仿生机器人控制网络的分层构建方法与流程
本发明属于机器人控制技术领域,涉及一种分层的机器人运动控制神经网络构建的方法。
背景技术:
中枢模式发生器(centralpatterngenerator,简称cpg)是生物运动控制神经网络的一个重要组成部分,它可以在没有外部感官信息反馈的情况下产生节律输出。为了利用中枢模式发生器进行仿生机器人的控制。许多研究人员进行了仿生机器人运动控制神经网络构建的研究。这些中枢模式发生器构建方法概括起来可以分为三类:振荡器cpg、生物神经元cpg和连接cpg。其中,振荡器cpg以其结构简单,运算量小等特点被广泛应用到仿生机器人的控制中,然而由于现有的振荡器都不同程度的存在一些缺点,在仿生机器人运动控制神经网络的构建方面还存在较多的困难。在这些振荡器中,matsuoka所提出的由两个具有疲劳特性漏积分器神经元通过相互抑制构成的振荡器成为仿生机器人运动控制神经网络构建中应用比较广泛的振荡器之一。然而,此振荡器在运动控制神经网络构建方面仍然存在较多的问题,如不能包含兴奋性连接、网络拓扑结构构建比较困难等。发明[一种新型神经元振荡器-201510507029.3和仿生机器人运动控制神经网络的构建方法-201510632804.8]提出了一个新的仿生机器人运动控制网络的设计方法,但是此方法针对某种(或某类)步态控制网络的构建具有较好的效果,在实现各种复杂运动步态,如各种节律运动步态、非节律运动步态的仿生机器人整体控制网络的构建方面还非常困难。
技术实现要素:
本发明的目的是针对现有技术的不足,提供一种仿生机器人控制网络的分层构建方法,该方法能够使得仿生机器人产生更加丰富、灵活和符合动物运动特点的运动步态。
本发明所要解决的技术问题是通过以下的技术方案来实现的。本发明是一种仿生机器人控制网络的分层构建方法,其特点是,其步骤如下:
(1)根据速度和运动步态特点,将仿生机器人的运动步态进行分组或分类;
(2)根据某一组或一类运动步态,构建仿生机器人某一层运动控制网络;
(3)将各层运动控制网络组合到一起,构成整个的仿生机器人运动控制网络;利用上层神经中枢产生控制信号,使整个运动控制网络的相应部分激活、失活与饱和,以产生仿生机器人的各种运动步态。
本发明所述的仿生机器人控制网络的分层构建方法,其进一步优选的技术方案是,步骤(2)中,构建的运动控制网络层被分为节律运动控制网络层和非节律运动控制网络层。
本发明所述的仿生机器人控制网络的分层构建方法,其进一步优选的技术方案是,节律运动和非节律运动控制网络层由产生某种运动步态的运动控制网络层构成。
本发明所述的仿生机器人控制网络的分层构建方法,其进一步优选的技术方案是,步骤(3)中,根据需要建立运动控制网络之间的突触连接关系。
本发明所述的仿生机器人控制网络的分层构建方法,其进一步优选的技术方案是,步骤(3)中,所述的中枢为脑干。
本发明所述的仿生机器人控制网络的分层构建方法,其进一步优选的技术方案是,对每一层的运动控制网络的构建方法如下:
该方法以新型神经元振荡器作为基础;
所述的新型神经元振荡器通过建立了一个神经元模型,然后将两个神经元之间通过抑制性突触相互连接,构成一个振荡器模型;所述的神经元模型,在具有疲劳特性的漏积分器神经元模型基础上,增加输出饱和和自兴奋性特性后,形成了一个神经元模型;其中,所述神经元模型的输出,采用非线性函数表示,且该非线性函数满足当x≥θ时,输出具有饱和特性,当x<θ时,神经元没有输出;
