梓醇在治疗糖尿病肝损伤的药物中的应用
1.本发明属于制药技术领域,具体涉及一种梓醇在改善ages致肝窦内皮细胞损伤中的应用。
背景技术:
2.哺乳动物糖尿病肝损伤(diabetic liver disease)是糖尿病的一种严重并发症。糖尿病和胰岛素抵抗也是糖尿病患者肝损伤的重要因素,糖尿病是一种由高糖状态引起的微血管并发症。在兽医领域中,使用1型糖尿病和2型糖尿病的分类方法,动物该病的发病的机制跟人类糖尿病的发病机制类似。糖尿病是犬、猫的常见病,犬猫糖尿病的发病率约占0.4%-1.2%,犬糖尿病几乎95%以上都是胰岛素依赖性糖尿病(1型糖尿病),非胰岛素依赖性糖尿病(2型糖尿病)极少见;猫糖尿病在1型糖尿病和2型糖尿病中各占约50%,严重状态下会合并酮症酸中毒而危及生命。糖尿病终末期会造成肝脏的损伤。根据世界糖尿病联盟(international diabetes federation)的报道,2011年糖尿病的发病人数达到3.66亿人次,预计到2030年,糖尿病的发病人数将达到5亿人次。流行病学研究表明,糖尿病患者患慢性糖尿病的风险增加肝脏疾病和肝细胞癌的患病风险。此外,1型糖尿病与高血糖有关的慢性肝损伤的风险,但其潜在机制仍基本未知。在2型糖尿病的发展过程中,胰岛素抵抗与炎症有关,是慢性肝病发病机制相关的重要因素。肝脏是哺乳动物代谢的中枢,也是糖尿病慢性并发症累及的主要靶器官之一。由于肝脏强大的代偿功能,糖尿病引起的早期肝损伤往往不易发现,而进展至肝硬化时,则失去了治疗的最佳时机;同时在血糖调节及糖的贮存、分布等方面肝脏同样发挥重要作用,而糖尿病性肝损伤往往会进一步使糖代谢紊乱程度恶化。因而糖尿病的治疗保肝护肝与控制血糖同等重要。
3.晚期糖基化终末产物(advanced glycation end products,ages)是糖尿病患者体内沉积于组织器官的毒性产物,它是在无酶的条件下,是由体内的还原糖醛基和蛋白质、脂肪酸或核酸的氨基基团之间发生的糖基化反应,又称为美拉德(maillard)反应。循环系统中过量的ages沉积于肝脏,会触发炎症、脏器纤维化以及氧化应激状态,造成肝脏的进一步损伤。galectin-3作为ages的受体之一,在介导组织器官损伤方面发挥重要作用。临床试验证明,2型糖尿病患者血清中galectin-3显著升高。galectin-3的水平和血浆中的血糖、c反应蛋白以及胰岛素拮抗的水平呈现明显的相关性,且高水平的galectin-3和糖尿病血管功能紊乱导致的并发症紧密关联。cd146是由一个胞外结构域(1-339个氨基酸),一个跨膜结构域(560-583个氨基酸)和一个胞浆内尾链(584-646个氨基酸)组成的ⅰ型整合素膜蛋白。cd146主要表达与平滑肌、大脑、毛囊、骨髓、激活的t淋巴细胞以及血管内皮细胞上galectin-3可以与cd146的n-糖基化位点结合,具备从而促进促炎因子的分泌,而这些促炎细胞因子对于内皮粘附,细胞增殖、迁移及血管新生都发挥着巨大的作用
14.。
4.近年来由糖尿病所致的肝损伤发病率正在迅速增长。糖尿病导致的肝脏损伤受到高度的关注,这类肝损伤通常以肝脏的炎症反应、肝硬化和脂质沉积为主要特征。在这些病理表现中,以肝脏的炎症反应最为显著,在糖尿病狗实验模型中,也发现了肝脏的病理损
