用于口服蛋白质递送的工具和方法与流程
发明领域
本发明涉及在宿主细胞如酵母细胞中产生重组蛋白的领域。更具体地,本发明涉及口服蛋白质递送领域。具体地,本发明提供了包含分泌重组多肽的重组宿主的培养基的口服药物制剂。
背景技术:
肽或蛋白质(包括激素,酶,配体或抑制剂),包括抗体,调节各种细胞功能。因此,它们在临床上可通过调节生理或病理过程来治疗或预防人类疾病。与小分子药物相比,肽或蛋白质对其靶标的高选择性可减少副作用和对宿主细胞的毒性。预期在癌症,代谢紊乱,胃肠疾病,口腔疾病,神经变性和感染性疾病的治疗中,用于治疗目的的蛋白质或肽的使用将持续增加。当前,蛋白质药物主要使用哺乳动物,植物,酵母或细菌细胞培养系统制造。这些表达的蛋白质必须被提取和纯化,这需要昂贵且复杂的工艺以及冷冻储藏和运输。生物制剂通常通过静脉内或皮下注射来递送,这是有效的,但对于患者,特别是对于慢性病患者而言并不理想。蛋白质药物的可注射形式常需要医护人员进行施用,从而导致频繁的医院就诊和降低的患者依从性。正在研究其他递送途径,例如透皮,鼻内,吸入和口服给药,但是通常认为口服递送是最期望的途径。尽管进行了数十年的努力,但是肽、蛋白质和抗体药物的口服递送仍然是主要的药学挑战,市场上只有极少数此类蛋白质。这尤其令人失望,因为生物制剂是制药市场的快速增长部分,在过去十年中其价值从360亿美元增至1630亿美元,增长了三倍。因此,尽管对患者方便,但是存在许多技术障碍,使得这种给药途径对大分子药物具有挑战性。当然,最重要的挑战是胃和肠中药物的酶促降解和ph依赖性降解。另外,沿胃肠道(gi)内衬排列的上皮细胞渗透性低,并且这些化合物存在内在不稳定性。已经开发了几种技术来促进大分子的口服递送。附着分子,例如聚乙二醇、抗体fc域或人血清白蛋白,可增加循环过程中肽在血清中的稳定性。另外,可以修饰肽药物以保护其免受血清蛋白酶和肽酶的侵害。这样的修饰包括n端乙酰化,c端酰胺化,使用非天然氨基酸和通过二硫键的环化。另外,在酶抑制剂的开发、吸收或渗透促进剂的使用、多肽在胶囊中的配制、粘在肠壁上的粘性聚合物的应用、以及载体分子在制剂中的掺入方面,已经取得了一些改进。尽管有这些技术,但是蛋白质和肽经口摄入时通常具有极低的生物利用度,低于2%。最近,我们证明,植物种子产生的抗体在胃小凹(gastriccanal)中存活并且在肠中具有生物活性(参见wo2014033313和virdiv.等人(2013)pnas,110,29,11809-11814)。植物生产系统尽管能够生产大量重组蛋白,但由于冗长而昂贵的监管程序,它们并不是生产可食用疫苗的最佳选择。低等真核生物是更期望的,其能够产生更高量的重组蛋白。使用低等真核宿主如酵母来产生可口服递送的治疗性蛋白质将是有利的。已经描述了低等真核细胞用于递送治疗性蛋白质,但是仅以能够产生治疗性蛋白质的完整重组细胞的形式来递送(参见例如zhang等人(2012)bmcbiotechnology12:97和wo2007039586)。期望能够仅使用包含分泌的多肽的培养基而不是重组酵母本身。甚至更期望的是,重要的是,不必从培养基中纯化治疗性多肽,而照原样使用该培养基。
发明概述
在本发明中,我们令人惊讶地证实,包含重组酵母培养基的干燥制剂可以用作口服递送制剂,所述培养基包含治疗性多肽,并且所述培养基已经通过膜分离工艺处理以降低渗透化合物的浓度。已证实,令人惊讶地,包含治疗性多肽的粉末状干燥培养基(通过冻干或通过喷雾干燥获得)可以免遭胃肠道(gut)降解。此外,还证实,粉末状的干燥制剂也可以在胃肠道中保持其生物活性。
在第一方面,本发明提供了一种干燥制剂,其包含口服容许的基质和来自重组宿主的培养基的混合物,所述培养基中可溶性渗透化合物的浓度已经在膜分离工艺中被降低,所述宿主产生分泌到所述培养基中的外源多肽。
在又一个具体方面,本发明提供了一种制剂,其包含来自重组酵母的培养基,所述酵母产生分泌到所述培养基中的外源多肽。
在第二方面,本发明提供了根据方面1的制剂,其中在制备所述制剂之前,通过在膜分离工艺中浓缩所述培养基来减少所述制剂的水含量和分子量低于5kda的可溶性分子。
在第三方面,本发明提供了根据方面1的制剂,其中通过在制备所述制剂之前干燥所述培养基来减少水含量和分子量低于5kda的可溶性分子。
在第四方面,本发明提供根据方面1的制剂,其中通过干燥所述制剂,降低所述制剂的水含量和分子量低于5kda的可溶性分子。
在第五方面,本发明提供了一种干燥制剂,其包含重组宿主的培养基,所述宿主产生分泌到所述培养基中的外源多肽。
在第六方面,本发明提供了根据第五方面的干燥制剂,其中所述重组宿主是酵母。
在第七方面,本发明提供根据方面5和6的干燥制剂,其中所述外源多肽与fc结构域融合。
在第八方面,本发明提供了根据方面7的干燥制剂,其中所述fc结构域是igafc结构域。
在第九方面,本发明提供根据方面5至8中任一项的干燥制剂,其中所述外源肽是预防性或治疗性肽,或者其中所述外源肽是疫苗或形成疫苗的一部分。
在第十方面,本发明提供了干燥的制剂,其包含对于重组宿主而言是外源的多肽,所述制剂是通过将所述多肽分泌到重组宿主的培养基中,然后干燥所述培养基而获得的。
在第十一方面,本发明提供了一种干燥制剂,其包含具有fc结构域的融合多肽,其通过将所述融合多肽产生到重组宿主的培养基中,然后干燥所述培养基而获得。
在第十二方面,本发明提供根据方面10或11的干燥制剂,其中所述重组宿主是原核宿主、植物细胞或真菌细胞,特别是丝状真菌。
在第十三方面,本发明提供了根据方面10或11的干燥制剂,其中所述重组宿主是酵母细胞。
在第十四方面,本发明提供根据方面5至13中任一项的干燥制剂,其中所述干燥通过喷雾干燥进行。
在第十五方面,本发明提供根据方面5至13中任一项的干燥制剂,其中所述干燥通过冻干进行。
在第十六方面,本发明提供根据方面5至13中任一项的干燥制剂,其进一步包含酵母细胞。
在第十七方面,本发明提供根据方面5至13中任一项的干燥制剂,其进一步包含富含蛋白质的粗粉(meal)。
在第十八方面,本发明提供了根据方面5至17中任一项的干燥制剂,其用作药物。
在第十九方面,本发明提供了根据方面5至17中任一项的干燥制剂,其用于治疗胃肠道疾病。
在第二十方面,本发明提供了根据方面5至17中任一项的干燥制剂,其用于治疗口腔疾病。
在第二十一方面,本发明提供了根据方面5至17中任一项的干燥制剂用作预防性产品。
在第二十二方面,本发明提供了根据方面5至17中任一项的干燥制剂,其用作疫苗。
在第二十三方面,本发明提供了根据方面5至17中任一项的干燥制剂,其用作功能性食品。
在第二十三方面,本发明提供了根据方面5至17中任一项的干燥制剂,其用作药用食品。
在第二十四方面,本发明提供了一种食品,其包含根据方面5至17中任一项的干燥制剂。
在第二十五方面,本发明提供了根据方面5至17中任一项的干燥制剂,其中所述多肽是il22igafc融合体。
在第二十七方面,本发明提供了一种口服药物制剂,其包含根据方面5至17中任一项的干燥制剂和药物赋形剂。
在第二十八方面,本发明提供了根据方面5至17中任一项的干燥制剂用作药物。
在第二十九方面,本发明提供了根据方面27的口服药物制剂用作药物。
附图说明
图1:通过固定化faeg的elisa,筛选和选择高表达性毕赤酵母克隆(a)和大豆种子(b)。图“c”和“d”分别显示了含有vhh-igafc的毕赤酵母上清液和大豆vhh-igafc表达种子提取物的代表性免疫印迹。
图2:基于elisa滴定的一个典型实例,用于比较毕赤酵母和种子产生的抗体的功能等效性。在该代表性图中,将虚线表示的毕赤酵母产生的v2a抗体(pichia-v2a)的曲线与实线表示的不同浓度的拟南芥产生的v2a(at-v2a)进行了比较。
图3:毕赤酵母和大豆产生的vhh-iga可以防止仔猪etec感染。实验示意图(a);攻击后f4-etec的排出(b);血清转化显示抗f4-etec血清igm(c),igg(d)和iga(e)滴度。
图4:图a.实施例3的实验设置;图b.4个不同组的每克粪便中f4+etec细菌的排出,直到第6天;图c.属于4个不同组的仔猪个体的抗etecigg的血清滴度;图d.属于4个不同组的仔猪个体的抗eteciga的血清滴度。
图5:在komagataellaphaffi(原名巴斯德毕赤酵母)中产生的gap启动子驱动的v2igafc-融合体表达的还原sds-page分析。图a)c-末端his标记的v2igafc经ni-imac纯化,1μg重组蛋白用pngasef处理(右泳道)或未处理(左泳道);图b)产v2igafc的komagataellaphaffi的粗上清液用pngasef处理(右泳道)或未处理(左泳道)。用pngasef处理后,分子量强烈降低至约38kda,这对应于非糖基化分子的理论分子量。。存在于单个n-糖基化位点上的高甘露糖基(hypermannosyl)结构使蛋白质的分子量增加30kda以上。
图6:当在komagataellaphaffi中由组成型gap启动子(在葡萄糖上培养)而不是由甲醇诱导性aoxi启动子(在甲醇上培养)产生v2igafc分子时,糖基化水平增加。gap启动子产生的蛋白质的平均mw约为70kda,而非糖基化的v2igafc蛋白质的预期mw为38.4kda。
发明详述
将根据具体实施方案并参考某些附图来描述本发明,但是本发明不限于此,而是仅由权利要求书来限定。权利要求中的任何附图标记不应被解释为限制范围。当然,应当理解,根据本发明的任何具体实施方案,并无必要可以实现所有方面或优点。因此,例如,本领域技术人员将认识到,本发明可以以实现或优化本文所教导的一个优点或一组优点的方式实施或实现,而不必实现本文所教导或建议的其他方面或优点。
当结合附图阅读下面的详细描述时,可以最好地理解本发明的组织和操作方法以及其特征和优点。参考下文描述的实施方案,本发明的各方面和优点将变得显而易见并得到阐明。在整个说明书中,提到“一个实施方案”或“实施方案”,是指结合该实施方案描述的特定特征、结构或特性将包括在本发明的至少一个实施方案中。因此,在整个说明书中各处出现的短语“在一个实施方案中”或“在实施方案中”无需,但可以,均指相同实施方案。类似地,应当理解,在本发明的示例性实施方案的描述中,有时将本发明的多种特征组合在单个实施方案、附图或其描述中,以简化公开并帮助理解多个发明方面中的一个或多个。然而,该公开方法不应解释为反映了这样一种意图,即所要求保护的发明需要比每个权利要求中明确叙述的特征更多的特征。相反,如所附权利要求所反映的,发明方面具有的特征可以少于单个之前公开的实施方案的所有特征。
