田鼠巴贝斯虫的体外增殖的制作方法
本申请要求于2016年9月19日提交的美国临时申请no.62/396,697以及于2017年5月15日提交的美国临时申请no.62/506,429的权益,其在此通过引用整体并入本文。
背景技术:
本公开一般涉及用于使田鼠巴贝斯虫(babesiamicroti)(“b.microti”)和相关的“类田鼠”血寄生虫物种持续体外增殖的方法。
相关技术的描述
田鼠巴贝斯虫最初由victorbabes于1885年描述,并且从那时起被越来越多地认为是人类病原体。
发明概述
本文公开了用于田鼠巴贝斯虫的体外增殖的方法。该方法包括在包含宿主红细胞的培养基中于存在补体来源和抗田鼠巴贝斯虫igm抗体来源的情况下孵育田鼠巴贝斯虫。
在一个变型中,使田鼠巴贝斯虫持续增殖一次传代以上。在另一个变型中,使田鼠巴贝斯虫持续增殖至少200小时。
在另一个变型中,igm抗田鼠巴贝斯虫抗体来源基本上不含igg抗体。抗田鼠巴贝斯虫igm抗体来源可以来自与宿主红细胞相同的物种。
在一个实施方案中,补体来源包括与宿主红细胞共培养的细胞或细胞系,其中细胞或细胞系分泌补体至培养基中。细胞或细胞系可选自:鼠pd388d1、仓鼠nil-8胚胎成纤维细胞、balb/c3t3成纤维细胞、肺泡ii型上皮细胞a549、犬dh82、人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在一个变型中,细胞或细胞系是选自以下的哺乳动物细胞或细胞系:人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在另一个变型中,细胞或细胞系来源于选自thp-1、hl60或aml-193的人单核细胞/巨噬细胞谱系。在另一个变型中,细胞或细胞系来源于人内皮细胞、人肝细胞、人胃肠道细胞、人上皮细胞、人肺泡细胞、鼠pd388d1、仓鼠nil-8胚胎成纤维细胞、balb/c3t3成纤维细胞或肺泡ii型上皮细胞a549。在一些实施方案中,补体来源包括外周血单核细胞(pbmc)。
在另一个实施方案中,细胞或细胞系使用transwell系统与宿主红细胞一起共培养或分别培养。
在一个实施方案中,igm抗体来源选自:脾脏淋巴细胞、b细胞、浆细胞、成浆细胞和杂交瘤。
在另一个实施方案中,可将补体来源外源性地添加至培养基。补体可通过自动装置添加。
本文公开了用于使田鼠巴贝斯虫相关的血寄生虫持续体外增殖的方法,所述方法包括在包含宿主红细胞的培养基中于存在补体来源和抗田鼠巴贝斯虫相关的血寄生虫igm抗体来源的情况下,孵育田鼠巴贝斯虫相关的血寄生虫。
田鼠巴贝斯虫相关的血寄生虫可选自:广义田鼠巴贝斯虫(b.microtisensulato)、杆状巴贝斯虫(b.rodhaini)、安妮泰勒虫(theileriaannae)(西班牙犬类病原体)和类田鼠巴贝斯虫某些种(b.microti-likespp)。
根据另一个实施方案,公开了包含田鼠巴贝斯虫的增殖的培养物。培养物包括于存在补体来源和抗田鼠巴贝斯虫igm抗体来源的情况下包含宿主红细胞的培养基。
增殖的培养物可以基本上不含igg抗体。
在增殖的培养物的一个实施方案中,宿主红细胞选自人、小鼠、仓鼠或其他许可性寄主。
在增殖的培养物的变型中,补体来源来自与宿主红细胞相同的物种。在另一个变型中,抗田鼠巴贝斯虫igm抗体来源来自与宿主红细胞相同的物种。
附图简述
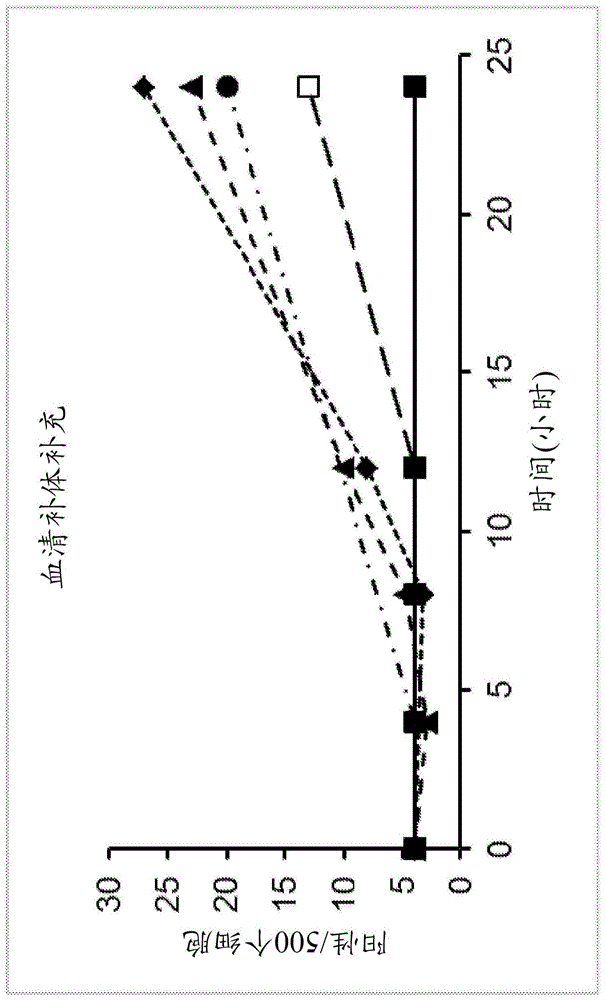
图1显示了血清补体补充对田鼠巴贝斯虫生长的影响。没有补体的对照由(■)描绘,而仅在4小时时补充2%补体(8x-是指比用于其他补体浓度的0.25%多8倍)的另一组对照由(●)描绘(即,其是其他补充剂浓度的8倍,并且仅在4小时标记处添加一次)。在4、8和12小时时补充0.25%补体由(▲)描绘,而(◆)描绘了在8和12小时时补充0.25%补体,并且(□)描绘了仅在12小时时补充0.25%。
图2a-图2d显示了体外培养的田鼠巴贝斯虫westport菌株,第18代的giemsa染色的薄涂片。比例尺=7μm。
图2a显示了指示峰复制的马耳他十字形式。
图2b显示了典型的环形,一个(左下)带有白色觅食液泡。
图2c显示了较低放大倍数的视野,显示了寄生的22个细胞中的4个。
图2d显示了单个细胞内的多个寄生虫。
发明详述
尽管增殖属于巴贝斯虫属各种物种的寄生虫取得了重大进展,但物种田鼠巴贝斯虫仍然是需要在仓鼠和/或小鼠中进行动物传代以增殖的那些物种之一。熟练的研究人员花费了多年的巨大努力,寻找这种单一物种在体外的持续增殖,但都没有成功。
