一种高温生物转化制备L-2-氨基丁酸的方法与流程
本发明属于属于生物化学
技术领域:
,尤其涉及一种高温生物转化制备l-2-氨基丁酸的方法。
背景技术:
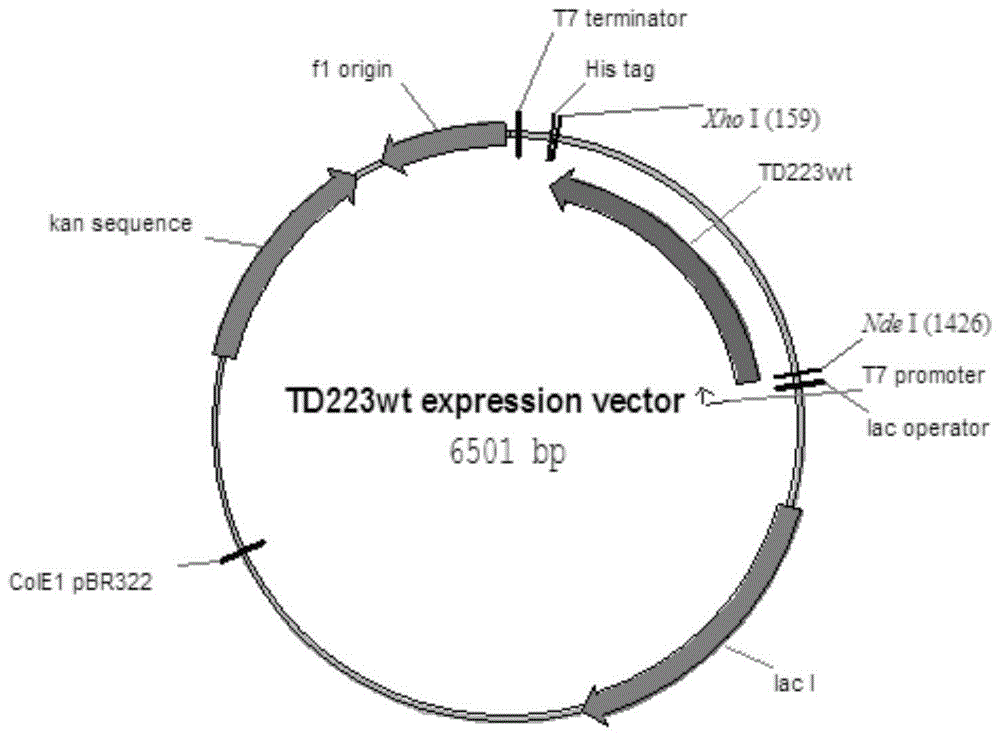
:l-2-氨基丁酸(l-aba)是一种非蛋白氨基酸,具有抑制人体神经信息传递,加强葡萄糖磷酸酯酶的活性和促进脑细胞代谢的作用。l-2-氨基丁酸的衍生物(s)-2-氨基丁酰胺可做为新型抗癫痫药物左乙拉西坦和布瓦西坦等手性药物的关键中间体,l-2-氨基丁酸的另一种衍生物(s)-2-氨基丁醇是抑菌抗结核药乙胺丁醇盐酸盐的关键中间体。这些药物的光学纯度对治疗的安全性和有效性至关重要。左乙拉西坦的r-对映体没有抗癫痫活性和(r,r)-形式的安非他命可引起失明。尽管乙胺丁醇和左旋乙拉西坦现在是仿制药,但在许多国家,高昂的药价造成大多数癫痫患者不能使用左乙拉西坦治疗。目前,随着左乙拉西坦等药物专利到期,市场普及程度提高,对其关键中间体l-2-氨基丁酸需求越来越大。l-2-氨基丁酸的生产方法主要有发酵法,化学合成法和生物催化法。发酵法现在存在的最大缺陷是产量低,产品纯度不够。目前的发酵法主要是针对苏氨酸代谢途径进行改造,使得代谢流向2-丁酮酸及-2-氨基丁酸,但报道出的数据均小于50克/升,结合l-2-氨基丁酸的市场价格现状(2018年四季度报价不足15美金)及市场容量(全球市场百吨级),目前的发酵效价不太具有放大生产的意义。如专利cn201510208576.1中发酵后期要补入100g/l苏氨酸,10g/l谷氨酸,反应实质上仍是全细胞生物催化反应,且产物最高记录仅为57g/l左右,转化率偏低。cn201210015308.4在苏氨酸高产菌株的基础上,通过过表达苏氨酸脱氨酶与亮基酸脱氢酶发酵生成产物,但该方法发酵时间需要60小时,且产物最高记录仅为20g/l。另外微生物发酵过程中会产生大量的代谢产物并且发酵培养基中也会带入大量的有机培养基及无机盐,对分离造成一定程度的影响,进而导致最终产品的纯度及手性纯度低于体外催化途径。如cn201610208600.6专利中,发酵结束后经陶瓷膜,超滤膜过滤后,再经离子交换树脂吸附洗脱,纯度仅为97.3%,仍低于98.5%的普通级l-2-氨基丁酸纯度要求且离子交换树脂的使用对于l-2-氨基丁酸目前的价格并不具有实用意义。化学法合成的产物多为消旋体,需要进一步拆分。另外,化学法需要使用昂贵的试剂,易生成副产物,同时使用多种有机溶剂,进一步增大了环保处理的成本。生物催化法主要有氨基酰化酶法,氨基酸氧化酶法,酰胺酶法,腈水解酶法,转氨酶法和氨基酸脱氢酶法等。其中又以转氨酶法和氨基酸脱氢酶法的催化底物浓度及生产效率较高,目前市场上的生物催化法主要以这两类为主。转氨酶法是指2-丁酮酸在转氨酶的催化作用下合成2-氨基丁酸。利用α转氨酶合成时,由于反应的可逆性仅获得58%~62%的产率,并且该过程复杂,需要引入乙酰乳酸合成酶、丙氨酸外消旋酶和d-氨基酸氧化酶移除副产物。l-2-氨基丁酸还可通过ω-转氨酶以苄胺或异丙胺作为氨基供体合成,该反应不可逆,但目前所报道的ω-转氨酶法普遍底物浓度偏低。专利cn201510900727.x以胺和酮类化合物为原料,通过立体选择性地转氨基作用,生产手性胺,但转化浓度偏低,最高的数据仅为46g/l,距离工业放大生产仍有不小差距。氨基酸脱氢酶为目前研究最深入的生物催化途径,专利cn201210066624.4和cn201010139227.6等报道了以l-苏氨酸为原料,先由苏氨酸脱氨酶转化成2-丁酮酸,再由亮氨酸脱氢酶催化2-丁酮酸制备l-2-氨基丁酸,但是cn201210066624.4催化1000g苏氨酸需要用到600g全细胞菌体,发酵成本上不具有生产意义。而cn201010139227.6的辅酶再生系统是葡萄糖,由于葡萄糖用量过大且葡萄糖酸与产物l-2-氨基丁酸的分子量相近,成熟简单的膜分离方法无法有效分离产物与副产物,增大了分离成本;该专利中酶参与的反应不超过30℃,反应温度低,反应速度慢,转化效率低。另外,现存报道中辅酶用量:底物用量几乎都大于1:1000,即实际生产中投入底物1吨至少需要消耗1公斤辅酶,而辅酶价格昂贵(如日本东方酵母进口95%规格nad价格约为8000元/公斤)间接的增加了l-2-氨基丁酸的生产成本。国内主要的l-2-氨基丁酸供应商采用的是综合性价比较高的苏氨酸脱氨酶,亮氨酸脱氢酶,偶联甲酸脱氢酶进行辅酶的循环使用,但过程中需要与苏氨酸等摩尔量的甲酸铵,且铵离子会一直存在于反应液中,提取结束后产生的大量废液中含有高浓度的铵盐,造成后期的环保成本巨大。专利cn201210066624.4公开了一种l-2-氨基丁酸的生物制备方法,该方法需在甲酸铵、辅酶磷酸吡哆醛的的存在下,在15℃-50℃条件下反应得到l-2-氨基丁酸。一方面该专利中催化1000g苏氨酸需要用到600g全细胞菌体,发酵成本上不具有生产意义;另一方面,引入了甲酸铵。加氨氮浓度质量大于500mg/l属于高浓度氨氮废水,由于nh4+中氮元素的氧化,会造成水体中溶解氧浓度降低,导致水质下降,对水生动植物的生存造成影响。水中氮素含量太多,致使光合微生物(大多数为藻类)的数量增加,即水体发生富营养化现象。因此当有nh4+存在时,水处理厂将需要更大的加氯量,从而增加处理成本。技术实现要素:本发明的目的解决现有技术中在制备l-2-氨基酸过程中需加入甲酸铵引入氨离子,增加环境负担及生产成本的问题,提供一种高效、高产、环保的高温生物转化制备l-2-氨基丁酸的方法。