在提及单数名词时使用不定冠词或定冠词的地方,例如“一”或“一个”,“该”,除非特别说明,否则包括该名词的复数形式。在本说明书和权利要求书中使用术语“包括”时,其不排除其他元素或步骤。此外,说明书和权利要求书中的术语第一,第二,第三等用于区分相似的元件,而非一定用于描述顺序或时间顺序。应当理解,如此使用的术语在适当的情况下是可互换的,并且本文描述的本发明的实施方案能够以不同于本文描述或示出的其他顺序来操作。提供以下术语或定义仅用于帮助理解本发明。除非本文特定限定,否则本文使用的所有术语具有与本发明领域的技术人员所理解的相同的含义。本领域技术人员尤其可以参考sambrook等人,《分子克隆:实验室手册》,第四版,冷泉港出版社,纽约州普莱恩斯维尤(2012年);和ausubel等人,《现代分子生物学实验方案》(增刊114),约翰·威利父子出版社,纽约(2016年),以了解本领域的定义和术语。本文所提供的定义不应被解释为具有小于本领域普通技术人员所理解的范围。
当涉及诸如量、持续时间之类的可测量值时,本文所用的“约”意在包括距规定值±20%或±10%,更优选±5%,甚至更优选±1%,且还更优选±0.1%的变化,只要这样的变化适合于执行所公开的方法。
如本文所用,“核苷酸序列”,“dna序列”,“dna元件”或“核酸分子”是指,核苷酸(即核糖核苷酸或脱氧核糖核苷酸)的任何长度的聚合形式。该术语仅指分子的一级结构。因此,该术语包括双链和单链dna以及rna。它还可以包括已知类型的修饰,例如甲基化、“帽子”、一个或多个天然存在的核苷酸用类似物的取代。“编码序列”是核苷酸序列,当置于适当调节序列的控制下时,其可以转录成mrna和/或翻译成多肽。编码序列的边界由5′-末端的翻译起始密码子和3′-末端的翻译终止密码子确定。“编码序列”可以包括但不限于mrna,cdna,重组核苷酸序列或基因组dna,而内含子在某些情况下也可以存在。
“直向同源物”是来自不同生物的基因,这些基因通过物种形成而起源,并且也源自共同的祖先基因。
术语“调控元件”,“控制序列”和“启动子”或“基因的启动子区域”在本文中全部可互换使用,并且应在广义上理解为指作为功能性dna序列单位的调节核酸序列,能够影响与其连接的序列的表达。术语“启动子”通常是指位于基因转录起点上游或与编码序列可操作连接的核酸控制序列,并且可以在置于适当的诱导条件下时,通过其序列的识别和rna聚合酶及其他蛋白质的结合,足以实现所述编码序列的转录。前述术语涵盖:源自经典真核基因组基因的转录调控序列(包括准确的转录起始所需的tata盒,带有或不带有ccaat盒序列)、和可以响应发育和/或外部刺激或以组织特异性方式改变基因表达的其他调控元件(即上游激活序列,增强子和沉默子)。该术语中还包括经典原核基因的转录调控序列,在这种情况下,可以包括-35框序列和/或-10框转录调控序列。术语“调控元件”还涵盖赋予、激活或增强核酸分子在细胞、组织或器官中表达的合成融合分子或衍生物。
术语“蛋白质”,“多肽”和“肽”在本文中可互换使用,进一步是指氨基酸残基的聚合物以及其变体和合成类似物。因此,这些术语适用于氨基酸聚合物,其中一个或多个氨基酸残基是合成的非天然存在的氨基酸,例如相应天然存在的氨基酸的化学类似物,以及天然存在的氨基酸聚合物。该术语还包括多肽的翻译后修饰,例如糖基化,磷酸化和乙酰化。“重组多肽”是指使用重组技术,即通过表达重组或合成多核苷酸制备的多肽。术语“表达”或“基因表达”是指一个或多个特定基因或特定基因构建体的转录。术语“表达”或“基因表达”特别是指将一个或多个基因或遗传构建体转录成结构rna(rrna,trna)或mrna,随后将其翻译成蛋白质或不翻译成蛋白质。该过程包括dna的转录和所得mrna产物的加工。
如本文所用,术语“重组宿主细胞”,“工程化细胞”,“表达宿主细胞”,“表达宿主系统”,“表达系统”或简单地“宿主细胞”旨在指,其中已经引入了重组载体和/或嵌合基因构建体的细胞。应当理解,这些术语不仅旨在指特定的对象细胞,而且还指该细胞的后代。由于突变或环境影响,某些修饰可能在后继世代中发生,所以这样的后代实际上可能与亲本细胞不同,但仍包括在本文所用的术语“宿主细胞”的范围内。重组宿主细胞可以是分离的细胞或在培养中生长的细胞系,细胞可以是原核细胞、真核细胞如动物细胞、植物细胞、真菌细胞如丝状真菌,优选细胞是重组酵母细胞。
如本文所用,术语“内源的”是指起源自生物体、组织或细胞内的物质(例如基因)。类似地,本文所用的“外源性”是指起源于生物体、组织或细胞之外的任何物质,但是其存在于该生物体、组织或细胞中(并且典型地可以成为有活性的)。
胃肠道(gi)对多肽是一种不利的环境,因为它在进化上经过了优化以分解养分并使病原体失活。胃中的高酸性ph值导致蛋白质的质子化及其解折叠,从而暴露出更多可被蛋白质降解酶识别的基序。胃中的酶(胃蛋白酶)、小肠中的酶(例如胰凝乳蛋白酶、氨肽酶和羧肽酶)以及由胰腺和胆汁产生的酶,将蛋白质切割成较小的片段和单个单位。因为治疗活性多肽(例如预防性、治疗性疫苗成分)也受这些过程影响,所以幸免于这些降解过程的级分通常是低且可变的,尤其是在存在食物的情况下。另外,为了达到系统区室,多肽药物需要克服旨在防止膳食和细菌抗原进入体内的多种障碍。为了进入上皮细胞层,多肽首先需要扩散通过覆盖肠上皮的粘液层。上皮是另一个重要的屏障,因为密封上皮细胞的紧密连接将细胞旁运输(即细胞之间的通过)限制到小于600da的小分子和离子。另外,跨细胞的通道是由胃腔表达的内吞性受体(例如,维生素b12受体,转铁蛋白受体)介导的,因此必须与相应配体缀合以用于药物递送。进入系统区室的另一个入口是派伊尔斑块(peyer’spatches)的吞噬性m细胞,该m细胞可以采集腔内抗原并可以摄入低微米范围内的特定底物。然而,胃肠(gut)上皮中的m细胞比例小,并且在物种之间差异很大,这使得根据动物数据对人类吸收进行预测变得复杂。鉴于上述的障碍,毫不奇怪的是,只有六种生物大分子被食品和药物管理局(fda)批准用于口服给药:两种局部给药和两种全身给药的肽,一种局部给药的非肽大环化合物,和一种局部给药的蛋白质混合物(moroze.等人(2016)advanceddrugdeliveryreviews101,108-121)。但是,目前正在对蛋白质、肽和核酸的几种口服制剂进行临床评估。通常,这些制剂包含以下赋形剂中的至少一种:肠溶衣和/或蛋白酶抑制剂,以防止药物降解;渗透增强剂,以实现大分子的细胞旁运输。在机械上,可以通过机械破坏紧密连接或质膜,降低粘液粘度和调节紧密连接调节性信号通路来实现吸收增强。临床上正在研发的生物大分子口服递送的其他策略包括:颊递送、利用载体介导的转胞吞作用、和局部递送至gi靶标。绝大多数的目前批准的口服药物和临床候选药物的分子量<1000da。超过此阈值,尽管经过近90年的反复试验,知识水平有了明显的提高,但低生物利用度、个体之间和个体内部的变异性、食物影响、以及提高生物利用度的赋形剂的长期安全性问题,仍然是口服递送的重要挑战。
本发明针对目前治疗性蛋白质口服递送的缺点提供了明确的解决方案。在本发明中,我们令人惊讶地发现,通过干燥在培养基中分泌治疗性蛋白质的重组酵母的培养基,其中所述培养基包含多数个大于5kda的大分子,而获得的干燥制剂,可以用于该干燥制剂的口服递送。令人惊讶地,该制剂不仅免于胃肠道中的蛋白水解和降解,而且干燥制剂中存在的治疗性蛋白还具有令人惊奇的生物学活性。本发明无需限制到特定的机制或作用,但据信至少一种机制是酵母细胞外介质可以在治疗蛋白周围充当保护膜,从而防止(或减慢)肠道中治疗性蛋白的蛋白水解。这不同于治疗性蛋白质在植物种子中表达的情况,其中干燥的种子基质保护治疗性蛋白质免于降解(参见wo2014033313)。另一个可能的机制是,该糖基化的治疗性蛋白质仅由(高)甘露糖糖结构构成(就其在重组酵母宿主中表达而言)。这些大体积的高甘露糖结构也可能保护治疗性肽免于胃肠道(gut)中的蛋白水解降解。图5描述了在巴斯德毕赤氏酵母中产生的v2a-igafc融合体上存在的大体积高甘露糖糖基化,该重组蛋白在本实施例中使用。我们的发现的主要优点是不需要纯化治疗性蛋白质,这意味着,包含原样培养基的制剂或包含存在于培养基中的多个大于5kda的大分子(包括酵母产生的自身蛋白质)和存在于培养基中的治疗性蛋白质的干燥制剂,可以用作口服药物产品。在以下实施方案中列出了这些优点。
在一个具体的实施方案中,本发明提供了包含重组宿主细胞的培养基的干燥制剂,所述宿主细胞产生分泌到所述重组宿主细胞的生长培养基(或培养基)中的外源蛋白质。重组宿主细胞可以是原核宿主(例如,乳球菌(lactococcus)、芽孢杆菌(bacillus)和其他细菌宿主),真核宿主(例如,植物细胞、动物细胞、真菌细胞,尤其是丝状真菌细胞和酵母细胞)。多肽可以是治疗性肽、预防性肽、或可以用于疫苗组合物中的肽。
在另一个具体的实施方案中,本发明提供了包含重组酵母的培养基的干燥制剂,所述酵母产生分泌到培养基中的外源蛋白质。“培养基”是指没有酵母细胞的发酵液(通常来自高密度酵母发酵液)。“培养基”在本领域中也被称为“生长培养基”。在下游加工中存在几种选择,以将酵母细胞与发酵液分离(也称为发酵液澄清技术),例如离心后进行深度过滤,离心后进行助滤剂增强的深度过滤以及微过滤技术。
显然,发酵液是非常复杂的汤或溶液。从根本上说,发酵液是营养的海洋,酵母在其中生长、繁殖并分泌治疗相关多肽。发酵液通常包含发酵营养成分,例如酵母蛋白胨(包括酵母提取物),酵母自溶物和无活性酵母。在b族维生素、核苷酸、矿物质、α-氨基氮含量和其他生物活性化合物方面,这些产品的含量有所不同。