所述的运动控制神经网络的构建方法,是利用新型神经元振荡器作为机器人关节的运动控制神经网络,其中一个神经元的输出作为机器人关节的曲肌控制信号,另一个神经元的输出作为关节的伸肌控制信号,然后,根据机器人各关节之间的运动关系和实际仿生生物神经控制环路的拓扑结构特点,利用抑制和兴奋性连接关系建立机器人关节振荡器之间的连接关系。
基于上述仿生机器人运动控制网络层的构建方法,既可以构建节律运动控制网络层也可以构建非节律运动控制网络层,不同之处在于所选取运动控制网络层的参数是不同的。
所述的神经元模型是采用以下两个微分方程组中的一个:
式中,x为神经元的膜电势;y为神经元的输出;s为神经元收到的所有外部输入;a为神经元收到的自我兴奋性反馈的连接权重,a>0;τ跟神经元膜电势相关的时间常数,τ>0;γ跟神经元疲劳过程相关的时间常数,γ>0;x′为反应神经元疲劳程度的变量;b为神经元的疲劳强度,b>0;θ为神经元的输出阈值,
两个神经元之间相互抑制,每个神经元具有一个自我兴奋性连接;具体模型如下:
式中,xi为第i神经元的膜电势;yi为第i神经元的输出;si为第i神经元所收到的外部输入;aij(j∈{1,2},j≠i)为神经元之间的连接权重,aij<0;aii为第i神经元收到的自我兴奋性反馈的连接权重;τi跟第i神经元膜电势相关的时间常数,τi>0;γi跟第i神经元疲劳过程相关的时间常数,γi>0;x′i为反应第i神经元疲劳程度的变量;bi为第i神经元的疲劳强度,bi>0;θi为第i神经元的输出阈值,
所述振荡器平衡状态
振荡器产生振荡输出时,其参数应满足的条件为:
振荡器产生非振荡输出时,其参数应满足的条件为:
(1)
(2)
(3)
或
(1)
(2)
(3)σi≥1,(i=1,2)
根据上述振荡器振荡输出和非振荡输出的条件,振荡器的振荡输出和非振荡输出可以通过调节自兴奋系数aii和疲劳系数bi来进行切换;
所述振荡器当输入满足si<εiθi,(i=1,2)时,振荡器的输出是不激活的,当
其中,所述振荡器的振荡频率和响应速度,通过跟神经元膜电势相关的时间常数τi,i=1,2和跟神经元疲劳过程相关的时间常数γi,i=1,2进行调节;
其中,所述振荡器的饱和输出和不激活以及振荡器节律输出和非节律输出的大小,可以通过振荡器的外部输入si,i=1,2进行调节。
由振荡器相互连接构成的仿生机器人运动控制神经网络的模型如下:
式中,n为神经元数量;xi为第i神经元的膜电势;yi为第i神经元的输出;si为第i神经元所收到的外部输入;aij(j∈{1,…,n},j≠i)为神经元之间的连接权重,aij>0表示兴奋性连接,aij<0表示抑制性连接;aii为第i神经元收到的自我兴奋性反馈的连接权重;τi跟第i神经元膜电势相关的时间常数,τi>0;γi跟第i神经元疲劳过程相关的时间常数,γi>0;x′i为反应第i神经元疲劳程度的变量;bi为第i神经元的疲劳强度,bi>0;θi为第i神经元的输出阈值,
所述仿生机器人运动控制神经网络,当其中存在一部分神经元的平衡状态
其中,ωi={j|aij>0,j∈λ3,j≠i};
这一部分神经元,即i∈λ3的神经元产生振荡输出需要满足的条件如下:
这一部分神经元,即i∈λ3的神经元产生非振荡输出需要满足的条件如下:
(1)
(2)
(3)
或
(1)
(2)
(3)σi≥1,(i∈λ3)
根据上述振荡输出和非振荡输出的条件,这一部分神经元,即i∈λ3的神经元的振荡输出和非振荡输出之间的切换可以通过改变自兴奋系数aii和疲劳系数bi实现,且这部分神经元输出随外部输入si,i∈λ3的变化而变化;
这一部分神经元,即i∈λ3的神经元的振荡频率和响应速度可以通过跟这部分神经元膜电势相关的时间常数τi,i∈λ3和跟神经元疲劳过程相关的时间常数γi,i∈λ3进行调节;