本发明的技术方案如下:一种高温生物转化制备l-2-氨基丁酸的方法,其特征在于,通过热稳定的苏氨酸脱氨酶能将l苏氨酸转化成2-丁酮酸,再通过亮氨酸脱氢酶及醇脱氢酶转化2-丁酮酸生成l-2-氨基丁酸,所述苏氨酸脱氨酶源于chryseobacteriumtakakiae(藻苔金黄杆菌)的野生型苏氨酸脱氨酶或其突变体,且所述突变体与seqidno.2具有90%以上相似性并具有以下特征中的一个或多个突变:h63n,i78v,e202s,r375k,所述苏氨酸脱氨酶突变体的序列为seqidno.5。优选的,所述苏氨酸脱氨酶或其突变体比大肠杆菌来源的苏氨酸脱氨酶具有更高的热稳定性。优选的,反应温度在45℃-60℃。优选的,辅酶循环依赖于醇脱氢酶且不添加甲酸铵。一种多核苷酸,其特征在于,所述多核苷酸的编码序列为seqidno.2或者seqidno.5的苏氨酸脱氨酶重组的多肽。优选的,所述多核苷酸的序列为seqidno.1或seqidno.4。一种重组质粒,其特征在于,包括表达载体连接序列为seqidno.1或seqidno.4的多核苷酸。一种宿主细胞,其特征在于,其包含如权利要求7的重组质粒。优选的,所述细胞是一种大肠杆菌。通过采用以上技术方案,本发明达到的技术效果如下:1、本发明公开的技术方案,利用热稳定的苏氨酸脱氨酶,避免使用甲酸铵引入铵离子,利用热稳定的醇脱氢酶循环辅酶再生,优势在于整个体系不引入无机盐,底物为各种醇类而副产物如丙酮等具有低沸点特性,可直接从产物中分离出去,对酶的活性没有任何影响。2、与目前发现的苏氨酸脱氨酶相比,本发明的重组苏氨酸脱氨酶突变体具有更好的热稳定性,本发明的苏氨酸脱氨酶突变体在45℃-60℃的活性更持久,具有更好的热稳定性,适用于较高温度下催化l苏氨酸生成2-丁酮酸再生成l-2-氨基丁酸,因此更适合于工业化应用。3、本发明公开的技术方案实现了l-2-氨基丁酸的高产、高效,底物浓度达到250-300g/l为首次公开报道,辅酶用量:底物用量比可达1:1500,辅酶循环次数高,反应条件温和且有效减轻环保压力。附图说明图1为td223wt的表达质粒图谱;图2为td223的表达质粒图谱;图3为td223wt的蛋白胶图谱;图4为td223的蛋白胶图谱;图5为实例9用于l-2-氨基丁酸高温生物催化制备的20小时薄层色谱结果。上方显色条带为产物l-2-氨基丁酸,下方显色条带为底物苏氨酸。自左至右依次为:反应样;底物苏氨酸对照;图6为实例10用于l-2-氨基丁酸高温生物催化制备的20小时薄层色谱结果。上方显色条带为产物l-2-氨基丁酸,下方显色条带为底物苏氨酸。自左至右依次为:反应样;底物苏氨酸对照;图7为实例12td233用于l-2-氨基丁酸高温生物催化制备的12小时hplc图谱。3.146分钟为产物l-2-氨基丁酸;图8为实例13td233wt用于l-2-氨基丁酸高温生物催化制备的16小时hplc图谱。2.691分钟为底物苏氨酸;3.167分钟为产物l-2-氨基丁酸;图9为底物、产物、中间体标准品液相图谱。2.671分钟为底物苏氨酸;3.151分钟为产物l-2-氨基丁酸;7.529分钟为中间体丁酮酸;图10为底物、产物标准品薄层色谱结果。上方显色条带为产物l-2-氨基丁酸,下方显色条带为底物苏氨酸。具体实施方式一种高温生物转化制备l-2-氨基丁酸的方法,其特征在于,通过热稳定的苏氨酸脱氨酶能将l苏氨酸转化成2-丁酮酸,再通过亮氨酸脱氢酶及醇脱氢酶转化2-丁酮酸生成l-2-氨基丁酸,所述苏氨酸脱氨酶源于chryseobacteriumtakakiae的野生型苏氨酸脱氨酶或其突变体,且所述突变体与seqidno.2具有90%以上相似性并具有以下特征中的一个或多个突变:h63n,i78v,e202s,r375k,所述苏氨酸脱氨酶突变体的序列为seqidno.5。全长突变的苏氨酸脱氨酶对于保持酶的催化活性并不是必需的。相应地,应考虑苏氨酸脱氨酶突变体的截短的类似物和有催化活性的片段。例如,在一些实施方案中,c端或n端的数个氨基酸可以被删去。任何特定的截短的类似物或片段可以利用相应的试验来评估催化活性。同样的,额外的氨基酸残基可以被添加到一个或两个末端而不影响催化活性。额外序列可以是功能性的或非功能性的。例如,额外氨基酸序列可以被用来帮助纯化,做为标记,或执行一些其它的功能。因此,本公开内容的苏氨酸脱氨酶突变体可以是融合蛋白的形式,其中苏氨酸脱氨酶突变体(或其片段)诸如通过助溶标签(如sumo蛋白)、纯化标签(如结合金属的his标签)和细菌定位信号(如分泌信号)的实例而非限制的方式被融合到其它蛋白质。优选的,所述苏氨酸脱氨酶或其突变体比大肠杆菌来源的苏氨酸脱氨酶具有更高的热稳定性。优选的,反应温度在45℃-60℃。优选的,辅酶循环依赖于醇脱氢酶且不添加甲酸铵。一种多核苷酸,所述多核苷酸的编码序列为seqidno.2或者seqidno.5的苏氨酸脱氨酶重组的多肽。优选的,所述多核苷酸的序列为seqidno.1或seqidno.4。一种重组质粒,其特征在于,包括表达载体连接序列为seqidno.1或seqidno.4的多核苷酸。一种宿主细胞,其包含如前面所述重组质粒。优选的,所述宿主细胞是一种大肠杆菌。本发明提供一种苏氨酸脱氨酶突变体,其源于chryseobacteriumtakakiae的野生型苏氨酸脱氨酶,能够催化l苏氨酸生成2-丁酮酸再生成l-2-氨基丁酸。所述苏氨酸脱氨酶突变体,与seqidno.2的野生型苏氨酸脱氨酶相比表现出更强的热稳定性。苏氨酸脱氨酶突变体和编码这种突变体的多核苷酸可以使用本领域技术人员通常使用的方法制备。突变体可以通过使编码该酶的体外重组、多核苷酸诱变、dna改组、易错pcr和定向进化方法等获得。