本领域技术人员已知几种膜分离工艺(具有变化的膜孔径),其可用于降低可溶性渗透物组分的浓度并进一步提高保留的化合物(在此为目的重组多肽)的浓度。反渗透或超滤是由压力梯度驱动的膜分离过程,其中膜将溶剂与溶液的其他组分分离。膜构型通常是交叉流的。在反渗透的情况下,膜的孔径非常小,仅允许非常少量的非常低分子量的溶质(例如截断值100mw)穿过膜。超滤是由压力梯度驱动的另一种膜分离工艺,其中膜将液体中的溶解和分散组分根据其溶剂化大小和结构分级分离。膜构型通常是交叉流的。在超滤中膜的孔径大于反渗透工艺中的孔径,因此允许某些组分与水一起通过孔。这可以是使用10,000mw截留值的分离/分级过程。渗滤是超滤的另一种类型,其涉及使用微分子可渗透过滤器,基于组分的分子大小来去除或分离溶液的组分(可渗透分子,如盐,小蛋白,溶剂等)。在蛋白质纯化和分级分离过程中通常使用的另一种工艺是,向生长培养基中加入一定浓度的高盐。蛋白质在高离子强度下其溶解度显著不同,因此“盐析”是非常有用的程序,有助于纯化和浓缩生长培养基中存在的蛋白质。硫酸铵是一种高溶解度的无机盐,在水溶液中会解离成铵和硫酸。硫酸铵作为沉淀剂特别有用,因为它高度可溶,稳定蛋白质结构,具有相对较低的密度并且相对便宜。
因此,在另一个实施方案中,本发明提供了一种干燥制剂,其包含存在于重组真菌的培养基中的多个大于5kda的大分子,所述真菌产生融合至fc结构域的外源多肽,其分泌到所述培养基中。
在又一个实施方案中,本发明提供了一种干燥制剂,其包含存在于重组真菌的培养基中的多个大于10kda的大分子,所述真菌产生融合至fc结构域的外源多肽,其分泌到所述培养基中。
在又一个实施方案中,本发明提供了一种干燥制剂,其包含存在于重组真菌的培养基中的多个大于15kda的大分子,所述真菌产生融合至fc结构域的外源多肽,其分泌到所述培养基中。
在具体的实施方案中,干燥制剂中的fc结构域是igafc结构域。
在具体的实施方案中,干燥制剂中的外源肽是预防性或治疗性肽,或其中所述外源肽是疫苗或形成疫苗的一部分。
由于难以根据其技术和结构特征来定义本发明的干燥制剂,因此我们认为在权利要求中可以将这些制剂更为充分地定义为“可通过......获得的”制剂。因此,在另一个实施方案中,本发明提供了干燥制剂,其包含对于酵母是外源的蛋白质,所述制剂通过将所述蛋白质分泌到重组酵母的培养基中、然后干燥所述培养基而获得。在又一个实施方案中,本发明提供了一种干燥制剂,其包含对于酵母是外源的治疗性蛋白质,其通过将所述治疗性蛋白质分泌到重组酵母的培养基中、然后干燥所述培养基而获得。在又一个实施方案中,本发明提供了一种干燥制剂,其包含对酵母而言是外源的预防性蛋白,其通过将所述预防性蛋白分泌到重组酵母的培养基中、然后干燥所述培养基而获得。在又一个实施方案中,本发明提供了包含蛋白质的干燥制剂,所述蛋白质形成疫苗的一部分并是酵母外源的,所述制剂通过将所述蛋白质分泌到重组酵母的培养基中、然后干燥所述培养基而获得。在又一个实施方案中,本发明提供了一种干燥制剂,其通过如下方式获得:将存在于重组真菌宿主如丝状真菌或酵母细胞的培养基中的多个大于5kda的大分子添加到口服容许的基质中,然后干燥获得的混合物,所述培养基包含对于所述重组宿主而言外源的分泌多肽。
在另一个实施方案中,本发明提供了一种干燥制剂,其包含重组宿主的培养基和口服容许的基质的混合物,所述宿主产生分泌到所述培养基中的外源多肽。
在另一个实施方案中,本发明提供了一种干燥制剂,其包含重组宿主的培养基和口服容许的基质的混合物,所述宿主产生分泌到所述培养基中的外源多肽,并且其中所述制剂的水含量通过在制备所述制剂之前浓缩所述培养基而降低。
在另一个实施方案中,本发明提供了一种干燥制剂,其包含重组宿主的培养基和口服容许的基质的混合物,所述宿主产生分泌到所述培养基中的外源多肽,并且其中所述制剂的水含量通过在制备所述制剂之前干燥所述培养基而降低。
在另一个实施方案中,本发明提供了一种干燥制剂,其包含重组宿主的培养基和口服容许的基质的混合物,所述宿主产生分泌到所述培养基中的外源多肽,并且其中所述制剂的水含量通过干燥所述制剂而降低。
在又一个实施方案中,本发明提供了一种干燥制剂,其通过将存在于重组宿主的培养基中的多个大于5kda的大分子添加到口服容许的基质中、然后干燥获得的混合物而获得,其中所述重组宿主的培养基中包含对所述重组宿主而言外源的分泌的多肽。
在又一个实施方案中,本发明提供了一种干燥制剂,其通过将存在于重组真菌细胞例如重组丝状宿主或重组酵母细胞的培养基中的多个大于5kda的大分子添加到口服容许的基质中、然后干燥获得的混合物而获得,其中所述培养基包含分泌的多肽,所述多肽是重组真菌细胞外源性的。
在另一个实施方案中,本发明提供了一种干燥制剂,其通过将存在于重组真菌细胞例如重组丝状宿主或重组酵母细胞的培养基中的多个大于5kda的大分子添加到口服容许的基质中、然后干燥获得的混合物而获得,其中所述培养基包含对所述重组真菌细胞而言是外源的分泌多肽,其中所述干燥通过喷雾干燥进行。
在另一个实施方案中,本发明提供了一种干燥制剂,其通过将存在于重组真菌细胞例如重组丝状宿主或重组酵母细胞的培养基中的多个大于5kda的大分子添加到口服容许的基质中、然后干燥获得的混合物而获得,其中所述培养基包含对所述重组真菌细胞而言是外源的分泌多肽,其中所述干燥通过冻干进行。
如本文所定义的“口服容许的基质”是在食品工业中用作载体的产品,例如淀粉,麦芽糊精,豆奶蛋白,纤维素,果胶和瓜尔豆胶。在一个具体的实施方案中,口服容许的基质是可食用的基质。在另一个特定的实施方案中,口服容许的基质是可溶性食品级营养物或可溶性食品级基质或可溶性食品级物质。
干燥制剂
在许多情况下,具有粉末形式的干燥蛋白质是有利的,这有利于将其转变成胶囊、片剂和薄膜。本领域中可用许多干燥方法将蛋白质溶液转化成干粉形式。大多数干燥方法包括通过升华(例如冷冻干燥)或蒸发(例如喷雾干燥和流化床干燥)或沉淀(例如超临界流体技术)来除去溶剂。在这些方法中,喷雾干燥和冷冻干燥是迄今为止最常用的干燥蛋白质溶液的工业方法。冻干(等效术语是冷冻干燥)是从生物药中去除水分的一种加工方法,它可以增加这些产品的稳定性、耐温性和保质期。冻干已在行业内确立,但它需要昂贵的设备,这些设备会占用生产设施中的大量空间。冻干也可能需要数天的时间来完成,需要粉状产品的制造商必须将制粒步骤纳入该工艺中。因此,冻干可用于在除去重组酵母之后通过冻干酵母发酵液来获得干燥制剂。
因此,在另一个实施方案中,本发明提供了一种生产包含重组酵母的培养基的干燥制剂的方法,其中蛋白质存在于所述培养基中,方法包括通过冻干干燥所述培养基。
喷雾干燥是保存生物药的另一种技术,它是一种将液体制剂在一个步骤中转化为干粉的工艺。通常通过首先将溶液雾化成细小液滴,然后通过使用温暖的气体在大室内将其快速干燥,来执行此工艺。用旋风分离器收集所得的干燥颗粒。喷雾干燥使生物药在雾化步骤中暴露于剪切应力,这可能会使易不稳定的生物药化合物(如蛋白质)失稳定。复杂的生物分子更难喷雾干燥,因为它们对高剪切应力敏感。遇到的剪切应力大小取决于雾化器的类型和所使用的雾化压力。可以方便地使用在小于20psig的相对较低压力下操作的音速喷嘴,该音速喷嘴可最大程度地降低剪切应力并允许处理复杂的生物药。对蛋白质,酶,抗体,病毒和细菌等多种多样的生物药已经进行过喷雾干燥。该过程去除水分并限制生物药的流动性,从而大大降低了降解率。因此,在去除重组酵母后,可以使用喷雾干燥通过将酵母发酵液喷雾干燥来获得干燥制剂。
因此,在又一个实施方案中,本发明提供了一种生产包含重组酵母的培养基的干燥制剂的方法,其中蛋白质存在于所述培养基中,方法包括通过喷雾干燥来干燥所述培养基。
在特定的实施方案中,在进行喷雾干燥之前将二糖或表面活性剂添加到培养基中。记载了dissarcharide和表面活性剂可防止聚集(broadheadj.等人(1993)j.pharm.pharmacol.46(6)458-467),并且还可以提高蛋白质荷载力(protein-loadedpower)的储存能力(adlerm和leeg(1999)j.pharm.sci.88,199-208)。
在另一个实施方案中,可以在进行喷雾干燥之前将海藻糖和/或山梨糖醇添加到培养基中。据描述,存在30%重量的山梨糖醇实质性地减少喷雾干燥期间药物蛋白质的聚集,并且还改善干燥储存稳定性(maurym.等人(2005)eur.j.ofpharmaceuticsandbiopharmaceutics59,251-261),针对海藻糖描述了类似的效果。
因此,在特定实施方案中,在进行喷雾干燥过程之前应用超声粘度降低。超声粘度降低可以允许溶液中更高的颗粒负载量,这导致必须蒸发的液体体积的减少。超声粘度降低可导致降低的能耗并更高的生产量。
喷雾干燥在设备、设施和应用方面更容易放大,且成本更低。此外,喷雾干燥的周期时间为数小时而不是数天,因此操作成本可以低于冻干的成本。
食品
在又一个实施方案中,本发明提供了一种食品,其包含如前所述的干燥制剂。
在又一个实施方案中,本发明提供了一种食品,其包含如本文之前所述的干燥制剂,其中所述食品是功能性食品。
在又一个实施方案中,本发明提供了一种食品,其包含如本文之前所述的干燥制剂,其中所述食品是药用食品。
可以根据本发明制备几种食品。食品的非限制性列表包括代餐食品,汤,面条,冰淇淋,调味酱,敷料,涂抹酱,零食,谷物食品,饮料,面包,饼干,其他烘焙产品,糖果,棒,巧克力,口香糖,乳制品和减肥产品。在us8105592(第20页第62行到第23页第35行)中讨论了后面的产品及其制备方法。
在另一个实施方案中,本发明提供了包含真菌产生的igafc融合蛋白的口服药物组合物,其中与igafc蛋白融合的蛋白是预防性或治疗性蛋白或疫苗组分。
在另一个实施方案中,本发明提供了包含真菌产生的igafc融合蛋白的口服药物组合物,其中与igafc融合的蛋白是免疫球蛋白单可变结构域。
在又一个实施方案中,本发明提供了包含真菌产生的igafc融合蛋白的口服药物组合物,其中与igafc融合的蛋白是vhh结构域。
在又一个实施方案中,本发明提供了包含真菌产生的igafc融合蛋白的口服药物组合物,其中与igafc融合的蛋白是vhh结构域,并且其中vhh结构域包含人工引入的n-糖基化位点。
在又一个实施方案中,本发明提供了一种口服药物组合物,其包含由真菌产生的具有n-聚糖和/或o-聚糖修饰的蛋白质,其中所述糖蛋白分子量的至少5%,至少10%,至少20%,至少30%,至少40%,至少50%,至少60%,至少70%,至少80%或更多由所述n-或o-聚糖贡献。
在又一个实施方案中,本发明提供了一种口服药物组合物,其包含由真菌产生的具有n-聚糖和/或o-聚糖修饰的igafc-融合蛋白,其中所述糖蛋白分子量的至少5%,至少10%,至少20%,至少30%,至少40%,至少50%,至少60%,至少70%,至少80%或更多由所述n-或o-聚糖贡献。