所述仿生机器人运动控制神经网络,当其中一部分神经元的平衡状态
所述仿生机器人运动控制神经网络,当其中一部分神经元的平衡状态
上述结论的推导过程如下:
为了表述方便,假设
令
当神经网络(1)的平衡状态
其中,ωi={j|aij>0,j∈λ3,j≠i};
将运动控制神经网络(1)在平衡状态
其中,
由于矩阵迹与矩阵特征根的和相等。从矩阵a可以看出,当λ3≠φ时,线性化系统(5)存在正实部特征根的充分条件为:
根据李雅谱诺夫定理可知,当式(6)满足时对于任意初始状态,神经网络(1)的平衡状态
由于运动控制神经网络(1)的解是唯一存在且有界的,由此可以得到当运动控制神经网络(1)满足式(6),运动控制神经网络(1)存在振荡输出。
根据gerschgorincircletheorem,对线性化的模型(5)进行分析很容易得到,当运动控制神经网络(1)的参数满足
(1)
(2)
(3)
或
(1)
(2)
(3)σi≥1,(i∈λ3)
时,运动控制神经网络(1)的线性化系统是渐进稳定的。
根据李雅谱诺夫第一定理,对于任意初始状态,运动控制神经网络(1)在平衡状态
本发明提出了一个先分层构建仿生机器人控制网络,然后将其组合构建整个仿生机器人控制网络的方法,解决仿生机器人整体控制网络构建的难题。与现有方法相比,本发明所提出的仿生机器人分层运动控制网络构建方法能够使得仿生机器人产生更加丰富、灵活和符合动物运动特点的运动步态,尤其是既可以产生节律运动也可以产生非节律运动。
附图说明
图1是蝾螈机器人的结构示意图;
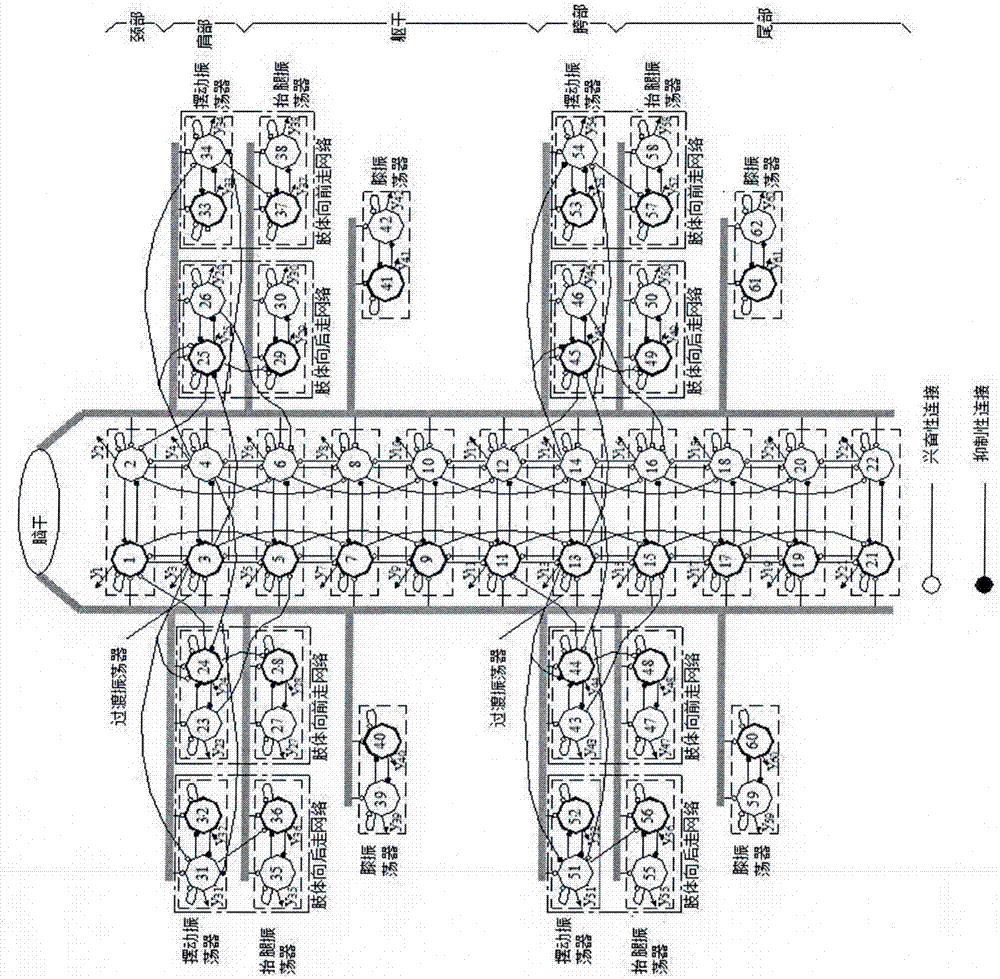
图2是蝾螈机器人低频行走控制网络结构示意图,高频游泳控制网络与低频行走控制网络的身体控制网络部分相同,细圆为伸肌神经元,粗圆为屈肌神经元;
图3蝾螈机器人向前和向后行走及其切换的步态仿真结果图(a)除过渡振荡器外,其余所有身体振荡器输出曲线图(b)蝾螈机器人实体模型一个向前行走周期的运动仿真图(c)除膝振荡器外,所有肢体振荡器的输出曲线图(d)蝾螈机器人实体模型一个向后行走周期的运动仿真图;
图4蝾螈机器人向前行走和游泳及其切换的步态仿真结果图(a)除过渡振荡器外,其余所有身体振荡器输出曲线图(b)蝾螈机器人实体模型一个向前行走周期的运动仿真图(c)除膝振荡器外,所有向前走网络肢体振荡器的输出曲线图(d)蝾螈机器人实体模型一个游泳周期的运动仿真图;
图5随着身体网络左右非对称输入逐渐增加时,蝾螈机器人游泳转弯运动的仿真图(a)输入到颈部和躯干部身体网络的非对称外部输入信号曲线图(b)除过渡振荡器外,其余所有身体振荡器输出曲线图(c)蝾螈机器人实体模型游泳转弯运动的仿真图;
图6蝾螈机器人向前行走转弯步态的仿真图(a)和(b)输入到身体网络和右前肢摆动振荡器的非对称外部输入信号曲线图(c)当非对称输入信号被输入到身体网络和右前肢摆动振荡器时,除过渡振荡器外,其余所有身体振荡器输出曲线图(d)当非对称输入信号被输入到身体网络和右前肢摆动振荡器时,除膝振荡器外,其余所有向前走网络肢体振荡器输出曲线图(e)当非对称输入信号仅被输入到右前肢摆动振荡器时,蝾螈机器人实体模型向前行走转弯仿真图(f)当非对称输入信号仅被输入到身体网络时,蝾螈机器人实体模型向前行走转弯仿真图(g)当非对称输入信号仅被输入到身体网络和右前肢摆动振荡器时,蝾螈机器人实体模型向前行走转弯仿真图;
图7是蝾螈机器人的结构示意图;
图8是蝾螈机器人节律控制网络结构示意图,高频控制网络与低频控制网络的身体控制网络部分相同,细圆为伸肌神经元,粗圆为屈肌神经元;
图9是蝾螈机器人非节律控制网络结构示意图,细圆为伸肌神经元,粗圆为屈肌神经元。