为了更好的解释本发明,下面结合实施例对本发明进行进一步的阐述。在本实施例中所用到的仪器、试剂,除非有特殊说明,均为市售产品。实施例1野生型苏氨酸脱氨酶基因序列的获得通过全基因合成的方法,对基因的二级结构以及密码子偏好性进行调整,以实现在大肠杆菌中的高表达。利用primerpremier(http://primer3.ut.ee/)和optimizer(http://genomes.urv.es/optimizer/)进行设计,并保证退火温度(tm)差异控制在3℃以内,引物长度控制在60base以内,并将获得的引物加双蒸水溶解后,加到如下的反应体系中,使得各引物的终浓度为30nm,首尾引物的终浓度为0.6μm。2mmdntpmix(2mmeachdntp)5μl10×pfubuffer5μlpfudnapolymerase(10u/μl)0.5μlddh2o使得反应体系总体积至50μl将配制好的pcr反应体系置于博日xpcycler基因扩增仪中,按下列程序进行扩增:98℃30s,55℃45s,72℃120s,35x。将pcr得到的dna片段进行切胶纯化,利用同源重组的方法克隆进pet30a的ndei/xhoi位点。挑取单克隆进行测序。测序成功的dna序列为seqidno.1,命名为td223wt,其表达质粒图谱和蛋白胶图谱分别图如图1和图3所示,其对应的氨基酸序列为seqidno.2。实施例2苏氨酸脱氨酶突变体基因序列的获得通过全基因合成的方法,对基因的二级结构以及密码子偏好性进行调整,以实现在大肠杆菌中的高表达。利用primerpremier(http://primer3.ut.ee/)和optimizer(http://genomes.urv.es/optimizer/)进行设计,并保证退火温度(tm)差异控制在3℃以内,引物长度控制在60base以内,并将获得的引物加双蒸水溶解后,加到如下的反应体系中,使得各引物的终浓度为30nm,首尾引物的终浓度为0.6μm。2mmdntpmix(2mmeachdntp)5μl10×pfubuffer5μlpfudnapolymerase(10u/μl)0.5μlddh2o使得反应体系总体积至50μl将配制好的pcr反应体系置于博日xpcycler基因扩增仪中,按下列程序进行扩增:98℃30s,55℃45s,72℃120s,35x。将pcr得到的dna片段进行切胶纯化,利用同源重组的方法克隆进pet30a的ndei/xhoi位点。挑取单克隆进行测序。测序成功的dna序列为seqidno.4,命名为td223,其表达质粒图谱和蛋白胶图谱分别如图2和图4所示,其对应的氨基酸序列为seqidno.5。实施例3大肠杆菌来源苏氨酸脱氨酶基因序列的获得根据qjz14922所示源自escherichiacoli的野生型蛋白序列,委托苏州金唯智生物对该蛋白的编码序列进行全基因合成,并克隆至pet30a中,得到对照蛋白表达质粒ec07,其对应的氨基酸序列为seqidno.3。实施例4摇瓶表达测试挑取含有表达载体的大肠杆菌单菌落接种于10ml高压灭菌后的培养基中:胰蛋白胨10g/l,酵母提取物5g/l,磷酸氢二钠3.55g/l,磷酸二氢钾3.4g/l,氯化铵2.68g/l,硫酸钠0.71g/l,七水硫酸镁0.493g/l,六水氯化铁0.027g/l,甘油5g/l,葡萄糖0.8g/l,添加卡那霉素至50mg/l。30℃,250rpm过夜培养。次日取1l三角瓶,按1:100的接种比例接入到100ml高压灭菌后的培养基中:胰蛋白胨10g/l,酵母提取物5g/l,磷酸氢二钠3.55g/l,磷酸二氢钾3.4g/l,氯化铵2.68g/l,硫酸钠0.71g/l,七水硫酸镁0.493g/l,六水氯化铁0.027g/l,甘油5g/l,葡萄糖0.3g/l,添加卡那霉素至50mg/l。于30℃中培养至菌体od5-6,立刻将三角瓶置于25℃摇床中,250rpm培养1小时。加iptg至终浓度0.1mm,并于25℃,250rpm继续培养16小时。培养结束后,将培养液于4℃,12000g下离心20分钟收集湿菌体。然后将菌体沉淀用蒸馏水清洗两次,收集菌体,-70℃保存。同时取少量菌体进行sds-page检测。实施例5分批补料发酵分批补料发酵在计算机控制的生物反应器(上海国强)中进行,反应器容量为15l,工作体积为8l,所用到的培养基为酵母抽提物24g/l,蛋白胨12g/l,葡萄糖0.4%,2.31g/l磷酸二氢酶和12.54g/l磷酸氢二钾,ph7.0。初级接种菌种制备200ml培养物,od2.0时接入。在整个发酵过程中,温度保持在37℃,发酵过程中溶解氧浓度由搅拌速率(rpm)和通气供应级联控制在30%自动控制,而培养基的ph值由50%(v/v)正磷酸和30%(v/v)氨水维持在7.0。发酵过程中,当出现大幅的溶氧回升时,开始补料。补料溶液含有9%w/v蛋白胨、9%w/v酵母提取物、14%w/v甘油。当od600约为50.0(湿重约为100g/l)时,控制温度为28℃,并用0.15mmiptg诱导表达。实施例6热稳定实验将实施4中制备的苏氨酸脱氨酶加入45℃,55℃水浴锅中温育120分钟,取出后置于4℃保存以测试酶活残余,并以未处理的样品酶活做为100%。同时以大肠杆菌来源的野生型苏氨酸脱氨酶相同条件处理后做为对照。