在又一个实施方案中,本发明提供了一种口服药物组合物,其包含由真菌产生的具有n-聚糖和/或o-聚糖修饰的igafc-融合蛋白,其中所述糖蛋白分子量的至少5%,至少10%,至少20%,至少30%,至少40%,至少50%,至少60%,至少70%,至少80%或更多由所述n-或o-聚糖贡献,并且其中与igafc融合的蛋白是免疫球蛋白单可变结构域。
值得注意的是,主要由高甘露糖基结构组成的n-糖基化结构可以使得在酵母中重组产生的igafc-融合蛋白的分子量增加50%以上。这在图5中得到了证明(请参见糖基化的v2a-igafc融合蛋白与去糖基化的v2a-igafc融合蛋白之间的差异)。同样值得注意的是,当igafc融合蛋白在酵母中在组成型gap启动子控制下产生时,与在甲醇氧化酶诱导型启动子(aox启动子)控制下产生的igafc融合蛋白相比,高糖基化的丰度可以甚至更高(参见图6)。
在另一个实施方案中,本发明提供了真菌产生的igafc融合蛋白用于口服施用。
在又一个实施方案中,本发明提供了真菌产生的igafc融合蛋白用于口服施用,其中与igafc融合的蛋白是免疫球蛋白单可变结构域。
在又一个实施方案中,本发明提供了真菌产生的igafc融合蛋白用于口服施用,其中与igafc融合的蛋白是vhh结构域。
在又一个实施方案中,本发明提供了真菌产生的igafc融合蛋白用于口服施用,其中与igafc融合的蛋白是vhh结构域,并且其中vhh结构域包含人工引入的n-糖基化位点。
在又一个实施方案中,本发明提供了具有n-聚糖和/或o-聚糖修饰的真菌产生的蛋白质用于口服施用,其中所述糖蛋白分子量的至少5%,至少10%,至少20%,至少30%,至少40%,至少50%,至少60%,至少70%,至少80%或更多由所述n-或o-聚糖贡献。
并非将本发明限制于特定机制,我们认为由真菌细胞在重组(融合)蛋白上产生的高甘露糖基化的聚糖结构保护了胃肠中的重组蛋白,使得这种高甘露糖基化的重组蛋白适用于口服递送。
术语“免疫球蛋白单可变结构域”(缩写为“isvd”),等同于术语“单可变域”,定义了其中抗原结合位点存在于单个免疫球蛋白结构域上并由单个免疫球蛋白结构域形成的分子。这将免疫球蛋白单可变结构域与“常规”的免疫球蛋白或其片段分开,在“常规”的免疫球蛋白或其片段中两个免疫球蛋白结构域,特别是两个可变结构域,相互作用形成抗原结合位点。通常,在常规的免疫球蛋白中,重链可变域(vh)和轻链可变域(vl)相互作用形成抗原结合位点。在这种情况下,vh和vl的互补决定区(cdr)都将对抗原结合位点产生贡献,即,总共6个cdr将参与抗原结合位点的形成。
鉴于以上定义,常规4链抗体(例如igg,igm,iga,igd或ige分子;本领域已知)的抗原结合结构域、或衍生自此常规4链抗体的fab片段,f(ab′)2片段,fv片段(例如二硫键连接的fv或scfv片段)或双链抗体(diabody)(均是本领域已知)的抗原结合结构域,通常不被视为免疫球蛋白单可变域,因为在这些情况下与抗原的相应表位的结合通常不由一个(单个)免疫球蛋白结构域来实现,而是由一对(缔合的)免疫球蛋白结构域,例如轻链和重链可变结构域来实现,即,由vh-vl免疫球蛋白结构域对来实现,其联合起来与相应抗原的表位结合。
相反,免疫球蛋白单可变结构域能够与抗原表位特异性结合,而无需与另外的免疫球蛋白可变结构域配对。免疫球蛋白单可变结构域的结合位点由单个vh/vhh或vl结构域形成。因此,免疫球蛋白单可变结构域的抗原结合位点由不超过三个的cdr形成。
这样,单可变结构域可以是轻链可变结构域序列(例如,vl序列)或其合适的片段;或重链可变结构域序列(例如,vh序列或vhh序列)或其合适的片段;只要它能够形成单个抗原结合单元(即基本上由单个可变域组成的功能性抗原结合单元,由此该单个抗原结合域无需与另一个可变结构域相互作用即可形成功能性抗原结合单元)。
在本发明的一个实施方案中,免疫球蛋白单可变结构域是重链可变结构域序列(例如,vh序列);更具体地,免疫球蛋白单可变结构域可以是衍生自常规四链抗体的重链可变域序列或衍生自重链抗体的重链可变域序列。
例如,免疫球蛋白单可变结构域可以是(单)结构域抗体(或适用作(单)结构域抗体的氨基酸序列)、“dab”或dab(或适用作dab的氨基酸序列)或纳米抗体(如本文所定义,包括但不限于vhh);其他单可变域,或其任何一个的任何合适的片段。
特别地,免疫球蛋白单可变结构域可以是
“vhh结构域”,也称为vhh,vhh域,vhh抗体片段和vhh抗体,最初被描述为“重链抗体”(即,“不含轻链的抗体”;hamers-castermanet等人(1993)nature363:446-448)的抗原结合免疫球蛋白(可变)结构域。选择术语“vhh结构域”是为了将这些可变结构域与常规4链抗体中存在的重链可变结构域(在本文中称为“vh结构域”或“vh结构域”)和存在于常规4-链抗体中的轻链可变结构域(在本文中称为“vl结构域”或“vl结构域”)区分开来。对于vhh和纳米抗体的进一步描述,参考muyldermans的综述文章(reviewsinmolecularbiotechnology74:277-302,2001)以及以下专利申请(这些文献被作为一般背景技术提及):布鲁塞尔自由大学的wo94/04678,wo95/04079和wo96/34103;联合利华的wo94/25591,wo99/37681,wo00/40968,wo00/43507,wo00/65057,wo01/40310,wo01/44301,ep1134231和wo02/48193;vlaamsinstituutvoorbiotechnologie(vib)的wo97/49805,wo01/21817,wo03/035694,wo03/054016和wo03/055527;algonomicsn.v.和ablynxn.v.的wo03/050531;加拿大国家研究委员会的wo01/90190;抗体研究所的wo03/025020(=ep1433793);以及ablynxnv的wo04/041867,wo04/041862,wo04/041865,wo04/041863,wo04/062551,wo05/044858,wo06/40153,wo06/079372,wo06/122786,wo06/122787和wo06/122825;以及ablynxnv的其他公开的专利申请。也参考这些申请中提到的其他现有技术,特别是国际申请wo06/040153第41-43页提及的参考文献列表,该列表和参考文献通过引用并入本文。如这些参考文献中所述,纳米抗体(特别是vhh序列和部分人源化的纳米抗体)尤其可以通过在一个或多个构架序列中存在一个或多个“hallmark残基”来表征。纳米抗体的进一步描述,包括纳米抗体的人源化和/或驼源化以及其他修饰、部分或片段、衍生物或“纳米抗体融合物”、多价构建体(包括接头序列的一些非限制性示例)和用于延长纳米抗体的半衰期的不同修饰及其制备,可以见于例如在wo08/101985和wo08/142164中。对于纳米抗体的进一步一般描述,参考本文引用的现有技术,例如wo08/020079(第16页)中的描述。
“结构域抗体”,也称为“dabs”、“域抗体”和“dabs”(术语“域抗体”和“dabs”由葛兰素史克集团公司使用作为商标),已经描述在例如ep0368684,ward等人(nature341:544-546,1989),holt等(tendsinbiotechnology21:484-490,2003)和wo03/002609以及例如wo04/068820,wo06/030220,wo06/003388和domantisltd.的其他公开专利申请。结构域抗体基本上对应于非驼类哺乳动物,尤其是人4链抗体的vh或vl结构域。为了以单抗原结合结构域(即,不与vl或vh结构域配对)结合表位,需要对这种抗原结合特性进行特异性选择,例如,通过使用人单vh或vl结构域序列的文库。像vhh一样,结构域抗体具有约13至约16kda的分子量,并且如果衍生自完整的人序列,则不需要人源化用于例如在人类中的治疗用途。
还应注意,单可变结构域可衍生自某些鲨鱼物种(例如,所谓的“ignar结构域”,参见例如wo05/18629)。
因此,在本发明的意义上,术语“免疫球蛋白单可变结构域”或“单可变结构域”包括衍生自非人类来源的多肽,优选衍生自驼科动物,优选驼科动物重链抗体。如前所述,它们可以被人源化。此外,该术语包括衍生自非驼科动物来源例如小鼠或人类的多肽,它们已经“驼源化”,例如如davies和riechmann(febs339:285-290,1994;biotechnol。13:475-479,1995;prot.eng.9:531-537,1996)和riechmann和muyldermans(j.immunol.methods231:25-38,1999)中描述的。
在又一个实施方案中,本发明提供了一种生产干燥制剂的方法,所述干燥制剂包含重组酵母的培养基,其中蛋白质存在于所述培养基中,所述方法包括:
i)培养重组酵母,所述重组酵母包含蛋白质并允许将所述蛋白分泌到培养基中,
ii)将培养基与重组酵母细胞分离,
iii)在膜分离过程中浓缩所述培养基,和
iv)干燥所述浓缩的培养基。
产生重组酵母的方法是本领域众所周知的。简而言之,在重组酵母中存在包含编码重组蛋白的嵌合基因的表达载体。表达载体可以整合到基因组中,或者可以在重组酵母中自主复制。整合到宿主染色体中的载体由于在没有选择的情况下其有丝分裂的稳定性而被广泛使用。然而,一些酵母系统存在游离型表达载体。表达载体通常包含强酵母启动子/终止子和酵母选择标记盒。大多数酵母载体可以在大肠杆菌中繁殖和扩增以有利于克隆,并因此也包含大肠杆菌复制起点和氨苄青霉素选择标记。最后,许多酵母表达载体包括可选地克隆基因在有效分泌前导序列下游的能力(例如,交配因子或ost1序列的有效分泌前导序列(fitzgeraldi&glickbs(2014)microbialcellfactory13,1)),由此可以有效地引导异源蛋白质从细胞中分泌出来。嵌合基因可以包含与编码信号序列的核酸序列可操作地偶联的启动子,该信号序列编码序列与编码有用多肽的重组基因可操作地偶联。启动子可以是组成型启动子或诱导型启动子。
在又一个实施方案中,本发明提供了一种生产干燥制剂的方法,所述干燥制剂包含重组酵母的发酵液,其中蛋白质存在于所述培养基中,所述方法包括:
i)培养重组酵母,所述重组酵母包含蛋白质并允许将所述治疗性蛋白质分泌到培养基中,
ii)干燥所述发酵液(包含重组酵母和培养基)。
在另一个实施方案中,本发明提供了进一步包含酵母细胞的本发明干燥制剂。
在另一个实施方案中,本发明提供了进一步包含非重组酵母细胞的本发明干燥制剂。
在又一个实施方案中,本发明提供了一种生产干燥制剂的方法,所述干燥制剂包含重组酵母的培养基和非重组酵母细胞,其中外源蛋白质存在于所述重组酵母的培养基中,所述方法包括:
i)培养重组酵母,所述重组酵母包含外源蛋白质并允许将所述蛋白质分泌到培养基中,
ii)分离重组酵母细胞与培养基,
iii)在膜分离过程中浓缩所述培养基,
iv)向培养基中添加非重组酵母细胞
v)干燥所述培养基。
在又一个实施方案中,本发明提供了一种生产干燥制剂的方法,所述干燥制剂包含重组酵母的培养基和非重组酵母细胞,其中外源蛋白质存在于所述重组酵母的培养基中,所述方法包括:
i)培养包含外源蛋白质的重组酵母,并允许将所述外源蛋白质分泌到培养基中,
ii)分离重组酵母细胞与培养基,
iii)干燥所述培养基,
iv)将非重组酵母细胞的干燥制剂加入到步骤iii)中获得的干燥培养基中。
在另一个实施方案中,本发明提供了本发明的干燥制剂,其进一步包含与酵母发酵液中存在的蛋白质不同的富含蛋白质的制剂。
在又一个实施方案中,本发明提供了一种生产干燥制剂的方法,所述干燥制剂包含重组酵母的培养基和富含蛋白质的制剂,其中外源蛋白质存在于所述培养基中,所述富含蛋白质的制剂不同于酵母发酵液中存在的蛋白质,方法包括::
i)培养包含外源蛋白质的重组酵母,并允许将所述蛋白质分泌到培养基中,
ii)从培养基中分离重组酵母细胞,
iii)向培养基中添加富含蛋白质的制剂,
iv)干燥所述培养基。
在又一个实施方案中,本发明提供了一种生产干燥制剂的方法,所述干燥制剂包含重组酵母的培养基和非重组酵母细胞,其中治疗性蛋白质存在于所述培养基中,所述方法包括:
i)培养包含治疗性蛋白的重组酵母,并允许将所述治疗性蛋白分泌到培养基中,
ii)从培养基中分离重组酵母细胞,
iii)干燥所述培养基,
iv)将富含蛋白质的制剂的干燥制剂加入到步骤iii)中获得的干燥培养基中。
在又一个实施方案中,本发明提供了包含重组酵母的培养基的干燥制剂,所述酵母产生分泌到培养基中的外源性fc融合蛋白。
在又一个实施方案中,本发明提供了包含重组酵母的培养基的干燥制剂,所述酵母产生分泌到培养基中的外源性fc融合蛋白,并且其中所述fc结构域是igafc结构域。
由于难以根据其技术特征在结构上描述本发明的干燥制剂,因此更合适的是将干燥制剂定义为“可通过……获得”的权利要求形式。因此,在另一个实施方案中,本发明提供了干燥制剂,其包含在fc结构域和多肽之间的外源融合蛋白,所述制剂是通过将所述外源蛋白分泌到重组酵母的培养基中、然后干燥所述培养基而获得的。
fc融合蛋白
fc区(片段可结晶区)是免疫球蛋白的尾部区域,可与称为fc受体的细胞表面受体和补体系统的某些蛋白质相互作用。根据特别设想的实施方案,fc融合蛋白中的fc区是来自免疫球蛋白g(igg)同种型的fc区。这可以是任何igg亚类(人类中的igg1、2、3、4)。对于igg,如iga和igd同种型,fc区由两个相同的蛋白质片段组成,该片段来自抗体两条重链的第二和第三恒定域。在另一个实施方案中,融合蛋白中的fc部分衍生自iga抗体。如本文所用,“fc融合蛋白”是融合蛋白,其中fc区与蛋白质或肽融合。一类特定的含fc蛋白是可以结合抗原的含fc蛋白。实例是其中fc区连接至结合部分(例如,纳米抗体,fab区,f(ab′)2区)的抗体或融合蛋白。另外,本发明不限于人序列。例如,fc区可是小鼠或驼科的,恒河猴,狗,牛,豚鼠,绵羊,猪,山羊,马,大鼠,兔,猫或任何其他哺乳动物的fc区。fc区甚至可来自非哺乳动物(例如鸡)。在具体的实例中,结合部分可以是非抗体支架。非抗体支架大致分为两个结构类别,即结构域大小的化合物(分子量为6-20kda)和受约束的肽(2-4kda)。结构域大小的支架包括亲和体(affibody),affilins,anticalins,atrimers,darpins,fn3支架(例如adnectins和centyrins),fynomers,kunitz域,pronectins和obodies,而avimers,双环肽和cys-knots与肽相关(请参见vazquez-lombardiretal(2015)drugdiscoverytoday20,10,1271中的全面综述)。
药物制剂
在一个具体的实施方案中,本发明的干燥制剂可以用任何可得的软胶囊或硬胶囊技术包囊以产生固体口服药物剂型,其可以进一步包含肠溶衣或延迟释放包衣。
在另一方面,可将干燥制剂溶解在液体中以获得乳液(参见moreiratc等人(2016)colloidssurfb.biointerface143:399-405)。因此,在特定方面,药物制剂是液体。一方面,根据本发明的药物制剂是液体,并且包含少于10%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于9%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于8%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于7%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于6%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于5%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于4%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于3%(w/w)的水。一方面,本发明的药物制剂是液体,并且包含少于2%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含少于1%(w/w)的水。一方面,根据本发明的药物制剂是液体,并且包含小于0%(w/w)的水。
在本发明的某些方面,药物制剂可包含通常存在于药物制剂中的其他赋形剂,此类赋形剂的实例包括但不限于抗氧化剂,抗微生物剂,酶抑制剂,稳定剂,防腐剂,调味剂,甜味剂和其他成分,如在handbookofpharmaceuticalexcipients,rowe等人编,第7版,pharmaceuticalpress(2012)中描述的那样,其通过引用并入本文。
这些另外的赋形剂的量可为总药物制剂的约0.05-5重量%。抗氧化剂,抗微生物剂,酶抑制剂,稳定剂或防腐剂通常占药物制剂总重量的至多约0.05-1%。甜味剂或调味剂通常提供总药物制剂重量的至多约2.5%或5%。
根据本发明的口服药物制剂可以配制成固体剂型。
根据本发明的口服药物制剂可以配制成固体剂型,并且可以选自胶囊,片剂,糖衣丸,丸剂,锭剂,散剂和颗粒剂。
根据本发明的口服药物制剂可以配制成多颗粒剂型。
根据本发明的口服药物制剂可以被配制成多颗粒剂型,并且可以选自软胶囊或硬胶囊、带肠溶衣的软硬胶囊中的丸、微粒、纳米颗粒、液体或半固体填充制剂。
一方面,口服药物制剂可以用一种或多种包衣例如肠溶衣制备,或者根据本领域众所周知的方法配制成延迟释放制剂。
一方面,根据本发明的药物制剂用于制备药物。
如本文所用,术语“表面活性剂”是指可以吸附在表面和界面处的任何物质,特别是洗涤剂,例如但不限于液体-空气,液体-液体,液体-容器或液体-任何固体的界面。
本文所用的术语“药”、“治疗性”或“药物”是指药物制剂中使用的活性成分,可用于预防、治疗或疫苗应用,因此也指在本专利申请中定义为“大分子治疗性”或“治疗性大分子”或“预防性大分子”或“疫苗大分子”的物质。
预防/治疗/疫苗
显然取决于肽的性质,包含多肽的本发明干燥制剂可以用于多种疾病。例如,当肽是治疗性肽时,则几种疾病包括但不限于神经退行性疾病,癌症,血液学疾病,免疫学疾病,心脏疾病,肝脏疾病,呼吸系统疾病,吸收不良疾病,糖尿病,病毒感染,真菌感染,细菌感染,眼部疾病,罕见的代谢紊乱和高血压。
在一个具体的实施方案中,本发明提供了用于治疗胃肠道疾病的本发明的干燥制剂。胃肠道疾病的非限制性实例包括肠易激综合症,便秘,痔疮(例如内痔),肛裂,憩室病,结肠息肉,结肠癌,感染性结肠炎,溃疡性结肠炎,克罗恩病,缺血性结肠炎,放射性结肠炎和肠粘膜炎。
在另一个具体的实施方案中,本发明提供了用于治疗颊(或口腔)疾病的本发明干燥制剂,例如唇疱疹(coldsores),口疮(cankersores),鹅口疮(thrush),粘膜白斑病(leukoplakia),口干症(drymouth),牙龈病,口臭,龋齿,牙周疾病(例如牙龈炎),口腔念珠菌病,口腔单纯疱疹病毒感染,口腔人乳头瘤病毒感染,复发性口腔溃疡,口腔和咽喉癌以及口腔粘膜炎。
存在于本发明的干燥制剂中的治疗性蛋白质包括单克隆抗体,生长因子,白介素等。在另一方面,外源多肽可用于疫苗目的。特别地,外源多肽可以单独用作疫苗或可以形成疫苗组合物的一部分。
重组酵母
在一个具体的实施方案中,用于制备本发明的干燥制剂的重组酵母是已获得gras状态的酵母物种。gras代表“一般被视为安全”。具有gras状态的酵母包括酵母,如酿酒酵母(saccharomycescerevisiae),巴斯德毕赤酵母(pichiapastoris),多形汉逊酵母(hansenulapolymorpha),解脂耶氏酵母(yarrowialipolytica)和乳酸克鲁维酵母(kluyveromyceslatis)。重组酵母中治疗性蛋白质的产生是本领域技术人员众所周知的。对于酵母巴斯德毕赤酵母,参见例如julienc(2006)bioprocessinternational,january,p22-31的综述;对于酿酒酵母(saccharomycescerevisiae),参见例如nielsenj(2013)bioengineered4:4,207-211的综述;对于多形汉逊酵母(hansenulapolymorpha),参见coxh.等(2000)yeast,第16卷,13,pp.1191-1203的综述;对于乳酸克鲁维酵母(kluyveromyceslactis),参见vanooyenajj等人(2006)femyeastres6,381-392的综述;对于解脂耶氏酵母(yarrowialipolytica),参见madzakc等人(2004)j.biotechnol.109(1-2):63-81的综述。
