人制瘤素M抗体及使用方法与流程
本发明涉及能够中和与人细胞上的膜受体结合的制瘤素M产生的生物活性的人抗体和用途。
背景技术:
制瘤素M(OSM)是由单核细胞、巨噬细胞、中性粒细胞和激活的T-淋巴细胞分泌的IL-6家族细胞因子的28kDa多功能成员(Tanaka&Miyajima,RevPhysiolBiochemPharmacol(《生理学、生物化学和药理学综述》)149:39-53,2003)。分泌的OSM的羧基末端附近的蛋白水解裂解产生完全活性形式的OSM,其长度为209个氨基酸,具有两个N-连接的糖基化位点。OSM属于IL-6家族细胞因子,IL-6家族细胞因子包括(IL-6、IL-11、白血病抑制因子(LIF)、心肌营养素-1、睫状神经营养因子(CNTF)和心肌营养素样细胞因子(CLC)),它们共享共同受体亚基gp130蛋白质。在人体中,OSM通过由gp130和LIFRα亚基或gp130和OSMRβ亚基组成的受体异源二聚体传递信号。与其他IL-6家族的其他细胞因子相比,OSM在不存在任何附加膜结合辅助受体的情况下直接结合gp130(Gearing等人,Science(《科学》)255:1434-1437,1992)。在OSM结合到gp130之后,OSMRβ或LIFRα被募集从而形成高亲和力的信号传导复合物(Mosley等人,JBiolChem(《生物化学杂志》)271:32635-32643,1996)。任一受体的激活导致通过JAK/STAT通路的信号传导(Auguste等人,JBiolChem(《生物化学杂志》)272:15760-15764,1997)。OSM主要由免疫系统来源的细胞产生,并且由于其信号传导受体分布广泛,其已与多种生物活性相关,包括细胞生长调节、神经发育和细胞外基质组成的调节。如其名称所暗示,制瘤素M与致癌过程相关。然而,OSM也参与炎症中早期事件和导致有害病症例如肺纤维化的肥厚性通路。因此,需要提供能够阻断受体信号传导(gp130信号传导)事件的对人OSM特异的人抗体,该信号阻断抗体可对gp130表达细胞施加临床可用的毒害细胞、抑制细胞或免疫调节效果。
技术实现要素:
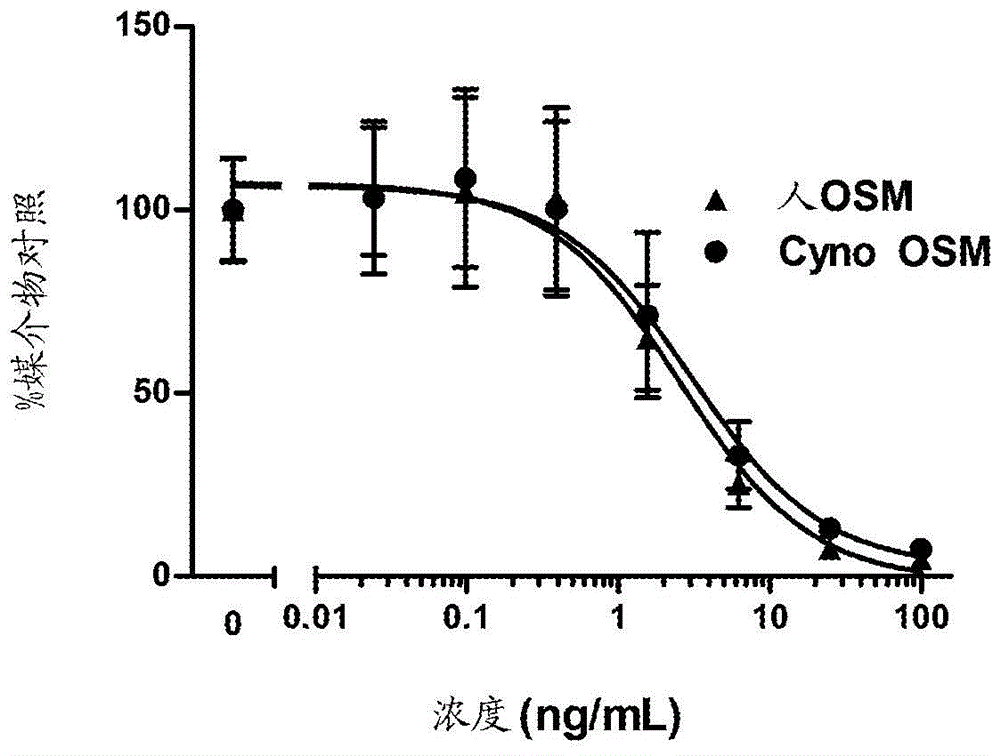
本发明提供能够阻断与一种或多种生物活性相关的活性的OSM结合单克隆抗体,所述生物活性与宿主受试者的细胞、组织或器官上的OSM与OSM结合受体相互作用相关。提供了示例性OSM结合单克隆抗体的氨基酸序列,其可由用于在宿主细胞中表达的核酸编码。本发明的一种或多种OSM单克隆抗体限定OSM表面上的表位,当OSM被本发明的抗体紧密结合时,其被阻止与信号传导复合物的受体组分即gp130和LIFRα或gp130和OSMRβ相互作用,从而阻止配体连接驱动的信号传导和下游生物活性。本发明的一个方面是与人OSM蛋白反应的分离的抗体,所述抗体具有如下所述单克隆抗体的抗原结合能力,该单克隆抗体包含具有如SEQIDNO:13-18单独示出的氨基酸序列或在如SEQIDNO:1-3示出的FR1-CDR1-FR2-CDR2-FR3的指定位置处的抗原结合域,如SEQIDNO:27-29和47表示的CDR3;或包含具有如SEQIDNO:23-26单独示出的氨基酸序列或在如SEQIDNO:5-8及其变体示出的指定位置处的抗原结合域和如SEQIDNO:19-22表示的CDR3。在一个具体实施例中,人OSM结合抗体包含选自SEQIDNO:49-55的可变域。在本发明的另一个实施例中,用作全长IgG结构的单克隆抗体结合域具有衍生自人IgG恒定域或其特定变体的恒定域,并且用作药物制剂中的治疗分子以阻止OSM与展示OSM受体组分的细胞结合。在另一个实施例中,结合域被构造为抗体片段,该抗体片段用作能够阻止OSM与展示OSM受体组分的细胞结合的治疗分子。在本发明的一个方面,提供了治疗制瘤素M相关的病症的药学上可接受的制剂、递送体系、或试剂盒或方法,其包含本发明的一个或多个OSM结合域,诸如但不限于如SEQIDNO:29和47提供的13-28和30-46及变体。附图说明图1示出用于构建展示在pIX外壳蛋白上的Fab文库的种系基因序列,其中每个HV域由根据SEQIDNO:1-3(1=IGHV1-69,2=IGHV3-23,且3=IGHV5-51)的差异化FR1-CDR1-FR2-CDR2-FR3接着可变长度的差异化H-CDR3区以及J区(FR4,SEQIDNO:4)组成;并且每个LV域由根据SEQIDNO:5-8(5=IGKV1-39(O12),6=IGKV3-11(L6),7-IGKV3-20(A27),且8=IGKV4-1(B3))的差异化FR1-CDR1-FR2-CDR2-FR3接着根据SEQIDNO:9的CDR3接着J区(FR4,SEQIDNO:10)组成。图2A为CHO细胞来源的重组人和食蟹猴制瘤素M抑制A375-S2细胞增殖的剂量-反应图,其通过BrdU掺入而测量并归一化成在仅媒介物存在的情况下的对照。图2B为示出由L180(SEQIDNO:53)和H17(SEQIDNO:54)可变域组成的抗体M71减轻OSM对A375-S2增殖的抑制的能力的图,其中OSM以2ng/ml的浓度存在。图3为示出20μg/ml的M64、M71、M55和M69将35SO4-吸收(增加的蛋白聚糖合成的量度)增加至在不存在抗体的情况下观察到的水平以上的效应的柱形图,其中非特异性同种型抗体是藻酸盐珠中的人软骨细胞和能够分泌OSM的人巨噬细胞的共培养物中的对照。图4A为示出在NHLF细胞中人OSM刺激的pSTAT3的剂量反应的图,其中发现EC50为大约1ng/ml。图4B为示出在2ng/mlOSM存在的情况下抗体M71中和pSTAT3信号传导的能力的图。图5A和5B为示出在用OSM刺激之后并且在用或未用指定浓度的M71抗体预处理的情况下在各个小鼠的血清中检测的IP-10(A)和MCP-1(B)的量的散点图。图6A和6B为示出在静脉内(A)或皮下(B)施用3mg/KgM71或M71的Fc变体之后在食蟹猴中随时间推移而变化的血清浓度的图。具体实施方式本说明书中所引用的所有出版物(包括但不限于专利和专利申请)均以引用方式并入本文,如同在本文中完整给出。缩写BSA=牛血清白蛋白;CDR=互补决定区;Cyno=食蟹猴(食蟹猴(Macacafascicularis));DN=糖尿病性肾病;ECD=细胞外域;FR=框架;H=重链;IPF=间质性肺关节炎;L=轻链;Ig=免疫球蛋白;Mab=单克隆抗体;OSM=制瘤素M;OA=骨关节炎;PBS=磷酸盐缓冲盐水;RA=类风湿性关节炎;VL=可变轻链;VH=可变重链。定义如本文所用,“抗体”包括全抗体和任何抗原结合片段或其单链。因此,抗体包括任何包含这样的分子的蛋白或肽,所述分子包含免疫球蛋白分子的至少一部分,诸如但不限于重链或轻链的至少一个互补决定区(CDR)或其配体结合部分、重链或轻链可变区、重链或轻链恒定区、框架区(FR)、或其任何部分,或可整合进本发明的抗体中的结合蛋白的至少一部分。术语“抗体”还旨在涵盖抗体、其消化片段、指定部分或变体,包括抗体模拟物或包含模拟抗体或其指定片段或部分的结构和/或功能的抗体部分,包括单链抗体和单域抗体及其片段。功能片段包括针对预选靶标的抗原结合片段。涵盖于抗体的“抗原结合部分”术语中的结合片段的例子包括(i)Fab片段,即由VL、VH、CL和CH域组成的一价片段;(ii)F(ab′)2片段,即包含通过铰链区二硫键连接的两个Fab片段的二价片段;(iii)由VH和CH域组成的Fd片段;(iv)由抗体的单个支臂的VL和VH域组成的Fv片段,(v)由VH域组成的dAb片段(Ward等人,(1989)Nature341:544-546);以及(vi)分离的互补决定区(CDR)。此外,虽然Fv片段的两个域VL和VH由单独的基因编码,但它们可使用重组方法通过合成的连接肽连接,从而将它们制成单个蛋白质链,所述连接肽使得它们可制成单个蛋白质链,其中VL和VH区配对形成一价分子(称为单链Fv(scFv);参见例如Bird等人(1988)Science(《科学》)242:423-426以及Huston等人(1988)Proc.Natl.AcadSci.USA(《美国科学院院刊》)85:5879-5883)。此类单链抗体也旨在涵盖于抗体的“抗原结合部分”术语中。使用本领域的技术人员已知的常规技术获得这些抗体片段,并筛选出效用方式与完整抗体相同的片段。相反地,可用scFv构造文库筛选抗原结合力,之后使用常规技术将其与编码人种系基因序列的其他DNA拼接在一起。这种文库的一个例子是“HuCAL:HumanCombinatorialAntibodyLibrary(人组合抗体文库)”(Knappik,A.等人,JMolBiol(《分子生物学杂志》)(2000)296(1):57-86)。术语“CDR”是指抗体的互补决定区或高变区氨基酸残基,这些氨基酸残基参与或负责抗原结合。人IgG抗体亚型的高变区或CDR包含来自轻链可变域中的残基24-34(L-CDR1)、50-56(L-CDR2)和89-97(L-CDR3)以及重链可变域中的残基31-35(H-CDR1)、50-65(H-CDR2)和95-102(H-CDR3)的氨基酸残基(如Kabat等人(1991SequencesofProteinsofImmunologicalInterest(1991个免疫学关注的蛋白序列),第5版,PublicHealthService,NationalInstitutesofHealth,Bethesda,Md.)所述)和/或来自轻链可变域中的高变环(即残基26-32(L1)、50-52(L2)和91-96(L3))以及重链可变域中的26-32(H1)、53-55(H2)或当前H2Chothia定义的52-57和96-101(H3)的那些残基(如(Chothia和Lesk,J.Mol.Biol.(《分子生物学杂志》)196:901-917(1987))所述)。Chothia和Lesk将结构保守的HV称为“正则结构”。框架或FR1-4残基是那些可变域残基而不是且排除高变区。Chothia和Lesk的编号体系通过在以小写字母符号表示的指定残基处显示扩展来考虑环中的残基的编号差别,例如30a、30b、30c等。最近,已发展并广泛采用了通用编号体系,即国际免疫遗传学信息体系(internationalImMunoGeneTicsinformation)(IMGT)(LaFranc等人,2005.NuclAcidsRes.(《核酸研究》)33:D593-D597)。本文中,CDR以氨基酸序列以及重链或轻链中的位置通过顺序编号来表示。由于CDR在免疫球蛋白可变域的结构中的“位置”在物种间是保守的且存在于称为环的结构中,因此通过使用根据结构特征对齐可变域序列的编号体系,容易鉴定CDR和框架残基。这种信息用于将来自一个物种的免疫球蛋白的CDR残基移接和置换到来自通常是人抗体的受体框架中。术语“成熟”应用于为了改变多肽性能的目的在抗体可变区进行的定向变化。如本领域已知和本文所述的,V区序列中的很多位置可影响抗原的识别。实际上,抗体通过体细胞突变的渐进过程来获得高亲和力和特异性。可在体外模拟该过程以允许平行选择和靶向变异,同时保持每个抗体链的序列完整性,以使得它们反映物种(在这种情况下是人)抗体,同时增强亲和力或生物物理参数如溶解度或抗氧化性。进行定向变化或“成熟”的过程通常在编码序列水平上进行并可通过制作用于选择增强的性能的子文库来实现。如本文所用,“OSM”是指制瘤素M多肽或包含编码OSM多肽的编码序列的多核苷酸。人OSM是人osm基因(基因5008)的产物。术语“表位”是指能与抗体特异性结合的蛋白决定簇。表位通常由分子的化学活性表面基团例如氨基酸或糖侧链组成,且通常具有特定的三维结构特征以及比电荷特征。构象表位和非构象表位的区别在于与前者而不是后者的结合在变性溶剂存在的情况下会丧失。如本文所用,KD是指解离常数,具体地讲,是指抗体对预定抗原的KD,并且是抗体对特定靶标的亲和力的测量。高亲和力抗体对预定抗原的KD为10-8M或更低,更优选10-9M或更低,并且更优选10-10M或更低。KD的倒数是KA,为缔合常数。