一种针对PCSK9的抗高血脂蛋白疫苗的制作方法
本发明属于生物医药领域,涉及一种针对PCSK9的抗高血脂蛋白疫苗。
背景技术:
心血管疾病是影响人类健康的一项重大疾病,每年大约有30%的人死于心血管疾病,WHO 2014年调查结果显示心血管疾病是全球的首要致死原因。高胆固醇血症与冠心病的发生有密切的联系,其主要表现为血浆低密度脂蛋白胆固醇(LDL-C)的增加。高血脂的传统治疗药物主要是他汀类药物,但他汀类药物具有较大的毒副作用,很多患者对他汀药物不耐受。
前蛋白转换酶枯草溶菌素9(PCSK9)是新发现的治疗高血脂的新靶点,其致病机制是:PCSK9的催化域与低密度脂蛋白受体(LDLR)的EGF-A域结合,阻断LDLR运输低密度脂蛋白胆固醇(LDL-C)的功能,且PCSK9-LDLR复合物将LDLR运输到溶酶体内,使LDLR降解,而PCSK9从复合物中分离并重新利用,从而导致机体内LDLR水平降低,LDL-C水平上升,引发高血脂症。此外,PCSK9的C末端虽然不直接与LDLR结合,但其可影响PCSK9与LDLR的结合,从而间接的影响LDLR的功能。图1为PCSK9的晶体结构及与LDLR的作用位点。近年来已有多个靶向PCSK9的单克隆抗体进入临床研究阶段,2015年7月,FDA批准了安进的PCSK9抑制剂evolocumab单克隆抗体和赛诺菲与再生元制药的alirocumab单克隆抗体上市,这标志着以PCSK9为靶点的药物进入了新纪元,开始挑战他汀类药物作为治疗高血脂传统药物的霸位,进一步说明了PCSK9作为治疗高血脂新靶点的可行性和广阔情景。
抗PCSK9单克隆抗体的作用机制是在体内直接与PCSK9结合,中和PCSK9的生物学作用。目前上市和进入临床研究阶段的PCSK9单克隆抗体都是通过封闭PCSK9与LDLR的EGF-A的结合区域,即完全封闭PCSK9的功能。但完全抑制PCSK9,将会导致肝再生能力的下降,因此限制了其对于肝损伤病人的应用。此外单克隆抗体采用真核系统进行表达,表达量较低,纯化工艺复杂,成本较高,且作用时间短,需反复注射,可能激发机体产生抗抗体而对药物耐受。
技术实现要素:
:
本发明的目的是提供一种治疗高血脂的新型蛋白疫苗,为治疗高血脂提供一种新的有效选择。
本发明的技术方案之一是提供了一种针对PCSK9的抗高血脂的蛋白疫苗。该疫苗是以PCSK9蛋白催化域片段多肽为主要活性成分。
其中,上述疫苗中的PCSK9蛋白催化域片段多肽的氨基酸序列为SEQ No.1所示。
进一步的,上述蛋白疫苗还含有PCSK9的C末端片段多肽为活性成分。
其中,上述疫苗中的PCSK9的C末端片段多肽的氨基酸序列为SEQ No.2所示。
其中,上述疫苗中所述的PCSK9的C末端片段多肽和PCSK9蛋白催化域片段多肽可位于同一包装中或分别位于独立包装中。
其中,上述疫苗中所述的PCSK9的C末端片段多肽和PCSK9蛋白催化域片段多肽的配比为1︰0.01~100。
进一步的,上述疫苗中所述的PCSK9的C末端片段多肽和PCSK9蛋白催化域片段多肽的配比为1︰0.1~10。
更进一步的,上述疫苗中所述的PCSK9的C末端片段多肽和PCSK9蛋白催化域片段多肽的配比为1︰0.5~5。
其中,上述疫苗中还含有免疫佐剂。所述的免疫佐剂可为氢氧化铝佐剂、CpG佐剂或脂质体佐剂等本领域常用的免疫佐剂中的至少一种。
此外,本发明还提供了PCSK9蛋白的催化域多肽片段在制备抗高血脂疫苗中的用途。所述PCSK9蛋白催化域片段多肽的氨基酸序列为SEQ No.1所示。
同时,本发明还提供了PCSK9蛋白的C末端多肽片段在抗高血脂疫苗中的用途。所述PCSK9蛋白的C末端多肽片段的氨基酸序列为SEQ No.2所示。
本发明的有益效果在于:本发明蛋白疫苗能刺激机体产生免疫反应,中和PCSK9。本发明的PCSK9蛋白疫苗治疗剂量低,作用时间持久,无需反复注射,可以避免单克隆抗体可能激发机体产生抗抗体而对药物耐受和成本高的缺点。本发明PCSK9疫苗无PCSK9的生物学活性,在体内不会与LDLR结合,避免LDLR被降解。本发明疫苗可以有效的诱导特异性的免疫反应,且产生靶向相应结构域的中和抗体,能有效降低低密度脂蛋白胆固醇(LDL-C)的含量,在作为高血脂治疗性疫苗方面具有很好的应用前景。
附图说明
图1、PCSK9的空间结构以及与与LDLR的作用位点示意图。
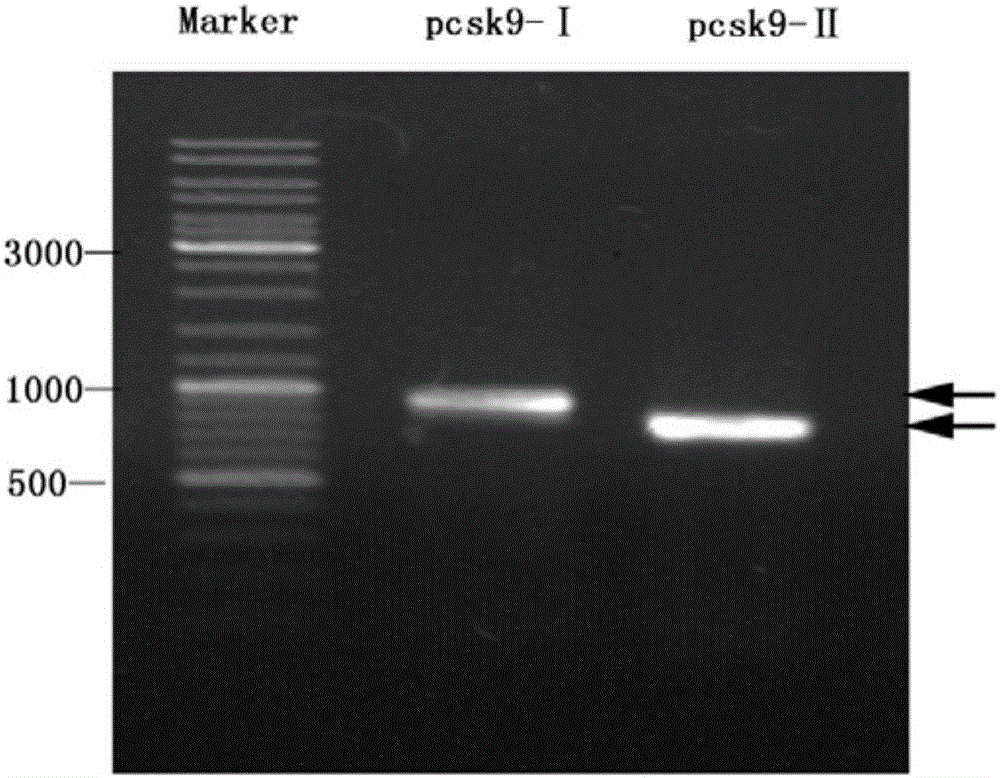
图2、获得小鼠催化域和C末端基因的PCR产物经1%琼脂糖凝胶电泳电泳分析的结果。图中序号说明:M)DNA分子量标准——10000bp标准;1)催化域PCR产物;2)C末端PCR产物;箭头显示催化域和C末端PCSK9基因位置。
图3、PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的鉴定及菌株的筛选;(A)表示PCSK9-Ⅰ蛋白;(B)表示PCSK9-Ⅱ蛋白。
图4、PCSK9-Ⅰ和PCSK9-Ⅱ蛋白纯度的鉴定;
(A)考马斯亮蓝染色鉴定PCSK9-Ⅰ和PCSK9-Ⅱ蛋白;
(B)Western blot验证PCSK9-Ⅰ和PCSK9-Ⅱ蛋白。
图5、rmPCSK9、PCSK9-I、PCSK9-II、PCSK9-I+PCSK9-II蛋白对LO2细胞LDLR和PCSK9的影响。