本公开涉及用于在一个或多个宿主细胞中体外增殖田鼠巴贝斯虫的方法。在一些实施方案中,本公开涉及用于使田鼠巴贝斯虫持续体外增殖的方法。田鼠巴贝斯虫的“持续的”体外增殖或培养意指增殖超过单次传代。在一些实施方案中,“传代”意指稀释具有原始数量的寄生虫的培养物(即,原始感染水平、感染百分比或“寄生虫血症”)并允许稀释培养物中寄生虫的数量通过细胞分裂增加直到寄生虫的数量恢复到原始水平。例如,培养物可以1:4稀释,即至寄生虫原始数量的1/4。因此,在一些实施方案中,超过单次传代意指在1:4(或基于感染水平如由本领域普通技术人员确定的另一稀释因子)稀释后,培养物恢复其原始(“传代前”)感染百分比。在一些实施方案中,“持续的”是指增殖数次传代。在一些实施方案中,“持续的”是指增殖超过约1至约100次传代。在一些实施方案中,“持续的”是指长期增殖约1天至约7天、约1周至约4周、约1个月至约12个月和/或约1年至约5年。
在一些实施方案中,短期内“持续的”体外增殖是指至少200小时的增殖。在一些实施方案中,短期内“持续的”体外增殖是指约200小时的增殖。在一些实施方案中,短期内“持续的”体外增殖是指超过200小时的增殖。在一些实施方案中,短期内“持续的”体外增殖是指约100小时至约10,000小时的增殖。
不受任何理论的束缚,据信田鼠巴贝斯虫持续增殖的失败是由于寄生虫不能有效地将其细胞内分裂转化为随后的未感染宿主细胞的渗透导致的。例如,当来自仓鼠和/或小鼠的寄生虫用igm和补体涂覆时,即使寄生虫经历了数倍的细胞分裂,如果涂覆母体寄生虫的igm和/或补体不降解或如果没有igm和补体可用于涂覆子寄生虫,则子寄生虫不能渗透新的未感染的宿主细胞。
不受任何理论束缚,持续增殖要求感染的宿主细胞用需要定期添加的未感染的宿主细胞在新鲜培养基中进行稀释。在一些实施方案中,寄生虫可以通过裂解感染的宿主细胞(如宿主红细胞)来进行纯化,然后与未感染的宿主红细胞一起返回培养物。然而,对于持续增殖而言,这不是必需的。
在一些实施方案中,体外培养开始时使用非常丰富的基础培养基,其中添加了两种不常见的组分。在一些实施方案中,hl-1培养基基础培养基(lonza,walkersville,md)补充有20-40%未加热的胎牛血清、2mm谷氨酰胺二肽(gibco)、100mg/ml硫酸庆大霉素、1mg/mlalbumaxii(gibco)、1xht补体(gibco)、1xhb-101补体(irvinescientific,irvine,ca)、1mml-半胱氨酸-hcl、0.02mm浴铜灵-二磺酸(bcs,sigmachemical)和1x抗生素-抗真菌药(gibco)。在一些实施方案中,培养包括添加和/或混合人和/或仓鼠血清与胎牛血清(fbs),但附加效果不保证将产生额外费用。在一些实施方案中,将两种不常见的组分添加至基础培养基,即补体来源和igm来源。
在一些实施方案中,向该基础培养基中添加来自-80℃储存的等分试样的快速冷冻的仓鼠或人血清(innovativeresearch,novi,mi)。该血清以“仓鼠补体”和“人补体”出售。血清用作补体来源。在一些实施方案中,所使用的稀释通过在24孔组织培养盘中使用连续稀释的试验来确定。在一些实施方案中,短期实验使用1:400稀释液(约0.25%新鲜血清),尽管通常在此血清水平(即补体来源)处观察到低水平的持续感染。通过频繁的血清(补体)补充,例如每4小时或更短,获得更高水平的感染(图1)。在一些实施方案中,在约1-2%的血清水平下,24小时循环可以在没有补充或者每12小时更换一次培养基的情况下完成(图1)。
在一些实施方案中,将igm来源添加到基础培养基。在一些实施方案中,从用田鼠巴贝斯虫感染的阳性供体单位(如人)和仓鼠来获得针对田鼠巴贝斯虫的igm类多克隆抗体的来源血清或血浆。在一些实施方案中,来源血清或血浆是针对田鼠巴贝斯虫的igm类单克隆抗体的来源。
鉴于从活的田鼠巴贝斯虫感染的仓鼠获得血液作为多克隆igm抗体来源的局限,在一些实施方案中,可以使用来自田鼠巴贝斯虫感染的balb/c小鼠的脾淋巴细胞的克隆的igm单克隆抗体。例如,immunopreciseantibodies,ltd.,victoria,bc供应田鼠巴贝斯虫感染的balb/c小鼠的脾淋巴细胞,以产生杂交瘤融合物,以产生克隆的igm单克隆抗体。在一些实施方案中,可以使用igm的一个或多个其它来源。非限制性实例包括b细胞、浆细胞、成浆细胞、杂交瘤等。因此,在一些实施方案中,此类克隆的igm单克隆抗体的使用应使田鼠巴贝斯虫的体外增殖简化并标准化。在一些实施方案中,使用多克隆igm抗体。在一些实施方案中,使用单克隆igm抗体。在一些实施方案中,使用多克隆的和单克隆的igm抗体。在一些实施方案中,igm抗体来自相同的物种(即,同系)。在一些实施方案中,igm抗体可来自不同的物种(即同种异体)。在一些实施方案中,红细胞、补体和igm获自同一宿主。在一些实施方案中,红细胞、补体和igm获自一个或多个不同的宿主(如人、仓鼠和小鼠)。
抗田鼠巴贝斯虫igg抗体与igm抗体竞争。igg抗体在体内使田鼠巴贝斯虫易受调理素作用(opsonize),导致通过巨噬细胞清除田鼠巴贝斯虫。在一些实施方案中,巨噬细胞的清除导致了培养物中寄生虫产量低。
