一种基于能量变化的植物组织抗脱水能力的检测方法与流程
本发明属于抗旱育种和农作物信息检测技术领域,具体涉及一种基于能量变化的植物组织抗脱水能力的检测方法。
背景技术:
干旱是指长期降水偏少,造成空气干燥,土壤缺水,使植物体内的水分发生亏缺,影响正常生长发育而减产,干旱造成的减产是干旱和半干旱地区粮食生产的主要障碍之一。抗旱性鉴定就是按植物抗旱能力的大小进行鉴定评价,由于植物在漫长的进化过程中以多种方式来抵抗和适应干旱,所以形成了多种抗旱机制。
其中植物组织的抗脱水能力在植物对干旱环境的适应方面发挥着重要作用。植物组织的抗脱水能力可通过水势变化的快慢进行比较,然而,叶片水势受环境变化容易快速波动;植物叶片的气孔导度、蒸腾速率、叶片含水率、茎直径的变化等即时指标同样经常被作为间接获得植物组织抗脱水能力的检测方法,其测量或者耗费时间,或者缺乏精确度。红外热成像分析同样受环境因子影响较大。植物电生理已广泛用于植物生长发育及水分状况的诊断,其测定易受夹持装置与叶片接触面积及夹持力的影响,造成测量误差。
因此,亟需建立一种既不受环境影响,又可以快速检测且具有较高精确度的测量方法,定量比较不同植物组织的抗脱水能力。
技术实现要素:
本发明要解决的技术问题是,提供一种基于能量变化的植物组织抗脱水能力的检测方法,可以通过力学指标准确检测植物组织的能量变化情况,进而反映其水分状况,简便快捷,为植物抗旱性检测技术提供科学支撑。
一种基于能量变化的植物组织抗脱水能力的检测方法,包括以下步骤:
步骤一,选取待测植物长势良好且一致的组织若干,设置不同形变梯度,测量各形变下植物组织的最大承压力;
步骤二,构建形变量与最大承压力之间的线性模型,得到植物组织的弹性模量;
步骤三,对所选取待测植物组织进行干燥失水处理;
步骤四,设置固定形变,测量植物组织在不同失水时刻的内力变化曲线,得到探头下压距离;
步骤五,计算探头下压过程中累积对植物组织所做的功,即为能量;
步骤六,计算不同失水时刻间能量的变异系数;
步骤七,定量比较不同植物组织的抗脱水能力。
进一步,所述步骤一中不同形变梯度以及步骤四中固定形变的设置均须以植物组织未受不可逆损坏为前提。
进一步,所述步骤二中的线性模型为胡克定律公式f=kx,其中f为压力,x为形变,k为弹性模量。
进一步,所述步骤四中的探头下压距离为:测量组织内力变化曲线的施压探头与植物组织接触开始到下压至所设定的固定形变的位置为止,其间所运行的距离。
进一步,所述步骤五中,功的计算公式为:w=fs,其中w为功,f为压力,s为探头下压距离。
进一步,所述步骤五中,探头下压过程中,压力随着探头下压距离的增加而增加,其变化符合胡克定律,可采用定积分法计算探头累积对植物组织所做的功,即
进一步,所述步骤六中,能量的变异系数的计算公式为:
进一步,所述步骤七中,不同植物组织的抗脱水能力通过步骤六中所计算的变异系数进行定量比较。
本发明的优点如下:
1)本方法基于组织力学指标计算组织抗脱水能力,结果不受测定仪器周围环境因子的限制,结果精确、可靠性高。
2)本方法基于组织弹性模量的测定,计算组织被施压过程中压力所做的功,根据能量守恒原理,来分析植物组织的水分变化情况,能够更好地将水分引起的组织能量的变化与植物的抗脱水能力联系起来,更好地反映植物对干旱环境的适应性。
3)本方法基于力学特性计算获取植物组织的水分变化状况,将物理学与植物生理学有机结合,植物生理指标的测试过程更为简捷。
附图说明
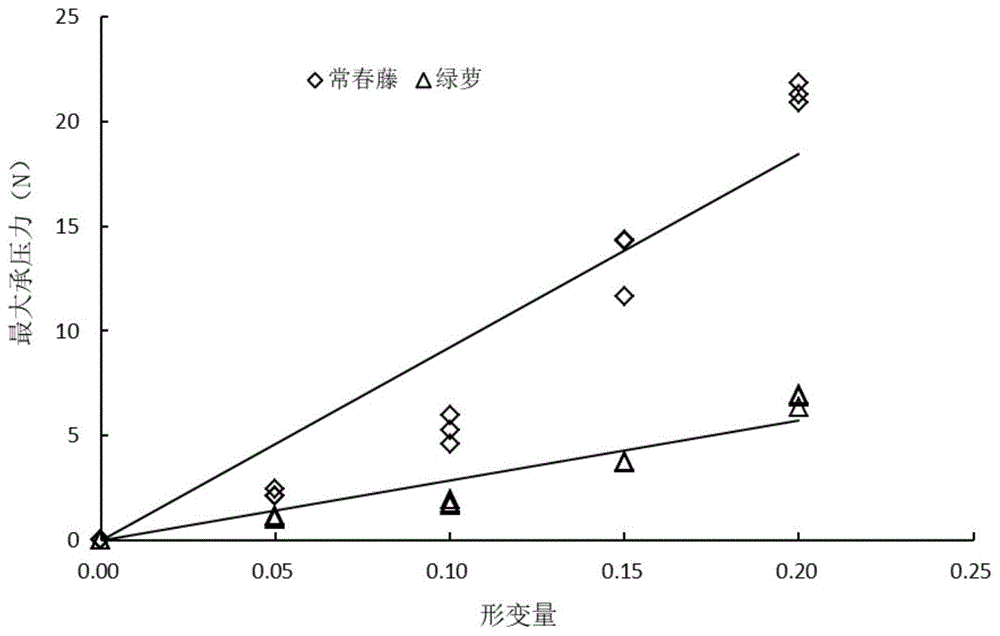
图1为利用胡克定律公式拟合的形变量与最大承压力之间的关系曲线图;
图2为0失水时刻常春藤茎的内力变化曲线图。
具体实施方式
下面将结合附图对本发明的具体方案做进一步的说明。
本发明的基本原理为:
植物经过长期的进化,从外部形态、解剖构造以及从内部生理生化特性等方面都形成了在高水势下的延迟脱水和低水势下的忍耐脱水能力。细胞壁机械强度越高,维持组织膨压的能力就越强,通过力学指标测定水分引起的植物组织的能量变化情况,能够反映细胞壁的抗破裂能力,表征植物抗脱水能力。
植物组织由细胞构成,构成细胞的细胞壁是一个弹性膜。因此,植物组织细胞整体上可被视为弹性材料,受力过程中,在植物组织遭受破坏前,外部压力与组织形变之间符合胡克定律公式。
胡克定律公式为:
f=kx(1)
其中f为压力,x为形变,k为弹性模量。
功的计算公式为:
w=fs(2)
其中w为功,f为压力,s为测量组织内力变化曲线的施压探头与植物组织接触开始到下压至所设定的固定形变的位置为止,其间所运行的距离。
探头下压过程中,压力随着下压距离的增加而实时增加,其变化符合胡克定律,可通过方程f=ks计算。
设置固定形变情况下,对植物组织施压,探头下压过程中对植物组织累积所做的功可采用定积分法进行计算,即:
其中b为探头与植物组织接触开始到下压至所设定的固定形变的位置后,所运行的最大距离。
植物组织失水过程中,会引发其内部生理及能量的变化。植物组织的机械强度越高,细胞壁的抗破裂能力越强,其抗脱水能力就越强。对组织施压过程中,同等形变下,压力对组织所做的功就越多。因此,同等形变下,不同失水时刻压力对植物组织所做的功即能量消耗之间的变化越剧烈,反映其水分变化越显著,组织抗脱水能力就越弱。变化幅度可通过变异系数进行计算,公式如下:
其中cv为变异系数,sd为标准偏差,mn为不同失水时刻的压力对组织所做的功的平均值。
通过不同植物组织间cv的比较,即可判定其抗脱水能力,反映抗旱性。
本发明的具体实施过程如下:
步骤一,取正常生长的待测植物的新鲜组织,并用湿布包住植株枝干基部,减缓水分散发;迅速返回实验室,清理所述组织表面灰尘后,用剪刀截取所述新鲜组织上长势较为一致的片段若干,放入湿布中保湿。设置不同形变梯度,利用质构仪测量各形变下组织的最大承压力;其中,各个形变量的设置须以组织未受不可逆损坏为前提。