图6、免疫后小鼠血清中抗PCSK9-Ⅰ和PCSK9-Ⅱ蛋白抗体滴度;(A)小鼠血清中抗PCSK9-Ⅰ蛋白抗体滴度;(B)小鼠血清中抗PCSK9-Ⅱ蛋白抗体滴度。
图7、免疫后小鼠血清中抗体特异性鉴定;图中序号说明:M)蛋白分子量标准—170KD标准;1)PCSK9-Ⅰ蛋白疫苗血清抗rmPCSK9蛋白;2)PCSK9-Ⅱ蛋白疫苗血清抗rmPCSK9蛋白;3)PCSK9-Ⅰ+PCSK9-Ⅱ蛋白疫苗血清抗rmPCSK9蛋白。
图8、蛋白疫苗免疫后产生中和抗体,阻断PCSK9与LDLR的结合;1泳道:Control,未进行任何处理组;2泳道:rmPCSK9蛋白处理组;3泳道:PCS血清处理组;4泳道:PCSK9-Ⅰ蛋白疫苗血清组;5泳道:PCSK9-Ⅱ蛋白疫苗血清组;6泳道:PCSK9-Ⅰ+PCSK9-Ⅱ蛋白疫苗血清组。
图9、蛋白疫苗免疫后小鼠血清血脂水平的变化情况。血清低密度脂蛋白胆固醇(LDL-C)的变化;血清总胆固醇(TC)的变化;血清高密度脂蛋白胆固醇(HDL-C)的变化;血清甘油三酯(TG)的变化。
具体实施方式
目前靶向PCSK9的单克隆抗体有2个已经上市,抗PCSK9单克隆抗体的作用机制是在体内直接与PCSK9结合,中和PCSK9的生物学作用。目前上市和进入临床研究阶段的PCSK9单克隆抗体都是通过封闭PCSK9与LDLR的EGF-A的结合区域,即完全封闭PCSK9的功能。但完全抑制PCSK9,将会导致肝再生能力的下降,因此限制了其对于肝损伤病人的应用。此外单克隆抗体采用真核系统进行表达,表达量较低,纯化工艺复杂,成本较高,且作用时间短,需反复注射,可能激发机体产生抗抗体而对药物耐受。
本发明从另一个思路,考虑疫苗可能通过主动免疫,产生针对靶分子的中和抗体,可能会对肥胖起到预防和治疗的目的;且疫苗与单克隆抗体相比,无需多次注射,即可达到治疗目的,降低了成本,适用于更多的支付体系。因此,本发明考虑开发以PCSK9为靶点的治疗性疫苗,考虑利用同源同种抗原不易产生免疫排斥,以及蛋白不同结构域功能不同的性质,制备靶向PCSK9催化域和C末端的结构域蛋白疫苗,通过注射PCSK9不同结构域的多肽片段作为疫苗,产生靶向PCSK9不同结构域的抗体而中和机体自身的PCSK9,阻断PCSK9的生物学功能。
本发明设计并制备了靶向PCSK9不同结构域的蛋白疫苗,包括靶向PCSK9催化域的蛋白疫苗PCSK9-Ⅰ和靶向PCSK9C末端的蛋白疫苗PCSK9-Ⅱ。
同种不同结构域的PCSK9蛋白疫苗主要特点是:它由来源于免疫对象的同一种属的PCSK9蛋白的不同结构域组成,利用同一种属的蛋白不易发生免疫排斥,可以避免免疫排斥反应;利用PCSK9蛋白不同结构域的功能不同,使机体产生靶向PCSK9不同结构域的中和抗体,从而达到治疗疾病的目的,并且避免个体对完全阻断PCSK9的不耐受。例如制备靶向小鼠PCSK9催化域和C末端的蛋白疫苗,靶向催化域的PCSK9-Ⅰ蛋白疫苗来自小鼠PCSK9蛋白催化域,包含296个氨基酸,其氨基酸序列为SEQ No.3所示;靶向C末端的PCSK9-Ⅱ蛋白疫苗来自小鼠PCSK9蛋白C末端,包含242个氨基酸,氨基酸序列为SEQ No.3所示。而使用人的靶向催化域的PCSK9-Ⅰ蛋白疫苗来自人PCSK9蛋白催化域,其氨基酸序列为SEQ No.1所示;靶向C末端的PCSK9-Ⅱ蛋白疫苗来自人PCSK9蛋白C末端,氨基酸序列为SEQ No.2所示。
同种不同结构域的PCSK9蛋白疫苗制备方法是:利用cDNA序列进行聚合酶链式反应(PCR)合成可溶型PCSK9-Ⅰ和PCSK9-ⅡcDNA片段,载入原核表达载体(如大肠杆菌的表达载体pET32a等),测序鉴定后经诱导表达、分离纯化,得到目的蛋白。可用蛋白疫苗免疫小鼠,检测其免疫原性和治疗效果。
本发明还提供了上述蛋白疫苗的用途及体内治疗效果和安全性评价,获得了很好的结果
此外,本发明还提供了上述蛋白疫苗的制备方法。该方法包括如下步骤:
1.构建不同结构域的PCSK9基因的载体
利用cDNA序列进行PCR反应扩增PCSK9催化域和C末端的cDNA片段,并通过核酸内切酶酶切和DNA连接酶连接构建至表达载体(pET32a)上,分别记作pET32a-pcsk9-Ⅰ和pET32a-pcsk9-Ⅱ。
2.不同结构域PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的表达和纯化
将重组表达质粒转化至大肠杆菌菌株BL21(DE3)中进行表达,分别筛选出高表达PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的菌株,重组蛋白蛋白主要以包涵体的形式存在,包涵体蛋白经不同浓度的尿素洗涤后得到粗蛋白,进一步通过复性和离子交换层析纯化后得到相对较纯的PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白,经免疫印迹鉴定正确。
进一步的对上述PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白进行体外生物学活性检测。良好的蛋白疫苗应当不具有生物学活性,不能直接与配体结合,影响配体的功能。