不受任何特定理论的束缚,据信抗田鼠巴贝斯虫igg抗体对培养物的增殖速率是有害的。不受任何特定理论的限制,据信于存在巨噬细胞的情况下增殖时,igg抗体通过寄生虫的滋养体阶段的巨噬细胞引起调理素作用和吞噬作用。因此,不受任何特定理论的限制,据信培养物中的igg百分比与寄生虫产率百分比成反比,这是由于因igg易受调理素作用的寄生虫不能渗透/感染新鲜红细胞,并且当培养物中不存在巨噬细胞时,与使寄生虫能够穿透/感染新鲜红细胞的igm抗体相竞争。
因此,在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源基本上不含igg抗体。在一些实施方案中,通过使用本领域已知的一种或多种可商购获得的igg去除试剂盒,使抗田鼠巴贝斯虫igm抗体来源基本上不含igg抗体。在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源不具有igg或不具有可检测的igg。在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源具有少于约0.0001%、0.001%、0.01%、0.1%或1%的igg。在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源具有少于约0.1%、0.2%、0.3%、0.4%、0.5%、0.6%、0.7%、0.8%、0.9%、1%、5%或10%的igg,或由任何两个上述值限定的范围内的值。在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源具有约0.0001至10%的igg之间的任何百分比。在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源完全不含igg抗体。因此,在一些实施方案中,抗田鼠巴贝斯虫igm抗体来源具有0%的igg抗体。
在一些实施方案中,来自血清或血浆的igm作为冷球蛋白沉淀。在一些实施方案中,通过在2-8℃下针对合适的透析缓冲液进行过夜透析来进行沉淀。在一些实施方案中,透析缓冲液是磷酸钾缓冲液,10mm,ph5.4。在一些实施方案中,本领域普通技术人员可以使用一种或多种其他透析缓冲液。在一些实施方案中,将冷球蛋白用冷透析缓冲液洗涤,然后溶解在缓冲盐水中。在一些实施方案中,乙酸盐缓冲盐水(10mm乙酸盐ph5.4)用于溶解冷球蛋白。由于甚至半纯化的igm也是公知的不稳定,因此将其与等体积的igm稳定稀释剂(2x-指的是其他实施例的2倍)混合并保持在2-8℃。igm稳定稀释剂(1x)由50mm乙酸盐缓冲液(ph5.5)中的20%山梨糖醇和1m甘氨酸组成。由于其对红细胞的负面影响,这种稳定的igm溶液不能在培养基中以低于1:50的稀释度使用,并且其所使用的稀释度通过使用连续稀释的多次体外培养试验来确定。在一些实施方案中,在长期经验之后,使用稀释度从1%下降到大约等于稳定的igm的ifa滴度(1:400)。
在一些实施方案中,支持巴贝斯虫物种的体外增殖的多克隆igm的浓度为约0.001mg/ml至约100mg/ml。在一些优选的实施方案中,支持巴贝斯虫物种的体外增殖的多克隆igm的浓度为约0.01mg/ml至约10mg/ml。在一些实施方案中,浓度为0.001mg/ml至约100mg/ml的多克隆igm支持巴贝斯虫体外增殖约2天至约4天,其倍增时间约为24小时。在一些优选的实施方案中,浓度为约0.01mg/ml至约10mg/ml的多克隆igm支持巴贝斯虫体外增殖约4天,其倍增时间为约12小时。
在一些实施方案中,支持巴贝斯虫物种的体外增殖的单克隆igm的浓度为约0.005mg/ml至约500mg/ml。在一些优选的实施方案中,支持巴贝斯虫物种的体外增殖的单克隆igm的浓度为约0.05mg/ml至约50mg/ml。在一些实施方案中,浓度为约0.005mg/ml至约500mg/ml的单克隆igm支持巴贝斯虫体外增殖约2天至约4天,其倍增时间约为24小时。在一些优选的实施方案中,浓度为约0.05mg/ml至约50mg/ml的单克隆igm支持巴贝斯虫体外增殖约4天,其倍增时间约为12小时。
在一些实施方案中,将igm抗体的不同克隆的共混物(即igm抗体的多于一个克隆的组合)用于巴贝斯虫的体外生长。在一些实施方案中,与单独使用的igm抗体的不同克隆相比,igm抗体的不同克隆的共混物更有效地支持巴贝斯虫的体外生长。在一些实施方案中,与单独使用的igm抗体的不同克隆相比,igm抗体的不同克隆的共混物更为有效约10%至约90%。在一些实施方案中,与单独使用的igm抗体的不同克隆相比,igm抗体的不同克隆的共混物更为有效约10、20、30、40、50、60、70、80、81、82、83、84、85、86、87、88、89或90%,或者在由任何两个上述值限定的范围内的值。在一些实施方案中,不同igm抗体的克隆混合物用于巴贝斯虫的体外生长。在一些实施方案中,igm抗体的不同克隆的最佳浓度可由本领域普通技术人员使用本领域已知的标准技术(如elisa)确定。