如本文所用,术语“kdis”或“k2”或“kd”意指特定抗体-抗原相互作用的解离速率。“KD”是解离速率(k2,也称为“解离率(off-rate)(koff)”)与缔合速率(k1)或“结合率(on-rate)(kon)”的比率。因此,KD等于k2/k1或koff/kon,且以摩尔浓度(M)表示。它遵循下列原则,即KD越小,结合越强。因此与10-9M(或1nM)相比较,10-6M(或1微摩尔)的KD表明为弱结合。如本文所用,术语“单克隆抗体”或“单克隆抗体组合物”是指单分子组合物的抗体分子制剂。单克隆抗体组合物对特定表位呈现出单一的结合特异性和亲和力。该术语也包括“重组抗体”和“重组单克隆抗体”,因为所有抗体都是通过重组方法制备、表达、生产或分离的,例如(a)从动物或杂交瘤分离的抗体,所述抗体通过抗体分泌动物细胞和融合配偶体来制备的;(b)从经转化以表达抗体的宿主细胞分离的抗体,例如从转染瘤分离;(c)从重组的、组合的人或其他物种的抗体文库分离的抗体以及(d)用任何其他方法制备、表达、生产或分离的抗体,所述方法涉及将免疫球蛋白基因序列与其他DNA序列拼接。如本文所用,“分离的抗体”意指这样的抗体:基本上不含有其他具有不同抗原特异性的抗体。然而,与人OSM表位、同种型或变体特异性结合的分离的抗体可对其他相关抗原,例如来自其他物种的抗原(例如OSM物种同源物)具有交叉反应性。此外,分离的抗体可基本上不含其他细胞物质和/或化学物质。在本发明的一个实施例中,具有不同特异性的“分离的”单克隆抗体组合物被混合在成分明确的组合物中。如本文所用,“特异性结合”、“免疫特异性结合”和“免疫特异性地结合”是指抗体与预定抗原的结合。通常,抗体以10-7M或更小的解离常数(KD)结合,并且与预定抗原结合的KD为其与除预定抗原以外的非特异性抗原(例如BSA、酪蛋白或任何其他特定多肽)结合的KD的至少1/2。短语“识别抗原的抗体”和“对抗原特异的抗体”在本文中可与术语“与抗原特异性结合的抗体”互换使用。如本文所用,“高度特异性”结合是指抗体对特定靶表位的相对KD至少小于该抗体与其他配体结合的KD的1/10。如本文所用,“类型”是指由重链恒定区基因编码的抗体种类(例如IgA、IgE、IgM或IgG)。一些抗体种类还涵盖也由重链恒定区编码的亚类或“同种型”(例如IgG1、IgG2、IgG3和IgG4)。抗体可进一步被在恒定区域中的特定残基处与蛋白质连接的寡糖修饰,所述寡糖进一步增强抗体的生物功能。例如,在人抗体同种型中,IgG1、IgG3以及现存较少的lgG2呈现出与鼠lgG2a抗体一样的效应子功能。所谓“效应子”功能或“效应子阳性”是指抗体包含与抗原特异性结合域有所区别的结构域,该结构域能与受体或其他血液组分,例如补体相互作用,导致例如巨噬细胞募集,以及引起由抗体的抗原结合域所结合的细胞破坏的事件。抗体具有通过效应分子的结合而介导的几种效应子功能。例如,补体的C1组分结合抗体会激活补体系统。补体的激活对细胞病原体的调理和溶解是重要的。补体的激活会刺激炎症反应,且还可涉及自身免疫性超敏反应。另外,抗体通过Fc区结合细胞,即抗体Fc区上的Fc受体位点结合细胞上的Fc受体(FcR)。有许多种对不同类别的抗体有特异性的Fc受体,所述抗体包括IgG(γ受体)、IgE(η受体)、IgA(α受体)以及IgM(μ受体)。将抗体结合到细胞表面上的Fc受体会引发许多重要的和各式各样的生物反应,包括吞噬和破坏抗体包裹颗粒,清除免疫复合物,杀伤细胞裂解抗体包裹的靶细胞(被称为抗体依赖性细胞介导的细胞毒作用,或ADCC),释放炎症介质,胎盘转移和调节免疫球蛋白产生。术语“多肽”是指包含由肽键连接形成多肽的氨基酸残基的分子。小于50个氨基酸的小多肽可称为“肽”。多肽也可称为“蛋白质”。综述本发明提供能够结合并中和OSM蛋白质和裂解产物的生物活性的分离的Mab。具体地讲,OSM结合本发明的mAb能阻断OSM与gp130结合或阻止OSM结合的gp130募集LIFRa或OSMRb。不论哪种情况,本发明的OSMmAb能够阻断OSM驱动的gp130受体信号传导。已报道在多种病理环境下STAT3的磷酸化会导致成纤维细胞过度产生胶原(Lim等人,Oncogene(《癌基因》)23(39):5416-25,2006;Huang等人,JCellBiochem(《细胞生物化学杂志》)81(1):102-13,2001)。这些性能展示这些抗体对RA、OA和对纤维化适应症例如特发性肺纤维化(IPF)和糖尿病性肾病(DN)的潜在治疗价值。人OSM基因产物OSM(NCBI登录号NP_065391)是长度为252个氨基酸的pre-pro-多肽(SEQIDNO:11),具有长度为25个氨基酸的信号肽和在残基234和235之间的蛋白水解裂解位点。它是分泌性蛋白,具有在残基31至152和74至192之间形成两个内部二硫键的五个半胱氨酸残基(KallestadJC等人,JBiolChem.(《生物化学杂志》)1991年5月15日;266(14):8940-5)。在残基100和217处存在两个潜在的N-连接糖基化位点,并且当在真核细胞中产生时,蛋白质是糖基化的。人OSM在残基105处具有游离巯基。cynoOSM蛋白的序列在公共领域无法得到,但存在衍生自带注释的基因序列(NW_001095169)的1867bpmRNA(NCBINo.XM_001110148)的自动计算生成的记录。为了获得cynoOSM序列,从cynoPBMC分离RNA,然后通过RT-PCR由该cDNA扩增基因并测序。如申请人的共同待审申请(美国序列号12/648430)中所公开,发现经克隆序列(SEQIDNO:12)的预测翻译与预测的恒河猴(Macacamullata)(Rhesus)序列有99.6%相同,与人OSM蛋白序列有92%相同,并且与小鼠OSM蛋白序列有41%相同。因此,本发明针对能够抑制由OSM结合的gp130信号传导引起的下游生物活性的人源OSM结合Mab的鉴定,并且其中Mab表现出如下能力:在OSM存在的情况下恢复细胞增殖、抑制组织外植体中OSM驱动的关节(关节)间基质的软骨细胞退化、有效地中和人肺成纤维细胞中OSM依赖性STAT3磷酸化以及阻止OSM诱导的细胞因子释放。1.本发明的抗体组合物本发明的OSM中和抗体是这样的抗体,其在体外、原位和/或体内抑制、阻断或干扰至少一种OSM活性或OSM受体结合并且不促进、刺激、诱导、或竞争OSM活性或配体结合,并且抗体结合不模拟OSM驱动的OSM受体连接(具体地讲是gp130与OSM相互作用)的下游效应,例如宿主细胞中的信号转导。合适的OSM中和抗体、指定部分或变体也可任选地影响至少一种OSM活性或功能,例如但不限于:RNA、DNA或蛋白合成;蛋白释放;细胞激活、增殖或分化;抗体分泌;OSM受体信号传导;OSM裂解;OSM结合、OSM或gp130诱导、合成或分泌。本发明基于能够抑制OSM结合或OSM对LIFR的募集之后的gp130信号传导的抗人OSM单克隆抗体的发现。对展示在与pIX外壳蛋白连接的丝状噬菌体颗粒上的Fab文库形式的抗体结合域(参见WO29085462A1并在下文中进一步描述)选择结合OSM的能力。使用gp130的竞争测定用于区分那些Fab,所述Fab在与OSM结合时阻止OSM结合gp130。作为另外一种选择,Fab在与OSM结合时能阻止LIFR募集到gp130结合。使用基于细胞(A375,人黑素瘤细胞)的测定来鉴定多个能够抑制gp130介导的表达OSM的宿主细胞的pSTAT3激活的候选抗体。本文所述的OSM结合抗体识别活性形式的人OSM蛋白上的至少两个不同区,这指示另外发现OSM上多个适合靶向抗体或其他具有类似阻断功能的能力的化合物。因此,本文中作为氨基酸序列提供的抗体结合域的表达和纯化还提供这样的工具,其提供用于选择表现出OSM中和活性的新分子的方法。在一个实施例中,抗人OSM抗体具有结合区,所述结合区包含具有如SEQIDNO:49-55所示的氨基酸序列的轻链可变(VL)或重链可变(VH)区,并且所述抗体或其结合部分免疫特异性地结合OSM。在本发明的另一个实施例中,包含具有SEQIDNO:54或55的重链,并且其抗原结合部分结合OSM蛋白且另外具有本发明抗体的特定功能性能,例如:1.以小于100pM的KD结合人OSM2.以小于500pM的KD结合cynoOSM3.能够在2ng/ml人OSM存在的情况下将A375-S2细胞的增殖恢复不存在OSM的情况下的90%水平,4.能够在2ng/mlcynoOSM存在的情况下将A375-S2细胞的增殖恢复不存在OSM的情况下的90%水平,5.抑制组织外植体中OSM驱动的关节(关节)间基质的软骨细胞退化,6.有效地中和正常人肺成纤维细胞(NHLF)中OSM依赖性STAT3磷酸化,或7.阻断小鼠中用OSM系统性刺激之后的细胞因子释放。在本发明的另一个方面,如本文所述的表现出以上提及的生物活性的一些或全部的抗体的结构特征,具体地讲,定名为M55和M71结合域的Mab被用于制备结构相关的人抗OSM抗体,这些抗体保留本发明抗体的至少一种功能性能,例如与OSM结合。更具体地,M55和M71的一个或多个CDR区(例如SEQIDNO:1和8的特定残基)可与已知的人框架区和CDR例如SEQIDNO:13-28、30-46重组地组合,从而产生本发明另外的重组工程化的人抗OSM抗体。在一个实施例中,本发明的抗体具有如下序列,包括IGVH1-69(SEQIDNO:1)或IGVH5-51(SEQIDNO:3)的FR1、2和/或3,其中来自选自SEQIDNO:13-22的CDR的一个或多个残基存在于SEQIDNO:1或3的CDR位置,同时仍保留抗体结合OSM的能力(例如保守替换)。因此,在另一个实施例中,工程化的抗体可由一个或多个CDR组成,所述CDR例如与SEQIDNO:13-22中列出的CDR或如SEQIDNO:29或47给出的L-CDR3的变体有90%、95%、98%或99.5%相同。除了仅结合OSM,还可对诸如以上所述的那些工程化抗体选择其对本发明抗体的其他功能特性的保留,例如抑制OSM蛋白或其裂解产物与GP130阳性细胞结合的能力,这种结合将导致对体内GP130阳性细胞的增殖的抑制。可通过例如标准ELISA测试本发明的人单克隆抗体与OSM的结合。2.OSM中和抗体的产生可通过多种技术产生表现出所需生物活性谱的OSM中和抗体,如本文中以M5、M6、M9、M10、M42、M45、M53、M54、M55、M62、M63、M65、M66、M67、M68、M69、M71和M83举例说明的,这些例子包含具有文库框架SEQIDNO:1、3、8的指定重链和轻链序列并具有SEQIDNO:13-28、30-46的CDR。在另一个实施例中,由本发明的抗体结合的表位,包含SEQIDNO:11的残基51-227中的最少五个至全部或因此得到的核酸编码序列,可用来免疫受试者,以便在宿主中直接产生本发明的抗体,达到治疗、预防或缓解与OSM的产生相关的疾病或疾病症状的目的。在一个实施例中并如本文所举例说明的,人抗体选自噬菌体文库,其中噬菌体包含人免疫球蛋白基因,并且该文库将人抗体结合域表达为例如单链抗体(scFv),表达为Fab,或一些其他的呈现配对或不配对的抗体可变区的构造(Vaughan等人,NatureBiotechnology(《自然生物技术》)14:309-314(1996);Sheets等人,PITAS(USA)95:6157-6162(1998);Hoogenboom和Winter,J.Mol.Biol.(《分子生物学杂志》),227:381(1991);Marks等人,J.Mol.Biol.(《分子生物学杂志》),222:581(1991))。本发明的人单克隆抗体也可用噬菌体展示法制备,以用来筛选人免疫球蛋白基因的文库。此类用于分离人抗体的噬菌体展示法在本领域是成熟的。参见例如授予Ladner等人的美国专利No.5,223,409;No.5,403,484;和No.5,571,698;授予Dower等人的美国专利No.5,427,908和No.5,580,717;授予McCafferty等人的美国专利No.5,969,108和No.6,172,197;以及授予Griffiths等人的美国专利No.5,885,793;No.6,521,404;No.6,544,731;No.6,555,313;No.6,582,915和No.6,593,081。