伤,研究表明一些抗炎药物能够发挥降低血糖,增强胰岛素敏感性和保护糖尿病导致的肝损伤的效果。在体内高糖状态下,ages是通过糖的酮基或醛基与蛋白质的氨基之间的非酶糖基化反应产生的一种毒性产物。ages能够促进炎症反应的生成和活性氧(reactive oxygen species,ros)的生成,高糖加速ages在血浆和组织的形成和积累,从而导致糖尿病血管并发症。研究报道,肝窦内皮细胞清除受体主要介导ages的内吞摄取功能,ages大类积累会损伤肝窦内皮细胞的清除受体的功能,从而进一步导致细胞的损伤。ages通过不同类型的受体发挥其功能,半乳糖凝集素-3(galectin-3)是其受体之一,在肝细胞中表达,主要介导肝脏细胞保护的功能。本发明中着重基于ages对于galectin-3的直接作用,用慢病毒载体成功建立rlsecs中galectin-3过表达。
5.galectin-3是一种β半乳糖基结合凝集素,参与细胞粘附、细胞迁移、细胞凋亡和血管生成等生物学功能,其在肝脏巨噬细胞(库普弗细胞)和内皮细胞广泛表达。本发明用过慢病毒过表达以及敲减galectin-3实验结果表明,梓醇需要通过galectin-3的抑制进一步发挥抗炎症细胞的迁移浸润作用。cd146是细胞粘附分子免疫球蛋白超家族的成员,也是血管内皮细胞表面的关键细胞粘附蛋白。本发明进一步研究发现梓醇通过抑制ages诱导的galectin-3和cd146的共定位结合偶联发挥作用,并不影响rlsecs细胞内两种蛋白的各自细胞内总表达量。
技术实现要素:
6.梓醇是从一种从生地黄根中提取的环烯醚萜苷类生物活性成分,然而,其与糖尿病病理产物ages沉积造成肝脏血管损伤作用及其血管内皮galectin-3的关系未曾报道,本发明验证了梓醇对于ages大鼠肝窦内皮细胞炎性损伤的保护作用,进而应用于制备治疗糖尿病肝损伤的药物。
7.本发明的目的是克服上述不足之处提供一种梓醇在制备治疗糖尿病肝损伤的药物中的应用。
8.进一步的,所述的糖尿病肝损伤是肝窦血管内皮损伤。
9.进一步的,所述的肝窦血管内皮损伤是由ages致肝窦内皮细胞损伤。
10.本发明实验结果显示,梓醇可降低大鼠肝窦内皮细胞促炎因子icam-1和mcp-1的分泌,抑制巨噬细胞的激活,降低巨噬细胞no的释放量,发挥显著的抗炎作用。与此同时,肝脏炎症发生的同时会有大量的肝脏巨噬细胞(库普弗细胞)浸润。
11.本发明的目的是通过以下方式实现的:
12.利用慢病毒lv-galectin-3-gfp转染rlsecs,在细胞内过表达galectin-3。将0、0.1、1、10μmol/l梓醇分别孵育rlsecs 48h后,cck-8法观察细胞增殖的影响。将10μmol/l梓醇孵育0、12、24、48、96h,同上法观察细胞增殖影响。设置空白对照组(control)、模型组(ages)、1μmol/l梓醇组、10μmol/l梓醇组、阳性对照gb1107组,给药组加入药物孵育48h后,观察以上各组的rlsecs形态变化,乳酸脱氢酶(lactate hydrogenase,ldh)法检测细胞的损伤程度,elisa法观察各组mcp-1、icam-1的分泌量。将巨噬细胞raw264.7种板,同上法分组并给药,48h后,griess法观察各组一氧化氮(nitric oxide,no)向细胞上清液的释放量。将raw264.7细胞种于transwell小室,rlsecs种于孔板底部,将细胞分为control组、模型组、梓醇组、梓醇+lv-galectin-3-gfp组、梓醇+lv-galectin-3-shrna组,分别给药及慢病
毒预孵30min后,在培养基中加入终浓度为200μg/ml的ages刺激,48h后用结晶紫法观察巨噬细胞的透膜细胞数量。将control组、ages组、10μmol/l梓醇组、阳性对照gb1107组的rlsecs孵育药物48h后,免疫荧光法观察galectin-3和cd146的共定位情况。将control组,ages组和10μmol/l梓醇组的蛋白样品通过免疫共沉淀法和western blotting法检测galectin-3和cd146的相互作用以及各自的表达量。