步骤二,构建形变量与最大承压力之间的线性模型,得到组织的弹性模量;线性模型为胡克定律,即公式(1)。
步骤三,对所选取待测植物组织进行干燥失水处理。
步骤四,设置固定形变,利用质构仪测量植物组织在不同失水时刻的内力变化曲线,得到探头下压距离;其中,固定形变的设置同样须以组织未受不可逆损坏为前提;探头下压距离为测量组织内力变化曲线的探头与植物组织接触开始到下压至所设定的固定形变的位置为止,其间所运行的距离。
步骤五,计算探头下压过程中累积对植物组织所做的功,即为能量;功的计算为公式(2);探头下压过程中,压力随着探头下压距离的增加而增加,其变化符合胡克定律,可采用定积分法计算探头累积对植物组织所做的功,即公式(3)。
步骤六,计算不同失水时刻间能量的变异系数;能量的变异系数的计算公式为公式(4)。
步骤七,根据步骤六中所计算的变异系数定量比较不同植物组织间的抗脱水能力。
实施例:
取正常生长的常春藤和绿萝的新鲜枝条,并用湿布包住植株枝干基部,减缓水分散发;迅速返回实验室,清理所述植物茎表面灰尘后,用剪刀截取所述新鲜枝条上长势较为一致的茎10mm共10段,放入湿布中保湿。设置不同形变梯度(0,5%,10%,15%,20%),测量各形变下茎的最大承压力(见表1)。
表1各形变下常春藤和绿萝茎的最大承压力(n)
利用胡克定律公式对形变量与最大承压力之间的关系进行拟合,拟合曲线如图1。常春藤和绿萝的拟合方程分别为f=92.23x,r2=0.90,p<0.0001以及f=28.54x,r2=0.91,p<0.0001。因此常春藤和绿萝茎的弹性模量k值分别为92.23和28.54。
对所选取待测植物茎进行干燥失水处理;设置固定形变为22%,分别测量两种植物茎在不同失水时刻的内力变化曲线,得到探头下压距离。以0失水时刻常春藤茎的内力变化曲线为例做图,见图2。得到两种植物不同失水时刻设置固定形变下探头的下压距离,见表2。
表2两种植物不同失水时刻设置固定形变下探头的下压距离(mm)
探头下压过程中,压力随着探头下压距离的增加而增加,其变化符合胡克定律,可采用定积分法,分别计算不同失水时刻下探头下压过程中累积对两种植物茎所做的功,见表3。
表3不同失水时刻下探头下压过程中累积对两种植物茎所做的功(10-3j)
由能量的变异系数的计算公式计算不同失水时刻间压力对植物茎所做的功即能量消耗的变异系数,见表4。
表4不同失水时刻间压力对植物茎所做的功即能量消耗的变异系数
由表4可知,不同失水时刻间压力对常春藤茎所做的功即能量消耗的变异系数低于绿萝,常春藤茎中的水分更为稳定,表明其具有较强的抗脱水能力,使其能够在干旱环境下维持较好的水分保持特性,体现出较好的抗旱性。这与实际情况相符。
以上说明仅仅为本发明的较佳实施例,本发明并不限于列举上述实施例,应当说明的是,任何熟悉本领域的技术人员在本说明书的教导下,所做出的所有等同替代、明显变形形式,均落在本说明书的实质范围之内,理应受到本发明保护。
- 还没有人留言评论。精彩留言会获得点赞!