通过称为生物淘选(biopanning)的多步骤程序选择并鉴定噬菌体克隆。生物淘选这样进行:将展示蛋白配体变体的噬菌体(噬菌体展示文库)与靶标孵育,通过洗涤技术移除未结合的噬菌体,然后特异性地洗脱结合的噬菌体。将洗脱的噬菌体任选地扩增,然后使其通过附加的结合和任选扩增的循环,所述扩增富集特异序列的库,这些特异序列有利于那些载有展示与靶标的最佳结合的抗体片段的噬菌体克隆。在若干轮之后,将各个噬菌体克隆进行表征,并通过确定噬菌体毒粒的相应DNA的序列来确定被克隆展示的肽的序列。Fab噬菌体-pIX文库在噬菌体展示技术的具体实施例中,Shi等人,JMolBiol(《分子生物学杂志》)397:385-396,2010;WO29085462A1和美国序列号12/546850中所述且本文将进一步详述的展示在pIX噬菌体外壳蛋白上的合成Fab文库,被用于从衍生自人种系基因的人IgG序列的所有组成成分中选择连接子。对由已知IGV和IGJ种系序列编码的四个VL和三个VH域构建文库,所述种系序列基于已观察到的这些序列在从天然来源分离的人抗体中存在的频率而选.择。选.择的VH、IMGT命名为IGHV1-69(SEQIDNO:1)、IGHV3-23(SEQIDNO:2)或IGHV5-51(SEQIDNO:3)。VH设计的多样性产生在CDR3区中具有可变长度的序列的重链,其中仍然为恒定长度的H-CDR1和H-CDR2中具有有限多样性的位置。在文库的所有成员中,框架4(H-FR4)保持不变(SEQIDNO:4)。VH169,IGHV1-69*01长度=98,CDR1=31-35,CDR2=50-66QVQLVQSGAEVKKPGSSVKVSCKASGGTFSSYX1ISWVRQAPGQGLEWMGX2IX3X4X5X6GTANYAQKFQGRVTITADESTSTAYMELSSLRSEDTAVYYCAR(SEQIDNO:1)其中,在169文库中,X1为A或G,X2为G或W,X3可为I或S,X4可为P或A,X5可为I或Y并且X6可为F或N。VH323,IGHV3-23*01长度=98,CDR1=31-35,CDR2=50-66EVQLLESGGGLVQPGGSLRLSCAASGFTFSX1YX2MX3WVRQAPGKGLEWVSX4IX5X6X7GX8STYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAK(SEQIDNO:2)其中,在323文库中,X1可为S、D、N或T;X2可为A、G或W;X3可为S或H;X4可为V、A、N或G;X5可为S、N、K或W;X6可为Y、S、G或Q;X7可为S或D;并且X8可为S或G。VH551,IGHV5-51*03长度=98,CDR1=31-35,CDR2=50-66EVQLVQSGAEVKKPGESLKISCKGSGYSFTX1YWIX2WVRQMPGKGLEWMGX3IX4PX5DSX6TRYSPSFQGQVTISADKSISTAYLQWSSLKASDTAMYYCAR(SEQIDNO:3)其中,在551文库中,X1可为S、N或T;X2可为S或G;X3可为I或R;X4可为D或Y;X5可为G或S;X6可为D或Y。具有(11个残基)WGQGTLVTVSS(SEQIDNO:4)的FR4或JH区已与以上序列连接形成完整重链可变区。靶向用于多样化的H-CDR1和H-CDR2位置通过以下来确定:1)种系基因的多样性;和2)在已知结构的抗体-抗原复合物中发现的与抗原接触的频率(Almagro,JMolRecognit.(《分子识别杂志》)17:132-143,2004)。选择的位置处的氨基酸多样性通过以下来确定:1)在种系中的用途;2)在人重排V基因中最常被观察到的氨基酸;3)预测将来自单碱基体细胞突变的氨基酸;和4)有助于抗原识别的氨基酸的生物化学和生物物理特性。文库整合VH(H3)的CDR3的多样性,模拟人抗体的所有组成成分(Shi等人,2010,同上),如下所示(式I),其中最终长度介于7和14个残基之间。在超过5000个人可变区的CDR3中,氨基酸甘氨酸(G)和丙氨酸(A)经常在所有位置中使用。此外,天冬氨酸(D)经常在第95位中使用,酪氨酸(Y)经常被编码于在J区段的规范区之前的位置。氨基酸苯丙氨酸(F)、天冬氨酸(D)和酪氨酸(Y)在第99-101位占优势,它们用于IgG的这些位置。由于这些位置通常充当H-CDR3的结构支撑且不那么容易被抗原和/或IgG的表面接近,所以固定第99位的氨基酸苯丙氨酸加亮氨酸(50/50比率)、第100位的天冬氨酸以及第101位的酪氨酸。因此,式I的序列插入在SEQIDNO:1、2或3和SEQIDNO:4之间形成完整VH。-(D)-(N)n(N+O)m(F)DY-(I)其中:(D)=富含Asp(D)和Gly(G)的位置。(N)n=富含Ala(A)和Gly(G)的位置,n=3-7。(O)m=富含Ala(A)、Gly(G)和Y(Tyr),m=1-4。(F)=Phe(F)优势位置。多种形式的文库涵盖与也衍生自人种系所有组成成分的固定或多样化的轻链配对。在本发明中,四个轻链文库VLκ基因(Kawasaki等人,2001.EurJImmunol(《欧洲免疫学杂志》)31:1017-1028以及Schaeble&Zachau,1993BiolChemHoppeSeyler(《HoppeSeyler生物化学》)374:1001-1022)为A27(IGKV3-20*01)、B3(IGKV4-1*01)、L6(IGKV3-11*01)和O12(IGKV1-39*01),其中括号中的基因名称为推测的相应IMGT基因。Fab通过双顺反子载体的表达展示在pIX上,其中VH-CH1域与外壳蛋白序列融合,并且VL-CLκ或VL-CLλ表达为与VH-CH1自缔合的游离多肽。CDR区以下划线表示。基于Vkappa(Vk)种系基因的轻链可变文库012;IGKV1-39*01,IGKV1D-39*01长度=88;CDR1=24-34,CDR2=50-56DIQMTQSPSSLSASVGDRVTITCRASQSISX1X2X3NWYQQKPGKAPKLLIYX4ASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYC(SEQIDNO:5)L6,IGKV3-11*01长度=88,CDR1=24-34,CDR2=50-56EIVLTQSPATLSLSPGERATLSCRASQSVX1X2X3LAWYQQKPGQAPRLLIYX4ASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYC(SEQIDNO:6)A27,IGKV3-20*01,长度=89,CDR1=24-35,CDR2=51-57EIVLTQSPGTLSLSPGERATLSCRASQSVX1X2X3X4LAWYQQKPGQAPRLLIYX5ASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYC(SEQIDNO:7)B3,DPK24,VKIVKlobeck;IGKV4-1*01长度=94,CDR1=24-40,CDR2=56-62DIVMTQSPDSLAVSLGERATINCKSSQSVLX1SSNNX2NX3LAWYQQKPGQPPKLLIYX4ASTRESGVPDRFSGSGSGTDFTLTISSLQAEDVAVYYC(SEQIDNO:8)每个可变区支架的特定位置处的多样性汇总于下表1中,其中氨基酸单字母代码被采用并交替出现在如图1所示的特定位置处。表1:所有文库中的VLCDR3具有七个残基,其中前两个残基为谷氨酰胺(Gln,Q)并且对应于Kabat残基95的残基是脯氨酸(Pro,P)。对于L-CDR3,所述序列符合QQX1X2X3X4PX5T(SEQIDNO:9),其中变异与下表中一样,残基位置根据Kabat。表2:由于基因间可变序列长度不同,所以可使用如Al-LazikaniB、LeskAM、ChothiaC,1997(Standardconformationsforthecanonicalstructuresofimmunoglobulins.(免疫球蛋白的规范结构的标准构象)JMolBiol(《分子生物学杂志》)273:927-948)中所定义的残基编号将高变环或CDR中的特定残基位置的多样性描述如下。在该体系中,通过对给定残基指定子位置a、b、c等来适应高变环的长度变化。使用框架4(FR4)区段例如JK4,FGQGTKVEIK(SEQIDNO:10)来形成完整人轻链可变区。在初始选择中已显示Fab对0.2至20nM的多种蛋白靶标的亲和力。用于改善结合参数的综合成熟过程的方法由改组VL或VH多样性或任选地定向或有限VL修饰组成,所述方法使用设计并用于如引用的出版物中所述、如本文中所教导的文库的载体和引物完成,并且与本领域已知的方法组合。OSM结合免疫球蛋白域的替代来源可通过多种方法来制备具有本文公开的人Mab的特征的OSM结合抗体或形成来源于免疫球蛋白域的结合片段,所述方法包括Kohler和Milstein(1975)Nature(《自然》)256:495的标准体细胞杂交技术(杂交瘤方法)。在杂交瘤方法中,如本文所述免疫小鼠或其他合适的宿主动物,例如仓鼠或猕猴,以引起淋巴细胞,所述淋巴细胞产生或能产生将特异性地结合到用于免疫的蛋白的抗体。或者,可将淋巴细胞在体外免疫。然后用适合的融合剂,例如聚乙二醇将淋巴细胞与骨髓瘤细胞融合,以形成杂交瘤细胞(Goding,MonoclonalAntibodies:PrinciplesandPractice(《单克隆抗体:理论与实践》),第59-103页(AcademicPress,1986))。如本文所述和/或本领域中已知的,还可以通过对能产生全套人抗体的转基因动物(如小鼠、大鼠、仓鼠、非人灵长类动物等等)进行免疫来任选地产生OSM中和抗体。可使用合适的方法,例如本文描述的方法,从这些动物分离产生人抗OSM的抗体的细胞并使其无限增殖化。作为另外一种选择,可将抗体编码序列克隆,导入合适的载体,并用来转染宿主细胞,从而通过本文教导的方法和本领域所知的那些方法表达和分离抗体。使用在其种系基因结构中载有人免疫球蛋白(Ig)基因座的转基因小鼠来提供高亲和力的完全人单克隆抗体的分离,该抗体针对多种靶标,包括正常人免疫系统对其具有耐受性的人自身抗原(Lonberg,N.等人,US5569825、US6300129和1994,Nature(《自然》)368:856-9;Green,L.等人,1994,NatureGenet.(《自然·遗传学》)7:13-21;Green,L.&Jakobovits,1998,Exp.Med.(《实验医学》)188:483-95;Lonberg,N和Huszar,D.,1995,Int.Rev.Immunol.(《国际免疫学综述》)13:65-93;Kucherlapati等人,US6713610;Bruggemann,M.等人,1991,Eur.J.Immunol.(《欧洲免疫学杂志》)21:1323-1326;Fishwild,D.等人,1996,Nat.Biotechnol.(《自然·生物技术》)14:845-851;Mendez,M.等人,1997,Nat.Genet.(《自然·遗传学》)15:146-156;Green,L.,1999,J.Immunol.Methods(《免疫学方法杂志》)231:11-23;Yang,X.等人,1999,CancerRes.(《癌症研究》)59:1236-1243;Brüggemann,M.和Taussig,MJ.,Curr.Opin.Biotechnol.(《生物技术当前述评》)8:455-458,1997;Tomizuka等人,WO02043478)。此类小鼠中的内源免疫球蛋白基因座可以被破坏或缺失,以消除动物产生由内源基因编码的抗体的能力。此外,诸如Abgenix,Inc.