应当理解,尽管本文已经讨论了根据本发明的工程化重组酵母细胞和方法的特定实施方案、特定构型以及材料和/或分子,但是可以进行形式和细节上的各种改变或修饰而不脱离本发明的范围和精神。提供以下实施例以更好地举例说明特定实施方案,并且不应认为它们限制了本申请。本申请仅受权利要求书的限制。
实施例
1,口服递送毕赤酵母产生的单体vhh-igafc融合蛋白可有效预防仔猪f4-etec感染
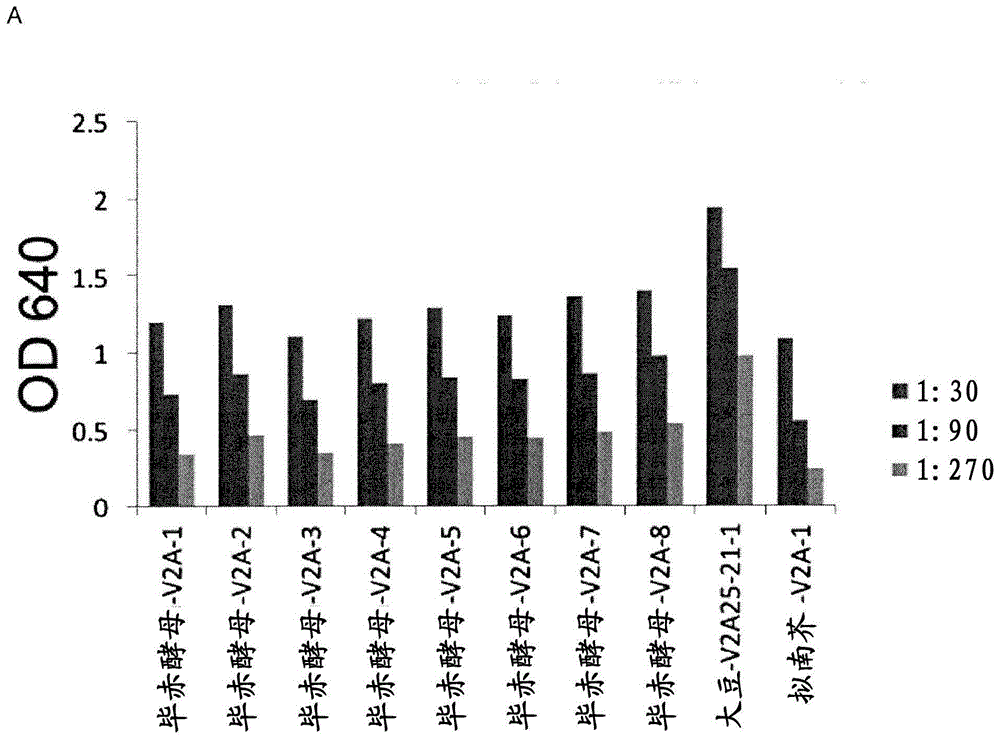
先前在拟南芥种子中生成和评估的单体vhh-igafc融合蛋白(分别称为v2a和v3a)(参见virdi等(2013)110,29,11809-11814),现在也已在酵母毕赤酵母和在大豆种子中产生。简而言之,产生了针对f4+etec菌毛的美洲驼仅重链抗体(vhh)。将选定的f4+etecvhhs嫁接到猪igab的密码子优化部分上。所得的vhh-igafc融合蛋白称为v2a和v3a。使用具有f4-etec尖端粘附抗原-faeg包被的孔的elisa,评估功能性vhh-igafc融合蛋白的表达水平,并用与辣根过氧化物酶偶联的抗猪iga检测。图1显示了筛选毕赤酵母克隆(见图1a)或大豆种子储备物(seedstocks)(见图1b)的典型示例。针对毕赤酵母产生的抗体v2a和v3a,各筛选了二十个单独的菌落。类似地,从每个转化的大豆事件,筛选了10-12粒种子。分别针对v2a和v3a筛选了5个此类事件。保留了高表达的种子储备物,其中一部分也用于产生t3纯合种子储备物。大豆产生的v2a和v3a的表达水平经计算为种子重量的约0.2%,这与拟南芥种子中的表达水平相似。分泌的毕赤酵母v2a和v3a的表达水平高达100mg/l(在用考马斯蓝染色的sds-page凝胶上分析)。但是,基于elisa的功能分析显示,1ml毕赤酵母上清液相当于1ml大豆或拟南芥种子提取物(5mg种子)(见图2)。在此elisa设置中,很可能,由于毕赤酵母聚糖引起的屏蔽作用,对毕赤酵母产生的vhh-igaf的定量检测受到阻碍。实际上,毕赤酵母上清液(参见图2c)和大豆种子提取物(参见图1d)的免疫印迹分析显示了差异糖基化的明显标志。毕赤酵母vhh-igafc迁移高于50kda
毕赤酵母和大豆产生的vhh-igafc融合蛋白可防止仔猪etec感染
在仔猪饲喂攻击实验中评估了毕赤酵母和大豆种子生产的v2a和v3a的vhh-igafc抗体混合物的体内功效(剂量为5mg/猪/天)。先前制备的由拟南芥产生的单体vhh-igafc(参见virdi等人,2013,pnas110,29,11809-11814)用作参考(拟南芥组),而不含抗体的饲料(亚麻饲料,图3a)作为阴性对照。每组中的六只f4-etec血清阴性和f4-受体(f4r)基因型阳性仔猪接受为期10天的实验饲料。在第三天,所有的仔猪连续两天(第0天和第1天)受到1010f4-etec细菌的攻击,并通过分析攻击菌株的每日排出,直至第14天,来监测感染的结果(图3b)。在第14天对仔猪实施安乐死,此时还测定了空肠、回肠和盲肠内容物中的f4-etec细菌(表1)。
表1:每头仔猪每克粪便中f4-etec的粪便排放量(log10)
值得注意的是:处死后的观察结果表明,拟南芥组的仔猪18患有脐疝,小肠极度绞窄,导致阻塞性通道。因此,仔猪18的数据不包括在排放图(图3b)和统计分析(数据记录在表1)中。
也在安乐死之后,使用肠绒毛肠上皮细胞进行的f4-etec粘附试验显示,每250μm细胞表面结合了41至85个细菌,双重确认了大量f4-受体(f4r)的表型表达。因此,除了18号仔猪之外,所有仔猪的数据都用于评估单体vhh-igafc的功效。
排放数据表明,在毕赤酵母、拟南芥种子或大豆种子中产生的单体vhh-igafcv2a和v3a混合物,以每天5mg/猪的剂量,成功预防了etec感染。vhh-igafc接受组的排放显著较低(参见图3b,表1)。在第2天,记录到在三组中最高的平均排放量,其中对于大豆、毕赤酵母和拟南芥组,分别为每克粪便中有4.5(log10)、3.7(log10)和4(log10)细菌。后一天,这三组的排放量下降了一个log。在第5天,大豆、毕赤酵母和拟南芥组中的平均排放量分别达到每克粪便2.6(log10)、2(log10)和2,7(log10)细菌。此后,这三组保持低的排放量,其中三组的一些小猪常具有低于可检测水平的排放量(每克粪便中2(log10)个细菌)(见表1)。相反,未在饲料中接受抗体的组(对照组)的仔猪具有高滴度攻击细菌的延长排放,从第3天到第6天平均为每克粪便中高于6.3(log10)细菌;并且此后在第7天(每克粪便中5.3(log10)个细菌)和第8天(每克粪便中3.3(log10)个细菌)下降,到第9天降至检测水平以下。长时间的高排放表明,攻击菌株可以有效地建立感染并成功定殖于对照组的小肠中。而拟南芥组、毕赤酵母组和大豆组中接受饲料中vhh-igafc抗体的仔猪,在攻击后都立即显示出f4-etec的迅速下降。这表明,这些饲料中的单体vhh-igafc阻止了f4-etec附着在肠上皮细胞上、定植和建立感染。抗f4-etec血清转化进一步证实了这一点(见图3c)。由于f4-etec病原体有限地暴露于免疫系统,因此,毕赤酵母、大豆和拟南芥组中的大多数仔猪均产生了较低的免疫应答,而对照组的抗f4-etec血清igg、igm和iga的平均滴度稳步上升到第7天,并继续上升到第14天。
排放和血清转化结果清楚地表明,由拟南芥、大豆或毕赤酵母中生产的vhh-igafc抗体——v2a和v3a——按等比例组成的针对f4-etec的单体vhh-igafc制剂,当以5mg剂量在饲料中递送时,是有效的。此外,在毕赤酵母产生的抗体的情况下,体内功效结果充分地证实了,对带有分泌的vhh-igafcs的培养基的加工和配制,适于稳定地掺入和口服递送基于饲料的毕赤酵母产生的分子,用于胃肠适应症。
实施例1的材料和方法
vhh-igafc的表达
拟南芥:先前公开的表达单体v2a-igafc和v3a-igafc融合蛋白的拟南芥品系(virdi等人(2013)pnas110,29,11809-11814),在温室中扩大规模,产生约100克的v2a-igafc和v3a-igafc生产种子,在攻击实验中配制用于拟南芥组的含抗体的饲料。
大豆:将分别带有抗体v2a和v3a的vhh-igafc融合基因的质粒pev2a和pev3a(virdi等人(2013)pnas110,29,11809-11814),根据
毕赤酵母:使用引物对alfa-v2(ctctctcgagaagagagaggccgaagctcaggtgcagctgc)以及iga-noti(cctcttgagcggccgccctttagtagcatatgccttctg),pcr扩增vhh-igafcv2a和v3a的vhh-igafc融合基因,如先前由demeyert.等针对vhh-igg所描述的((2015)plantbiotechnolj13,938-947),这些引物带有限制性酶切位点avai和noti,通过这些位点,将抗体基因克隆在pppt4_alpha_s表达载体中并与α-交配因子一起在读框中(naatsaaril.等(2012)plosone7,e39720)。使用酶pmei线性化各个表达载体,并通过电穿孔将其导入巴斯德毕赤酵母中(jacobspp等人(2009)natprotoc.4,58-70)。在铺有100μg/ml
仔猪攻击实验
经比利时根特大学兽医学院动物护理和伦理委员会批准(伦理档案号ec2015/47),根据比利时动物福利法规进行了仔猪攻击实验。对于用毕赤酵母和大豆生产的单体iga进行的实验,从比利时梅勒农业和渔业研究所(ilvo)的农场,购买了来自未接种疫苗的母猪的仔猪(比利时地方品种x英国地方品种)。从2-3周龄的仔猪采集血样,以监测血清中抗etec抗体的水平(由ilvo的机构伦理委员会批准,档案号-2016/267)。通过muc-13pcr(goetstouwers等(2014)plosone9,e105013)测定,选择了如下仔猪,所述仔猪对f4-etec呈血清阴性且对muc-13基因呈阳性(纯合和杂合显性),这与f4-etec受体(f4r)的存在相关。每个实验组由六个仔猪组成。断奶后,将仔猪带到专门的猪圈,并根据仔猪的所在窝、基因型和体重,适当地将其随机分配到喂养组中。每组的平均起始重量为7.2kg。如先前所述进行攻击(virdi等人(2013)procnatacadsciusa.110,29,11809-11814)。简而言之,在用碳酸氢盐缓冲液中和胃内ph30分钟后,在镇静下通过胃内插管,连续数天用1010f4-etec细菌(gis26rstrep菌株)攻击仔猪。攻击的第一天在时间轴中记为第0天。从攻击前三天开始,将含抗体的饲料施用10天(图3,a)。从攻击当天至第12天以及在安乐死当天,收集粪便样品,在带有链霉素选择(1mg/ml)的血液琼脂平板上监测f4-etec攻击菌株gis26rstrep的排放。在第-1天,第7天和第14天采集血样以监测f4-etec特异性igg,iga和igm滴度。在图3a中示意性地表示了具体的改动、样品收集和动物操作。
饲料准备:
对于种子产生的抗体,将经过精确称重的各个种子(大豆或拟南芥)压碎,然后分两步与基础猪饲料混合,以确保完全均匀。首先以小体积产生浓缩的预混合物,然后将其用更多的猪饲料稀释以制备实验饲料(表2)。使用刀磨机(retschgrindomixgm200)研磨种子,之前使用干冰将种子和研磨室冷却。为了在每个实验中在所有组中保持成比例的营养,使用亚麻籽替代拟南芥种子(表2)。类似地,为了考虑额外的大豆蛋白,将野生型大豆种子以与产生iga的大豆种子等比例的量添加到除大豆iga组之外的组中(表2)。
表2:饲料配方。星号(*)是指,来自每个生产批次的四批冷冻干燥浆料(由经过滤和缓冲液交换的毕赤酵母培养基和仔猪饲料制成)的汇合干燥毕赤酵母混合物的总重量。占总饲料的百分比在括号中显示。