实施例7酶活检测取6个5ml离心管,分别标号1-6,分别加入稀释好的丁酮酸溶液0μl,40μl,80μl,100μl,120μl,160μl,然后每管补加0.1m,ph7.0的磷酸盐缓冲液补足3ml,混匀后在230nm下检测并记录吸光度的值;根据上述测得值,得到丁酮酸的标准曲线y=k*x,其中y为吸光度的值,x为丁酮酸的浓度(umol),曲线的r2>99.5%;用纯水按一定的倍数稀释酶液(参考稀释倍数:40-100倍),稀释的倍数使每分钟吸光值变化0.02-0.04之间为合适;取5ml离心管,按以下比例取样加入离心管,迅速混合,立即倒入比色皿。检测试剂用量40mm苏氨酸2.95ml稀释后的酶液50μl在230nm处检测吸光度的变化,每隔1min记录一个值,每分钟的变化率基本相同,其中0min的吸光度为s0,3min的吸光度为s3酶活定义:一个酶活力单位定义为在测定条件下,每分钟产生1umol,2-丁酮酸所需要的酶量。酶活检测结果如下:实施例8.生物转化反应在100ml三角摇瓶中加入5ml纯水,2.2g饲料级苏氨酸(含量大于98%),1.6ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.9,再加入终浓度0.2mmnad粗品(10%含量,购自南京朗恩生物科技有限公司),1mgplp,混匀。最后加入120μltd223,100μl亮氨酸脱氢酶ld161(购自南京朗恩生物科技有限公司),100μl醇脱氢酶ad151(购自南京朗恩生物科技有限公司),置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度大于220g/l。反应起始会有大量的不溶物在瓶底,但在3小时已无明显底物沉淀,整个反应变得澄清,显示出所用苏氨酸脱氨酶在前3小时反应速度非常快;此时取出反应液冷却后,会析出较多的晶体,此为产物l-2-氨基丁酸在低温下溶解度下降结晶所致。实施例9.生物转化反应在100ml三角摇瓶中加入7.0ml纯水,2.5g饲料级苏氨酸(含量大于98%),2.0ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.8,再加入终浓度0.25mmnad粗品,0.166mgplp(取13μl1%浓度的plp母液),混匀。最后加入63μltd223,84μl亮氨酸脱氢酶ld161,84μl醇脱氢酶ad151,置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为250g/l。如图10所示,图10为底物、产物标准品薄层色谱结果,上方显色条带为产物l-2-氨基丁酸,下方显色条带为底物苏氨酸。如图5所示,通过与图10对比可知,上方显色条带为产物l-2-氨基丁酸,下方显色条带为底物苏氨酸。自左至右依次为:反应样;底物苏氨酸对照;20小时点板已无底物剩余。实施例10.生物转化反应在100ml三角摇瓶中加入7.0ml纯水,3g饲料级苏氨酸(含量大于98%),2.4ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.8,再加入终浓度0.3mmnad粗品,0.2mgplp(取20μl1%浓度的plp母液即可),混匀。最后加入100μltd223,100μl亮氨酸脱氢酶ld161,100μl醇脱氢酶ad151,置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为300g/l,此时会有大量的底物沉淀在瓶底不溶。反应2小时已无明显沉淀,反应体系呈澄清透明状态,5.5小时可见摇瓶底部出现大量糊状沉淀,此为生成的l-2-氨基丁酸浓度超过溶解度而析出。如图6所示,通过与图10对比可知,上方显色条带为产物l-2-氨基丁酸,下方显色条带为底物苏氨酸。自左至右依次为:反应样;底物苏氨酸对照;20小时点板已无底物剩余。实施例11.生物转化反应在100ml三角摇瓶中加入7.9ml纯水,1.2g饲料级苏氨酸(含量大于98%),0.5ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.0,再加入终浓度0.2mmnad粗品,0.05mgplp,混匀。最后加入40μltd223,33μl亮氨酸脱氢酶ld161,33μl醇脱氢酶ad151,置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为120g/l。反应3小时后,补入0.3ml异丙醇。10小时取样液相检测,l-2-氨基丁酸浓度约97g/l。实施例12.生物转化反应在100ml三角摇瓶中加入7.2ml纯水,1.5g饲料级苏氨酸(含量大于98%),1.2ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.9,再加入终浓度0.15mmnad粗品,0.75mgplp,混匀。最后加入90μltd223,75μl亮氨酸脱氢酶ld161,75μl醇脱氢酶ad151,置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为150g/l。分别反应6小时,12小时后取样检测,结果显示l-2-氨基丁酸浓度分别为109g/l,123g/l。图7为本实施例用于l-2-氨基丁酸高温生物催化制备的12小时hplc图谱,图9为底物、产物、中间体标准品液相图谱。由图9可知,2.671分钟为底物苏氨酸;3.151分钟为产物l-2-氨基丁酸;7.529分钟为中间体丁酮酸。对比图9,由图7可知,3.146分钟为产物l-2-氨基丁酸;未明显检出底物苏氨酸和中间体丁酮酸的峰;反应12小时后,底物、中间体基本转化为l-2-氨基丁酸。实施例13.