鉴定一种物质对昆虫行为是否有影响及有何种影响的方法与流程
本发明涉及鉴定一种物质对昆虫行为是否有影响和/或有何种影响的方法。
背景技术:
昆虫在其体内可以合成成千上万的小分子化合物或高分子物质,这些物质,如何确定这些物质在其体内行驶何种功能,目前还没有成熟的方法来确定。
例如,在蛾类昆虫的性信息素通讯中,主要是由雌虫释放某种特异性(特定成分及比例)的性信息素,而雄虫在远距离通过触角上数以万计的性信息素感器对其进行感受并产生趋向行为,寻找到雌虫并与之交配。随着化学分析技术的发展,大量的蛾类昆虫的性信息素成分本鉴定,通过行为学以及田间诱捕实验进一步鉴定出此类化合物的行为学功能,但是对于其如何驱使雄虫特定的行为学反应,目前还没有较为系统的研究方法来确定。
很多种类的蛋白都参与了昆虫感知外界的气味分子的过程,气味受体是其中重要的一种。特定的气味分子会激活特定的气味受体,进而昆虫会产生相应的行为,因此有必要鉴定开拓一系列较为完善的实验方法来研究蛾类昆虫感知性信息素的机理研究。
技术实现要素:
鉴于现有技术的缺陷,本发明提供了鉴定一种物质对昆虫行为是否有影响和/或有何种影响的方法,包括1)确定所述物质;2)利用所述物质刺激所述昆虫后,分析所述昆虫的电生理和/或行为学反应。
在一个具体实施方式中,所述物质为所述昆虫自身产生的物质。
在一个具体实施方式中,优选所述物质包括信息素和/或信息素拮抗剂。
在一个具体实施方式中,更优选所述信息素选自顺-11-十六碳烯醛和/或顺-9-十六碳烯醛;所述信息素拮抗剂选自顺-9-十四碳烯醛和/或顺-11-十六碳烯醇。
在一个具体实施方式中,步骤1)还包括1-2)测定所述昆虫产生所述物质的含量和/或相对含量。
在一个优选地具体实施方式中,步骤1)还包括1-2)测定所述昆虫在不同龄期产生所述物质的含量和/或相对含量。
本发明还提供了一种鉴定由昆虫自身产生的物质对昆虫行为是否有影响和/或有何种影响的方法,包括1-1)确定所述物质;1-2)测定所述昆虫产生所述物质的含量和/或相对含量;2-1)确定在昆虫体内与所述物质相关的基因;2-2)以野生型昆虫的虫卵为材料,敲除至少一个与所述物质相关的基因,获得突变型昆虫;3-1)对比野生型昆虫和突变型昆虫对所述物质的电生理反应。
在一个具体实施方式中,优选所述物质包括信息素和/或信息素拮抗剂。
在一个具体实施方式中,更优选所述信息素选自顺-11-十六碳烯醛和/或顺-9-十六碳烯醛;所述信息素拮抗剂选自顺-9-十四碳烯醛和/或顺-11-十六碳烯醇。
在一个具体实施方式中,优选测定所述昆虫在不同龄期产生所述物质的含量和/或相对含量。
在一个具体实施方式中,优选利用CRISPR/Cas9系统敲除至少一个与所述物质相关的基因。
在本发明的研究过程中,利用CRISPR/Cas9系统敲除与所述物质相关的基因并非易事,特别是在昆虫中没有较为成熟的CRISPR/Cas9系统敲除方法的情况下,需要发明人探索可能的受限条件。最终,发明人确定了sgRNA和Cas9mRNA的浓度至关重要。发明人注意到并非注射任意浓度的sgRNA和Cas9mRNA都能使敲除顺利完成,而是敲除过程对两者的浓度要求非常敏感。因此,在一个具体实施方式中,向所述昆虫的卵中注射的sgRNA的浓度为150μg/μl,注射的Cas9mRNA的浓度为1000μg/μl。
在一个具体实施方式中,优选利用单感器记录仪纪录所述野生型和所述突变型对所述物质的电生理反应。