毕赤酵母:
根据基于elisa的当量测试(图2),毕赤酵母提取物1毫升相当于5毫克生产vhh-igafc的拟南芥种子粉末溶于一毫升提取缓冲液中。基于该比例,为了向6只仔猪提供5mg剂量的毕赤酵母产生的vhh-igafcs,为期10天,产生了30l毕赤酵母培养基的必需量。按照上述标准表达方案(在bmgy培养基中生长48小时,然后在bmmy培养基中诱导48小时),进行4批7.5l的每周培养,制成该30l的毕赤酵母培养物。在每次运行结束时,通过离心收集培养基,将无细胞的上清液通过渗滤浓缩至2-1.5l,然后使用装有5kdaomegatmcentramate过滤器盒的centramatetm500s切向流过滤系统(palllifescience),用具有18.75mmnacl的磷酸钠缓冲液(ph6)进行缓冲液交换。向所得的含有毕赤酵母产生的vhh-igafc的约2-1.5l蛋白质溶液中(在四批试验的每批结束时获得),加入等重量的商品猪饲料,并用手持式桨叶混合以避免起泡,并且使用冷冻干燥(epsilon2-10dlsc-martin-christ,德国)将浆液冻干47小时。4个批次的所得的干粉(称为毕赤酵母预混物)总共为5.368kg,然后将其与猪饲料混合以得到18kg最终的带有毕赤酵母生产的vhh-igafc的饲料(表2)。
统计分析:
使用线性混合模型,使用来自sas的混合程序(sas系统的9.4版,适用于windows7,64bit,版权所有
2.评估在毕赤酵母中产生的igafc与人il-22的融合蛋白
材料:
重组鼠tnf(mtnf)是内部在大肠杆菌产生的,比活为9.46×107iu/mg。
动物:
从geertvanloo教授获得a20条件性基因敲除小鼠(a20iec-ko)。a20iec-ko小鼠的肠上皮细胞(iec)缺失a20(vereecke,l.等人(2010)实验医学杂志207,1513–1523)。为了进行实验,仅使用8-12周大的雌性小鼠。将小鼠笼养在vib炎症研究中心(irc)的专门无病原体设施的单独通风的笼子中。所有小鼠实验均根据机构、国家和欧洲动物法规进行。动物实验方案经根特大学伦理委员会批准。
饲料:
从荷兰bio-service购买了标准的粉状小鼠饲料(
体外测试:
为了测试生产培养基中不同il-22形式的生物活性,在进行生物测定之前,对培养基取样并在无菌pbs中稀释培养基。为了测试不同il-22形式在干燥饲料中的生物活性的保留,将饲料以1:1(v/v)溶于pbs。然后将浆液以13.000g离心。将上层水相转移至新管中,并使用低蛋白结合的0.22μm注射器式过滤器(millipore)进行过滤除菌。然后在进行生物测定之前,将滤液在无菌pbs中稀释。
从美国典型培养物保藏中心(atcc)订购了人类colo-205结肠癌细胞,并根据数据表中提供的指南进行培养。简而言之,将细胞系作为半贴壁细胞在补充了10%胎牛血清(fbs)的rpmi1640(gibco)中于37℃5%co2下培养。为了传代,收集在悬浮液中生长的细胞,并按照标准组织培养程序用胰蛋白酶消化贴壁的细胞。
为了使用colo-205分析法测定il-22的生物活性,将细胞以3.0×105细胞/ml(100微升/孔)的密度接种在96孔u型底板中。使细胞适应24小时,然后用一系列稀释的含il-22的级分刺激细胞。允许所有刺激进行过夜。作为对照,使用商业上可获得的生产重组hil-22(无载体)的大肠杆菌(biolegend)的稀释系列。第二天,将板在4℃下400g离心10分钟,并收集上清液。使用hil-10duosetelisa(r&d系统)测定上清液中的il-10。在graphpadprism6中分析数据。基于剂量反应曲线确定比活性,用于确定ec50。
实验模型:
为了测试il-22的保护作用,向c57bl/6a20iec-ko小鼠(每组n=8)管饲10ml/kg的液体饮食(相当于200μl),其中含相当于20、10、5、1μg的每种形式的il-22,或管饲不含il-22的等效饲料作为对照。
为了诱发实验性结肠炎,在管饲后1小时,腹膜内(i.p.)给予亚致死剂量的250μg/kg的mtnf。给对照组小鼠(模拟)腹膜内注射等体积的0.9%nacl。每小时监测一次体温和存活率。在平行实验中,4小时后对小鼠进行安乐死以进行组织学分析和caspase活性测定。
组织学:
动物处死后,将整个结肠取出,从盲肠至肛门,并测量结肠长度作为炎症的标志。测量后,从小肠中取出部分并固定在4%多聚甲醛(pfa)中。石蜡包埋和切片后,组织用苏木精/曙红染色以进行组织学检查。使用tunel染色的原位细胞死亡检测试剂盒(roche),通过荧光显微镜分析了细胞凋亡。
血清分析:
分别使用小鼠il-6duosetelisa试剂盒(r&dsystems)和小鼠ccl2/je/mcp-1duosetelisa试剂盒(r&dsystems),测定血清中的血清促炎细胞因子il-6和mcp-1。通过在hitachi747分析仪(diagnostica,boehringermannheim)上的常规光度测试,分析丙氨酸氨基转移酶活性(alt)和天冬氨酸氨基转移酶活性(ast)。
结果:
模拟处理的a20iec-ko小鼠没有表现出任何困扰的表型迹象。相比之下,接受tnf注射的小鼠在注射后2小时立即显示出明显的tnf毒性症状,包括体温过低和严重腹泻。另外,在注射tnf后5至9小时之间,小鼠开始死亡。用含il-22的饲料(il-22igafc或il-22)进行胃内治疗的小鼠,仅显示体温的轻度降低,不出现腹泻。此外,用任何il-22形式治疗的小鼠均未在tnf攻击后死亡。
从组织学上讲,仅接受tnf处理但未接受il-22的小鼠在回肠和空肠具有严重损伤,特征在于广泛的上皮损害和隐窝-绒毛结构几乎完全丧失。相反,接受任何il-22形式的小鼠均不显示任何损伤迹象,清楚地保持了屏障完整性。在细胞水平上,il-22处理后缺乏tunel染色,而在仅用tnf处理的小鼠中,除了已经脱离并在肠腔中发现的细胞外,绒毛上皮内衬中的凋亡细胞高度丰富。
我们通过测量促炎细胞因子il-6和mcp-1进一步评估了全身效应。对于对照小鼠和接受il-22形式的小鼠,两者的水平是可比较的,而仅接受tnf但未接受il-22处理的小鼠具有升高的ii-6和mcp-1水平。此外,我们还评估了肝脏转氨酶水平(ast和alt),因为有报道说tnf也影响肝脏的生理功能。实际上,用tnf处理的小鼠在血清中具有升高的ast和alt水平,而il-22处理的小鼠则没有。
实施例2的材料和方法
菌株,培养基和试剂
大肠杆菌(e.coli)mc1061或dh5α用于标准分子生物学操作。为了进行质粒繁殖,将大肠杆菌在lb肉汤(0.5%酵母提取物,1%胰蛋白胨和0.5%nacl)中培养,所述lb肉汤中补充了适当的抗生素:50μg/ml羧苄青霉素(duchefabiochemie),50μg/ml卡那霉素(sigma)aldrich),50μg/ml潮霉素b(duchefabiochemie)或50μg/ml
巴斯德毕赤酵母nrrl-y11430菌株(同名komagataellaphaffi)由a.glieder(奥地利格拉茨技术大学)提供。该菌株被称为野生型。酵母培养物在液体ypd(1%酵母提取物,2%蛋白胨,1%d-葡萄糖)中或在固体ypd-琼脂(1%酵母提取物,2%蛋白胨、1%d-葡萄糖,2%琼脂)上生长,通过使用适当的抗生素进行选择:100μg/ml
为了蛋白质表达,将培养物在bmgy(1%酵母提取物,2%蛋白胨,100mmkh2po4/k2hpo4、1.34%ynb,1%甘油,ph5.5)中在摇动孵育器(28℃,225rpm)中生长。为了诱导蛋白质表达,将细胞切换至含有1%meoh的bmmy(1%酵母提取物、2%蛋白胨、100mmkh2po4/k2hpo4、1.34%ynb,ph5.5)。为保持诱导并补偿蒸发,每8-12小时向培养物中掺加1%meoh。在诱导结束时,通过离心(1.500g,4℃10分钟)收获培养物。立即分析样品,或在液氮中速冻然后保存在-20℃。
hil-22表达载体的构建
使用genscript的专利算法对成熟人白介素22的开放阅读框(hil-22,uniprotkb登录号q9gzx6,残基34-179)进行密码子优化以用于巴斯德毕赤酵母中的表达,并合成订购。将hil-22编码序列克隆在pkai61ea-yegfp表达载体中,与酿酒酵母α-交配因子在读框中,但不包括ea重复序列(schoonooghes等人(2009)bmcbiotechnology9,70)。最终表达载体pkai-hil22包含在强甲醇诱导型aox1启动子控制下的hil-22转基因,并具有
作为替代的分泌信号,使用引物ost1saccharopaox1fw和ost1saccharorv从酿酒酵母基因组dnapcr扩增酿酒酵母ost1序列(fitzgeraldi和glickbs(2014)microbialcellfactories,13,1)。
hil-22-higa_fc-6xhis表达载体的构建
使用模块克隆策略(moclo)克隆所有构建体。该系统是为酿酒酵母开发的(leeme等(2015)acssynthbiol4,975-986),但在内部进行了改造以用于毕赤酵母中。简而言之,moclo系统利用标准化的进入载体(entryvector)来亚克隆所需构建体的各个“部分”。这些载体中的每个部分的侧翼是不同的ii型限制酶位点。通过在单个反应中合并这些进入载体并进行连续的限制性酶切消化和t4连接反应,使具有相容末端的部分组装并连接成环状质粒,以产生最终表达载体。通过合并等摩尔量(约20飞摩尔)的扩增片段和moclo进入载体pytk001或合成的但相同的载体pptk081,生成了所有进入载体。使用该标准的模块克隆(“moclo”)方案,并涉及在合并的序列中添加ii型限制酶bsmbi(1u;neb)、t4连接酶(1u;neb)和10xt4连接酶缓冲液(neb)。在pcr循环仪中进行了42℃2分钟和16℃5分钟的24个循环,然后在50℃下进行了10分钟的最终消化步骤,并在80℃下进行了另外10分钟的热变性步骤,然后保持在12℃。将得到的载体引入大肠杆菌mc1061感受态细胞,然后将其铺在lb氯霉素培养基上。由于pytk001/pptk081受体进入载体具有gfp缺失盒(gfpdropoutcassette),可以进行绿白筛选,从绿色假阳性克隆中区分出包含(正确)质粒的菌落。分离质粒,并使用引物pp001和pp002通过sanger测序法验证目的序列。