在一些实施方案中,至少两种具有不同表位特异性的igm单克隆抗体用于支持体外巴贝斯虫以与多克隆igm相同的水平生长。在一些实施方案中,使用具有不同表位特异性的2至约20的igm单克隆抗体来支持体外巴贝斯虫以与多克隆igm相同的水平生长。
在一些实施方案中,杂交瘤上清液是单克隆igm的来源。在一些实施方案中,将杂交瘤上清液稀释并用作单克隆igm的来源。在一些实施方案中,将杂交瘤上清液稀释20%v/v至约60%v/v,并用作单克隆igm的来源。在一些实施方案中,支持体外巴贝斯虫生长的杂交瘤上清液中igm的浓度为约0.2mg/ml至约200mg/ml。在一些实施方案中,支持体外巴贝斯虫生长的经稀释的杂交瘤上清液中igm的浓度为约2mg/ml至约4mg/ml。在一些实施方案中,支持体外巴贝斯虫生长的经稀释的杂交瘤上清液中igm的浓度为约4mg/ml至约6mg/ml。在一些实施方案中,支持体外巴贝斯虫生长的经稀释的杂交瘤上清液中igm的浓度为约6mg/ml至约12mg/ml。
在一些实施方案中,可以使用一种或多种商业级igm纯化方法来产生本文所述方法的实施方案的毫克量。非限制性实例包括基于固定的甘露聚糖结合蛋白的试剂盒(如nevens,j.r.,等人,affinitychromatographicpurificationofimmunoglobulinmantibodiesutilizingimmobilizedmannanbindingprotein,jchrom(1992)597:247-256;nethery,a.,等人,single-steppurificationofimmunoglobulinmonc1q-sepharose,jimmunolmeth(1990)126:57-60;ohta,m.,等人,themechanismofcarbohydrate-mediatedcomplementactivationbytheserummannan-bindingprotein,jbiolchem(1990)265:1980-1984中所述,其在此通过引用整体并入本文),基于羟基磷灰石的纯化(bioprocessintl.com/upstream-processing/biochemicals-raw-materials/igm-purification-with-hydroxyapatite-349783/,其在此通过引用整体并入本文)和基于琼脂糖的纯化(如labs.mcb.harvard.edu/gaudet/resources_files/gehealthcare_chromatography/don%27t%20move/18103746aa.pdf中所述,其在此通过引用整体并入本文)。单克隆抗体的分离和纯化的其他进展对于本领域普通技术人员来说是已知的,例如,如bioprocessintl.com/upstream-processing/biochemicals-raw-materials/igm-purification-with-hydroxyapatite-349783/中所述,其在此通过引用整体并入本文。
在一些实施方案中,该方法包括共培养方法。在一些实施方案中,共培养方法允许培养物在不需要每天添加补体的情况下发展。在一些实施方案中,共培养方法包括一种或多种细胞(如,原代细胞)或细胞系(如,“原代”或传代细胞系)作为一种或多种补体因子的来源。在一些实施方案中,补体因子是有效的补体因子。在一些实施方案中,补体来源是持续产生补体的细胞或细胞系。在一些实施方案中,补体来源是原代细胞或细胞系。在一些实施方案中,原代细胞或细胞系持续产生补体。在一些实施方案中,只要每2-3天更换一次培养基,原代细胞或细胞系就会无限地产生补体。在一些实施方案中,原代细胞或细胞系产生并分泌补体因子。在一些优选的实施方案中,产生补体的原代细胞或细胞系来自与培养物中使用的红细胞来源相同的物种。在一些实施方案中,补体来源是持续产生补体的单核细胞性白血病细胞系。在一些实施方案中,将作为一种或多种补体因子的来源的细胞或细胞系与宿主红细胞一起或分别用过滤器介导的交界部(如,使用transwell系统)共培养。
在一些实施方案中,补体来源是补体因子c3的来源。不受任何理论的束缚,据信寄生虫结合的igm分子继而被补体因子c3的一个或多个分子结合,然后附着至红细胞上的一个或多个c3受体。不受任何理论束缚,据信针对田鼠巴贝斯虫的igm类抗体固定补体因子c3,所述补体因子c3启动“替代补体途径”。在该途径中,c3结合五聚体igm分子并自发切割成c3a和c3b。对于c3b(和c4b)而言,存在igm-c3复合物可以附着的红细胞受体(cr1)。这种结合的绝对多重性是必需的并且允许寄生虫进入红细胞。红细胞受体(cr1)嵌入流体膜中并迁移至初始附着位点以使整体效应倍增数倍。