图10是节律和非节律网络协调控制示意图;
图11是蝾螈机器人行走运动仿真结果图;
图12是蝾螈机器人行走和游泳以及步态切换的仿真结果图。
具体实施方式
下面结合附图和具体实施方式对本发明进行详细说明。
具体实施方式:
实施例1,一种仿生机器人控制网络的分层构建方法,其步骤如下:
1)根据速度和运动步态特点等,将仿生机器人的运动步态进行分组或分类;
2)根据某一组(或一类)运动步态,构建仿生机器人某一层运动控制网络;这些运动控制网络层可以被分为节律运动控制网络层和非节律运动控制网络层;节律运动和非节律运动控制网络层又可以由产生某种运动步态的运动控制网络构成;
3)将各层网络组合到一起,如有必要适当建立它们之间的突触连接关系,构成整个的仿生机器人运动控制网络;利用上层神经中枢(如脑干)产生控制信号,使整个运动控制网络的相应部分激活、失活与饱和,以产生仿生机器人的各种运动步态。
实施例2、仿生机器人运动控制网络的分层构建方法实验:
仿生机器人运动控制网络的分层方法对于不同的仿生机器人其分层方法也是不一样的。下面针对不同的动物仿生机器人的举例说明运动控制网络的分层过程:
(1)辐鳍鱼仿生机器人运动控制网络的分层构建
研究表明辐鳍鱼在低速运动时主要采用胸鳍等进行节律推进运动,随着胸鳍拍动频率的增加,辐鳍鱼的游动速度也逐渐增加;当辐鳍鱼游动速度增加到一定程度后,胸鳍靠着身体保持静止,辐鳍鱼主要采用身体和尾鳍节律摆动运动进行高速游泳运动;在静止和起动等运动状态,当辐鳍鱼进行姿态保持和调整以及起动时,其运动还存在较多的非节律运动。因此,可以将辐鳍鱼仿生机器人的运动控制网络分为低速和高速节律游泳运动控制网络层以及非节律运动控制网络层,从而构成整个仿生机器人的运动控制网络。
(2)蝾螈仿生机器人运动控制网络的分层构建
研究表明,蝾螈在陆地上运动时主要通过身体和四肢的相互协调进行低速行走运动;当蝾螈从陆地到水中后,四肢向后靠着身体保持静止,通过身体的摆动运动进行高速运动;蝾螈的运动除了节律运动外还存在较多的非节律运动(如,捕食、监视、原地转弯等),此外,蝾螈在陆地上节律行走的过程同时也需要保持身体具有一定的刚度,以抵抗重力的作用。因此,蝾螈的运动控制网络可以分为低速行走节律运动控制网络层、高速游泳节律运动控制网络层和非节律运动控制网络层,以构成整个蝾螈仿生机器人的运动控制网络。
(3)四足动物仿生机器人运动控制网络的分层构建
四足动物,如马、狗、猫等的运动步态也是随着运动速度的增加逐渐变化的,四足动物的步态随着运动速度从低到高可以被分为:行走、疾走、慢跑和疾驰四类。每一种步态,动物四个肢体的相对运动相位是不同的,需要构造不同结构的运动控制网络来实现,并将它们分别作为四足动物仿生机器人的节律控制网络层。考虑到四足动物的非节律运动以及抵抗重力的作用,建立仿生机器人的非节律运动控制网络层。将所有运动控制网络层组合的到一起,从而构成整个仿生机器人的运动控制网络。
2、蝾螈仿生机器人节律控制网络的分层构建
蝾螈作为生物从水中进化到陆地的一个代表性物种,有关其仿生机器人运动控制网络的构建在各类仿生机器人运动控制网络的构建中是具有重要代表性的。因此,下面以蝾螈仿生机器人运动控制网络的构建来说明本发明关于仿生机器人运动控制网络的分层构建过程。
据蝾螈机器人的结构(如图1所示),构建了一个具有两个相互抑制神经元的控制网络,作为关节控制的振荡器,其中一个神经元的输出作为伸肌控制信号,另一个神经元的输出作为屈肌控制信号。考虑到蝾螈在从陆上行走转换为水中游泳后身体的摆动速度会明显增加的特点,将蝾螈机器人的节律控制网络分为了高频的游泳控制网络和低频陆上行走控制网络分别进行蝾螈机器人的游泳和行走控制。其中,低频控制网络如图2所示,高频控制网络与低频控制网络的身体控制网络部分相同。