结果发现,与lv-gfp组相比,lv-galectin-3-gfp组转染至rlsecs 48h后呈现高表达。与0μg/mlages组相比,200μg/mlages组细胞活力下降显著(p<0.01),后续实验用此浓度进行造模刺激。与空白对照组相比,0.1、1和10μmol/l梓醇组的细胞增殖率轻微升高但无显著差异(p>0.05);与孵育0h相比,10μmol/l梓醇孵育48h有显著差异(p<0.05)。与ages组相比,梓醇组改善ages导致的细胞损伤,显著降低rlsecs上清液ldh的活力、mcp-1和icam-1的释放量(p<0.05)。与ages组相比,梓醇组抑制巨噬细胞raw264.7的极化作用,显著降低细胞no的释放量(p<0.05)。通过慢病毒载体过表达和敲低rlsecs和raw264.7的galectin-3,证明梓醇通过抑制该分子改善巨噬细胞的浸润作用,进一步证实梓醇通过促进galectin-3和cd146分子的解偶联作用发挥肝窦血管内皮保护作用。
13.由此,梓醇通过抑制galectin-3改善ages致肝窦血管内皮细胞的损伤以及促炎因子的释放,降低巨噬细胞的激活和no的分泌,并通过促进galectin-3和cd146分子的解偶联作用发挥肝窦血管内皮的保护作用,通过离体实验证明其发挥改善糖尿病ages沉积造成的肝损伤的作用机制,进而可以有效应用于制备治疗糖尿病肝损伤的药物。
附图说明
14.图1为lv-galectin-3-gfp对于rlsecs的转染作用图(比例尺:200μm);
15.图2梓醇对于ages诱导的rlsecs形态和ldh活性的影响图;
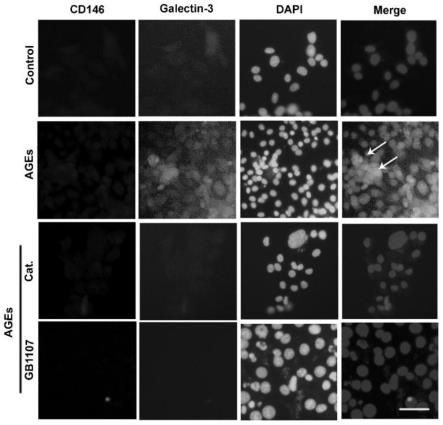
16.a,光镜形态学观察结果;b,rlsecs ldh活力检测统计结果图;
17.图中,肩标不同字母表示差异显著(p<0.05);肩标相同字母或无字母标注表示差异不显著(p>0.05);
18.图3梓醇对于ages诱导的raw 264.7细胞形态和no含量的影响图;
19.a,光镜形态学观察结果;b,raw 264.7细胞no含量检测统计结果图;
20.a,结晶紫染色观察结果;b,raw 264.7细胞迁移细胞量统计结果图;
21.图4梓醇对于ages诱导的raw 264.7细胞迁移的影响图;
22.图5梓醇对于ages诱导的rlsecs细胞膜上galectin-3和cd146共定位的影响图;
23.图6梓醇对于ages诱导的rlsecs上galectin-3和cd146的co-ip和western blotting检测图。
具体实施方式
24.以下通过具体试验例对本发明进行进一步解释说明。
25.1.材料与方法
26.1.1材料
27.1.1.1药物与试剂
28.梓醇(cas号:2415-24-9,批号:z00511803015)购自成都瑞芬思生物科技有限公
司;高糖dmem(货号:c11995500bt)购自美国hyclone公司;cck-8试剂盒(货号:c0038)、ldh试剂盒(货号:20220504)均购自上海碧云天生物技术有限公司;lv-gfp、lv-galectin-3-gfp、lv-galectin-3-shrna,购自北京合生基因科技有限公司;一氧化氮(no)测定试剂盒(货号:20220322)购自南京建成生物工程研究所;胎牛血清(fbs,货号:ab20214230)购自上海诺娃医药科技有限公司公司;青链霉素混合液(100
×
)双抗(货号:p1400)、ripa组织/细胞裂解液(货号:p0013)、5
×