根据具体情况将上述获得的蛋白制成疫苗,或者加入本领域常用的各种佐剂制成疫苗。PCSK9-Ⅰ和PCSK9-Ⅱ制成疫苗时,可以制作成在一个包装内的单一制剂,也可各自制成制剂,分别包装,一起配合使用。
而实例中用不同浓度的PCSK9-Ⅰ和PCSK9-Ⅱ重组多肽(如2、4、6、8μg/ml)处理高表达PCSK9蛋白的人正常肝细胞LO2,检测PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白对细胞LDLR表达水平的影响,检测重组蛋白在体内是否直接与LDLR作用,从而导致有害的介导LDLR的降解。结果表明PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白对细胞LDLR的表达水平无影响,
本发明的研究结果证实由同源同种的靶向PCSK9具体结构域的抗体疫苗可以降低免疫排斥反应;靶向具体结构域的蛋白疫苗,可以避免完全阻断PCSK9带来的不耐受和毒副作用。本领域技术人员通过上述实例,有足够的理由相信在人身上使用人的PCSK9-Ⅰ和PCSK9-Ⅱ结构域的蛋白疫苗也能起到同等的抗高血脂的作用。此外,常用的佐剂,如用氢氧化铝或CpG为佐剂即可刺激免疫反应产生中和抗体,达到降血脂的目的,降低了成本和滥用佐剂带来的毒副作用。以下将结合实施例和附图详细说明本发明,应当说明的是,实施例是对本发明的技术方案的进一步说明,并不表明本发明仅限于这些实施例。
实施例1 PCSK9不同结构域基因的获得和载体的构建
图2显示了PCSK9催化域和C末端的核苷酸序列及相应蛋白氨基酸序列。
以小鼠cDNA序列为模板进行PCR反应扩增催化域(pcsk9-Ⅰ)和C末端(pcsk9-Ⅱ)cDNA片段,并通过核酸内切酶酶切和DNA连接酶连接构建至表达载体(pET32a)上,分别记作pET32a-pcsk9-Ⅰ和pET32a-pcsk9-Ⅱ。
其中pET32a-pcsk9-Ⅰ载体的构建如下:
以小鼠pcsk9的cDNA为模板,设计并合成相应的引物。
上游:GGGGTACCAGCATCCCATGGAACCTGGA(SEQ No.7);下游:CGGTCA ATGGGTGCTGGGGGGC AGTG(SEQ No.8)。其中斜体部分为EK酶位点,下划线部分为KpnI酶切位点,斜体部分为Xho I酶切位点。经PCR反应,得到片段的大小与预期相同(891bp左右)(图2)。回收PCR产物片段后,插入载体pET32a(+)(购于Novagen公司)中,通过Invitrogen公司测序后,结果显示pET32a-pcsk9-Ⅰ的序列与小鼠催化域的序列(SEQ No.4)一致,证明pET32a-pcsk9-Ⅰ载体构建成功。
pET32a-pcsk9-Ⅱ载体的构建如下:
以小鼠pcsk9的cDNA为模板,设计并合成相应的引物。
上游:GGGGTACCGAGACAGGCGGGCAGCTGCT(SEQ No.9);下游:CGGTCACTGAACCCAGGAGGCCT(SEQ No.10);其中斜体部分为EK酶位点,下划线部分为Bgl II酶切位点,斜体部分为Xho I酶切位点),经PCR反应,得到片段的大小与预期相同(729bp左右)(图2)。回收PCR产物片段后,插入载体pET32a(+)(购于Novagen公司)中,通过Invitrogen公司测序后,结果显示pET32a-pcsk9-Ⅱ的序列与小鼠C末端的序列(SEQ No.6)一致,证明pET32a-pcsk9-Ⅱ载体构建成功。
实验结果表明:
本实施例通过PCR反应扩增PCSK9催化域的DNA序列pcsk9-Ⅰ和PCSK9C末端的DNA序列pcsk9-Ⅱ;并成功的插入表达载体pET32a(+)中,构成pET32a-pcsk9-Ⅰ和pET32a-pcsk9-Ⅱ重组质粒,经酶切和测序验证正确,表明重组质粒构建成功。
实施例2不同结构域PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的表达和纯化
将重组表达质粒转化至大肠杆菌菌株BL21(DE3)中进行表达,经0.01M IPIG,25℃,220rpm诱导表达8h,筛选出高表达PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的菌株,电泳结果表明得到的目的蛋白大小与预计的相似(图3)。对重组蛋白进行可溶性鉴定,主要以包涵体的形式存在。包涵体蛋白经洗涤、复性和离子交换层析后得到相对较纯的PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白,经免疫印迹鉴定正确(图4),且产量可达1-2mg/ml。
实验结果表明:重组表达质粒转化至大肠杆菌菌株BL21(DE3)中表达出PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白,经鉴定以包涵体的形式存在。包涵体蛋白经洗涤、复性和离子交换层析后纯化PCSK9-Ⅰ和PCSK9-Ⅱ重组蛋白,并经蛋白免疫印迹鉴定正确。
实施例3抗高血脂PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗活性检测