在一些实施方案中,预期可以在共培养中使用可以在有或没有刺激的情况下分泌或者合成并分泌补体因子c3的任何细胞或细胞系。在一些实施方案中,补体来源是可以在有或没有刺激的情况下分泌或者合成并分泌补体因子c3的任何细胞或细胞系。在一些实施方案中,可以在有或没有刺激的情况下分泌或者合成并分泌补体因子c3的细胞或细胞系选自:鼠pd388d1、仓鼠nil-8胚胎成纤维细胞、balb/c3t3成纤维细胞、肺泡ii型上皮细胞a549、犬dh82、人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在一些实施方案中,补体来源包括外周血单核细胞(pbmc)。在一些实施方案中,细胞或细胞系分泌或者合成并分泌补体因子c3的刺激是本领域熟知的。例如,在没有限制的情况下,有许多方法可以刺激巨噬细胞或其他补体分泌细胞或细胞系,这取决于特定的细胞类型和基于由特定细胞类型进行补体分泌的天然和/或药物诱导剂。例如,在一些实施方案中,对于巨噬细胞/单核细胞细胞系,pma(佛波醇12-肉豆蔻酸酯13-乙酸酯)用于诱导巨噬细胞表型,并且苯肾上腺素用于进一步刺激趋化因子和补体的产生。
在一些实施方案中,用于共培养方法的一个或多个产生补体的细胞或细胞系是鼠pd388d1、仓鼠nil-8胚胎成纤维细胞、balb/c3t3成纤维细胞、肺泡ii型上皮细胞a549、犬dh82、人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在一些实施方案中,补体来源选自:人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在一些实施方案中,细胞或细胞系是哺乳动物细胞或哺乳动物细胞系。在一些实施方案中,细胞或细胞系是人细胞或人细胞系。在一些实施方案中,人细胞或细胞系是人补体的来源。在一些实施方案中,细胞或细胞系是非人细胞或非人细胞系。在一些实施方案中,补体来源包括外周血单核细胞(pbmc)。
在一些实施方案中,对于每个细胞或细胞系,通过实验确定所需的产生补体的细胞数量。在一些实施方案中,基于所需结果,对每个细胞或细胞系通过实验确定所需的产生补体的细胞数量。在一些实施方案中,共培养物中产生补体的细胞数量为约1x103/ml至约1x105/ml。在一些实施方案中,共培养物中产生补体的细胞的数量为约1x105/ml至约1x108/ml。在一些实施方案中,共培养物中产生补体的细胞的数量为约1x108/ml至约1x1010/ml。在一些实施方案中,共培养物中产生补体的细胞的数量为约1x102/ml、1x103/ml、1x104/ml、1x105/ml、1x106/ml、1x107/ml、1x108/ml、1x109/ml或1x1010/ml,或由上述任何两个值限定的范围内的值。在一些实施方案中,细胞或细胞系具有活跃的代谢和/或生化表型。在一些实施方案中,细胞或细胞系具有活跃的代谢和/或生化表型,例如类似于巨噬细胞。
在一些实施方案中,培养物中的一个或多个细胞或细胞系不提供一种或多种补体因子。在一些实施方案中,还考虑使用除细胞或细胞系之外的其他补体来源。非限制性实例包括血浆、血清和可商购获得的纯化的补体蛋白。在一些实施方案中,将一种或多种补体因子外源性添加至培养物。例如,在一些实施方案中,将补体因子c3或c3b外源性添加至培养物。在一些实施方案中,将一种或多种补体因子手动添加至培养物。在一些实施方案中,以自动方式将一种或多种补体因子添加至培养物。在一些实施方案中,使用自动化机器以自动方式将一种或多种补体因子添加至培养物。在一些实施方案中,使用自动化机器以自动方式将一种或多种补体因子添加至培养物,其中机器可以被编程为以定时方式添加一种或多种补体因子。在一些实施方案中,机器可以被编程为以定时方式添加一个或多个特定量的一种或多种补体因子。
在一些实施方案中,补体因子作为一种或多种冷冻沉淀提供,其被分配到进入培养物的培养基流中。在一些实施方案中,将一种或多种冷冻沉淀分配到进入培养物的培养基流中,并以所需浓度提供在培养物中。在一些实施方案中,将一种或多种冷冻沉淀手动分配到进入培养物的培养基流中,并以所需浓度提供在培养物中。在一些实施方案中,使用机器以自动方式将一种或多种冷冻沉淀分配到进入培养物的培养基流中,并以所需浓度提供在培养物中。
在一些实施方案中,培养物中感染的红细胞(表示为“寄生虫血症”)的分数范围为约0.001%至约80%。在一些实施方案中,补体来源的存在将寄生虫血症改善了约4倍至约10倍。在一些实施方案中,寄生虫血症的改善是基于增殖的田鼠巴贝斯虫的菌株。在一些实施方案中,田鼠巴贝斯虫的菌株选自:狭义田鼠巴贝斯虫(b.microtisensustricto),这包括菌株gi、westport和rmns;广义田鼠巴贝斯虫(类田鼠巴贝斯虫),这包括菌株aw1(kobe)、ho234(hobetsu)。并且其他类田鼠巴贝斯虫物种包括罗氏巴贝斯虫(b.rodaini)和“西班牙狗/犬病原体”(以前被称为安妮泰勒虫)。在一些实施方案中,本公开可适用于巴贝斯虫的其它物种。非限制性实例包括啮齿动物寄生虫杆状巴贝斯虫、巴贝斯虫的犬物种和赤狐巴贝斯虫(b.vulpes)。在一些实施方案中,本领域普通技术人员也可以对本文提供的任何方法进行调整以用于任何类田鼠巴贝斯虫的某些种。其他类田鼠巴贝斯虫寄生虫也包括在其中。例如,不受任何理论的束缚,据信田鼠组将在完整的基因组测序和分析后作为新的属出现。因此,还包括该新的属中的其他寄生虫,其在可获得额外测序数据后被归类和/或被重新归类。