(Freemont,Calif.)和Medarex(SanJose,Calif.)之类的公司可从事使用如上所述的技术提供针对所选抗原的人抗体。可使用任何合适的技术例如重组蛋白制备技术来制备用作淘选策略中的靶配体和用作免疫原性抗原的多肽。呈纯化蛋白或蛋白混合物(包括全细胞或细胞提取物或组织提取物)形式的靶配体或其片段,或在免疫的情况下抗原,可在动物体内由编码所述抗原或所述抗原的一部分的核酸从头形成。本发明的分离的核酸可用本领域熟知的(a)重组方法、(b)合成技术、(c)纯化技术或它们的组合进行制备。编码单克隆抗体的DNA易于使用本领域已知的方法来分离和测序,所述已知的方法为例如通过使用能特异性地与编码人抗体区的重链和轻链的基因结合的寡核苷酸探针。在杂交瘤生成的情况下,此类细胞可用作此类DNA的来源。作为另外一种选择,使用其中编码序列和翻译产物相关联的展示技术,例如噬菌体或核糖体展示文库,简化了连接子和核酸的选择。噬菌体选择后,可分离来自噬菌体的抗体编码区,并用来产生完整抗体,包括人抗体或任何其他所需的抗原结合片段,并且表达在任何所需的宿主内,所述宿主包括哺乳动物细胞、昆虫细胞、植物细胞、酵母和细菌。人抗体本发明还提供结合人OSM的人免疫球蛋白(或抗体)。这些抗体也可表征为工程化或适应获得的。所述免疫球蛋白具有基本上来自人种系免疫球蛋白的一个或多个可变区并且包括已知参与抗原识别的残基的定向变异,所述残基例如CDR或结构上限定的高变环的残基。一个或多个恒定区,如果存在,也基本上来自人免疫球蛋白。人抗体对OSM表现出的KD为至少约10-6M(1μM)、约10-7M(100nM)、10-9M(1nM),或更小。为了影响亲和力的改变,例如改善人抗体对OSM的亲和力或降低人抗体对OSM的KD,可以对CDR残基或其他残基进行替换。用于产生结合OSM的人抗体的来源优选为本文提供的序列,这些序列为可变区、框架和/或CDR,标记为SEQIDNO:13-55,使用展示在丝状噬菌体颗粒上的全套人源Fab鉴定为能够结合人OSM并与食蟹猴OSM交叉反应。如果人可变域框架采取与CDR从其产生的亲本可变框架相同或相似的构象,则将CDR中的任何者替换进入任何人可变域框架很可能导致保持其正确的空间取向。在最终Mab中将配对的重链和轻链可变框架区可以衍生自相同或不同人抗体序列。人抗体序列可为天然存在的人抗体序列,衍生自人种系免疫球蛋白序列,或者可为几个人抗体和/或种系序列的共有序列。合适的人抗体序列可通过计算机比较小鼠可变区的氨基酸序列和已知人抗体的序列来进行鉴定。分别进行重链和轻链的比较,但是每种比较的原则是相同的。就实验方法而言,已发现它特别便于构建可变序列文库,可以根据所需的活性、结合亲和力或特异性来对此可变序列文库进行筛选。用于构建此类变体文库的一种形式为噬菌体展示载体。作为另外一种选择,可使用用于使编码可变域中靶残基的核酸序列随机化或多样化的替代已知方法来产生变体。确定是否需要进一步取代以及选择用于取代的氨基酸残基的另一种方法,可使用计算机建模来完成。用来产生免疫球蛋白分子三维图像的计算机硬件和软件可广泛获得。通常,分子模型的生成起始于免疫球蛋白链或其结构域的已解算的结构。将要建模的链与已解算的三维结构的链或结构域进行氨基酸序列相似性的比较,并且选择表现出最高序列相似性的链或结构域作为分子模型构建的起始点。对已解算的起始结构进行修饰,以允许被建模的免疫球蛋白链或结构域中的实际氨基酸与起始结构中的那些实际氨基酸之间的不同。然后将修饰的结构组装进复合免疫球蛋白中。最后,通过能量最小化以及通过证实所有原子彼此处于合适的距离且键长和键角在化学可接受限度内来改良模型。由于密码的简并性,多种核酸序列将编码每个免疫球蛋白氨基酸序列。所需的核酸序列可通过固相DNA从头合成或通过早期制备的所需多核苷酸的变体的PCR诱变来生成。在本申请中描述的所有编码抗体的核酸明确地包括在本发明中。如本文所述产生的人抗体的可变区段通常连接到人免疫球蛋白恒定区的至少一部分。所述抗体将包含轻链和重链恒定区。重链恒定区通常包含CH1、铰链、CH2、CH3结构域,并且有时还包含CH4结构域。所述人抗体可包含来自任何抗体种类,包括IgM、IgG、IgD、IgA和IgE,和来自任何亚类(同种型),包括IgG1、IgG2、IgG3和IgG4的任何类型恒定区。当需要人源化抗体呈现细胞毒活性时,恒定区通常为补体结合性恒定区,并且种类通常为IgG1。当不需要此类细胞毒活性时,恒定区可以是IgG2类。人源化抗体可包含来自不止一个种类或同种型的序列。将可任选地与恒定区连接的编码人源化轻链可变区和重链可变区的核酸插入表达载体中。轻链和重链可克隆在相同或不同的表达载体中。将编码免疫球蛋白链的DNA区段可操作地连接到确保免疫球蛋白多肽表达的一个或多个表达载体中的对照序列。此对照序列包括信号序列、启动子、增强子和转录终止序列(参见Queen等人,Proc.Natl.Acad.Sci.(《美国科学院院刊》)USA86,10029(1989);WO90/07861;Co等人,J.Immunol.(《免疫学杂志》)148,1149(1992),所述文献全文以引用方式并入本文中用于所有目的)。3.使用抗OSM的抗体的方法如以下详细描述的,本发明显示具有M5、M6、M9、M10、M42、M45、M53、M54、M55、M62、M63、M65、M66、M67、M68、M69、M71和M83的可变域的分离的单克隆抗体结合OSM上的重叠表位并且展示体外和/或体外OSM抑制活性。重要的是,选择的MAb的反应性包括如下能力:剂量依赖性地阻断OSM与gp130的相互作用、降低在gp130存在的情况下的OSM信号传导、降低OSM刺激的A375细胞增殖、阻止巨噬细胞刺激的软骨细胞胶原生成、或降低体内OSM导致的细胞因子释放。考虑到本发明所述的单克隆抗体的特性,抗体或其抗原结合片段适合作为治疗剂和预防剂来治疗或预防人和动物中OSM相关的病症。一般来讲,使用将包括将治疗或预防有效量的一种或多种本发明的单克隆抗体或抗原结合片段,或选择的具有类似结合谱和生物活性的抗体或分子施用给敏感受试者或表现出其中已知OSM活性具有病理后果的病症(例如免疫疾患或肿瘤生长和癌细胞转移)的受试者。可以施用所述抗体的任何活性形式,包括Fab和F(ab′)2片段。优选地,所用抗体与受者物种相容,使得在受试者中对MAb的免疫反应不会造成不能接受的短循环半衰期或引起对MAb的免疫反应。施用的MAb可表现出一些辅助功能,例如与受试者的Fc受体结合和激活ADCC机制,以便使用细胞溶解或细胞毒机制排除靶细胞群,或可将它们进行工程化以限制或消除这些辅助功能,从而保留靶细胞群。个体治疗可包括施用治疗有效量的本发明的抗体。抗体可以如下所述的试剂盒提供。抗体可以例如相等量的混合物形式使用或施用,或可以单独地按序提供或一起施用。在向患者提供在受试患者中能结合OSM的抗体或其片段或能阻断OSM的抗体时,施用剂的剂量会根据诸如患者的年龄、体重、身高、性别、一般身体状况、既往病史等因素而有所不同。在一个相似的方法中,本发明的单克隆抗体的另一个治疗用途为,使用针对本发明的一个单克隆抗体培育的抗独特型抗体来主动免疫患者。用模拟表位结构的抗独特型进行免疫可引起主动抗OSM反应(Linthicum,D.S.和Farid,N.R.,Anti-idiotypes,Receptors,andMolecularMimicry(抗独特型、受体和分子模拟)(1988),第1-5和285-300页)。同样,可通过施用作为疫苗组分的一个或多个抗原表位和/或致免疫表位来引起主动免疫。可口服或非肠道服用一定量的疫苗,该量足够在预防性或治疗上实现受试者产生对抗此生物功能区的保护性抗体。可用纯化形式的抗原肽/致免疫肽、肽的片段或肽的修饰形式主动免疫宿主。可将不对应初始蛋白序列的一个或多个氨基酸加入初始肽或截短形式的肽的氨基端或羧基端。此类额外的氨基酸可用于将该肽偶联到另一个肽、偶联到大载体蛋白或偶联到支撑物。可用于这些目的的氨基酸包括:酪氨酸、赖氨酸、谷氨酸、天冬氨酸、半胱氨酸以及它们的衍生物。可使用替代的蛋白修饰技术,例如NH2端乙酰化或COOH端酰胺化,以提供另外的将该肽偶联或融合到另一个蛋白或肽分子或支撑物的方法。旨在将能够阻断不需要的OSM生物活性的抗体以足够实现减轻、治愈或缓解OSM相关症状或病变的量提供给受试者。如果试剂的剂量、给药途径等足以影响此类反应,则认为该量为“实现”症状减轻的足够量或“治疗有效量”。可通过分析受试者的受影响的组织、器官或细胞,如通过成像技术、或通过体外分析组织样品来测定对抗体施用的反应。如果试剂的存在导致受试患者的生理出现可检测的改变,那么该试剂为生理上显著的。治疗应用可使用本发明的OSM中和抗体、其抗原结合片段或特定变体在细胞、组织、器官或动物(包括哺乳动物和人)中测量或引发效应,以诊断、监测、调节、治疗、缓解、帮助预防由OSM或表达OSM的细胞介导、影响或调节的病症的发病率或减轻该病症的症状。因此,本发明提供了使用至少一种本发明的OSM抗体调节或治疗细胞、组织、器官、动物或患者中的至少一种OSM相关疾病的方法,如本领域已知的或如本文所述的。已知在多种涉及炎症的疾病状态中OSM被上调,并且OSM已牵连多种生物作用,包括骨形成、软骨退化、胆固醇吸收、疼痛和炎症。具体适应症在下文讨论。适应症本发明人已证明OSM介导软骨破坏,并且已显示OSM导致来自组织外植体的关节间基质中的软骨细胞退化。OSM也促进细胞因子释放,例如TNFα,其能促进软骨释放胶原,如T.Cawston等人(1998,ArthritisandRheumatism(《关节炎与风湿病》),41(10)1760-1771)和以下事实所示,所述事实是:目前举例为M71结合域的抗体能够阻断系统性细胞因子释放。本发明的抗体M55、M64、M69和M71显示出如下能力:将巨噬细胞-软骨细胞共培养体系中的蛋白聚糖合成增加到不存在OSM特异性抗体的情况下所见水平以上。本发明人已进一步证明,施用本发明的抗OSM的中和抗体将抑制体内OSM驱动的细胞因子和趋化因子释放,例如IL-6、IP-10和KC。IP-10即干扰素γ诱导蛋白10kDa或可诱导的小细胞因子B10是在人中由CXCL10基因(C-X-C基序趋化因子10(CXCL10))编码的蛋白。多种作用已归因于CXCL10,例如单核细胞/巨噬细胞、T细胞、NK细胞和树突细胞的化学趋向性以及促进T细胞粘附内皮细胞。KC,现在称为趋化因子(C-X-C基序)配体1(CXCL1)是属于CXC趋化因子家族的小细胞因子,其之前被称为GRO1癌基因、GROα、中性粒激活蛋白3(NAP-3)和黑素瘤生长刺激活性α(MSGA-α)。在人中,该蛋白由CXCL1基因编码。CXCL1由巨噬细胞、中性粒细胞和上皮细胞表达,并具有中性粒细胞趋化活性。根据本发明,因此提供了使用选自M5、M6、M9、M10、M42、M45、M53、M54、M55、M62、M63、M65、M66、M67、M68、M69、M71和M83的抗体或抗体片段来制造用于治疗或预防关节蛋白聚糖退化疾病例如骨关节炎、炎性关节病或炎性疾患的药物。OSM的拮抗剂的一个具体用途是用来制造用于阻止或降低软骨释放胶原的药物。本发明还提供用于治疗或预防炎性关节病或炎性疾患的方法,所述方法包括给患有此类疾患的患者施用有效量的阻断OSM与gp130结合的此类抗体。本发明的抗体可用于治疗促炎过程的制剂中,在促炎过程中,OSM直接或间接地(例如通过释放炎性细胞因子)导致组织或器官尤其是皮肤、肺和关节发病。此类病状包括骨关节炎、类风湿性关节炎、银屑病性关节炎、强直性脊柱炎、神经性关节病、反应性关节炎、肩袖撕裂关节病、风湿热、赖特尔综合征、进行性系统性硬化、原发胜胆汁性肝硬化、天疱疮、类天疱疮、坏死性血管炎、重症肌无力、多发性硬化症、红斑狼疮、多发性肌炎、肉样瘤病、肉芽肿病、血管炎、恶性贫血、CNS炎性疾患、抗原-抗体复合物介导的疾病、自身免疫性溶血性贫血、桥本氏甲状腺炎、格雷夫斯病、雷诺综合症、肾小球肾炎、皮肌炎、慢性活动性肝炎、乳糜泻、AIDS的自身免疫性并发症、萎缩性胃炎、和阿狄森氏病、内毒素血症或败血性休克(败血病)、或败血病症状的一种或多种以及其他类型的急性和慢性炎症。