(1)低频行走控制网络构建
从图2可以看出,蝾螈机器人的低频控制网络可以被分为身体控制网络和四肢控制网络两部分。蝾螈机器人的身体控制网络包括十一个振荡器,其中每个身体关节由一个振荡器控制。此外,为了协调身体控制网络与四肢控制网络之间的关系,在身体控制网络的肩部和胯部增加了两个过渡振荡器。蝾螈机器人节律控制网络采用了下行占优的兴奋性连接来建立身体振荡器之间的连接关系,如图2所示。蝾螈机器人四肢的控制网络是相似的,每条腿有三个控制振荡器,摆动振荡器控制着腿的前后摆动运动,抬腿振荡器控制着腿的抬起和落下,膝振荡器控制着膝关节的摆动运动。由于蝾螈向前行走的时候,其四条腿是对角线同步的,两个前肢(或两个后肢)之间的摆动运动则是反相的。为了利用身体网络来协调前后肢的运动,实现四肢对角线同步的运动步态,将身体肩部过渡振荡器的屈肌神经元通过抑制性突出被连接到了右前肢摆动振荡器的屈肌神经元,通过兴奋性突触被连接到了左前肢摆动振荡器的屈肌神经元。肩部过渡振荡器的伸肌神经元与前肢之间的连接与屈肌神经元和前肢之间的连接是关于蝾螈的身体纵剖面对称的。此前肢摆动振荡器与身体振荡器之间的连接关系起到了使左右前肢相互反相运动的作用。由于向前行走时蝾螈的身体呈现一个驻波,因此,将前肢摆动振荡器的伸肌神经元通过兴奋性突触被连接到了与其接近的躯干振荡器的身体同侧神经元。利用蝾螈身体呈驻波的特点,同时考虑到蝾螈四肢运动的对角线同步以及左右后肢之间的反相运动,建立了后肢摆动振荡器与身体胯部过渡振荡器之间的连接,此连接关系与前肢摆动振荡器和肩部过渡振荡器之间的连接关系是相似的,仅突触的特性进行了对应的改变。后肢摆动振荡器的伸肌神经元则通过兴奋性突触被连接到了与其接近尾部振荡器的身体同侧神经元,屈肌神经元则通过兴奋性突触被连接到了与其接近躯干振荡器的身体同侧神经元。由于蝾螈腿部向前摆动的过程中,需要抬腿以减小地面摩擦力对腿部向前摆动的影响,腿部向后摆动的时候又需要下压腿部以增加抓地力,因此,将四肢摆动振荡器屈肌神经元的输出通过兴奋性突触连接到了抬腿振荡器的屈肌神经元。
除了向前行走外,蝾螈也可以向后行走,而且其四肢的运动也是对角线同步的。因此,与向前行走相似,蝾螈机器人每个肢体增加了一个摆动振荡器和抬腿振荡器。为了使得肢体摆动振荡器的伸肌神经元比屈肌神经元先激活,来自于身体肩部过渡振荡器同侧神经元兴奋性突触和来自于身体肩部过渡振荡器对侧神经元的抑制性突触被连接到了向后走网络肩关节摆动振荡器的伸肌神经元。来自于身体胯部过渡振荡器对侧神经元兴奋性突触和来自于身体胯部过渡振荡器同侧神经元的抑制性突触被连接到了向后走网络胯关节摆动振荡器的伸肌神经元。此外,对于每个肢体的向后走网络,每个肢体的摆动振荡器的伸肌神经元被连接到了抬腿振荡器的屈肌神经元通过单向的兴奋性突触。在向后行走过程中,当肢体向后摆动的时候,也同时抬起;相反,肢体抓住地面,从而实现向后行走。与蝾螈脑干对脊髓控制网络的控制相似,通过规划蝾螈机器人控制网络的外部输入信号便可产生各种节律运动步态。
(2)高频游泳运动控制网络
高频游泳运动时,蝾螈通过身体的节律摆动运动产生推进力,向前游泳运动。因此,蝾螈的高频游动运动控制网络主要包括身体控制网络部分,其结构与低频行走运动控制网络的身体控制网络部分相同。将上述低频行走运动控制网络层和高频游泳运动控制网络层组合到一起便构成了机器人的整个节律运动控制网络。
(3)蝾螈仿生机器人的步态产生和控制