生物转化反应在100ml三角摇瓶中加入7.2ml纯水,1.5g饲料级苏氨酸(含量大于98%),1.2ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.9,再加入终浓度0.15mmnad粗品,0.75mgplp,混匀。最后加入90μltd223wt,75μl亮氨酸脱氢酶ld161,75μl醇脱氢酶ad151,置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为150g/l。分别反应6小时,12小时,16小时后取样检测,结果显示l-2-氨基丁酸浓度分别为98g/l,112g/l,122g/l。对比图9,由图8可知,3.167分钟为产物l-2-氨基丁酸;未明显检出底物苏氨酸和中间体丁酮酸的峰,底物基本完全转化为l-2-氨基丁酸。由实施例12和实施例13可知,相同的条件下,本实施公开的苏氨酸脱氨酶突变体比来源于chryseobacteriumtakakiae的野生型苏氨酸脱氨酶的具有更高的酶催化转化效率。实施例14.生物转化反应在100ml三角摇瓶中加入7.2ml纯水,1.5g饲料级苏氨酸(含量大于98%),1.2ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.9,再加入终浓度0.15mmnad粗品,0.75mgplp,混匀。最后加入90μl大肠杆菌来源苏氨酸脱氨酶,75μl亮氨酸脱氢酶ld161,75μl醇脱氢酶ad151,置于50℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为150g/l。分别反应6小时,12小时后取样检测,结果显示l-2-氨基丁酸浓度分别为6g/l,6g/l。说明催化第一步反应的苏氨酸脱氢酶在高温条件下迅速失活,从而中止了后续的催化反应。同时证明了大肠杆菌来源的苏氨酸脱氨酶无法有效应用于高温催化反应。实施例15.放大反应体系生产在20立方反应罐中加入14吨自来水,2.7吨饲料级苏氨酸(含量大于98%),2.15立方异丙醇,用25%氨水(w/w)调节ph至8.9,再加入终浓度1.78公斤nad辅酶,0.3公斤磷酸吡哆醛,混匀后控制温度至48℃-52℃。最后加入160公斤td223,135公斤亮氨酸脱氢酶ld161,135公斤醇脱氢酶ad151,最后加入少量自来水定容至18立方,搅拌速度设为30转/分钟。总体系约18立方,底物浓度约为150g/l。反应10小时后取样检测,结果显示l-2-氨基丁酸浓度达到118g/l。可见由于高温反应,即使辅酶用量:底物用量比仅为1:1500,仍可有效生成产物。实施例16.生物转化反应取2个100ml三角摇瓶中加入7.2ml纯水,1.5g饲料级苏氨酸(含量大于98%),1.2ml工业异丙醇,取少量25%氨水(w/w)调节ph至8.9,再加入终浓度0.15mmnad粗品,0.75mgplp,混匀。最后加入90μltd223,75μl亮氨酸脱氢酶ld161,75μl醇脱氢酶ad151,分别置于45℃,60℃摇床速度设为180rpm反应。总体系约10ml,底物浓度约为150g/l。分别反应9小时后取样检测,结果显示l-2-氨基丁酸浓度分别为121g/l,119g/l。实施例16.薄层色谱点板取反应液100μl,加纯水稀释一定倍数。取1μl上清tlc点板,展开剂正丁醇:乙酸:水=4:1:1。跑板达到4/5取出吹干,茚三酮显色。结果如图10所示,下边条带是底物,上边条带是产物。实施例17.hplc检测条件色谱柱:c185μm250mm×4.6mm;流动相:4%的甲醇及96%的0.1%h3po4(ph3.0);流速:0.7ml/min;检测波长:210nm;柱温:30℃。以上所述,仅为本发明进一步的实施例,但本发明的保护范围并不局限于此,任何熟悉本
技术领域:
的技术人员在本发明所公开的范围内,根据本发明的技术方案及其构思加以等同替换或改变,都属于本发明的保护范围。sequencelisting<110>南京诺云生物科技有限公司<120>一种高温生物转化制备l-2-氨基丁酸的方法<130>2020<160>5<170>patentinversion3.3<210>1<211>1263<212>dna<213>人工序列<400>1atgaacaacaccctgaccttcccgaccctggaatctatcatccaggctggtaaatctatc60gaaaacgttgttaactacaccccgctgcagtacaacgctcgtctgtctgaaaaattcggt120gctcacatctacctgaaacgtgaagacctgcagccggttcgttcttacaaactgcgtggt180gcttaccacaaaatcaaatctctgttcaacgaaggtaaaacctctgaaggtatcgtttgc240gcttctgctggtaaccacgctcagggtgttgctttctcttgcaaacagctgcagatcaaa300ggtaccatcttcatgccggttaccaccccgaaacagaaactggaacaggttgaaatgttc360ggtggtcacttcgttgaaatcaaactgttcggtgacaccttcgacgcttctaaaaacgct420gctctggacttcgctgaaaccttcggtgctgctttcatccacccgttcgacgacgttcag480atcatcgaaggtcaggctaccgttgctctggaaatcctgggtcagcagaaagaagctatg540gacttcgttttcatcccgatcggtggtggtggtctggcttctggtatctctaccgttttc600aaagaactgtctgctgaaacccgtctgatcggtgttgaaccgaaaggtgctccgtctatg660aaaatctctatcgaaaacaaaatcaacaccgaactgccggaaatcgaccgtttcgttgac720ggtgctgctgttaaaaaagttggtgacctgaccttcgaaatctgccgtaacaccctgtct780gaatgcatctctgttgacgaaggtaaaatctgcaacaccatcctgcagctgtacaacaaa840gacgctgttgttctggaaccggctggtgctctgtctatctctgctctggaccagttccgt900aaccgtatcaaaggtaaaaacgttgtttgcatcgtttctggttctaacaacgacatcacc960cgtatggaagaaatcaaagaacgtgctctgctgtacaacggtctgaaacactacttcatg1020gttaaattcccgcagcgtccgggtgctctgaaagacttcgttctgaacgttctgggtgtt1080aacgacgacatcacccacttcgaatacaccaaaaaaaactctcgtgaaaccgctctggct1140atcgttggtatcgaactgtctgacccgtctgacttcgaaggtctgcgtcagcgtatgcag1200gctctggactacctggaatcttacctgaacgaaaacccggacgttctgaacatgctggtt1260taa1263<210>2<211>420<212>prt<213>chryseobacteriumtakakiae<400>2metasnasnthrleuthrpheprothrleugluserileileglnala151015glylysserilegluasnvalvalasntyrthrproleuglntyrasn202530alaargleuserglulyspheglyalahisiletyrleulysargglu354045aspleuglnprovalargsertyrlysleuargglyalatyrhislys505560ilelysserleupheasngluglylysthrsergluglyilevalcys65707580alaseralaglyasnhisalaglnglyvalalaphesercyslysgln859095leuglnilelysglythrilephemetprovalthrthrprolysgln100105110lysleugluglnvalglumetpheglyglyhisphevalgluilelys115120125leupheglyaspthrpheaspalaserlysasnalaalaleuaspphe130135140alagluthrpheglyalaalapheilehispropheaspaspvalgln145150155160ileilegluglyglnalathrvalalaleugluileleuglyglngln165170175lysglualametaspphevalpheileproileglyglyglyglyleu180185190alaserglyileserthrvalphelysgluleuseralagluthrarg195200205leuileglyvalgluprolysglyalaprosermetlysileserile210215220gluasnlysileasnthrgluleuprogluileaspargphevalasp225230235240glyalaalavallyslysvalglyaspleuthrphegluilecysarg245250255asnthrleuserglucysileservalaspgluglylysilecysasn260265270thrileleuglnleutyrasnlysaspalavalvalleugluproala275280285glyalaleuserileseralaleuaspglnpheargasnargilelys290295300glylysasnvalvalcysilevalserglyserasnasnaspilethr305310315320argmetglugluilelysgluargalaleuleutyrasnglyleulys325330335histyrphemetvallyspheproglnargproglyalaleulysasp340345350phevalleuasnvalleuglyvalasnaspaspilethrhispheglu355360365tyrthrlyslysasnserarggluthralaleualailevalglyile370375380gluleuseraspproseraspphegluglyleuargglnargmetgln385390395400alaleuasptyrleuglusertyrleuasngluasnproaspvalleu405410415asnmetleuval420<210>3<211>514<212>prt<213>大肠杆菌<400>3metalaaspserglnproleuserglythrprogluglyalaglutyr151015leuargalavalleuargalaprovaltyrglualaalaglnvalthr202530proleuglnlysmetglulysleuserserargleuaspasnvalile354045leuvallysarggluaspargglnprovalhisserphelysleuarg505560glyalatyralametmetalaglyleuthrglugluglnlysalahis65707580glyvalilethralaseralaglyasnhisalaglnglyvalalaphe859095serseralaargleuglyvallysalaleuilevalmetprothrala100105110thralaaspilelysvalaspalavalargglypheglyglygluval115120125leuleuhisglyalaasnpheaspglualalysalalysalaileglu130135140leuserglnglnglnglyphethrtrpvalpropropheasphispro145150155160metvalilealaglyglnglythrleualaleugluleuleuglngln165170175aspalahisleuaspargvalphevalprovalglyglyglyglyleu180185190alaalaglyvalalavalleuilelysglnleumetproglnilelys195200205valilealavalglualagluaspseralacysleulysalaalaleu210215220aspalaglyhisprovalaspleuproargvalglyleuphealaglu225230235240glyvalalavallysargileglyaspgluthrpheargleucysgln245250255glutyrleuaspaspileilethrvalaspseraspalailecysala260265270alametlysaspleuphegluaspvalargalavalalagluproser275280285glyalaleualaleualaglymetlyslystyrilealaleuhisasn290295300ileargglygluargleualahisileleuserglyalaasnvalasn305310315320phehisglyleuargtyrvalsergluargcysgluleuglyglugln325330335argglualaleuleualavalthrileprogluglulysglyserphe340345350leulysphecysglnleuleuglyglyargservalthrglupheasn355360365tyrargphealaaspalalysasnalacysilephevalglyvalarg370375380leuserargglyleuglugluarglysgluileleuglnmetleuasn385390395400aspglyglytyrservalvalaspleuseraspaspglumetalalys405410415leuhisvalargtyrmetvalglyglyargproserhisproleugln420425430gluargleutyrserpheglupheprogluserproglyalaleuleu435440445argpheleuasnthrleuglythrtyrtrpasnileserleuphehis450455460tyrargserhisglythrasptyrglyargvalleualaalapheglu465470475480leuglyasphisgluproaspphegluthrargleuasngluleugly485490495tyraspcyshisaspgluthrasnasnproalapheargphepheleu500505510alagly<210>4<211>1263<212>dna<213>人工序列<400>4atgaacaacaccctgaccttcccgaccctggaatctatcatccaggctggtaaatctatc60gaaaacgttgttaactacaccccgctgcagtacaacgctcgtctgtctgaaaaattcggt120gctcacatctacctgaaacgtgaagacctgcagccggttcgttcttacaaactgcgtggt180gcttacaacaaaatcaaatctctgttcaacgaaggtaaaacctctgaaggtgttgtttgc240gcttctgctggtaaccacgctcagggtgttgctttctcttgcaaacagctgcagatcaaa300ggtaccatcttcatgccggttaccaccccgaaacagaaactggaacaggttgaaatgttc360ggtggtcacttcgttgaaatcaaactgttcggtgacaccttcgacgcttctaaaaacgct420gctctggacttcgctgaaaccttcggtgctgctttcatccacccgttcgacgacgttcag480atcatcgaaggtcaggctaccgttgctctggaaatcctgggtcagcagaaagaagctatg540gacttcgttttcatcccgatcggtggtggtggtctggcttctggtatctctaccgttttc600aaatctctgtctgctgaaacccgtctgatcggtgttgaaccgaaaggtgctccgtctatg660aaaatctctatcgaaaacaaaatcaacaccgaactgccggaaatcgaccgtttcgttgac720ggtgctgctgttaaaaaagttggtgacctgaccttcgaaatctgccgtaacaccctgtct780gaatgcatctctgttgacgaaggtaaaatctgcaacaccatcctgcagctgtacaacaaa840gacgctgttgttctggaaccggctggtgctctgtctatctctgctctggaccagttccgt900aaccgtatcaaaggtaaaaacgttgtttgcatcgtttctggttctaacaacgacatcacc960cgtatggaagaaatcaaagaacgtgctctgctgtacaacggtctgaaacactacttcatg1020gttaaattcccgcagcgtccgggtgctctgaaagacttcgttctgaacgttctgggtgtt1080aacgacgacatcacccacttcgaatacaccaaaaaaaactctaaagaaaccgctctggct1140atcgttggtatcgaactgtctgacccgtctgacttcgaaggtctgcgtcagcgtatgcag1200gctctggactacctggaatcttacctgaacgaaaacccggacgttctgaacatgctggtt1260taa1263<210>5<211>420<212>prt<213>人工序列<400>5metasnasnthrleuthrpheprothrleugluserileileglnala151015glylysserilegluasnvalvalasntyrthrproleuglntyrasn202530alaargleuserglulyspheglyalahisiletyrleulysargglu354045aspleuglnprovalargsertyrlysleuargglyalatyrasnlys505560ilelysserleupheasngluglylysthrsergluglyvalvalcys65707580alaseralaglyasnhisalaglnglyvalalaphesercyslysgln859095leuglnilelysglythrilephemetprovalthrthrprolysgln100105110lysleugluglnvalglumetpheglyglyhisphevalgluilelys115120125leupheglyaspthrpheaspalaserlysasnalaalaleuaspphe130135140alagluthrpheglyalaalapheilehispropheaspaspvalgln145150155160ileilegluglyglnalathrvalalaleugluileleuglyglngln165170175lysglualametaspphevalpheileproileglyglyglyglyleu180185190alaserglyileserthrvalphelysserleuseralagluthrarg195200205leuileglyvalgluprolysglyalaprosermetlysileserile210215220gluasnlysileasnthrgluleuprogluileaspargphevalasp225230235240glyalaalavallyslysvalglyaspleuthrphegluilecysarg245250255asnthrleuserglucysileservalaspgluglylysilecysasn260265270thrileleuglnleutyrasnlysaspalavalvalleugluproala275280285glyalaleuserileseralaleuaspglnpheargasnargilelys290295300glylysasnvalvalcysilevalserglyserasnasnaspilethr305310315320argmetglugluilelysgluargalaleuleutyrasnglyleulys325330335histyrphemetvallyspheproglnargproglyalaleulysasp340345350phevalleuasnvalleuglyvalasnaspaspilethrhispheglu355360365tyrthrlyslysasnserlysgluthralaleualailevalglyile370375380gluleuseraspproseraspphegluglyleuargglnargmetgln385390395400alaleuasptyrleuglusertyrleuasngluasnproaspvalleu405410415asnmetleuval420当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1