在一些实施方案中,根据培养条件,巴贝斯虫的倍增时间可以在小于约12小时至约24小时的范围内。在一些实施方案中,倍增时间为约8小时至约24小时。在一些实施方案中,倍增时间约为6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23或24小时,或由任何两个上述值限定的范围内的值。
在一些实施方案中,巴贝斯虫物种在遗传上和通过进入红细胞的常用方式相关,并因此,通过本文描述的方法的实施方案适于持续增殖。因此,在一些实施方案中,本文描述的方法的实施方案包括目前在巴贝斯虫属内的密切相关的类田鼠巴贝斯虫(田鼠组)血寄生虫的整个组,包括狭义和广义田鼠巴贝斯虫。这些目前包括日本菌株aw1(kobe)、ho234(hobetsu)、赤狐巴贝斯虫(以前的安妮泰勒虫或“西班牙狗”)、罗氏巴贝斯虫(babesiarodhaini)、猫巴贝斯虫(babesiafelis)等。在一些实施方案中,任何类田鼠巴贝斯虫的某些种适于通过本文所述方法的实施方案持续增殖。
另外的实施方案
在一些实施方案中,用于田鼠巴贝斯虫的体外增殖的方法,该方法包括在包含宿主红细胞的培养基中于存在补体来源和抗田鼠巴贝斯虫igm抗体来源的情况下孵育田鼠巴贝斯虫。在该方法的一些实施方案中,使田鼠巴贝斯虫持续增殖一次传代以上。在该方法的一些实施方案中,使田鼠巴贝斯虫持续增殖至少200小时。在该方法的一些实施方案中,使田鼠巴贝斯虫持续增殖至少200小时。在该方法的一些实施方案中,抗田鼠巴贝斯虫igm抗体来源基本上不含igg抗体。在该方法的一些实施方案中,抗田鼠巴贝斯虫igm抗体的来源来自与宿主红细胞相同的物种。在该方法的一些实施方案中,补体的来源包括与宿主红细胞共培养的细胞或细胞系,并且其中细胞或细胞系分泌补体至培养基中。在该方法的一些实施方案中,细胞或细胞系选自:鼠pd388d1、仓鼠nil-8胚胎成纤维细胞、balb/c3t3成纤维细胞、肺泡ii型上皮细胞a549、犬dh82、人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在该方法的一些实施方案中,细胞或细胞系是选自以下的哺乳动物细胞或细胞系:人thp-1、人hl60、人hela229、hsb-2淋巴细胞性白血病和单核细胞性白血病细胞系。在该方法的一些实施方案中,细胞或细胞系来源于选自thp-1、hl60或aml-193的人单核细胞/巨噬细胞谱系。在该方法的一些实施方案中,细胞或细胞系源自人内皮细胞、人肝细胞、人胃肠道细胞、人上皮细胞、人肺泡细胞、鼠pd388d1、仓鼠nil-8胚胎成纤维细胞、balb/c3t3成纤维细胞或肺泡ii型上皮细胞a549。在该方法的一些实施方案中,细胞或细胞系是外周血单核细胞。在该方法的一些实施方案中,细胞或细胞系使用transwell系统与宿主红细胞一起共培养或分别培养。在该方法的一些实施方案中,igm抗体来源选自:脾脏淋巴细胞、b细胞、浆细胞、成浆细胞和杂交瘤。在该方法的一些实施方案中,将补体来源外源性地添加至培养基。在该方法的一些实施方案中,补体来源通过自动装置添加。在该方法的一些实施方案中,抗田鼠巴贝斯虫igm抗体是多克隆的或单克隆的。在该方法的一些实施方案中,田鼠巴贝斯虫的倍增时间为约12小时至约24小时。
在一些实施方案中,用于田鼠巴贝斯虫相关的血寄生虫的持续体外增殖的方法,该方法包括在包含宿主红细胞的培养基中于存在补体来源和抗田鼠巴贝斯虫相关的血寄生虫igm抗体来源的情况下孵育田鼠巴贝斯虫相关的血寄生虫。在用于持续体外增殖的方法的一些实施方案中,田鼠巴贝斯虫相关的血寄生虫选自:广义田鼠巴贝斯虫、杆状巴贝斯虫或“安妮泰勒虫”(西班牙犬类病原体)以及类田鼠巴贝斯虫的某些种。在用于持续体外增殖的方法的一些实施方案中,抗田鼠巴贝斯虫相关的血寄生虫igm抗体是多克隆的或单克隆的。在用于持续体外增殖的方法的一些实施方案中,田鼠巴贝斯虫相关的血寄生虫的倍增时间为约12小时至约24小时。
在一些实施方案中,增殖的培养物包含田鼠巴贝斯虫或田鼠巴贝斯虫相关的血寄生虫,增殖的培养物包含于存在补体来源和抗田鼠巴贝斯虫igm抗体或抗田鼠巴贝斯虫相关的血寄生虫igm抗体来源的情况下的包含宿主红细胞的培养基。在一些实施方案中,增殖的培养物基本上不含igg抗体。在增殖的培养物的一些实施方案中,宿主红细胞选自人、小鼠、仓鼠或其他许可性寄主。在增殖的培养物的一些实施方案中,在增殖的培养物的一些实施方案中,补体来源来自与宿主红细胞相同的物种。在增殖的培养物的一些实施方案中,抗田鼠巴贝斯虫igm抗体来源或抗田鼠巴贝斯虫相关的血寄生虫igm抗体来源来自与宿主红细胞相同的物种。在一些实施方案中,增殖的培养物还包含与宿主红细胞共培养的细胞或细胞系,并且其中细胞或细胞系分泌补体至培养基中。在增殖的培养物的一些实施方案中,抗田鼠巴贝斯虫igm抗体或抗田鼠巴贝斯虫相关的血寄生虫igm抗体是多克隆的或单克隆的。在增殖的培养物的一些实施方案中,田鼠巴贝斯虫或田鼠巴贝斯虫相关的血寄生虫的倍增时间为约12小时至约24小时。