上样缓冲液(货号:p0015)、丙烯酰胺(acr-bis)(货号:a853851)、1.5m tris,ph=8.8(货号:st789)、1m tris,ph=6.8(货号:st768)、temed(货号:st728)、封闭用羊血清(货号:sl038)、dapi溶液(即用型)(货号:c0065)、丽春红染色液(货号:p0012)均购自北京solarbio公司;mcp-1elisa试剂盒(货号:ml028554)、icam-1elisa试剂盒(货号:ml023954)均购自上海酶联生物科技有限公司;预染蛋白marker(10-180kd)(批号:01114357)、supersignal
tm
west pico plus化学发光底物(货号:34580)均购自thermo fisher scientific公司;hrp-羊抗小鼠igg(货号:ba1050)、hrp-羊抗兔igg(货号:ba1054)、cy3-羊抗兔igg(货号:ba1032)、fitc-羊抗小鼠igg(货号:ba1101)购自武汉博士德生物工程有限公司;4%多聚甲醛溶液(货号:bl539a)购自兰杰柯科技有限公司;β-actin抗体(货号:23660-1-ap)、galectin-3抗体(货号:60207-1-ig)、cd146抗体(货号:66153-1-ig)购自proteintech公司。
29.1.1.2细胞株
30.大鼠肝窦内皮细胞(编号:bncc359686)购自北京北纳创联生物技术研究院;小鼠巨噬细胞(raw264.7 macrophage)(目录号:scsp-5036)购自中国科学院细胞库。
31.1.2方法
32.1.2.1慢病毒包装和转染试验
33.用rt-pcr扩增galectin-3的编码序列。pcr片段和pgc-fu质粒经ecor i酶切,然后与t4dna连接酶连接,构建质粒plv-galectin-3-gfp。将质粒转化到dh5α大肠杆菌中进行鉴定。引物(galectin-3)序列如下:正向:5'-atgctgatcacaatcatagg-3',反向:5'-tcaccagcgcttcccacgcc-3'。使用100μl lipofectamine 2000,将20μg pgc fu质粒与目标cdna、15μg phelper1.0质粒和10μg phelper2.0质粒共转染293t细胞,以生成重组慢病毒lv-galectin-3-gfp。以上过表达病毒和lv-galectin-3-shrna由北京合生基因科技有限公司包装,shrna-galectin-3靶序列为5'-gcaacacgaagcaggacaata-3'。向96孔板中接种密度为5
×
103个rlsecs,40%-50%的融合率,从孵箱中取出96孔板,确定细胞均生长良好。弃去旧培养液,将病毒加入96孔板中,每孔100μl。以等量的生理盐水(saline)作为空白对照组,以及空病毒(lv-gfp)作为阴性对照组,在慢病毒染后48h观察细胞中荧光表达情况。
34.1.2.2cck-8检测细胞的增殖效应
35.每孔加入100μl 5
×
103个细胞进行培养进行以下实验:(1)给予0.1、1、10μmol/l梓醇孵育细胞;(2)将10μmol/l梓醇分别孵育12、24、48及96h,以上所有实验以不含fbs的dmem培养基作为空白对照组。孵育48h后每孔加入10μlcck-8溶液。在细胞培养箱内继续孵育1h,在450nm测定od值。以确定最佳给药浓度和最适给药时间。
36.1.2.3ldh实验检测细胞的损伤效应
37.将1
×
106/ml rlsecs接种于96孔板,将细胞分为control组、ages组、1μmol/l梓醇组、10μmol/l梓醇组、阳性药(gb1107)组,用含10% fbs的dmem培养基培养24h,更换为不含
