgcagca540
gtagttgcaaaacaggactactctggaattgtcaatgacaataccaaagattggtttgtg600
aaagccgcagtctatcaggaatatgcagataaatggagagctgaagttacacctatgact660
ggtaaacgactaatggatgcccaaagagttactgctggttacattcaattatggttcgac720
acttacggtgacaggtaa738
<210>4
<211>245
<212>prt
<213>人工序列(artificialsequence)
<400>4
trpseralagluaspprohisilegluthrvalasnserhisleutrp
151015
ilevalasnargalaileaspilemetserargasnthrthrleuval
202530
lysglnaspargvalalaglnleuasnglutrpargthrgluleuglu
354045
asnglyiletyralaalaasptrpgluasnprotyrtyraspaspser
505560
thrphealaserhisphetyraspproaspasnglylysthrtyrile
65707580
prophealalysglnalalysgluthrglyalalystyrphelysleu
859095
alaglyglusertyrlysasnglyaspmetlysglnalaphephetyr
100105110
leuglyleuserleuhistyrleuglyaspvalasnglnpromethis
115120125
alaalaserphethraspleusertyrproglnglyphehisserlys
130135140
tyrgluasnphevalaspthrilelysaspasntyrlysvalthrasp
145150155160
glyasnglytyrtrpasntrplysglythrasnprogluglutrpile
165170175
hisglyalaalavalvalalalysglnasptyrserglyilevalasn
180185190
aspasnthrlysasptrpphevallysalaalavaltyrglnglutyr
195200205
alaasplystrpargalagluvalthrprometthrglylysargleu
210215220
metaspalaglnargvalthralaglytyrileglnleutrppheasp
225230235240
thrtyrglyasparg
245
- 还没有人留言评论。精彩留言会获得点赞!