尤其能从本发明的方法受益的那些患者是患有由大肠杆菌(E.coli)、乙型流感嗜血杆菌(HaemophilusinfluenzaB)、脑膜炎奈瑟氏菌(Neisseriameningitides)、葡萄球菌或肺炎球菌导致的感染的那些患者。具有败血病风险的患者包括遭受烧伤、伤口、肾或肝功能衰竭、创伤、烧伤、免疫功能不全(HIV)、造血瘤、多发性骨髓瘤、Castleman病或心脏粘液瘤的那些患者。其他与OSM相关并可接受用本发明的抗体的治疗或预防疗法病症包括纤维化疾病例如肺纤维化、糖尿病性肾病、特发性肺纤维化、系统性硬化、以及肝硬化。使用本发明的抗体治疗或预防的另一种适应症是涉及背根神经节的神经元的伤害性疼痛。施用和给药本发明提供优选为含水磷酸盐缓冲盐水或混合盐溶液的OSM中和抗体的稳定制剂,以及保存溶液和制剂以及适用于药物和兽用药用途的多用途保存制剂,所述制剂包含于药学上可接受的制剂中的至少一种OSM中和抗体。合适的媒介物及其制剂,包含其他人蛋白如人血清白蛋白在内,描述于例如Remington:TheScienceandPracticeofPharmacy(制药科学与实践),第21版,Troy,D.B.编辑,LipincottWilliams和Wilkins,Philadelphia,PA2006,第5部分,PharmaceuticalManufacturing(《药物制造》)第691-1092页,特别参见第958-989页。在本文所述的稳定或保存制剂或溶液中的OSM中和抗体可以根据本发明通过多种递送方法施用给患者,所述递送方法包括静脉内(I.V.);肌肉内(I.M.);皮下(S.C.);经皮;经肺;透粘膜;使用在植入物、渗透泵、药筒、微型泵中的制剂;或技术人员理解、本领域熟知的其他方式。例如,可通过下列位点特异性施用到体室或体腔中:关节内、支气管内、腹内、囊内、软骨内、腔内、体腔内、小脑内、脑室内、结肠内、颈内、胃内、肝内、心肌内、骨内、骨盆内、心包内、腹膜内、胸膜内、前列腺内、肺内、直肠内、肾内、视网膜内、脊柱内、滑膜内、胸内、子宫内、膀胱内、病灶内、阴道、直肠、口腔、舌下、鼻内或经皮方式。通常,如果施用全身剂量的抗体,希望提供给受者剂量在约1ng/kg-100ng/kg、100ng/kg-500ng/kg、500ng/kg-1μg/kg、1μg/kg-100μg/kg、100μg/kg-500μg/kg、500μg/kg-1mg/kg、1mg/kg-50mg/kg、50mg/kg-100mg/kg、100mg/kg-500mg/kg(受者的体重)范围内的剂量,但也可施用较低或较高的剂量。当然,本发明的拮抗剂的合适剂量将根据诸如待治疗的疾病或疾患、给药途径和待治疗的个体的年龄和体重以及拮抗剂的性质等因素而有所不同。虽然不受任何具体剂量的束缚,但据信例如对于非肠道给药,0.01至20mg/kg的本发明的抗体(或其他大分子)的日剂量(通常以如上文所述的药物组合物的部分存在)可适于治疗典型成人。治疗可以单剂量疗法进行,或优选地以多剂量疗法进行,在多剂量疗法中,最初治疗过程可以为给予1-10次单独的剂量,接着为了维持和/或增强反应,有必要在后续的时间间隔给予其他剂量,例如在1-4个月时给予第二种剂量,并且如果需要,在几个月后给予一种或多种后续的剂量。合适的治疗疗程的例子包括:(i)0、1个月和6个月,(ii)0、7天和1个月,(iii)0和1个月,(iv)0和6个月,或其他足以引起预期会减轻疾病症状或减轻疾病严重程度的所需反应的疗程。本发明的抗体可单独使用,或与免疫抑制剂例如类固醇(泼尼松等)、环磷酰胺、环孢菌素A或嘌呤类似物(例如甲氨蝶呤、6-巯嘌呤等)、或抗体例如抗-淋巴细胞抗原抗体、抗-白细胞抗原抗体、TNF拮抗剂如抗-TNF抗体或TNF抑制剂如可溶性TNF受体、或试剂例如NSAID或其他细胞因子抑制剂组合使用。序列表本发明现结合以下具体的、非限制性实例进行描述。实例1:试剂和测定法为了选择和表征OSM结合抗体,形成人和CynoOSM的构建体以用于哺乳动物细胞表达。人OSM(NP_065391,由NM_020530编码)为252个氨基酸前体,其加工成227个氨基酸的全长分泌性蛋白(SEQIDNO:11),该分泌性蛋白为前蛋白,其进一步加工成更完全活性的成熟形式,即氨基酸1-184。人OSMcDNA从OriGene(Cat.No.SC121421)订购,将来自OriGene克隆的人OSM的ORF通过PCR扩增,并将信号肽(鼠IgG1)与用于蛋白纯化的六-His标签和用于定点蛋白生物素酰化的Avi标签(SEQIDNO:56)一起引入。选择后者是为了避免存在于与受体相互作用的OSM附近的赖氨酸残基的随机化学生物素酰化。使用SuperscriptIII第一链合成体系(InVitrogen)从cynoPBMC’sRNA克隆食蟹猴OSM以获得cDNA,然后使用由人OSM序列设计的UTR引物进行PCR扩增,如美国专利申请序列号12/648430中所述。表达的全长蛋白示于SEQIDNO:12中,其中裂解的活性形式由184个残基即51-227表示。使用标准方法将人和CynoOSM的前体和成熟形式在HEK293中表达并纯化。使用大肠杆菌来源的市售人OSM(R&DSystems,Cat.No.295-OM)作为对照在A375-S2细胞增殖和pSTAT信号传导测定中测试蛋白的功能活性。Mab当使用对照抗体时,使用指定CNTO6234的人IgG1同种型抗体。化学生物素酰化使用靶向细胞因子上的胺残基的NHS-酯化学(EZ-连接磺基-NHS-LC-生物素酰化试剂盒,Pierce,#21435)将重组人OSM生物素酰化。将生物素偶联反应优化,使靶标标记效率为每摩尔抗原一摩尔生物素。后者使结合和功能活性的损失最小化,同时确保蛋白群的近乎完全标记。在反应完成时,使用包括在EZ-连接磺基-NHS-LC-生物素酰化试剂盒(Pierce)中的Zeba脱盐离心柱从游离生物素试剂和残余离去基团中纯化蛋白。大约80%的起始材料被回收。使用HABA测定(Pierce生物素定量试剂盒,#28005)来测量生物素结合水平,该水平表明为每摩尔人OSM大约一摩尔生物素。使用Octet仪器(FortéBIO)来验证链霉抗生物素蛋白偶联和gp130-Fc(R&DSystems,Cat.No.671-GP)对生物素酰化蛋白的结合。Octet测量结果显示生物素酰化的人OSM结合gp130的特征图与未标记的起始材料的特征图基本上相同。体外靶向生物素酰化15个残基的Avi标签(GLNDIFEAQKIEWHE)(SEQIDNO:56)具有与内源性BirA底物BCCP类似的生物素-受体动力学(Beckett等人,1999,ProteinScience(《蛋白科学》))。当与所关注的蛋白连接时,具有一个受体赖氨酸残基的Avi标签将仅在一个位置被生物素酰化。使用从Avidity商购的生物素-蛋白连接酶和试剂在体外将重组CynoOSM位点特异性地进行生物素酰化。使用一价链霉抗生物素蛋白亲和树脂纯化生物素酰化的CynoOSM。通过SDS-PAGE和SEC-HPLC评价所得蛋白的质量。使用HABA测定(Pierce生物素定量试剂盒,#28005)来测量生物素结合水平,该水平表明为每摩尔cynoOSM大约一摩尔生物素。使用Octet仪器(FortéBIO)来验证链霉抗生物素蛋白偶联和gp130-Fc嵌合体对生物素酰化蛋白的结合。Octet测量结果显示生物素酰化的CynoOSM结合gp130的特征图与未标记的起始材料的特征图基本上相同。固相免疫测定初始噬菌体-Fab淘选使用以2μg/ml的于TBS中的生物素酰化的人OSM或cynoOSM涂覆的NEUTRAVIDINTMELISA板(Pierce)。在4℃下孵育过夜,封闭和洗涤之后,加入来自每轮淘选的多克隆噬菌体库的1∶100稀释液。用对pVIII,即M13噬菌体主要外壳蛋白(GEHealthcare,Cat.No.27-9421-01)特异的HR缀合的单克隆检测结合的噬菌体,然后加入化学发光底物POD(Roche,Cat.No.11582950001),并在PerkinElmer仪器上读数。使用来自大肠杆菌上清液的分泌性可溶Fab-His蛋白进行各个克隆的初步筛选。使用包含可溶Fab-His蛋白的细菌上清液进行ELISA格式中的结合。将黑色MaxiSorp板(Nunc,Cat.No.437111)用1μg/ml的羊抗人Fd(CH1)抗体(TheBindingSite,Cat.No.PC075)涂覆并在4℃下孵育过夜。在将板洗涤和封闭之后,加入50μl未稀释的细菌上清液(包含Fab-His蛋白)并使之在室温下孵育1小时,同时温和摇振。将板洗涤,并将生物素酰化的人或CynoOSM(20nM)加入到捕集的Fab中。在室温下1小时之后,加入SA-HRP(Invitrogen,Cat.No.43-4323)并如上进行化学发光检测。通过计算,用20nM浓度的人OSM进行的初步结合ELISA筛选将允许检测具有在纳摩尔亲和力范围内的亲和力的克隆。表位分箱(EpitopeBinning)表位分箱是为了基于结合特征划分MAb而进行的竞争测定法,其使用人IgG1转化的mAb和商业抗体MAB29进行,所述商业抗体已知为OSMRβ/LIFRα募集阻断剂(R阻断剂)。向384孔多阵列板(MesoScaleDiscovery(MSD),L25XA-4)的每个孔中加入2.5μg/ml的于磷酸盐缓冲盐水(PBS)pH7.4(Sigma,P3813)中的抗人Fc(JacksonImmuno,709-005-149)。将板在4℃下孵育过夜,然后在室温下用50μl的MSD阻断剂A封闭1小时。将384多阵列板洗涤三次(PBSpH7.4,0.05%Tween20(Scytek,PBT010),并向每个孔中加入1.0μg/ml测试mAb溶液,然后在室温下在效价板摇动器上以水平6摇振1小时。将板如前洗涤三次。平行地,在单独的96孔板(COSTAR,3357)中在MSD测定缓冲液(1∶3的封闭缓冲液与PBSpH7.4,0.05%Tween20)将5μg/ml竞争mAb溶液与1μg/ml生物素酰化OSM(R&DSystems,295-OM-010/CF)混合,并在室温下在效价摇动器上以水平3摇振1小时。将竞争抗体和生物素酰化OSM的前复合物加入到384多阵列板的每个孔中并在室温下摇振(在效价板摇动器上以水平6)1小时。然后将板洗涤三次并向每个孔中加入链霉抗生物素蛋白磺基标签(streptavidinsulfoTAG)(MSD,R32Ad-S)并在室温下摇振(在效价板摇动器上以水平6)摇振30分钟。将板洗涤三次并向每个孔中加入用蒸馏水按1∶4稀释的读数缓冲液T(MSD,R92TC-1)。将板在MSDS6000仪器上读数。基于在生物素酰化OSM存在且不存在竞争mAb的情况下对测试mAb获得的信号(最大信号)以及来自自身竞争的信号(最大信号减少2-4倍)来解读数据。当在竞争mAb存在的情况下的值在来自自身竞争的值的三个标准偏差内时,指定表位竞争。A375细胞A375细胞(ATCC:CRL-1619)为衍生自恶性黑素瘤的人上皮细胞。将A375-S2细胞(ATCC:CRL-1872,对IL-1敏感的CRL-1619亚系)在T-175培养瓶(Corning;Cat.No.431080)中培养于补充有10%FBS(Gibco)的完全生长培养基(DMEM/GlutaMax-I,Gibco)中,并在每三至四天大约80%铺满时以1∶20传代培养比率进行传代培养。A375-S2细胞增殖(BrdU掺入)测定使用化学发光ELISA通过BrdU掺入测量由人或食蟹猴OSM造成的A375-S2细胞增殖的减少。制瘤素M减少A375-S2人黑素瘤细胞系的增殖(Zarling等人,PNAS83:9739-9743)。该细胞系用于在抗体发现的所有阶段评价抗-制瘤素M抗体。在37℃下在95%O2-5%CO2中将A375-S2细胞维持在T-150组织培养瓶(BDFalcon35-5001)中的DMEM(Gibco11995)+10%FBS(Gibco16140)+1%青霉素/链霉素(Gibco15140)中并每周按1∶10分离两次。