fbs的dmem培养基饥饿培养24h,给药组加入相应含药培养基孵育30min后,在培养基中加入终浓度为200μg/ml的ages刺激,培养48h后,收集细胞培养液上清,按照ldh试剂盒说明书检测450nm处的od值,计算ldh活性。
38.1.2.4elisa法检测mcp-1和icam-1的含量
39.将1
×
106/ml rlsecs接种于96孔板,按照2.3项下的分组和实验方法,收集细胞培养液上清,分别按照mcp-1和icam-1elisa试剂盒实验方法,检测mcp-1和icam-1的含量。
40.1.2.5griess法检测no的含量
41.将1
×
106/ml raw264.7接种于96孔板,按照2.3项下的分组和实验方法,收集细胞培养液上清,按照no试剂盒说明书检测540nm处的od值,计算no含量。
42.1.2.6transwell实验检测巨噬细胞的迁移浸润
43.将浓度为1
×
106个ml-1
的rlsecs接种至24孔板内,并置于孵育箱中,待融合度达到90%以上后,更换为不含fbs的dmem培养基饥饿培养24h。然后将细胞分为control组、模型组、梓醇组、梓醇+lv-galectin-3-gfp组、梓醇+lv-galectin-3-shrna组,每组设3个复孔。向24孔板中插入transwell小室,分别给药及慢病毒预孵30min后,在培养基中加入终浓度为200μg/ml的ages刺激。之后另取对数生长期raw264.7以1
×
106个ml-1
接种于5μm的transwell小室中,与rlsecs共培养48h后,使用镊子小心取出小室,吸干小室内液体并将小室浸入多聚甲醛中室温固定30min。之后取出小室,用小棉花团吸干多聚甲醛,加入结晶紫染液室温染色30min。用纯净水洗涤数次,每次5min。取出小室,吸去上室液体,用小棉球小心擦去上室内部膜表面上的细胞,然后将小室置于荧光显微镜下观察。
44.1.2.7western及免疫共沉淀co-ip实验检测cd146、galectin-3的表达及复合物的量
45.取对数生长期的细胞种入六孔板中,每孔1
×
106个细胞,将细胞分为空白组、模型组、梓醇组,每组三个复孔,待六孔板中细胞融合度达到100%时,吸出上清,用1
×
pbs洗两遍,并吸净。六孔板每孔加入蛋白裂解液120μl,冰上裂解30min,以12000r/min离心10min,吸取上清液,加入5
×
上样缓冲液,沸水浴5min。将提取各组蛋白上样,100v恒定电压电泳后转膜,用5%bsa封闭1h后,一抗孵育:cd146(1:1000)、galectin-3(1:1000)、β-actin(1:1000),放置于4℃冰箱过夜。洗膜:用pbst洗膜4次,每次15min。二抗孵育:用pbst稀释山羊抗兔igg抗体(1:5000),稀释山羊抗小鼠igg-hrp抗体(1:5000),室温轻摇孵育1h。用pbst洗膜4次,每次15min。ecl显影液反应1min,曝光。以β-actin为内参对照。用目的蛋白灰度值/β-actin灰度值做统计,将3次实验结果的平均值作为蛋白含量的相对值。
46.按照以上提蛋白方法获取细胞蛋白,取上清液与预孵育蛋白质g-琼脂糖珠(sigma-aldrich)然后离心以获得目标上清液。将上清液与25μl蛋白质g-琼脂糖珠在4℃下共孵育1h,然后离心以去除非特异性粘附在珠上的蛋白质,并获得用于以下ip实验的目标上清液。蛋白g-琼脂糖珠与抗体(兔galectin-3抗体,1:100,;兔cd146抗体,1:100)孵育3-4h。添加抗体结合蛋白g-琼脂糖珠和目标上清液孵育过夜。通过离心分离免疫复合物,用0.05mol/lhepes缓冲液洗涤4次,并在上样缓冲液中通过100℃加热洗脱结合蛋白。通过免疫印迹分析蛋白表达量。