基于本文的教导,本领域普通技术人员将理解如何实现本发明的全部范围。例如,基于本文的教导,本领域普通技术人员将能够执行长期持续增殖。
实施例
以下实施例是非限制性的,并且本领域普通技术人员预期的其他变型包括在本公开的范围内。
实施例1
购买仓鼠红细胞(biochemedservices,winchester,va)作为具有edta抗凝剂的全血,并且在附近的医疗中心以2周的间隔从本发明人抽取人o型红细胞作为具有edta抗凝剂的全血。
在37℃,含5%co2的空气气氛(优选)或微需氧固定相(masp)气氛(5%co2,2%o2和93%氮气)中,使用模块化孵育室(billups-rothenberginc.,delmar,ca),在24孔细胞培养板(sarstedt)中建立并维持培养物。
随着使用masp气氛的感染百分比增加,往往存在所谓的危机或退化形式、小密集的红细胞内体的趋势。这些培养物从masp气氛转移到含5%co2的空气,氧气是最明显的变量。令人惊讶的是,额外的氧气在一夜之间恢复了正常环形和多个梨状裂殖子的优势,这表明危机形式可能在条件改善时能够再生。在仓鼠感染的晚期阶段也在体内观察到这种现象。
实施例2
在24孔培养板中使用体外培养进行滴定以确定补体和igm抗体的最佳浓度。含人红细胞的培养物与人补体和人igm抗体配对。同样,将含仓鼠红细胞的培养物与仓鼠补体和仓鼠igm抗体配对以获得最佳效果。在添加红细胞和工作培养物(减去可变组分)之前,将可变因子(补体或igm抗体)在培养孔内以小体积连续稀释。对于4孔滴定,将单个生长的培养孔用无菌pbs洗涤两次,并将细胞加入已经补充有另外的新鲜红细胞和滴定的可变组分的4个培养孔中。对于8孔滴定,将两个培养孔一起处理以产生1:4分裂。然后加入储备培养基使每孔的总体积达到1.25ml。过夜孵育后,去除各种培养基,在dulbeccopbs(dpbs)中以1:100稀释1ml等份的红细胞,并在12孔ifa载玻片上点样1ml等分试样以用于测试。根据所测试的组分,载玻片用以下一种或多种染色:抗-igm荧光缀合物、抗-补体c3荧光缀合物或抗田鼠巴贝斯虫igg(具有二级缀合物)。基于田鼠巴贝斯虫的大约20-24小时的体内倍增速率,滴定通常表现出平台期,随后在更高的稀释度下在每400x显微镜视野中荧光寄生虫下降50%。
实施例3–补体滴定
为了追踪随时间推移的补体活性,汇集4孔培养物,在pbs中洗涤两次,然后用未感染的红细胞和新鲜培养基分开以产生具有相同起始材料的完整24孔培养板。将培养物补充人补体,短暂混合并以不同的间隔返回孵育箱(图1)。
图1显示了血清互补补充对田鼠巴贝斯虫生长的影响。没有补体的对照用(■)描绘,而另一组仅在4小时时补充2%补体(8x)的对照用(●)描绘。在4、8和12小时时补充0.25%补体由(▲)描绘,而(◆)描绘了在8和12小时时的补充,并且(□)描绘了仅在12小时时补充0.25%。通过24小时对人细胞和组分的孔进行取样,并用igg免疫血清和抗-igg缀合物染色。
补体保持活跃的短时间表明了使用比平时多数倍的策略以避免每3-4小时必须补充一次,但这一额外的补体活性小于8小时或者将导致红细胞裂解增加而没有伴随的寄生虫增殖增加。得到的血影红细胞间接用免疫血清染色并直接用抗c3缀合物染色,表明了补体介导的裂解。
实施例4
实施例1和2中描述的滴定以类似的方式单独和多次使用,用以优化单克隆抗田鼠巴贝斯虫igm抗体。田鼠巴贝斯虫的倍增时间允许单日(过夜)测定或更长时间(涉及更多,但非常有用)以确定单个或混合克隆的效用。与多克隆抗体功能相似的此类克隆的可用性使得该技术可以更广泛地被研究人员接受。
具有igm单克隆抗体的培养物表明igm单克隆抗体有效并且田鼠巴贝斯虫可以在无血清培养基中成功生长。培养基包含rpmi1640-hepes(50%)和leibovitzl15(50%),其中含有谷氨酰胺、抗生素和2x胰岛素-转铁蛋白-硒补充剂,用于thp-1细胞。倍增时间小于24小时(接近12小时)。
实施例5
经培养并且目前正在培养扩增的菌株中的一个来自没有复杂的巴贝斯虫病的供体,所述供体用薄涂片诊断出患有特征性寄生虫血症并且通过ifa检测证明针对田鼠巴贝斯虫的抗体滴度增加了大于4倍。薄涂片寄生虫血症记录为0.12%(每800个计数红细胞1个裂殖子),并且在体外8次传代(1:4培养分裂)后,不同培养孔中的感染率为4-6%。gi菌株至少每3-4天传代一次持续超过两个月,但随方法的每一次改进,百分比增加了。最终这种菌株(分型为westport菌株)在冷冻前达到19代,增加了2750亿倍,在此期间培养物从冷冻原种中复活3次。图2显示了在第18代,体外培养的田鼠巴贝斯虫westport菌株的薄涂片。