在一个具体实施方式中,所述方法还包括在步骤2-2)之后的步骤3-2)确定所述野生型和所述突变型的行为学反应。
在一个优选地具体实施方式中,在步骤2-2)之后的步骤3-2)为利用Y型管行为学确定所述野生型和所述突变型的行为学反应。其中步骤3-2)可以在步骤3-1)之前或之后。
在一个具体实施方式中,所述方法还包括在步骤2-2)之后的步骤3-3)利用交配行为学实验分别统计突变组和野生组的不同龄期交配成功率,并统计交配后雌蛾的产卵量以及幼虫孵化率。其中步骤3-3)可以在步骤3-1)和/或3-2)之前或之后。
在一个具体实施方式中,其特征在于,通过爪蟾卵母体外表达系统表达所述物质的相关的基因,通过双电机电压钳记录所述相关的基因的表达产物对所述物质的电流反应值,进行电生理分析。其中,所述物质的相关的基因可以是所述物质的靶标受体基因。
在一个具体实施方式中,所述昆虫包括鳞翅目(Lepidoptera)昆虫中的至少一种;优选所述鳞翅目昆虫包括夜蛾科(Noctuidae)昆虫和/或螟蛾科(Pyralidae)昆虫中的至少一种;更优选所述夜蛾科昆虫包括铃夜蛾属(Helicoverpa)昆虫中的至少一种;最优选所述铃夜蛾属昆虫包括棉铃虫(Helicoverpa armigera)。
在一个具体实施方式中,利用GC-MS测定所述物质的含量和/或相对含量。
在一个具体实施方式中,所述野生型昆虫的虫卵为所述昆虫产卵后1小时以内的虫卵。
在本发明的一个具体实施方式中,通过测定所述昆虫在不同龄期产生性信息素成分的相对含量,首次发现性信息素拮抗剂参与调控所述昆虫的最佳生殖状态的交配时机,为进一步鉴定昆虫自身产生的化合物调控不同生理状态下行为趋向提供理论依据。因此,在本发明的中,还提供了一种利用昆虫自身产生的信息素拮抗剂能够调控所述昆虫交配时机在防治昆虫中的应用。
在一个具体实施方式中,优选所述信息素拮抗剂选自顺-9-十四碳烯醛和/或顺-11-十六碳烯醇。
在一个具体实施方式中,利用昆虫在不同时间产生信息素的含量和产生信息素拮抗剂的含量的不同而调控所述昆虫交配时机在防治昆虫中的应用。
本发明通过CRISPA/Cas9技术可以对所述昆虫的一个或多个功能基因位点进行高效的编辑。在虫卵注射前的处理过程中,通过低温处理延迟虫卵的发育,用以保障注射的有效性。注射过程中,筛选出最佳打靶效率的sgRNA和Cas9mRNA的浓度比例(150μg/μl sgRNA和1000μg/μl Cas9mRNA)。
本发明结合生物信息学、生物化学,行为学和电生理学技术全面解析了所述现象的分子与神经机制,提供一套较为完善研究方法,用于研究外界以及昆虫自身产生的化合物调控其特定的行为的机理。
附图说明
图1为1-4日龄雌虫腺体中顺-11-十六碳烯醇(Z11-16:OH)和顺-11-十四碳烯醛(Z9-14:Ald)的相对含量的鉴定图谱;A为1-4日龄雌虫腺体化合物鉴定的离子色谱图;B为Z11-16:OH(左图)和Z9-14:Ald(右图)在1-4日龄雌虫腺体中的相对含量值统计图。通过GC-MS鉴定1-4日龄期的雌虫释放的性信息素成分的含量变化。
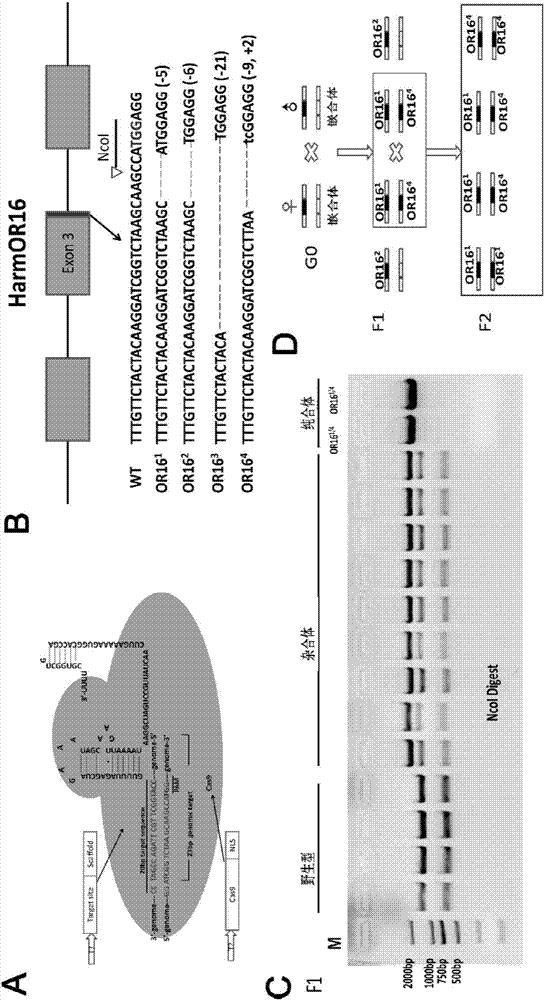
图2为CRISPR/Cas9敲除OR16的流程图;A为CRISPR/Cas9技术的作用原理图;B为OR16基因的靶标位点展示图;C为限制性内切酶检测F1代突变体的电泳图;D为纯合突变体筛选的流程图。