利用上述运动控制网络,蝾螈仿生机器人可以产生向前行走、向后行走、游泳、转弯运动及其相互转换。当向前行走的时候,高频游泳网络和四肢的向后走网络被失活,在低频行走网络的控制下,蝾螈仿生机器人就产生向前行走。当四肢的向前走网络失活,向后走网络激活后,蝾螈机器人就产生向后行走。当低频行走网络的身体部分失活,四肢的向后走网络失活,向前走网络的伸肌神经元饱和而屈肌神经元失活,从而使得蝾螈机器人四肢向后靠着身体,身体则产生节律摆动实现游泳运动。在游泳运动和行走运动过程中,当非对称输入被施加到蝾螈仿生机器人的身体网络或四肢网络时,蝾螈机器人便可以产生相应的转弯运动。
对于蝾螈机器人的节律运动控制网络,假设所有低频控制网络的振荡器参数都是相同的,并选取振荡器的参数为:τi=0.04、γi=0.044、εi=2、σi=0.1、aii=2.12、bi=0.08、θi=-0.2、
根据设计步骤(2),蝾螈机器人节律控制网络的激活、失活和饱和输出是可以通过改变控制网络的外部输入来实现的。对于游泳、向前行走和向后行走的节律控制网络,当仅仅神经元i处于失活或饱和输出,而其它神经元都处于节律输出时,神经元i失活的外部输入si的上界和饱和的外部输入si的下界如表1、2和3所示。
基于上述运动控制网络参数以及运动步态的产生方法,选取向前行走、向后行走和游泳运动步态时蝾螈机器人运动控制网络的外部输入如表4和5所示。转弯步态运动的外部输入则是在对应向前行走和游泳步态外部输入的基础上增加相应的偏移量。在上述参数和外部输入作用下,蝾螈机器人向前行走、向后行走、游泳、转弯及其相互转换的仿真结果如图3-6。
表1对于向前行走节律控制网络神经元i失活的外部输入si上界和饱和的外部输入si下界
表2对于向后行走节律控制网络神经元i失活的外部输入si上界和饱和的外部输入si下界
表3对于游泳节律控制网络神经元i失活的外部输入si上界和饱和的外部输入si下界
表4向前走和向后走步态时的外部输入
表5向前走和游泳步态时的外部输入
从图3-6可以看出,蝾螈机器人能够产生向前行走、向后行走、游泳、转弯及其相互转换等运动步态,它们是与实际蝾螈的运动步态一致的。
3、包含非节律运动控制网络的蝾螈机器人运动控制网络构建
对于上述实施实例2,为了使上述蝾螈仿生机器人在行走的过程中保持站立且身体具有一定的刚度,使其身体和肢体肌肉具有一定刚度,从而使得蝾螈仿生机器人在上述节律运动控制网络的控制下能够进行节律行走运动。但是,由于蝾螈机器人身体和肢体肌肉的固有刚度并不是可控制的,因此,蝾螈机器人只能保持站立,而不能使蝾螈机器人产生四肢弯曲趴下的步态。
为了适应各种环境,蝾螈的运动是非常灵活、多变的,可以被分为节律运动和非节律运动。蝾螈由趴在地面到起立行走的过程便是由节律和非节律运动组合而成的。蝾螈站立起来的过程一般被看作是非节律运动,此时蝾螈神经控制系统的主要作用是抵抗重力作用,使蝾螈站立起来.蝾螈站立起来后行走的过程一般被认为是节律运动,然而此时神经控制系统的作用则比较复杂,它除了要产生节律控制信号来控制蝾螈行走外,同时还要抵抗重力保持蝾螈呈站立的姿势。此时,如果仅用能单纯产生节律运动的控制神经网络来控制蝾螈的行走运动是非常困难的。为了解决这一问题,本发明将蝾螈机器人的仿脊髓控制网络控制分为节律和非节律控制网络两部分。根据蝾螈随运动速度变化而运动步态不同的特点,蝾螈机器人节律运动控制网络又被分为了低频行走控制网络层和高频游泳控制网络层,然后将各层网络组合构成整个机器人的运动控制网络。在上层神经中枢信号的控制下,蝾螈机器人利用这一运动控制网络,能够实现各种运动步态。每层网络的具体设计过程如下:
(1)节律运动控制网络