为了进行增殖测定,使细胞胰蛋白酶化(0.25%,Gibco25200)以将它们从T150瓶中移除,然后以2000细胞/孔的密度培养于黑色TC涂覆的板(BDFalcon353948)的内部48个孔中的200μLDMEM/FBS/青霉素-链霉素中。在37℃/95%O2-5%CO2下孵育过夜之后,移除培养基并以180μL的新鲜培养基替代。准备一个单独的板,该板包含10X终浓度的所有测试溶液。从该板转移20μL到细胞板中的对应孔中以形成适当的实验条件。每种实验条件以一式三份进行测试。在任何评价中和的实验中,将抗体和制瘤素M一起孵育至少1小时,然后加入到细胞中。然后将板在37℃/95%O2-5%CO2下孵育另外72小时。此时,进行化学发光BrdU细胞增殖ELISA(Roche11669915001)。将BrdU标记试剂加入到培养物中并保持4小时。然后移除培养基,并将100μL的固定溶液加入到每个孔中。在室温下30分钟之后,移除溶液并加入100μL/孔的抗-BrdU-POD溶液。在室温下2小时之后,将板用PBS-Tween洗涤并将100μL的SuperSignalPico(ThermoScientific37069)加入到每个孔中。然后使用PerkinElmerVictor3仪器读取发光。进行初始实验以确定自身产生的CHO细胞来源的重组人和食蟹猴制瘤素M抑制增殖的剂量-反应。从起始浓度100ng/ml到以1∶5稀释得到的终浓度0.0244ng/ml测试Osm。相比于用媒介物对照处理的细胞计算增殖抑制百分比。此实验的结果示于图2A中。随着人或cyno制瘤素M的浓度增加,BrdU掺入相应减少。基于抗增殖活性程度和效应的EC50,人和食蟹猴制瘤素M的抗增殖活性是不可区别的。基于这些实验,使用2ng/ml浓度的制瘤素M来评价和比较抗-制瘤素M抗体,因为该浓度将增殖抑制大约80%,从而给出在其中确定抗体剂量反应的较大窗口。在2ng/ml的人或cyno制瘤素M存在的情况下评价抗体的完全剂量范围。此外,在使用最高浓度的抗-制瘤素M抗体时,同种型对照包含在实验中。测定窗口由未处理的对照孔(最大增殖)和与单独2ng/ml制瘤素M孵育的孔(最小增殖)之间的差异限定,并且在任何抗体浓度下的中和百分比限定在该窗口内。使用A375-S2磷酸-STAT3测定的pSTAT3信号传导OSM通过结合细胞表面受体gp130并诱导受体与OSMRβ异源二聚化以开始细胞内信号传导级联放大来抑制A375-S2细胞的生长,所述信号传导级联放大包括信号传导分子STAT3的激活(磷酸化)(Kortylewski等人,Oncogene(《癌基因》)18:3742-3753,1999)。STAT3信号传导的破坏中止A375-S2细胞的OSM生长抑制,表明STAT3激活是OSM信号传导中的关键步骤(Heinrich等人,Biochem.J.(《生物化学杂志》)374:1-20,2003)。已显示A375-S2细胞中的STAT3磷酸化是依赖OSM浓度的并且可利用商业试剂盒来测量受刺激的细胞中的STAT3磷酸化。选择A375-S2细胞中OSM诱导的STAT3磷酸化的中和作为Mab候选物的初步筛选测定。对于OSM诱导的STAT3磷酸化的抗体中和,将A375-S2细胞以25,000细胞/孔接种到96孔组织培养板中(Corning;Cat.No.3596)的200μl完全生长培养基中并孵育24小时。将细胞用包含5ng/ml人OSM(重组的、哺乳动物细胞来源的OSMN1-1)的溶液处理,该溶液已在室温下与1∶5连续稀释的实验mAb(从10μg/m起始)孵育3小时。对照包括未处理的细胞、刺激的细胞(仅5ng/mlOSM)以及hIgG1同种型对照mAb。除非另外指明,否则所有处理均以一式三份进行。按照制造商规程使用磷酸-STAT3全细胞裂解液试剂盒(MSD;Cat.No.K150DID-1,LotNo.K0010570)进行pSTAT3含量的分析。简而言之是,将细胞在200μl/孔体积中处理10分钟;移除处理溶液并通过多通道移液管加入50μl的细胞MSD裂解缓冲液。将板在轨道摇动器(300RPM)上放置5分钟。然后,将每种裂解液转移30l到MSD磷酸-STAT396孔板中。将板密封并在室温下在轨道摇动器(300RPM)上放置1小时,用150μl的MSD洗涤缓冲液洗涤三次,然后将25μl的第二检测抗体缀合物(抗-pSTAT3-Ru(bpy)32+)加入到每个孔中,再次密封并在室温下在轨道摇动器(300RPM)上孵育1小时。将板如前洗涤并将150μl的MSD读数缓冲液(三丙胺溶液)加入到每个孔中。将板在MSDSECTORImager6000仪器上读数。获得行内成熟的、亲本和对照mAb的完全EC50剂量-反应曲线并将其绘制成归一化的百分比pSTAT3信号。通过表面等离子体共振进行的亲和力测量(Biacore)使用所述的人或CynoOSM构建体,用表面等离子体共振(SPR)与Biacore3000光学生物传感器(Biacore)测量结合亲和力。生物传感器表面这样来制备:使用用于胺偶联化学的制造商说明书将抗小鼠(Jackson,Cat.No.315-005-046)和抗人(Jackson,Cat.No.109-005-098)的抗IgGFc抗体混合物偶联到CM-5芯片(Biacore,Cat.No.BR-1000-14)的羧甲基化葡聚糖表面。大约19,000RU(反应单位)的抗OSM抗体被固定在四个流通池的每一个中。在25℃下在运行缓冲液(DPBS+0.005%P20+3mMEDTA)中进行动力学实验。用运行缓冲液制备100nM至0.412nM的连续稀释人和CynoOSMECD。约200RU的mAb被捕集在传感器芯片的流通池2至4上。流通池1用作参考表面。捕集mAb后,以50uL/min注射抗原三分钟(缔合相),随后是10分钟的缓冲液流(解离相)。通过两个18秒的脉冲以50μl/min注射100mMH3PO4(Sigma,Cat.No.7961),使芯片表面再生。使用BIAevaluation软件(Biacore,3.2版)处理收集的数据。首先采用如下方式将数据减去两个参照值:从分析物注射得到的曲线中扣除参照值,再从该曲线中扣除缓冲液注射生成的曲线。使用整体拟合的1∶1结合模型进行数据的动力学分析。每个mAb的结果以Ka(结合率)、Kd(解离率)和KD(亲和常数)的格式记录。实例2:OSM结合FAB的选择从头Fab-pIX文库已描述于Shi等人,JMolBiol(《分子生物学杂志》)397:385-396,2010;WO09085462A1;美国序列号12/546850;并且本文中上述被指定169、323和551,其参考使用的重链人种系框架:IMGT命名中的IGHV1-69(SEQIDNO:1)、IGHV3-23(SEQIDNO:2)或IGHV5-51(SEQIDNO:3)。所述三个重链文库框架与如下四个轻链文库VLκ框架组合:A27(IGKV3-20*01(SEQIDNO:5))、B3(IGKV4-1*01(SEQIDNO:6))、L6(IGKV3-11*01(SEQIDNO:7))和O12(IGKV1-39*01(SEQIDNO:8))。在文库中,通过添加包含重链中的SEQIDNO:4和轻链中的SEQIDNO:10的J区(FR4)而使FabV区完整。重链CDR3具有7-14个残基的可变长度。每个文库的完整V区的例子示于图1中并被编号,并且CDR区根据Kabat显示。初始组的抗OSM噬菌体展示命中使用市售的糖基化人OSM鉴定。按照用于噬菌体选择的公布方案(Marks和Bradbury,AntibodyEngineering(《抗体工程》)第248卷:161-176,HumanaPress,2004)使用捕集在顺磁性链霉抗生物素蛋白(SA)珠(Invitrogen,Cat.No.112.05D)上的生物素酰化的人OSM(R&DSystems,Cat.No.295-OM)对Fab-pIX噬菌体展示文库进行淘选。简而言之,将生物素酰化的人OSM加入到已预吸附在未缀合的珠上的噬菌体文库中使最终浓度为100nM并在温和旋转下孵育1小时。加入封端的SA珠并孵育15分钟以捕集生物素酰化的OSM与结合的噬菌体。将磁力捕集的噬菌体/抗原/珠复合物用1ml的TBST洗涤5次并用1mlTBS洗涤1次。在移除最终TBS洗涤液之后,加入1ml的按指数律生长的TG1细胞(Stratagene,Cat.No.200123)并在37℃下且不摇振的情况下孵育30分钟。将经感染的细菌铺涂在LB/琼脂(1%葡萄糖/100μg/ml羧苄青霉素)板(Teknova,Cat.No.L5804)上并在37℃下孵育过夜。刮取细菌菌苔并制备甘油原种[15%甘油/羧苄青霉素(100μg/ml)/2xYT]并在-80℃下保存。为了制备用于第二轮淘选的噬菌体,将25ml的2xYT/羧苄青霉素(100μg/ml)用25μl的细菌甘油原种接种并在37℃下培养直至OD600为大约0.5。将辅助噬菌体VCSM13(Stratagene,Cat.No.200251)以大约10∶1的感染复数加入到培养物中并在37℃下且不摇振的情况下孵育30分钟。细菌自旋向下,沉淀物重悬于诱导培养基(2xYT/羧苄青霉素/卡那霉素/IPTG)中并在30℃下培养过夜。将噬菌体用2%PEG/0.25MNaCl(终浓度)沉淀并重悬于2ml的PBS中。将第一轮噬菌体在4℃下保存并用于进行第二轮淘选。淘选参数为:第1轮,100nM抗原,在室温下孵育1小时,用TBST洗涤5次然后用TBS洗涤1次;第2轮,10nM抗原,在室温下孵育1小时,用TBST洗涤10次/用TBS洗涤1次;以及第3轮,1nM抗原,在4℃下孵育16小时(过夜),用TBST洗涤10次/用TBS洗涤1次。使用ELISA监测成功,其中Fab被抗人Fd(CH1)抗体捕集,加入20nM的生物素酰化的人OSM并用SA-HR检测结合的OSM。如展示的Fab的三十(30)个独特的重链-轻链配对被鉴定,通过ELISA显示,它们与CynoOSM交叉反应。重链代表来自169(IGHV1-69衍生的)和551(IGHV5-51衍生的)文库的序列,并与代表所有四个文库种系来源(A27、B3、L6和O12)的轻链可变区组合。实例3:osm结合MAB的表征四螺旋束构造的OSM的特征在于由相对松散的环连接的四个α螺旋区段,这些螺旋区段定名为A、B、C和D。OSM通过位于螺旋A和C中的表面(位点II)与gp130相互作用,所述表面经确定包括SEQIDNO:1的氨基酸残基Q16、Q20、G120、N123、N124的接触(Deller等人,Structure(《结构》)8(8):863-874,2000;Liu等人,Int.J.Mol.Med.(《国际分子医学杂志》)23:161-172,2009)。据信,负责OSM与OSMRβ和LIFRα相互作用的表面(位点III)主要由位于螺旋D中的残基限定(Deller等人,同上)。目的是选择OSM的高亲和力连接子,其能够通过阻止OSM与gp130结合(位点II或B阻断剂)或阻止OSM结合的gp130募集LIFRa或OSMRb(位点III或R阻断剂)来阻止OSM驱动的gp130信号传导。将30个初始选择的OSM结合Fab中的29个克隆到用于转化为全长人IgG1Mab的载体中。表征测定为(1)用于鉴定表位簇或“箱”的竞争性结合,(2)通过表面等离子体共振的亲和力测量(Biacore),以及(3)阻断pSTAT3信号传导的能力。已使用如实例1中所述的哺乳动物细胞产生的(糖基化)人和cyno蛋白进行所有的筛选和测定。结果基于pSTAT3和ELISA结合测定中mAb相对于对照MAB295(R&DSystems)的分级选择的mAb子集的亲和力测量数据示于表3中。表3:选择的抗OSM第一轮mAb子集的亲和力。分箱测定的结果显示M5、M6和M9彼此竞争但不与MAB295竞争。M10与MAB295竞争。通过pSTAT3测定,这四个mAb也显示为gp130信号传导的功能中和剂。因此,得出如下结论:M10为R阻断剂,M5、M6和M9阻断OSM与gp130的结合(B阻断剂)。治疗先导物的靶标亲和力由明确地竞争OSM-gp130相互作用的需要决定,OSM-gp130相互作用的亲和力(KD)已测量为大约1nM。