以高表达PCSK9蛋白的人正常肝细胞LO2为靶细胞,检测PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的生物学活性。在高营养(DMEM+10%FBS)的条件下培养LO2细胞密度至80%左右,将培养基更换为含10μM普伐他汀的低营养(DMEM+5%FCS)条件下培养18h,诱导细胞LDLR的表达。18h后,将培养基更换为高营养(DMEM+10%FBS)培养基,同时加入不同浓度的PCSK9-Ⅰ和PCSK9-Ⅱ蛋白(2、4、6、8μg/ml),同时以6μg/ml商品化的rmPCSK9为阳性对照,以不加蛋白的为空白对照,孵育8h后,Western blot检测细胞裂解液中LDLR与PCSK9的水平,结果见图5。
实验结果表明:rmPCSK9、PCSK9-Ⅰ和PCSK9-Ⅱ蛋白对细胞PCSK9的表达水平均无影响。用商品化的rmPCSK9处理LO2细胞后,细胞LDLR蛋白水平下降,且几乎完全抑制细胞LDLR的表达;而用自己经大肠杆菌表达复性的PCSK9重组蛋白对LDLR的水平无影响,说明其在体外无生物学活性。这种无生物学活性的蛋白疫苗可以避免PCSK9蛋白进去体内而发挥其生物学作用,与LDLR结合,从引发高血脂症。
实施例4抗高血脂PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗打破免疫耐受,产生抗体
将6~8周龄雌性BALB/c小鼠随机分为4组:对照组,PCSK9-Ⅰ蛋白免疫组、PCSK9-Ⅱ蛋白免疫组、PCSK9-Ⅰ+PCSK9-Ⅱ蛋白免疫组,每组7只。以Al(OH)3作为佐剂,每只小鼠免疫100ug重组蛋白,500ug Al(OH)3,其中PCSK9-Ⅰ+PCSK9-Ⅱ蛋白免疫组为50ugPCSK9-Ⅰ蛋白+50ug PCSK9-Ⅱ蛋白,用PBS将Al(OH)3和重组蛋白混合物稀释至200ul,充分混匀。小鼠腹股沟皮下多点免疫,两周一次,共4次。在首次免疫前及每次免疫1周后经小鼠眶内静脉丛取血,分离血清。通过ELISA检测血清中抗PCSK9-Ⅰ和PCSK9-Ⅱ蛋白的抗体滴度,结果见图6。
通过Western blot检测小鼠血清抗体的特异性,以商品化纯度较高的rmPCSK9蛋白(全长)为抗原,以PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗免疫小鼠后的抗血清为一抗,检测抗血清能否与rmPCSK9蛋白(全长)结合,结果见图8。
图6表明,PCSK9-Ⅰ和PCSK9-Ⅱ蛋白均能刺激机体产生免疫反应。PCSK9-Ⅰ蛋白疫苗产生抗PCSK9-Ⅰ蛋白的抗体,抗体中位数是2560000,而未产生抗PCSK9-Ⅱ蛋白的抗体。PCSK9-Ⅱ蛋白疫苗产生抗PCSK9-Ⅱ蛋白的抗体,抗体中位数是1280000,未产生抗PCSK9-Ⅰ蛋白的抗体。而PCSK9-Ⅰ+PCSK9-Ⅱ蛋白疫苗既产生抗PCSK9-Ⅰ蛋白的抗体,又产生抗PCSK9-Ⅱ蛋白的抗体,说明本发明设计的不同结构域PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗能够靶向PCSK9蛋白不同结构域。图7表明PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗均能产生特异性的抗rmPCSK9蛋白(全长)的抗体。
实施例5抗高血脂PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗中和抗体检测
以高表达PCSK9蛋白的人正常肝细胞LO2为靶细胞,研究蛋白疫苗免疫后能否产生中和PCSK9生物学功能的抗体。在高营养(DMEM+10%FBS)的条件下培养LO2细胞密度至80%左右,将培养基更换为含10μM普伐他汀的低营养(DMEM+5%CS)条件下培养18h,诱导细胞LDLR的表达。18h后,将培养基更换为高营养(DMEM+10%FBS)培养基,同时加入rmPCSK9蛋白与抗血清的复合物(2ugrmPCSK9蛋白+20ul血清),以只加rmPCSK9蛋白的为阴性对照,以不加蛋白的为空白对照。孵育LO2细胞24h后,Western blot检测细胞裂解液中LDLR与PCSK9的水平,结果见图8。
实验结果表明:用rmPCSK9处理LO2细胞,LDLR水平明显下降,与PBS组血清相比,PCSK9-Ⅰ蛋白疫苗免疫组、PCSK9-Ⅱ蛋白疫苗免疫组和PCSK9+PCSK9-Ⅱ蛋白疫苗免疫组LDLR水平均上升,其中PCSK9+PCSK9-Ⅱ蛋白疫苗免疫组LDLR的水平显著上升,比PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗单独免疫组的LDLR水平明显上升。而各组PCSK9水平无明显变化。说明蛋白疫苗免疫后血清中的抗体可以阻断PCSK9与LDLR的结合,中和PCSK9的生物学功能。
实施例6抗高血脂PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗降低小鼠血清血脂水平
在前述模型第4次免疫小鼠2周后,小鼠眼眶静脉丛取血(取血前禁食24h),分离血清,用日立血生化仪检测血清低密度脂蛋白胆固醇(LDL-C)、总胆固醇(TC)、高密度脂蛋白胆固醇(HDL-C)、甘油三酯(TG)水平的变化,结果见图9。实验结果表明:
PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗免疫小鼠后。与对照组(PBS)相比,PCSK9-Ⅰ免疫组、PCSK9-Ⅱ免疫组、PCSK9-Ⅰ+PCSK9-Ⅱ免疫组血清LDL-C的水平均显著降低(P<0.05),其中PCSK9-Ⅰ+PCSK9-Ⅱ免疫组血清LDL-C水平降低了67%。说明同种不同结构域PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗可显著降低血清血脂水平,而且具有协同作用。其中,单独使用靶向催化域或C末端的蛋白疫苗即可显著降低血清LDL-C的水平,说明对于部分肝损伤不能完全抑制PCSK9的病人,采用部分抑制PCSK9的功能的策略即可起到治疗作用,避免了个体对完全阻断PCSK9的不耐受。
PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗免疫BALB/c小鼠后,小鼠血清TC、TG、HDL-C均略微下降,但尚无显著性差异(P>0.05),说明PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗主要是通过阻断PCSK9与LDLR的结合,而调节血清LDL-C的变化。
实施例7抗高血脂PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗体内安全性评价
为了进一步评价抗高血脂PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗体内安全性,在实施例4的过程中每周称量一次小鼠体重,观察蛋白疫苗对小鼠体重的影响。同时,第4次免疫2周后处死小鼠,分离小鼠血清,对小鼠血清进行血生化分析,主要检测以下指标:ALT、AST、TP、Glu、BUN、UA、LDH、ALB。实验结果表明PCSK9-Ⅰ和PCSK9-Ⅱ蛋白疫苗无明显的毒副作用,对小鼠体重和ALT、AST、TP、Glu、BUN、UA、LDH、ALB等血生化指标无明显的影响,具有较好的体内安全性。
SEQUENCE LISTING
<110> 四川大学
<120> 一种针对PCSK9的抗高血脂蛋白疫苗
<130> A161035KN
<160> 10
<170> PatentIn version 3.3
<210> 1
<211> 300
<212> PRT
<213> Artificial
<220>
<223> 人PCSK9催化域氨基酸序列
<400> 1
Ser Ile Pro Trp Asn Leu Glu Arg Ile Thr Pro Pro Arg Tyr Arg Ala
1 5 10 15
Asp Glu Tyr Gln Pro Pro Asp Gly Gly Ser Leu Val Glu Val Tyr Leu
20 25 30
Leu Asp Thr Ser Ile Gln Ser Asp His Arg Glu Ile Glu Gly Arg Val
35 40 45
Met Val Thr Asp Phe Glu Asn Val Pro Glu Glu Asp Gly Thr Arg Phe
50 55 60
His Arg Gln Ala Ser Lys Cys Asp Ser His Gly Thr His Leu Ala Gly
65 70 75 80
Val Val Ser Gly Arg Asp Ala Gly Val Ala Lys Gly Ala Ser Met Arg
85 90 95
Ser Leu Arg Val Leu Asn Cys Gln Gly Lys Gly Thr Val Ser Gly Thr
100 105 110
Leu Ile Gly Leu Glu Phe Ile Arg Lys Ser Gln Leu Val Gln Pro Val
115 120 125
Gly Pro Leu Val Val Leu Leu Pro Leu Ala Gly Gly Tyr Ser Arg Val
130 135 140
Leu Asn Ala Ala Cys Gln Arg Leu Ala Arg Ala Gly Val Val Leu Val
145 150 155 160
Thr Ala Ala Gly Asn Phe Arg Asp Asp Ala Cys Leu Tyr Ser Pro Ala
165 170 175
Ser Ala Pro Glu Val Ile Thr Val Gly Ala Thr Asn Ala Gln Asp Gln
180 185 190
Pro Val Thr Leu Gly Thr Leu Gly Thr Asn Phe Gly Arg Cys Val Asp
195 200 205
Leu Phe Ala Pro Gly Glu Asp Ile Ile Gly Ala Ser Ser Asp Cys Ser
210 215 220
Thr Cys Phe Val Ser Gln Ser Gly Thr Ser Gln Ala Ala Ala His Val
225 230 235 240
Ala Gly Ile Ala Ala Met Met Leu Ser Ala Glu Pro Glu Leu Thr Leu
245 250 255
Ala Glu Leu Arg Gln Arg Leu Ile His Phe Ser Ala Lys Asp Val Ile
260 265 270
Asn Glu Ala Trp Phe Pro Glu Asp Gln Arg Val Leu Thr Pro Asn Leu
275 280 285
Val Ala Ala Leu Pro Pro Ser Thr His Gly Ala Gly
290 295 300
<210> 2
<211> 240
<212> PRT
<213> Artificial
<220>
<223> 人PCSK9 C末端氨基酸序列
<400> 2
Trp Gln Leu Phe Cys Arg Thr Val Trp Ser Ala His Ser Gly Pro Thr
1 5 10 15
Arg Met Ala Thr Ala Val Ala Arg Cys Ala Pro Asp Glu Glu Leu Leu
20 25 30
Ser Cys Ser Ser Phe Ser Arg Ser Gly Lys Arg Arg Gly Glu Arg Met
35 40 45
Glu Ala Gln Gly Gly Lys Leu Val Cys Arg Ala His Asn Ala Phe Gly
50 55 60
Gly Glu Gly Val Tyr Ala Ile Ala Arg Cys Cys Leu Leu Pro Gln Ala
65 70 75 80
Asn Cys Ser Val His Thr Ala Pro Pro Ala Glu Ala Ser Met Gly Thr
85 90 95
Arg Val His Cys His Gln Gln Gly His Val Leu Thr Gly Cys Ser Ser
100 105 110
His Trp Glu Val Glu Asp Leu Gly Thr His Lys Pro Pro Val Leu Arg
115 120 125
Pro Arg Gly Gln Pro Asn Gln Cys Val Gly His Arg Glu Ala Ser Ile
130 135 140
His Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys Lys Val Lys Glu
145 150 155 160
His Gly Ile Pro Ala Pro Gln Glu Gln Val Thr Val Ala Cys Glu Glu
165 170 175
Gly Trp Thr Leu Thr Gly Cys Ser Ala Leu Pro Gly Thr Ser His Val
180 185 190
Leu Gly Ala Tyr Ala Val Asp Asn Thr Cys Val Val Arg Ser Arg Asp
195 200 205
Val Ser Thr Thr Gly Ser Thr Ser Glu Gly Ala Val Thr Ala Val Ala
210 215 220
Ile Cys Cys Arg Ser Arg His Leu Ala Gln Ala Ser Gln Glu Leu Gln
225 230 235 240
<210> 3
<211> 297
<212> PRT
<213> Artificial
<220>
<223> 小鼠PCSK9催化域氨基酸序列
<400> 3
Ser Ile Pro Trp Asn Leu Glu Arg Ile Ile Pro Ala Trp His Gln Thr
1 5 10 15
Glu Glu Asp Arg Ser Pro Asp Gly Ser Ser Gln Val Glu Val Tyr Leu
20 25 30
Leu Asp Thr Ser Ile Gln Gly Ala His Arg Glu Ile Glu Gly Arg Val
35 40 45
Thr Ile Thr Asp Phe Asn Ser Val Pro Glu Glu Asp Gly Thr Arg Phe
50 55 60
His Arg Gln Ala Ser Lys Cys Asp Ser His Gly Thr His Leu Ala Gly
65 70 75 80
Val Val Ser Gly Arg Asp Ala Gly Val Ala Lys Gly Thr Ser Leu His
85 90 95
Ser Leu Arg Val Leu Asn Cys Gln Gly Lys Gly Thr Val Ser Gly Thr
100 105 110
Leu Ile Gly Leu Glu Phe Ile Arg Lys Ser Gln Leu Ile Gln Pro Ser
115 120 125
Gly Pro Leu Val Val Leu Leu Pro Leu Ala Gly Gly Tyr Ser Arg Ile
130 135 140
Leu Asn Ala Ala Cys Arg His Leu Ala Arg Thr Gly Val Val Leu Val