根据蝾螈机器人的结构(如图7所示),构建了一个具有两个相互抑制神经元的控制网络,作为关节控制的振荡器,其中一个神经元的输出作为伸肌控制信号,另一个神经元的输出作为屈肌控制信号。考虑到蝾螈在从陆上行走转换为水中游泳后身体的摆动速度会明显增加的特点,将蝾螈机器人的节律控制网络分为了高频的游泳控制网络和低频陆上行走控制网络分别进行蝾螈机器人的游泳和行走控制。其中,低频控制网络如图2所示,高频控制网络与低频控制网络的身体控制网络部分相同。
从图8可以看出,蝾螈机器人的低频控制网络可以被分为身体控制网络和四肢控制网络两部分。蝾螈机器人的身体控制网络包括十一个振荡器,其中每个身体关节由一个振荡器控制。此外,为了协调身体控制网络与四肢控制网络之间的关系,在身体控制网络的肩部和胯部增加了两个过渡振荡器。蝾螈机器人节律控制网络采用了下行占优的兴奋性连接来建立身体振荡器之间的连接关系,如图8所示。蝾螈机器人四肢的控制网络是相似的,每条腿有三个控制振荡器,摆动振荡器控制着腿的前后摆动运动,抬腿振荡器控制着腿的抬起和落下,膝振荡器控制着膝关节的摆动运动。由于蝾螈向前行走的时候,其四条腿是对角线同步的,两个前肢(或两个后肢)之间的摆动运动则是反相的。为了利用身体网络来协调前后肢的运动,实现四肢对角线同步的运动步态,将身体肩部过渡振荡器的屈肌神经元通过抑制性突出被连接到了右前肢摆动振荡器的屈肌神经元,通过兴奋性突触被连接到了左前肢摆动振荡器的屈肌神经元。肩部过渡振荡器的伸肌神经元与前肢之间的连接与屈肌神经元和前肢之间的连接是关于蝾螈的身体纵剖面对称的。此前肢摆动振荡器与身体振荡器之间的连接关系起到了使左右前肢相互反相运动的作用。由于向前行走时蝾螈的身体呈现一个驻波,因此,将前肢摆动振荡器的伸肌神经元通过兴奋性突触被连接到了与其接近的躯干振荡器的身体同侧神经元。利用蝾螈身体呈驻波的特点,同时考虑到蝾螈四肢运动的对角线同步以及左右后肢之间的反相运动,建立了后肢摆动振荡器与身体胯部过渡振荡器之间的连接,此连接关系与前肢摆动振荡器和肩部过渡振荡器之间的连接关系是相似的,仅突触的特性进行了对应的改变。后肢摆动振荡器的伸肌神经元则通过兴奋性突触被连接到了与其接近尾部振荡器的身体同侧神经元,屈肌神经元则通过兴奋性突触被连接到了与其接近躯干振荡器的身体同侧神经元。由于蝾螈腿部向前摆动的过程中,需要抬腿以减小地面摩擦力对腿部向前摆动的影响,腿部向后摆动的时候又需要下压腿部以增加抓地力,因此,将四肢摆动振荡器屈肌神经元的输出通过兴奋性突触连接到了抬腿振荡器的屈肌神经元。蝾螈为了提高行走的速度,有时会在前肢向后摆动的过程中通过前肢膝关节的内收增加向前行走的步长。为此,将前肢摆动振荡器伸肌神经元的输出通过兴奋性突触连接到了膝振荡器的屈肌神经元。与蝾螈脑干对脊髓控制网络的控制相似,通过规划蝾螈机器人控制网络的外部输入信号便可产生各种节律运动步态。
(2)非节律运动控制网络
考虑到蝾螈在非节律运动时关节的屈肌和伸肌的作用也是相互抑制的,因此,利用相互抑制的两神经元网络作为关节的非节律控制网络。由于不影响运动步态的实现,暂不建立关节非节律控制网络之间的连接关系,只为蝾螈机器人每个关节的每个自由度建立了一个两神经元相互抑制控制网络,然后,通过来自脑干的神经控制信号对蝾螈机器人的非节律控制网络进行控制,同时协调节律控制网络实现各种运动步态,具体的结构如图9所示。
(3)节律和非节律网络的协调控制
在不影响蝾螈机器人行走和游泳步态实现的情况下暂不建立节律控制网络和非节律控制网络之间的连接关系。另外,假设节律控制网络和非节律控制网络的关节控制网络的屈肌神经元(或伸肌神经元)的输出经过加和后输出到对应关节的屈肌(或伸肌),如图10所示。在脑干控制信号的作用下节律运动和非节律运动控制网络合作产生蝾螈机器人的起立行走运动步态。