47.1.2.8免疫荧光实验检测cd146、galectin-3的细胞原位表达
48.将浓度为2
×
104个ml-1
的rlsecs接种至24孔板内,细胞贴壁融合后,更换为不含
fbs的dmem培养基饥饿24h。然后将细胞分为空白组、bsa组、模型组、梓醇组,分别给予药物刺激。30min后,用终浓度为200μg/ml的ages刺激。48h后,吸出24孔板上清,用pbs洗涤两遍,并吸净。使用4%多聚甲醛于室温环境下固定15min,用pbs洗涤3遍,每次3min。使用0.1%triton-x室温通透细胞膜20min,洗涤同上用10%的羊血清溶液室温封闭1h。使用羊血清稀释一抗,按照分组加入一抗,4℃过夜。回收一抗,用pbs洗涤3次,每次5min。加入用pbs稀释的cy3-山羊抗小鼠igg,常温避光孵育1h。弃二抗,用pbs避光洗涤3次,每次5min。每孔加入dapi,染核5min。然后弃去dapi,用pbs避光洗涤4次,每次5min。将洗涤完的24孔板荧光显微镜下观察拍照。
49.1.3统计学分析
50.实验所得数据使用graphpad prism 6.0统计软件进行检验分析。所有定量数据均表示
51.为均数
±
标准误采用one-way anova post bonferroni法比较组间差异的显著性,p<0.05表示差异有统计学意义。
52.2.结果
53.2.1lv-galectin-3-gfp的对于肝窦内皮细胞的转染作用
54.由图1可知,lv-galectin-3-gfp转染rlsecs48 h后,荧光显微镜和白光视野下观察细胞gfp高表达。
55.2.2梓醇和ages对于rlsecs活力的影响
56.由表1可知,与空白对照组(0μmol/l)相比,0.1、1和10μmol/l梓醇组的细胞增殖率轻微升高但无显著差异(p>0.05);由表2可知,与孵育0h相比,10μmol/l梓醇孵育48h有显著差异且有一定时间依赖性(p<0.05)。
57.表1不同浓度梓醇对rlsecs细胞活力的影响
[0058][0059]
梓醇不同浓度比较,肩标不同大写字母表示差异显著(p<0.05);肩标相同字母或无字母标注表示差异不显著(p>0.05)。
[0060]
表2 10μmol/l梓醇不同孵育时间对rlsecs细胞活力的影响
[0061][0062]
梓醇不同孵育时间比较,肩标不同大写字母表示差异显著(p<0.05);肩标相同字母或无字母标注表示差异不显著(p>0.05)。
[0063]
2.3梓醇对于ages致rlsecs损伤作用的影响
[0064]
由图2a可知,与control组相比,ages模型组内皮细胞皱缩漂浮细胞较多;与ages组相比,1和10μmol/l梓醇组皱缩漂浮细胞显著减少且与阳性药gb1107组形态类似。由图2b可知,与control组相比,ages组细胞上清液ldh活性明显升高(p<0.01),与ages模型组相比,0.1和1μmol/l梓醇组明显抑制了ldh活性且有一定剂量依赖性(p<0.01),抑制效果与阳性药gb1107组无差异(p>0.05)。
[0065]
2.4梓醇对于rlsecs促炎因子分泌的影响
[0066]
由表3可知,与空白组相比,ages组的mcp-1的分泌水平显著提高(p<0.01);与ages组相比,1和10μmol/l梓醇组明显抑制了单核细胞趋化蛋白-1(mon ocyte chem otact ic pr otein 1,mcp-1)的分泌量并有一定剂量依赖性(p<0.01),且与gb1107组无显著差异(p>0.05);与空白组相比,ages组的细胞间黏附分子-1(intercellular cell adhesion molecule-1,icam-1)的分泌水平显著提高(p<0.01);与ages组相比,1和10μmol/l梓醇组明显抑制了mcp-1的分泌量并有一定剂量依赖性(p<0.01),且与gb1107组无显著差异(p>0.05)。