因此,期望对OSM的靶标亲和力为100pM或更低KD。发现为OSM-gp130相互作用阻断剂的中和MAbM5、M6和M9,加上M10被选择用于内联成熟,以便产生满足治疗候选物的100pM亲和力要求的衍生物。附加的所需特性是,mAb以在对人OSM的亲和力的五倍内的亲和力结合食蟹猴OSM(KD为500pM或更低)。发现这四个Mab的结合域的组成代表经定名的四个独特的重链和轻链对,如表4中所示。表4:MabHC文库HCIDLCIDM10169H2L2M6551H14L12M9551H17IGKV4-1(B3)M5551H135L111完整可变区序列包含用于形成本文上述噬菌体文库的种系序列,因此,固定的残基匹配未突变的亲本种系残基。就这一点而言,每个V区由定名为SEQIDNO:1-3或5-8的支架内的指定CDR1和CDR2;接着CDR3(表3和4)以及分别是轻链可变区和重链可变区(表5LC和表6HC)接着J区组成,重链的J区为SEQIDNO:4,轻链的J区为SEQIDNO:10。表5:HC可变区H2包含衍生自SEQIDNO:1的FR1-CDR1-FR2-CDR2-FR3,其中X1=A,X2=G,X3=I,X4=P,X5=I,并且X6=F,其中H-CDR2中有附加突变。来自文库551的HC(H14、H17和H135)包含衍生自SEQIDNO:3的FR1-CDR1-FR2-CDR2-FR3,其中X1=S,X2=S或G,X3=I,X4=Y,X5=G并且X6=Y或D。四个HC中的每一个由独特的CDR3(SEQIDNO:19-22)组成。表6:选择的Fab的LC可变区均衍生自B3文库并且包括在IMGT数据库中表示为IGKV4-1(B3)的种系序列,IGKV4-1(B3)被用作文库多样化的起始序列,如本文和引用的出版物所述。四个轻链中的三个在CDR1中有差异并具有相同的H-CDR2。四个选择的LC可变FR1-CDR1-FR2-CDR2-FR3的共有序列衍生自SEQIDNO:8,其中X1为Y、S或A;X2为K、E或N;X3为Y、W或F;并且X4总是为W。鉴定了两个独特的CDR3序列。L-CDR3可由共有序列(SEQIDNO:29)表示,该共有序列由式Q-Q-(S,Y)-(F,Y)-S-(F,T)-PLT表示。实例4:亲和力重新选择实例3中描述的四个V区配对即OSMM5、OSMM6、OSMM9和OSMM10被选择用于轻链重新选择以改善亲和力。为了以一种有效且迅速的方式使来自初步选择的大量抗体亲和力成熟,使用描述于Shi等人,JMolBiol(《分子生物学杂志》)397:385-396,2010和WO09085462A1以及美国序列号12/546850中的“内联”成熟方法。简而言之,将在起始几轮的选择中获得的抗原特异性克隆的VH区与对应VL支架的文库组合,在这种情况下为基于B3的V区(SEQIDNO:8)。形成三个新的文库,一个使用用于初步选择中的VL文库作为多种VL链的来源,而另外两个文库基于已知结构的抗原-抗体复合物的近期分析而设计(Raghuanthan等人,J.Mol.Recognit(《分子识别杂志》),2010)。基于最可能参与靶标蛋白的结合(也称为特异性决定残基用途(SDRU))的那些残基来选择用于多样化的残基。在Vκ轻链中,这已被确定为以Chothia和Lesk所定义的高变环为中心的三个接触区,它们根据靶标是蛋白、肽或小半抗原而稍有不同。表5示出用于亲和力成熟的VL文库,其中B3是与发现阶段期间所用相同的文库,文库2“聚焦SDRM的(SDRMfocused)”基于SDRU残基并聚焦多样性,而NNK为随机化文库。在该表中,“X”意指由NNK混合形成的任何氨基酸加上一个终止密码子。表7汇总了文库1、2和3中的多样性。在分析最终文库期间,鉴定了一些氨基酸,这些氨基酸不是初始设计的部分,因此是由于合成方法而引入。对于文库2,使用二核苷酸进行合成,这些氨基酸为:S(位置30c)、T(位置30d)、EK(位置30f)、IW(位置32)、TV(位置50)、I(位置92)、D(位置93)、以及F(位置96)。表7:由第三轮淘选输出制备Fab-His蛋白,并且采用单克隆Fab-HisELISA来鉴定具有比对应亲本Fab高的结合信号的各个命中。在这些分级ELISA中使用两个浓度的生物素酰化的人抗原:2nM和0.2nM。将每个亲本克隆的结合信号设定为100%。存在22个来自M6和M9的亲和力成熟的命中,当与包含初始重链和轻链V区的亲本Fab相比时显示出提高高达9倍(900%)的结合。重链和轻链的这些新配对中的一些被选择用于进一步评价。与MAB295对照和亲本mAbM6和M9的亲和力相比的人和食蟹猴OSM的内联成熟mAb的亲和力(KD)汇总于表8中,并且那些mAb的CDR组成示于表9中。在一些情况下,LCV与H14和H17均配对。基于它们的生物物理特性和功能效能指定为候选治疗先导物的某些mAb以灰色突出显示。表8:表9:较高亲和力Mab的LC-CDR表选择的候选物中的LC-CDR3多样性Q-Q-(S,Y)-(F,Y)-S-(F,T)-PLT(SEQIDNO:29)被降低,并且重新选择的高亲和力LC-CDR3的共有序列表示为QQY-(F,Y)-STP-(L,I)-T(SEQIDNO:47)。对于重新选择的mAb,H14和H17的VH都具有共同的FR1-CDR1-FR2-CDR2-FR3,其由SEQIDNO:48给出并包含SEQIDNO:14(CDR1)和SEQIDNO:17(CDR2)。降低人和CynoA375-S2细胞增殖的能力为了评价内联成熟的抗体,进行剂量-反应,以5μg/ml或1μg/ml起始,并以1∶5稀释分别降低至0.0016或0.00032μg/ml。M71导致的中和示于图2B中。随着抗体浓度增加,人和食蟹猴制瘤素M两者的抗增殖效应被中和。使用来自用人(开放符号)和食蟹猴(闭合符号)制瘤素M进行的三个单独实验的数据计算剂量-反应曲线。表10汇总了M55、M64、M69和M71抗人和食蟹猴制瘤素M的IC50(95%置信区间)。表10:人gp130竞争使用如实例1中所述的表面等离子体共振(SPR)与Biacore3000光学生物传感器(Biacore)在选择用于内联成熟的抗OSMmAb加上来自上表的选择的新连接子和人gp130之间进行竞争实验。通过按照制造商说明书将每种测试mAb偶联到CM-5芯片(Biacore,Cat#BR-1000-14)的羧甲基化葡聚糖表面来制备生物传感器表面。每种测试mAb大约4,000-15,000RU(反应单位)被固定在仪器的四个流通池的每一个中。在25℃下在运行缓冲液(DPBS+0.005%P20+3mMEDTA)中进行竞争实验。将人OSM(自身OSMN1-1)用运行缓冲液稀释至30nM并以3μl/min在具有固定的mAb的每一个流通池上注射3分钟。捕集人OSM后,注射300nM的竞争mAb或人gp130-Fc(缔合相)三分钟,随后是注射缓冲液流(解离相)3分钟。通过两个12秒的脉冲以50μL/min注射100mMH3PO4(Sigma,Cat#7961),使芯片表面再生。使用BIAevaluation软件3.2版(Biacore)处理收集的数据。首先,在注射人OSM时将传感图对齐。然后记录人OSM、竞争mAb或gp130-Fc的结合水平(RU)。当将竞争mAb或gp130-Fc注射在OSM表面上时结合水平(RU)上升,则表明与固定的测试mAb不存在竞争,反之亦然。流通池(Fc1等)固定的mAb沿着表10的横向样品行出现。M2之前显示与商业抗体MAB295竞争,已知MAB295不与结合OSM的gp130竞争。竞争结合测定的结果显示,M54、M55、M64、M69和M71与人gp130-Fc竞争OSM抗原(表11)。表11:人gp130竞争成熟的候选先导物mAb。在A375细胞中阻断pSTAT3的能力在存在或不存在5ng/mlhOSM的情况下,在A375细胞中进行M6和M9以及这些Mab的内联成熟的变体对pSTAT3的抑制,所计算的EC50值示于表12中。表12:pSTAT3抑制mAbIDEC50(ng/ml)95%置信区间曲线拟合R2M6529.524.35至35.820.9921M6737.730.93至45.920.9905M4550.042.45至58.910.9931M8371.259.59至85.050.9908MAB29572.146.91至110.70.9848M5478.566.11.至93.230.9930M6388.258.55至132.90.9819M6497.177.01至122.40.9902M68126.9107.2至150.10.9924M69141.7115.1至174.50.9914M66152.7111.1至209.80.9833M71159.6128.3至198.50.9885M55181.6153.0至215.60.9869M42191.393.67至390.70.9607M62233.5206.4至264.10.9947M53236.6196.3至285.20.9859M6650.3241.7至17500.9540M91218.0352.8至42080.9614M85-0.4454IL13-0.3438实例5:生物活性巨噬细胞-软骨细胞共培养测定在巨噬细胞-软骨细胞共培养体系中评价M71。已知分化的巨噬细胞产生制瘤素M(Hasegawa等人,Rheumatology(《风湿病学》)38:612-617,1999)。OSM可降低构成软骨基质的重要部分的高度硫酸化蛋白聚糖类多聚糖白聚糖(GAG)的合成。.使用重组HEK产生的His-Avi标记的人和食蟹猴制瘤素M来发现抗人制瘤素M抗体,并且使用这些分子以及来自R&D体系(295-OM)的细菌来源的重组人制瘤素M来评价这些抗体的活性。这些都与原生内源性人制瘤素M不相同,是因为his-avi标签(HEK产生的Osm)或缺少糖基化(细菌重组Osm)。已知巨噬细胞分泌制瘤素M(Grove等人,JLipidRes(《脂质研究杂志》)32:1889-97,1991)并且制瘤素M降低人软骨细胞中的蛋白聚糖合成(Sanchez等人,OAandCart.(《骨关节炎与软骨》)12:810-10,2004)。因此,使用巨噬细胞-软骨细胞共培养体系来确定抗人制瘤素M抗体中和内源性或原生人制瘤素M的能力。简而言之,将包含40,000个正常人关节软骨细胞的单一藻酸盐珠在分化的人巨噬细胞存在的情况下培养72小时。这些实验在剂量范围的抗制瘤素M抗体存在的情况下进行。在实验结束时,通过掺入放射性355O4来测量蛋白聚糖合成。从所有细胞中获得CD14+外周血单核细胞。将单核细胞以2.5×105细胞/孔培养在48孔板上的0.5ml巨噬细胞培养基(RPMI+具有10%热灭活的FBS、1%NEAA和1%青霉素-链霉素的谷氨酰胺)中。将细胞用100ng/ml的巨噬细胞-集落刺激因子(M-CSF)处理。48小时之后,置换培养基以移除非贴壁细胞。在第6天时,将含有M-CSF的巨噬细胞培养基的用不含M-CSF的巨噬细胞培养基置换。在第8天时,将巨噬细胞培养基用软骨细胞培养基(50%Ham’sF-具有10%胎牛血清的12/50%DMEM)置换并将单一藻酸盐珠软骨细胞培养物(ArticularEngineering#CDD-H-2200)加入到每个孔中。保留等分试样的巨噬细胞条件培养基并在-80℃下保存,以用于使用R&D体系人制瘤素MDuoSet(DY295)分析制瘤素M水平。将藻酸盐珠-巨噬细胞共培养物在20μg/ml的抗人制瘤素M抗体(M64,M71,M55,M69)存在的情况下或在剂量范围(5μg/ml至0.00076μg/ml;1∶3稀释)的抗体(M71andM55)存在的情况下维持。此外,人IgG1同种型对照(CNTO6234)以测试的抗制瘤素M抗体的最高浓度包含在板上。其他对照是仅软骨细胞(没有共培养)以及在2ng/ml人制瘤素M存在的情况下的软骨细胞。此外,维持仅包含巨噬细胞的孔以便测量制瘤素M产生。在共培养72小时之后,将10μCi/ml放射性35SO4(Perkin-ElmerNEX041H002MC)加入到每个孔中并保持另外20小时。