为了构建具有恒定iga结构域的hil-22融合构建体(paox1-hil-22-higa_fc-6xhis),使用与moclo系统相容的引物,通过pcr扩增α-mf/ost1/pre-hil-22序列。为了将hil-22序列与higa序列进行n端融合,将完整的α-mf-hil-22cds,ost1-hil-22cds和pre-hil-22cds视为moclo系统中的n端标签或3a部分。类似地,iga的恒定结构域(人,小鼠和猪)被视为目标基因或3b部分。
携带α-mf-hil-22,ost1-hil-22或pre-hil-22的moclo进入载体都是以上述pppt4_alpha_s表达载体为模板通过初始pcr扩增而产生的。用引物il-22forentryfw和il-22forentryrv扩增α-mf-hil-22和pre-hil-22序列,同时用引物il-22ost1forentryfw和il-22forentryrv扩增ost1-hil-22序列。然后通过bsmbi限制性酶切和t4连接将pcr片段引入pytk001/pptk081。
对人iga(homosapiens,uniprokb:p01877)和小鼠iga(musmusculus,uniprotkb:p01878)的fc区的序列进行密码子优化以用于在巴斯德毕赤酵母中表达,并在idt处订购合成为gblocks。然后用引物igaforentryfw和igaforentryrv或igamouseforentryfw和igamouseforentryrv扩增人和小鼠igagblock,以进行模块克隆。从先前产生的ev2a载体((virdiv等人(2013)pnas110,11809)用引物igapigforentryfwcorrect和igapigforentryrvnew扩增出猪iga的fc区(susscrofa,uniprotkb:k7zrk0)。将所有iga序列克隆到pytk001/pptk081中,作为moclo系统中的“3b部分”。
然后使用moclo系统将这些进入载体用于产生表达载体。每个表达载体是通过合并等摩尔量(约20飞摩尔)的以下部分(在addgene可得或将可得)生成的:
实施例2中使用的引物:
3.来自毕赤酵母产生的vhh-iga-fc融合蛋白的不同干燥过程的可食用制剂的体内功效比较
对于vhh-igafcs等生物治疗药物而言,为了在胃肠道中有效,口服和胃的稳定性至关重要。围绕抗体的基质(在分子水平)和干燥过程很可能在保护vhh-igafc抗体不在胃肠道中被消化和失效方面起重要作用。在这里,我们着手评估了不同干燥对毕赤酵母生产的vhh-igafcs在清除f4-etec感染方面的体内功效上的影响,其中将其与饲料基质一起冷冻干燥(参见实施例1)或在没有饲料基质的情况下冷冻干燥。此外,我们还评估了喷雾干燥过程,该过程基于涉及加热的替代原理;其中将麦芽糊精(matodextrin)用作基质/载体。毕赤酵母产生的抗f4-etecvhh-igafc(v2a和v3a)的三种不同工艺加工的饲料制剂,由相同的剂量构成;相当于每只仔猪每天0.5升发酵液或5毫克vhh-igafcs/天/仔猪。第四组在饲料中未接受vhh-igafc抗体,并作为阴性对照。选择24只f4-etec血清阴性和muc-13基因测试阳性的仔猪(这与f4-etec感染易感性相关),断奶后分成4组,每组6只。使这些仔猪适应固体食物,然后向它们提供组特异性实验饲料(见图4a)。提供实验饲料10天,在第三天所有仔猪用1010f4-etec细菌连续攻击两天(第0天和第1天)。通过分析粪便中每天排出的攻击菌株的菌落形成单位(cfu),直至第12天,来监测响应于饲料制剂所得到的感染效果(见图4b和表3)。
表3:每只仔猪每克粪便中f4-etec(log10)cfu的日排放量
注:cfu的log(10)检出限为2,破折号(-)表示未检测到细菌
注:仔猪2和仔猪9分别在第-5天和第2天死亡;原因是在死亡后调查中发现的大的胃溃疡。
阴性对照组的所有仔猪在第1天和第2天排出高cfu的f4-etec,并且至少有一半仔猪的排放保持到第3天。抗体接受组的总体排放很低。图4b显示了,直到第6天,每组的平均排放和平均值的标准误(sem),这反映了含有经差异处理的毕赤酵母生产的vhh-iga-fc融合物的三种膳食在预防f4-etec感染中的有效趋势。这些数据表明,基质(无论是冷冻干燥过程中的猪饲料,还是喷雾干燥中的麦芽糖糊精)可以引起更高的体内功效。血清抗etecigg(参见图4c)和iga(参见图4d)水平在接受饲料基质上冻干制剂的组中最低,这证实了排放的结果。总体而言,在显示血清转化的组中观察到各仔猪个体的变异(图4c和4d中误差线代表平均值的标准误)。
实施例3的材料和方法
基于毕赤酵母生产的vhh-iga-fc的饲料制剂
如实施例1攻击实验中所使用的,抗etecvhh-igafc的有效剂量为约5mgvhh-igafc,或更合适地,每只仔猪每天饲喂具有来自0.5l摇瓶生长培养物的干燥产物的饲料制剂。该剂量由两个抗f4-etecvhh-igafcs,即v2a和v3a,等分组成(见virdi等人(2013)110,29,11809-11814)。为准备类似剂量用于18只仔猪(三组,每组六只动物)接受毕赤酵母生产的vhh-igafcs;在摇瓶中生长分别表达v2a和v3a的毕赤酵母培养物各45l(总计90l)。如下表5所总结,进行了六批生产(在为期五日的工艺中,如实施例1,在bmgy培养基中生长48小时,然后在bmmy培养基中诱导48小时)。因此,每周生产15l培养物批次。每次运行结束时,离心收集培养基,将无细胞的上清液通过渗滤浓缩至1l,然后使用装有5kdaomegatmcentramate过滤器盒的centramatetm500s切向流过滤系统(palllifescience),用磷酸钠缓冲液(20mmna2hpo4、18.75mmnacl,ph6)进行缓冲液交换。将在六个批次的每批结束时获得的、含有毕赤酵母产生的vhh-igafc的1l保留物蛋白溶液(称作保留物)(表4),按以下三种特定方式进行干燥。
表4:每批毕赤酵母生产的vhh-iga-fc的干燥工艺,以配制相应的实验饲料
方式1:具有基质的冷冻干燥(类似于实施例1):将一升保留物(带有vkh-igafcv2a(批次1,表5)或v3a(批次2,表4)),与一公斤商业仔猪饲料(供应商:vanhuffel,9850poesele(nevele)belgium)用手持式桨叶混合以避免起泡,并使用冷冻干燥机(epsilon2-10dlsc-martin-christ,德国)将浆液冻干47小时。然后将分别包含vhh-iga-fcv2a和v3a的2个冷冻干燥批次(每个约1kg)的所得干燥粉末与猪饲料混合,以产生18kg最终的具有在基质上冷冻干燥的毕赤酵母生产的vhh-igafc的饲料(表4)。
方式2:无基质冷冻干燥:使用冷冻干燥机(epsilon2-10dlsc-martin-christ,德国)冻干一升保留物(带有vhh-igafcv2a(批次3)或v3a(批次4))47小时。然后将分别包含vhh-igafcv2a和v3a的2个冷冻干燥批次(每个约30g)的所得干燥粉末与猪饲料混合,以产生18kg最终的具有无基质冷冻干燥的毕赤酵母生产的vhh-igafc的饲料(表4)。
方式3:喷雾干燥:将一升带有vhh-igafcv3a的保留物(批次5,表4)和另一升带有vhh-igafcv2a的保留物(批次6,表5),与含有2.5kg麦芽糖糊精的23l磷酸钠缓冲液(20mmna2hpo4,18.75mmnacl,ph6)混合。使用工业搅拌器将全部25l液体充分混合5-7分钟,然后将其进料到喷雾干燥器中,参数设置为45℃预热饲料液体和进气温度170℃。在干燥过程中,平均出口空气温度为约80℃,并保持恒定的液体泵送速度。回收了约2.3kg的带有v2a和v3a的干燥粉末,将其与猪饲料混合在一起,从而得到18kg最终的具有毕赤酵母生产的喷雾干燥的vhh-igafc的饲料(表4)。
来自每个不同干燥工艺运行的干燥vhh-igafcs(见表5),即,在饲料上冻干(v2a批次1,v3a-批次2),无基质冷冻干燥(v2a批次3,v3a批次4),和使用麦芽糖糊精的喷雾干燥(v3a+v2a,批次5和批次6的合并),当在磷酸盐缓冲液中溶解并在elisa中进行评估后,观察到功能活性,与固定化的f4-faeg抗原结合。
对照饲料包括18kg基础饲料,没有任何抗体。将18kg饲料制剂分成10袋,每袋1.8kg,作为每组的每日饲料供给量,从第-3天到第7天在仔猪的饲料桶中提供(图4a)。
仔猪攻击实验:
经比利时根特大学兽医学院动物护理和伦理委员会批准(伦理档案号ec2017/122),根据比利时动物福利法规进行了仔猪攻击实验。仔猪(品种:杂种)购自比利时梅勒市农业和渔业研究所(ilvo)的农场(伦理档案号2017/306)。五头初产母猪回避了针对f4-etec的加强疫苗,以确保低乳源免疫。由这些母猪出生的仔猪在出生后每隔一天接受抗生素施用(duphamox0.1ml/只仔猪)三次,以抵抗f4-etec感染。此外,在出生的第15天从这些仔猪中取血样,以评估抗f4-etec血清滴度和muc13基因分型测定(goetstouwers等(2014)plosone9,e105013),这与f4-etec受体的存在相关。选择24个血清阴性且muc13f4-etec易感基因型纯合的仔猪,断奶后运输到根特大学兽医学院的圈中进行攻击实验。根据仔猪的所在窝、基因型和体重,将它们适当地随机分配到喂养组中。每组的平均起始重量为8.5kg。在第-12天和第-11天给仔猪施用肌内抗生素-duphamox(0.6ml/只仔猪);而在第-8天,-7天和-6天施用baytril(0.25毫升/只仔猪)以防止运输后细菌感染的偶然性。如先前所述进行攻击(virdi等人(2013)procnatacadsciusa.110,29,11809-11814)。简而言之,在用60ml碳酸氢盐缓冲液(在蒸馏水中1.4%nahco3w/v)中和胃ph后,在镇静下(1ml氮杂哌酮,
- 还没有人留言评论。精彩留言会获得点赞!