对于蝾螈机器人的节律运动控制网络,假设所有低频控制网络的振荡器参数都是相同的,并选取振荡器的参数为:τi=0.04、γi=0.044、εi=2、σi=0.1、aii=2.12、bi=0.08、θi=-0.2、
根据设计步骤(2),蝾螈机器人节律和非节律控制网络的激活、失活和饱和输出是可以通过改变控制网络的外部输入来实现的。对于游泳和行走的节律控制网络,当仅仅神经元i处于失活或饱和输出,而其它神经元都处于节律输出时,神经元i失活的外部输入si的上界和饱和的外部输入si的下界如表6和7所示。对于非节律控制网络,当仅仅神经元i处于失活或饱和状态,而其它神经元都激活且小于饱和输出时,神经元i失活的外部输入si(i=1,…,42)的上界为-0.4,神经元i饱和的外部输入si(i=1,…,42)的下界为181.2。
表6对于行走节律控制网络神经元i失活的外部输入si上界和饱和的外部输入si下界
表7对于游泳节律控制网络神经元,失活的外部输入si上界和饱和的外部输入si下界
利用蝾螈机器人动力学模型以及所设计运动控制网络,下面对蝾螈机器人的行走和游泳步态的产生以及它们之间相互切换的性能进行了仿真。
(1)陆上行走
蝾螈在陆上行走时,除了利用腿部的支撑抵抗重力保持站立外,身体也会保持一定的刚度,以增强运动的效率。因此,假设利用脑干产生的外部输入信号激活整个蝾螈机器人的非节律运动控制网络。此时由脑干产生的蝾螈机器人非节律控制网络外部输入为si=10,(i=1,…,42)。然后,利用由脑干产生的外部输入信号使高频游泳控制网络保持失活,激活低频的行走节律控制网络,机器人便可以产生节律的直立向前行走运动,其中低频行走节律控制网络的外部输入信号si(i=1,…,46)如表8所示,对应的仿真结果如11图所示。
表8低频行走网络的外部输入
从图11可以看出,蝾螈机器人在行走过程中其身体呈现一个驻波,四肢是对角线同步的。而且,在行走过程中当四肢向前摆动的过程中腿部会向上抬起,而向后摆动的时候会下压,增加抓地力,同时前肢的下臂内收增强蝾螈机器人向前行走的能力。
(2)游泳运动
当蝾螈从陆上行走到水中后,在浮力的作用下其四肢的抗重力作用减弱或消失。为此,当蝾螈游泳后假设蝾螈机器人四肢控制网络的非节律控制网络是失活的。对于蝾螈机器人的身体非节律控制网络,假设其外部输入与陆地上行走时保持相同。从行走切换到游泳运动后,通过设置蝾螈机器人运动控制网络的外部输入,使蝾螈机器人的高频节律运动控制网络被激活,此时高频节律运动网络外部输入si(i=1,…,22)为0.2,低频节律行走网络的身体控制网络被失活。四肢摆动振荡器的伸肌神经元、抬腿振荡器和膝振荡器的屈肌神经元产生饱和输出,对应振荡器的另一个神经元处于失活状态。这些外部输入信号的控制下,蝾螈机器人的四肢向上抬起,并向后紧紧靠着蝾螈机器人身体,以减少对向前游动的阻力,同时,机器人身体产生一个行波,推动蝾螈机器人向前游动。蝾螈机器人行走和游泳运动以及他们之间相互切换的仿真结果如图12所示。
从图12中可以看出,蝾螈机器人首先进行陆上行走运动,10秒时切换为游泳运动。当蝾螈陆上行走时身体和四肢的运动与上述陆上行走运动的仿真结果相似,符合蝾螈陆上行走步态的运动特点。游泳运动时身体呈现一个行波,推动蝾螈机器人向前游动。而且游泳运动的频率高于陆上行走运动的频率,四肢则向上抬起向后靠着身体,以减少阻力。上述仿真结果是与蝾螈运动形态和神经肌肉控制机理的研究结果相一致的。
- 还没有人留言评论。精彩留言会获得点赞!