图3为OR16突变型和野生型雄蛾的神经自发反应和对性信息素反应的图谱;A为C类型毛型感器下受体-神经表达模式图;B为突变体和野生型C类型感器的自发电生理反应图谱;C为突变体和野生型C类型感器下OR6和OR16受体神经的自发反应值统计图;D为C类型感器对1mg性信息素成分的电生理反应图;E为OR6和OR16受体神经对性信息素成分反应值统计图。
图4为突变型和野生型雄虫Y型管实验结果图。
图5为突变和野生组个体羽化后7d内交配发生的概率分布图。
图6为不同交配日龄下的雌虫繁殖后代的统计图。
图7为突变和野生组交配的雌虫繁殖后代的统计图。
具体实施方式
下面通过以确定棉铃虫信息素拮抗剂在其体内行驶功能的具体实施例说明本发明。以下实施例用于说明本发明,但不用来限制本发明的范围。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的实验材料,如无特殊说明,均为市售常规生化试剂。
在过去的几十年中,大量的棉铃虫的性信息素成分被鉴定出来,其中包括主要成分顺-11-十六碳烯醛(Z11-16:Ald)和顺-9-十六碳烯醛(Z9-16:Ald),前者是用于长距离的吸引雄蛾,后者是用于增加对雄蛾的吸引作用。此外性信息素成分中还包括一些次要组分,其中包括两种性信息素拮抗剂顺-11-十六碳烯醇(Z11-16:OH)和顺-9-十四碳烯醛(Z9-14:Ald)。前期的研究结果表明羽化2天的雌虫释放的Z11-16:Ald和Z9-16:Ald的量达到最大值,但是雌虫在羽化3天后才达到最佳交配状态。这种不相符的结果表明性信息素拮抗剂可能参与调控雌虫最佳生殖状态下的选着性交配。
鉴于此,本申请人首先通过GC-MS测定性信息素拮抗剂在不同龄期的雌虫腺体提取物的相对含量,结果表明,随着日龄的增加,只有Z11-16:OH的相对含量呈现动态变化,结合雌虫性发育状态以及交配率的变化,初步推断Z11-16:OH在腺体中含量是雄虫判断雌虫性发育状态及择偶的标准,即性未成熟的雌虫通过提高Z11-16:OH的含量趋避雄虫,性成熟的雌虫通过降低Z11-16:OH的含量召唤雄虫进行交配。为了进一步阐述其分子与神经机制,本申请人通过基因编辑系统CRISPR/Cas9敲除Z11-16:OH激活的OR16受体,结果发现,纯合突变体丧失对Z11-16:OH电生理反应以及趋避行为学反应,更重要的是,这些电生理以及行为学上缺陷导致了雄虫错误的认为性未成熟的雌虫已经做好了交配的准备,强行的和雌虫进行交配,这种未发育成熟的条件下的交配必然造成后代发育的贫瘠。因此,本发明鉴定出棉铃虫性信息素拮抗剂的一种新功能,即作为调控开关控制雌雄个体在最佳状态下进行交配及繁育后代,为开发新的害虫防治技术提供了理论基础。
本发明的DNA或RNA的浓度用微量紫外/可见分光光度计NanoDrop ND-2000(Nanodrop,美国)仪器定量。
实施例1
棉铃虫腺体成分的鉴定和分析
1.分别以1-4日龄处女雌蛾为试虫,在进入暗器后第6-7h提取性信息素,用手轻轻挤压雌蛾腹部,迫使其伸出性信息素腺体,用手术刀片将腺体切下,置于微型的玻璃管中,加入10μl的正己烷溶剂浸泡30min后进行气相色谱分析。
2.GC-MS分析:所用的气质联用仪为HP-6890毛细管气相色谱仪和Agilent 5973质谱仪,毛细管色谱柱的型号为Agilent J&W DB-1MS GC柱,30m,0.25mm,0.25μm。色谱工作条件为:起始温度40℃,保持4min,然后以10℃/分钟的升温速度升至280℃,然后保持20分钟。