组分和方法的商业可用性允许更准确地测试和开发用于巴贝斯虫病的治疗、药物和诊断测定,以及帮助对田鼠巴贝斯虫的生命周期进行基础研究。
实施例6
来自血清或血浆的igm通过在2-8℃下针对透析缓冲液(磷酸钾缓冲液,10mmph5.4)过夜透析而作为冷球蛋白沉淀。用冷透析缓冲液洗涤冷球蛋白,然后将其溶于乙酸盐缓冲盐水(10mm乙酸盐ph5.4)中。由于甚至半纯化的igm也是公知的不稳定,因此将其与等体积的igm稳定稀释剂(2x-指的是其他实施例的2倍)混合并保持在2-8℃。igm稳定稀释剂(1x)由50mm乙酸盐缓冲液(ph5.5)中的20%山梨糖醇和1m甘氨酸组成。由于其对红细胞的负面影响,这种稳定的igm溶液不能在培养基中以低于1:50的稀释度使用,并且其所使用的稀释度通过使用连续稀释的多次体外培养试验来确定。在一些实施方案中,在长期经验之后,使用稀释度从1%下降到大约等于稳定化的igm的ifa滴度(~1:400)。
实施例7
使用包括甘露聚糖结合蛋白柱的pierceigm纯化试剂盒(44897)从细胞培养上清液中纯化igm。100ml上清液产生约2mg的igm。与igg相比,igm抗体具有更低的亲和力(affinity)但更高的亲合力(avidity)。然而,与igg杂交瘤相比,igm杂交瘤倾向于分泌更大量的抗体。来自杂交瘤的igm分泌浓度为约30mg/ml至约50mg/ml。这与igg分泌相反,igg分泌为约5mg/ml至约20mg/ml。
实施例8
在一个实验中,在表现出对田鼠巴贝斯虫的反应性的igm单克隆抗体的82个克隆中,igm单克隆抗体的17个克隆也在体外能够增殖。igm单克隆抗体的这17个克隆在体外培养的第2天支持增殖为背景水平的3倍以上。17个克隆支持狭义田鼠巴贝斯虫和广义田鼠巴贝斯虫菌株的体外生长。
实施例9
在另一个实验中,在igm单克隆抗体的274个克隆中,igm单克隆抗体的96个克隆对田鼠巴贝斯虫的反应性范围从部分/弱反应性到强反应性。在96个克隆中,16个克隆在体外能够增殖。igm上清液的滴定范围从未稀释至约1:100,例如未稀释、1:2、1:10和1:100。
在16个克隆中,选择10个克隆,其也使得能够按倍增时间为24小时在体外增殖,并且在体外培养中在3天时,产生背景水平的至少4倍的增加(通过免疫荧光测定法来测定)。对于7个克隆,使用60%v/v稀释的培养上清液测试体外增殖。如通过elisa测定,60%v/v稀释的上清液中的igm浓度为约6mg/ml至约12mg/ml。
多克隆igm和单克隆igm的生长速率的比较表明,共混的igm克隆将是有益的,即与单独使用单个igm克隆相比,使用igm的克隆混合物将是有益的。
本公开中引用的所有参考文献通过引用整体并入本文。
本公开不限于本申请中描述的旨在作为各方面的说明的特定实施方案。在不脱离本发明的精神和范围的情况下,可以进行许多修改和变化,这对本领域技术人员来说是显而易见的。除了本文列举的那些之外,本公开范围内的功能等同的方法和装置对于本领域技术人员而言从前面的描述中是显而易见的。这些修改和变化旨在落入所附权利要求的范围内。本公开仅受所附权利要求的条款以及这些权利要求所赋予的等同物的全部范围的限制。应理解,本公开不限于特定的方法、试剂、化合物、组合物或生物系统,其当然可以变化。还应理解,本文使用的术语仅用于描述特定实施方案的目的,而不是限制性的。
本领域技术人员将理解,对于本文公开的这个和其他过程和方法,在过程和方法中执行的功能可以以不同的顺序实现。此外,概述的步骤和操作仅作为实例提供,并且一些步骤和操作可以是可选的,组合成更少的步骤和操作,或者扩展到附加的步骤和操作而不偏离所公开的实施方案的本质。
如本领域技术人员将理解的,出于任何和所有目的,诸如就提供书面描述而言,本文公开的所有范围还涵盖任何和所有可能的子范围及其子范围的组合。任何列出的范围可以容易地被识别为充分描述并且使得相同的范围被分解为至少相等的一半、三分之一、四分之一、五分之一、十分之一等。作为非限制性实例,本文讨论的每个范围可以容易地分解为下三分之一、中间的三分之一和上三分之一等。如本领域技术人员还将理解的,诸如“至多”、“至少”等的所有语言包括所引用的数字并且指代其中的范围,可以随后将所述范围分解为如上所述的子范围。最后,如本领域技术人员将理解的,范围包括每个单独的成员。因此,例如,具有1-3个细胞的组是指具有1、2或3个细胞的组。类似地,具有1-5个细胞的组是指具有1、2、3、4或5个细胞的组,以此类推。
从前述内容可以理解,本文已经出于说明的目的描述了本公开的各种实施方案,并且在不脱离本公开的范围和精神的情况下可以进行各种修改。因此,本文公开的各种实施方案不旨在是限制性的,真正的范围和精神由所附权利要求指示。
- 还没有人留言评论。精彩留言会获得点赞!