[0067]
表3梓醇对ages诱导rlsecs分泌促炎因子的影响
[0068][0069]
2.5梓醇对于ages致巨噬细胞激活作用的影响
[0070]
由图3a可知,与空白组相比,ages刺激组的细胞形态改变较为明显,具有促进巨噬细胞活化后伪足形成的作用;与模型组相比,1和10μmol/l梓醇组明显减少了细胞伪足形成且与阳性药gb1107组形态类似。由图3b可知,与空白组相比,模型组ages显著增加巨噬细胞释放no的含量(p<0.01);与ages模型组相比,1和10μmol/l梓醇组明显降低了no的含量(p<0.01),且成一定剂量依赖性,抑制no释放的效果与阳性药组gb1107无显著差异(p>0.05)。
[0071]
2.6梓醇通过galectin-3对于巨噬细胞迁移的影响
[0072]
由图4a和4b可知,与空白组比较,ages刺激组巨噬细胞透膜数量增加(p<0.01);与ages组相比,梓醇组显著减少巨噬细胞的透膜数量(p<0.01);与梓醇组相比,敲减galectin-3组ages+梓醇+lv-gal-3-shrna组透膜率显著升高(p<0.01);与ages组相比,阳性药gb1107组显著减少巨噬细胞的透膜数量(p<0.01),且与梓醇组无显著性差异(p>0.05);与梓醇组相比,过表达galectin-3组梓醇+lv-gal-3-gfp组透膜数量略增加但无差异(p>0.05)。
[0073]
2.7梓醇对ages致rlsecs galectin-3、cd146的共定位的影响
[0074]
由图5可知,与空白组相比,ages组出现了cd146(红色)和galectin-3(绿色)重叠荧光重叠。与ages组相比,梓醇组显著抑制了cd146和galectin-3的复合物的形成,阳性药gb1107显著抑制了galectin-3的表达。
[0075]
2.8梓醇对于ages致galectin-3/cd146复合物表达的影响
[0076]
由图6和表4可知,与control组相比,model组cd146与galectin-3结合显著增多(p<0.01),与model组相比,10μmol/l梓醇组显著抑制cd146与galectin-3的结合(p<0.01);b图显示,与control组相比,model组galectin-3与cd146结合显著增多(p<0.01),与model组相比,10μmol/l梓醇组显著抑制galectin-3与cd146的结合(p>0.05);c图结果显示,model组cd146和galectin-3表达无差异,与model组相比,10μmol/l梓醇组cd146和galectin-3表达无差异(p>0.05)。
[0077]
表4梓醇对ages诱导rlsecsgalectin-3、cd146及galectin-3/cd146复合物的相对表达量
[0078][0079]
综上所述,梓醇可促进rlsecs的增殖,改善ages诱导的rlsecs损伤,降低rlsecs促炎因子的生成,抑制巨噬细胞的激活和no的释放,依赖于galectin-3降低巨噬细胞的迁移浸润数量,并且通过抑制galectin-3和cd146复合物的生成发挥改善内皮炎性损伤的作用。本发明提供了梓醇改善ages沉积致rlsecs炎性损伤的作用机制,为糖尿病肝损伤疾病中药的开发提供有力的理论依据,可用于中药临床防治糖尿病肝损伤疾病的治疗。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1