3.性信息素组分采用与标准化合物保留时间相比较,并结合GC-MS的方法定性;采用性信息素组分的气谱峰面积比较进行相对定量。结果见图1。结果表明2日龄雌虫开始合成性信息素成分,其中性信息素拮抗剂Z11-16:OH的相对含量到达了高峰,随后,Z11-16:OH在三日龄雌虫性腺成分中的含量开始迅速降低,四日龄雌虫腺体成分中近乎无法鉴定Z11-16:OH的含量。相比于Z11-16:OH,棉铃虫另一种性信息素拮抗剂Z9-14:OH在不同龄期雌虫的性腺成分中含量几乎不变。
实施例2
CRISPR/Cas9敲除HarmOR16(OR16)受体
本实验室通过爪蟾卵母细胞表达系统及双电极电压钳技术鉴定出感知Z11-16:OH的靶标受体是性信息素受体OR16。
CRISPR/Cas9敲除OR16的流程图见图2。
1.sgRNA的设计和体外合成:根据sgRNA靶标位点的设计准则(5’-GG(18N)NGG-3’),选择一段位于OR16第三外显子区域的小片段序列5’-ggatcggtctaagcaagccatgg-3’为靶标位点。同时sgRNA序列的最前端是T7启动子序列,用于sgRNA体外转录。合成sgRNA的双链DNA模板是通过长链PCR的扩增得到PCR反应体系:2×Premix PrimeSTAR Hs 25μL,HarmOR16-F1和HarmOR16-R1(10μM)1.5μL,不含核酸酶的水22μL。PCR反应条件:98℃预变性1min;98℃变性10s,60℃退火15s,72℃延伸30s,40个循环。PCR产物用全式金PCR产物回收试剂盒(按照试剂盒说明书操作)进行回收,回收后产物用T7Kit(Ambion)转录试剂盒(按照试剂盒说明书操作)转录成sgRNA。其中HarmOR16-F1:TAATACGACTCACTATAGGATCGGTCTA AGCAAGCCAGTTTTAGAGCTAGAAATAGC;HarmOR16-R1:CAAAATCTCG ATCTTTATCGTTCAATTTTATTCCGATCAGGCAATAGTTGAACTTTTTCACCGTGGCT CAGCCACGAAAA。
2.Cas9mRNA的合成:将前端带有SV40核定为星号(NLS)Cas9基因片段亚克隆到pTD1-T7载体上,pTD1-T7载体质粒进行单酶切线性化后,最后使用mMESSAGET7Kit(Ambion)(按照试剂盒说明书操作)试剂盒转录合成Cas9mRNA。
3.棉铃虫卵的微型注射:收集新鲜的卵置于载玻片上,并用双面胶固定。将合成好的Cas9mRNA和sgRNA按照一定比例混合后(Cas9mRNA的终浓度为1000ng/μl,sgRNA的终浓度为150ng/μl),通过eppendorf显微注射仪注射到虫卵中,每粒卵的注射量为2-10nL,上述操作必须在产卵后的1个小时内完成。总共注射大约1000粒卵,注射后的卵置于25℃和65%相对湿度的环境下培养至孵化。