145 150 155 160
Ala Ala Ala Gly Asn Phe Arg Asp Asp Ala Cys Leu Tyr Ser Pro Ala
165 170 175
Ser Ala Pro Glu Val Ile Thr Val Gly Ala Thr Asn Ala Gln Asp Gln
180 185 190
Pro Val Thr Leu Gly Thr Leu Gly Thr Asn Phe Gly Arg Cys Val Asp
195 200 205
Leu Phe Ala Pro Gly Lys Asp Ile Ile Gly Ala Ser Ser Asp Cys Ser
210 215 220
Thr Cys Phe Met Ser Gln Ser Gly Thr Ser Gln Ala Ala Ala His Val
225 230 235 240
Ala Gly Ile Val Ala Arg Met Leu Ser Arg Glu Pro Thr Leu Thr Leu
245 250 255
Ala Glu Leu Arg Gln Arg Leu Ile His Phe Ser Thr Lys Asp Val Ile
260 265 270
Asn Met Ala Trp Phe Pro Glu Asp Gln Gln Val Leu Thr Pro Asn Leu
275 280 285
Val Ala Thr Leu Pro Pro Ser Thr His
290 295
<210> 4
<211> 891
<212> DNA
<213> Artificial
<220>
<223> 小鼠PCSK9催化域编码核苷酸序列
<400> 4
agcatcccat ggaacctgga gcgaattatc ccagcatggc accagacaga ggaagaccgc 60
tcccctgatg gaagcagcca ggtggaggtg tatctcttag ataccagcat ccagggtgcc 120
catcgggaga ttgagggcag ggtcaccatc accgacttca acagcgtgcc ggaggaggat 180
gggacacgct tccacagaca ggcgagcaag tgtgacagcc acggcaccca cctggcaggt 240
gtggtcagcg gccgggatgc tggtgtggcc aagggcacca gcctgcacag cctgcgtgtg 300
ctcaactgtc aagggaaggg cacagtcagc ggcaccctca taggcctgga gtttattcgg 360
aagagtcagc taatccagcc ctcggggcca ctcgtggttc tgctgcccct ggccggtggg 420
tatagccgca tcctcaacgc tgcctgccgg cacctggcga ggactggggt ggtgctggtt 480
gcagcagctg ggaacttccg ggacgacgcc tgcctctact ccccagcttc tgctccagag 540
gtcatcacag tcggggccac gaatgcccag gaccagccag ttaccttggg gactttgggg 600
actaattttg gacgctgtgt ggatctcttt gcccccggga aggacatcat cggagcgtcc 660
agtgactgca gcacatgctt catgtcacag agtgggacct cacaggctgc tgcccacgtg 720
gccggcattg tggctcggat gctgagccgg gagcccacac ttaccctggc cgagctgcgg 780
cagaggctga tccacttctc taccaaagac gtcatcaaca tggcctggtt ccctgaggac 840
cagcaggtgc tgacccccaa cctggtggcc acactgcccc ccagcaccca t 891
<210> 5
<211> 242
<212> PRT
<213> Artificial
<220>
<223> 小鼠PCSK9C末端氨基酸序列
<400> 5
Glu Thr Gly Gly Gln Leu Leu Cys Arg Thr Val Trp Ser Ala His Ser
1 5 10 15
Gly Pro Thr Arg Thr Ala Thr Ala Thr Ala Arg Cys Ala Pro Glu Glu
20 25 30
Glu Leu Leu Ser Cys Ser Ser Phe Ser Arg Ser Gly Arg Arg Arg Gly
35 40 45
Asp Trp Ile Glu Ala Ile Gly Gly Gln Gln Val Cys Lys Ala Leu Asn
50 55 60
Ala Phe Gly Gly Glu Gly Val Tyr Ala Val Ala Arg Cys Cys Leu Val
65 70 75 80
Pro Arg Ala Asn Cys Ser Ile His Asn Thr Pro Ala Ala Arg Ala Gly
85 90 95
Leu Glu Thr His Val His Cys His Gln Lys Asp His Val Leu Thr Gly
100 105 110
Cys Ser Phe His Trp Glu Val Glu Asp Leu Ser Val Arg Arg Gln Pro
115 120 125
Ala Leu Arg Ser Arg Arg Gln Pro Gly Gln Cys Val Gly His Gln Ala
130 135 140
Ala Ser Val Tyr Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys Lys
145 150 155 160
Ile Lys Glu His Gly Ile Ser Gly Pro Ser Glu Gln Val Thr Val Ala
165 170 175
Cys Glu Ala Gly Trp Thr Leu Thr Gly Cys Asn Val Leu Pro Gly Ala
180 185 190
Ser Leu Thr Leu Gly Ala Tyr Ser Val Asp Asn Leu Cys Val Ala Arg
195 200 205
Val His Asp Thr Ala Arg Ala Asp Arg Thr Ser Gly Glu Ala Thr Val
210 215 220
Ala Ala Ala Ile Cys Cys Arg Ser Arg Pro Ser Ala Lys Ala Ser Trp
225 230 235 240
Val Gln
<210> 6
<211> 729
<212> DNA
<213> Artificial
<220>
<223> 小鼠PCSK9C末端编码核苷酸序列
<400> 6
gagacaggcg ggcagctgct ctgtaggacg gtgtggtcgg cacactcggg gcccactcga 60
acagctacag ctacagcccg ctgtgcccca gaagaggagc tgctgagctg ctccagcttc 120
tccaggagcg ggaggcgtcg tggtgattgg attgaggcca taggaggcca gcaggtctgc 180
aaggccctca atgcatttgg gggtgagggt gtctatgccg tcgcgagatg ctgcctggtt 240
ccccgtgcca actgcagcat ccacaacacc cctgcagcca gagctggcct ggagacccat 300
gtccactgcc accagaagga ccatgttctc acaggctgca gcttccattg ggaagtggaa 360
gaccttagtg tccggaggca gcctgcgctg aggtccagac gtcagcctgg ccagtgcgtt 420
ggccaccagg cggccagtgt ctatgcttcc tgctgccatg ccccagggct ggaatgcaaa 480
atcaaggagc atgggatctc aggtccttca gagcaggtca ctgtggcctg cgaagcagga 540
tggaccctga ctggatgcaa tgtgctccct ggggcatccc tcactctggg agcctacagc 600
gtggacaacc tgtgtgtggc aagagtccat gacactgcca gagcagacag gaccagtgga 660
gaagccacag tagctgctgc catctgctgc cggagccggc cttcagcaaa ggcctcctgg 720
gttcagtga 729
<210> 7
<211> 43
<212> DNA
<213> Artificial
<220>
<223> 扩增小鼠PCSK9催化域上游引物
<400> 7
ggggtaccga cgacgacgac aagagcatcc catggaacct gga 43
<210> 8
<211> 31
<212> DNA
<213> Artificial
<220>
<223> 扩增小鼠PCSK9催化域下游引物
<400> 8
cggaattctc aatgggtgct ggggggcagt g 31
<210> 9
<211> 43
<212> DNA
<213> Artificial
<220>
<223> 扩增小鼠PCSK9C末端下游引物
<400> 9
ggggtaccga cgacgacgac aaggagacag gcgggcagct gct 43
<210> 10
<211> 28
<212> DNA
<213> Artificial
<220>
<223> 扩增小鼠PCSK9C末端下游引物
<400> 10
cggaattctc actgaaccca ggaggcct 28
- 还没有人留言评论。精彩留言会获得点赞!