4.突变体的检测:为了检测注射后个体的突变效率,首先用天根全基因组提取试剂盒TIANamp Genomic DNA Kit(按照试剂盒说明书操作)提取全虫的基因组DNA。设计特异性引物扩出含有靶标位点的基因组片段(1800-2000bp)。PCR反应体系:2×Premix PrimeSTAR Hs 25μL,HarmOR16-F2和HarmOR16-R2(10μM)1.5μL,DNA模板2μL,不含核酸酶的水20μL。PCR反应条件:98℃预变性1min;98℃变性10s,60℃退火15s,72℃延伸2min,40个循环。本发现设计的靶标位点中含有一个酶切位点NcoI,便于使用限制性内切酶检测突变效率。吸取10μL的PCR扩增产物,然后加入1U单位的NcoI限制性内切酶酶切30min,进行电泳检测,单一条带的电泳结果表明是纯合突变,三条电泳条带表明是杂合突变,两条电泳条带表明是未发生突变。其中,HarmOR16-F2:AAGTACTGCGATTCATGCTACTTATCG;HarmOR16-R2:GT ATAATACACGCGATAGAATATCCGC。
5.纯合突变体的筛选:注射后的虫卵大概有10%个体能够生长到成虫期,我们随机配对单头雌雄个体进行交配、产卵,总计有40组,待每组中雌虫产完卵后,提取整头虫子的基因组,检测突变效率,成功筛选出一对父母本突变效率都很高的组合,并且产生250个F1代。F1代个体进入成虫期以后,对F1代个体进行上述配对筛选,最终筛选出一对父母本都是纯合体突变的组合,其产生F2代纯合个体用于电生理和行为学实验。
实施例3
单感器记录仪记录感器对性信息素成分的电生理反应
1.首先选取3-4日龄未交配的雄蛾个体放入Axygen 1.5mL离心管,雄蛾的头部露出离心管端口,通过牙腊将蛾子的下唇须固定在端口边缘,露出其中一根触角,固定在沾有双面胶的盖玻片上。
2.固定好的雄蛾用于单感器记录,我们用已经磨好的钨丝电极插入雄蛾的复眼,用作参比电极。连接嗅觉放大器(sytech)的记录电极也是通过磨好的钨丝电极插入单感器的基部,直到能够出现稳定电信号(信号的出现的频率要明显高于噪音信号)。以上记录的操作是在莱卡高分辨率显微镜Z16APO microscope at 920×magnification下进行。
3.为了方便刺激物的传递,我们首先吸取用石蜡油溶剂配制好的性信息溶液10μl滴加到大小为0.8cm×2.6cm的滤纸片上,将此滤纸片放入巴斯德管中,静止挥发5min后用作刺激源。刺激时,通过一个300毫秒的空气脉冲作用将巴斯德吸管气味以20ml/s的气流速度打入一个14厘米长的金属管控制器,最后通过一个经过加湿和过滤后的恒定气流将金属控制器中的刺激气味传递到固定好的触角表面。刺激反应后的信号通过前置放大器放大十倍进行采集,记录反应的信号图通过Autospike(Syntech)软件进行分析,分析感器下嗅觉神经的数量是通过信号冲动的增幅差异确定,反应值是通过统计刺激前一秒和后一秒信号的冲动的数量差值。结果见图3。记录结果表明,突变型和野生型雄虫C类型下着生的OR16和OR6神经元,可以通过信号的振幅大小区分,振幅大的信号代表OR16神经元,振幅小的信号代表OR6神经元。虽然突变体和野生型OR16神经都能维持正常的自发信号,但是突变体OR16神经自发信号的强度明显要低于野生型的OR16神经的自发信号强度。性信息素刺激反应结果表明OR16受体的敲出使OR16神经元对Z11-16:OH和Z9-14:Ald的电生理反应完全丧失,但是却没有影响OR6神经元对Z9-16:Ald和Z9-14:Ald的反应。
实施例4
Y型管行为学实验
挑选进入暗器4h的3-4日龄雄虫进行Y型管(直径6cm,主臂长60cm,分支臂长45cm)行为学实验。滤纸(2cm×2cm)放置在分支壁的末端,实验室气流以1L/min速度通过滤纸,一端分壁的滤纸上滴加10μg的混合物A(97%Z11-16:Ald+3%Z11-16:Ald)作为对照气味,另一端分壁的滤纸上滴加10μg的混合物B(A+5%Z11-16:OH)或混合物C(A+10%Z9-14:Ald)。将单头雄蛾放置在主臂端口,气流通过滤纸片、分支臂,最后到达主臂端口,5min以后观察雄蛾停落的位置。为了避免雄虫个体之间气味的交叉污染,每次操作完以后,Y型管要用75%酒精擦洗Y型管管壁,室温烘干后才能再次使用。
实施例5
棉铃虫交配行为学以及产卵行为的观察实验
1.设当晚羽化的成虫蛾龄为1,第二天的成虫蛾龄为2,以此类推,将当日羽化的棉铃虫成分别配对置于600mL的透明熟料杯中,杯底放一吸水棉球,每天定时补充饲料,然后观察其交配行为,观察从羽化当天一直延续到羽化后第7天,同时,在塑料杯的顶部用纱布遮盖,以便成功交配的雌虫在上产卵。由于成虫很少在光期交配,因此观察主要是在夜间(20:00—06:00)进行。结果见图5。本实验分为对照组和突变组,对照组由一对野生型雌雄虫个体配对,而突变组是由突变型雄虫和野生型雌虫配对。对照组交配行为学实验表明刚羽化的雌雄虫个体不进行交配,2日龄的个体开始出现少许的交配现象,3日的个体的交配率到达高峰,4日龄个体交配率开始大幅度下降,5、6日龄个体的不在进行交配。这种日龄动态交配分布规律与Z11-16:OH在腺体中相对含量完全吻合,进一步说明雌虫是通过Z11-16:OH含量调控雌雄之间的交配行为。与对照组相比,突变组的交配高峰期发生在羽化后第二天,这是由于突变雄虫无法感知Z11-16:OH,错误识别雌虫释放的信号,强行与其交配。
在上述交配实验中,如有发现雌雄成功交配,每天更换纱布,统计纱布上的卵粒数,直至雌蛾死亡。把产卵的纱布放入培养皿中保湿培养,直至幼虫孵化,统计孵化出幼虫的数量。研究结果发现3日龄交配的雌虫的生殖能力明显高于其他龄期交配的雌虫生殖能力(图6),突变组雌虫的生殖能力明显低于野生组雌虫的生殖能力(图7)。
LHA1760112 核 苷 酸 序 列 表
<110> 中国农业科学院植物保护研究所
<120> 鉴定一种物质对昆虫行为是否有影响及有何种影响的方法
<130> LHA1760112
<160> 4
<170> PatentIn version 3.5
<210> 1
<211> 57
<212> DNA
<213> 人工序列
<223>HarmOR16-F1
<400> 1
TAATACGACTCACTATAGGATCGGTCTAAGCAAGCCAGTTTTAGAGCTAGAAATAGC ;
<210> 2
<211> 80
<212> DNA
<213>人工序列
<223> HarmOR16-R1
<400> 2
CAAAATCTCGATCTTTATCGTTCAATTTTATTCCGATCAGGCAATAGTTGAACTTTTTCACCGTGGCTCAGCCACGAAAA;
<210>3
<211> 27
<212> DNA
<213> 人工序列
<223> HarmOR16-F2
<400> 3
AAGTACTGCGATTCATGCTACTTATCG;
<210> 4
<211> 24
<212> DNA
<213> 人工序列
<223> HarmOR16-R2
<400> 4
ATAATACACGCGATAGAATATCCGC。
- 还没有人留言评论。精彩留言会获得点赞!