使用CPC沉淀技术(MPBIomedicals(#190177)测量35SO4的掺入。在与35SO4孵育20小时之后,移除经标记的培养基并将每个珠用含Ca和Mg的DPBS洗涤两次。在第二次洗涤之后,将200μl的柠檬酸盐缓冲液(150mMNaCl,55mM柠檬酸钠,ph6.8)加入到每个珠。将板在37℃下孵育10至15分钟直至柱被溶解。将来自每个孔的100μl等分试样转移到用1%CPC预湿的Millipore多层筛96孔滤板,然后将10μl的10%CPC加入到每个孔中并保持5分钟。然后对板抽真空直至过滤器干燥。然后将每个孔用200μl的1%CPC洗涤2次,每次均抽真空直至过滤器干燥。然后将塑性底部从板移除并用密封物(PerkinElmer#6005185)置换。将闪烁液(50μl,PerkinElmer#6013621)加入到每个孔中并将板密封物施加到板的顶部。然后在顶部计数读出器(TopCountreader)上对板计数。在20μg/ml时,M64、M71、M55和M69将蛋白聚糖合成增加至在不存在抗体的情况下观察到的水平以上,而同种型对照无效果(图3)。在单独的实验中,M71以剂量依赖的方式将蛋白聚糖合成增加至单独软骨细胞表现出的水平(定义为100%中和),EC50为30ng/ml,而同种型对照无效果。这些数据显示,巨噬细胞来源的Osm降低共培养的软骨细胞中的蛋白聚糖合成,并且抗人制瘤素M抗体中和原生制瘤素M。人肺成纤维细胞磷酸-STAT3测定OSM诱导正常人肺成纤维细胞的增殖和胶原产生(Scaffidi等人,Br.J.Pharmacol(《英国药理学杂志》)136:793-801,2002)。成纤维细胞过度产生胶原是多种病理病症的关键特征(Lim等人,Oncogene(《癌基因》)23(39):5416-25,2006;Huang等人,JCellBiochem(《细胞生物化学杂志》)81(1):102-13,2001)。制瘤素M受体信号传导激活JAK-STAT通路,并且STAT3的磷酸化是信号传导通路中的早期事件(Auguste等人(1997)SignalingofTypeIIOncostatinMReceptor(II型制瘤素M受体的信号传导)JBiolChem(《生物化学杂志》)272:15760-15764)。使用R&D体系人/小鼠pSTAT3Duoset(DYC4607-5)在正常人肺成纤维细胞(NHLF)中确定制瘤素M产生pSTAT3的能力。然后使用该测定来确定M55和M71中和制瘤素M信号传导的能力。使用在Lonza专有的FGM-2(CC-3132)培养基上培养的来自Lonza的NHLF(CC-2512)进行这些实验。简而言之,将细胞以25,000细胞/孔接种在板上的FGM-2中并培养24小时。然后将细胞用制瘤素M或抗体加上制瘤素M处理10分钟。为了避免在该10分钟孵育期间的温度依赖效应,将用于准备处理物的所有溶液在37℃下预热并维持。在10分钟处理之后,吸掉培养基并用完全裂解缓冲液置换。裂解缓冲液(pH=7.2)由1%NP-40、1%脱氧胆酸钠、0.1%SDS、0.15MNaCl和0.01M磷酸钠组成,并在4℃下保存。使用时,完全裂解缓冲液这样来制备:将1片蛋白酶抑制剂混合物(Roche,11836153001)和110μLHALT磷酸酶抑制剂(ThermoScientific78420)加入到11ml的裂解缓冲液中。在10分钟裂解步骤之后,所得的裂解液准备用于pSTAT3检测。为了确定制瘤素M剂量-反应,将NHLF细胞用剂量范围的OSM(100ng/ml至0.024ng/mlw/1∶4稀释,一式三孔)处理。用预热的PBS+1%BSA准备稀释板,其中每个孔中Osm的浓度比处理需要的终浓度高10X,并包括仅有培养基的孔作为未处理的对照。将培养基从培养板完全移除并用180μl的预热FGM-2置换。将计时器开启10分钟,然后从稀释板中的每个孔转移20μl到培养板中的对应孔中。10分钟之后,通过抽吸完全移除细胞板中的处理溶液并用100μl的完全裂解缓冲液置换。为了最小化孵育时间的差异,将裂解缓冲液以与处理相同的顺序加入到孔中。然后将测定板在摇动器上放置10分钟。摇振之后,将裂解液在-80℃下冷冻用于以后测试或直接转移到涂覆有抗pSTAT3的ELISA板中。根据制造商说明书进行ELISA(R&D体系人/小鼠pSTAT3Duoset),不同的是1)仅将90μl的裂解液或标准物加入到每个孔中和2)SuperSignalPico(ThermoScientific37069)用作HR底物。在Victor3读板仪上对ELISA板读取发光。为了评价M55和M71在NHLF中和OSM信号传导的能力,在2ng/ml人OSM存在的情况下将细胞用剂量范围的抗OSM抗体(一式三孔,500ng/ml至0.005ng/ml,以1-10稀释或50ng/ml至1.563ng/ml,以1∶2稀释)处理。用PBS准备稀释板以用于20X终浓度的抗体剂量-反应处理,包括用于同种型对照(抗OSM抗体的最高浓度)的孔,而且未处理的对照和仅用OSM处理的细胞均不含抗体。单独用FGM-2以40ng/ml(20X终浓度)制备人制瘤素M。在稀释板中将20X抗体和OSM溶液按等体积混合以形成10X处理溶液,并将板在37℃下孵育1小时以允许OSM与抗体结合。1小时之后,将培养基从培养板移除并用180μl的预热FGM-2置换。将计时器开启10分钟,并从稀释板中的每个孔转移20μl到培养板中的对应孔中。10分钟之后,通过抽吸完全移除细胞板中的溶液并用100μl的完全裂解缓冲液置换。为了最小化孵育时间的差异,将裂解缓冲液以与处理相同的顺序加入到孔中。然后将测定板在摇动器上放置10分钟。摇振之后,将裂解液在-80℃下冷冻用于以后测试或直接转移到涂覆有抗pSTAT3的ELISA板中。根据制造商说明书进行ELISA(R&D体系人/小鼠pSTAT3Duoset),不同的是1)仅将90μl的裂解液或标准物加入到每个孔中和2)SuperSignalPico(ThermoScientific37069)用作HR底物。在Victor3读板仪上对ELISA板读取发光。人制瘤素M增加NHLF细胞中的pSTAT3,其中EC50为大约1ng/ml。制瘤素M剂量反应的实例提供于图4A中。对于该实例,EC50为0.90ng/ml,且95%置信区间为0.70至1.17ng/ml。抗制瘤素M抗体的中和能力在2ng/ml的制瘤素M存在的情况下确定,并且所有数据归一化成在2ng/mlOSM存在且不存在抗体的情况下的发光。图4B示出M71对OSM诱导的STAT3磷酸化的剂量依赖性中和。随着M71的浓度增加,STAT3磷酸化程度降低。由来自六个单独实验的数据计算剂量-反应曲线,并且对M71计算的IC50为8.9ng/ml,95%置信区间为6.9至11.6ng/ml。实例6:体内活性在全身施用人制瘤素M之后,对M71评价其阻断体内细胞因子产生的能力。腹膜内注射人制瘤素M增加某些血清细胞因子的水平,推测是通过与鼠白血病抑制因子受体相互作用。给小鼠全身(i.p.)施用人制瘤素M发展为用于评价抗制瘤素M单克隆抗体在体内环境下的中和能力的模型。给小鼠i.p.注射于200μl的PBS中的10μg人制瘤素M或PBS媒介物对照。1小时之后,用CO2使小鼠麻醉并通过末端心脏穿刺收集血液。使各个血液样品在冰上凝结20分钟,然后以3500rpm旋转10-15分钟。将血清样品冷冻保存,直至按照制造商说明书用Milliplex鼠MAP细胞因子/趋化因子多路复用(32)面板进行分析。样品分析显示,相比于媒介物对照,人制瘤素M显著增加鼠KC、IP-10、MCP-1、IL-6和嗜酸性粒细胞趋化因子的血清水平,对面板中的其他细胞因子无影响。这些数据显示,注射人制瘤素M诱导细胞因子释放,推测是通过与鼠白血病抑制因子受体相互作用(Richards等人,JImmunol.(《免疫学杂志》)159:2431-37,1997;Lindberg等人,MolCellBiol(《分子细胞生物学》)18:3357-3367,1988),所述受体可用于测试抗制瘤素M抗体的体内中和能力。在小鼠全身施用模型中评价M71和M55。简而言之,对小鼠皮下给药M71或M55(20、2.0或0.2mg/kg的人IgG1抗人Osm)、CNTO6234(huIgG1同种型对照,20mg/kg)或体积为10μl/g的PBS。24小时之后,然后给每只小鼠i.p.注射10μg的于PBS(SigmaD8357)中的自身产生的CHO细胞来源的重组人制瘤素Mw/0.1%小鼠血清白蛋白(SigmaA3559)或单独PBS-MSA媒介物对照(200μL总体积)。1小时之后,用CO2使小鼠麻醉并通过末端心脏穿刺收集血液。使各个血液样品在冰上凝结20分钟,然后以3500rpm旋转10-15分钟。将血清样品冷冻保存,直至使用Milliplex鼠MAP细胞因子/趋化因子多路复用(32)面板进行分析。按照制造商说明书分析血清样品。人制瘤素M诱导通过试剂盒检测的细胞因子中的如下五种的血清水平显著(非配对学生t检验)增加:嗜酸性粒细胞趋化因子、IL-6、IP-10、KC和MCP-1。以20mg/kg预先给药同种型对照CNTO6234对制瘤素M诱导的细胞因子释放没有影响。然而,预先给药M71显著降低IP-10、MCP-1、IL-6和嗜酸性粒细胞趋化因子(以2.0和20mg/kg给药)以及KC(以20mg/kg给药)的血清水平。0.2mg/kg的M71对任何细胞因子均没有影响。M71和同种型对照对IP-10和MCP-1的影响分别示于图5A和B中。观察到M55具有不那么稳健的中和作用,因为20和2.0mg/kg的中和作用仅见于IP-10。用20mg/kgM55时,IL-6、嗜酸性粒细胞趋化因子和MCP-1的血清水平降低,而用任何剂量的M55均未见KC水平降低。这些数据显示了抗制瘤素M抗体中和鼠全身施用模型中外源人制瘤素M的生物效应的能力。IP-10即干扰素γ诱导蛋白10kDa或可诱导的小细胞因子B10是在人中由CXCL10基因(C-X-C基序趋化因子10(CXCL10))编码的蛋白。多种作用已归因于CXCL10,例如单核细胞/巨噬细胞、T细胞、NK细胞和树突细胞的化学趋向性以及促进T细胞粘附内皮细胞。KC,现在称为趋化因子(C-X-C基序)配体1(CXCL1)是属于CXC趋化因子家族的小细胞因子,其之前被称为GRO1癌基因、GROα、中性粒激活蛋白3(NAP-3)和黑素瘤生长刺激活性α(MSGA-α)。在人中,该蛋白由CXCL1基因编码。CXCL1由巨噬细胞、中性粒细胞和上皮细胞表达,并具有中性粒细胞趋化活性。实例7:共结晶学将包含V区H17(SEQIDNO:51)和L180(SEQIDNO:55)的Fab片段与人OSM(SEQIDNO:10)残基26-212结晶。OSM与gp130细胞因子家族的其他成员共有其四螺旋束三维结构。四螺旋束构造的特征在于由相对松散的环连接的四个α螺旋区段,这些螺旋区段定名为A(残基10-37)、B(残基67-90)、C(残基105-131)和D(残基159-185)。OSM通过称为位点II的表位与gp130相互作用,位点II包括位于螺旋A和C中的氨基酸残基(Q16,Q20,G120,N123,N124)(Deller等人,Structure(《结构》)8(8):863-874,2000;Liu等人,Int.J.Mol.Med.(《国际分子医学杂志》)23:161-172,2009)。据信,位点III即负责OSM与OSMRβ和LIFRα相互作用的表位,主要由位于螺旋D中的残基限定(Deller等人,Structure(《结构》)8(8):863-874,2000)。结晶在20℃下通过沉滴蒸气扩散方法进行复合物的结晶,使用Oryx4蛋白结晶机械手(DouglasInstruments),分配等体积的0.2μL蛋白复合物(10.95mg/mL)和0.2μL储备溶液。进行多重结晶筛选。大部分的液滴保持澄清,反映出复合物的高溶解度。从0.1MMESpH6.5,2.4M硫酸铵和0.1MTrispH8.5,3.5M甲酸钠获得晶体。晶体结构溶液的结果与H14/L180Fab接触的OSM残基构成结合表位。与OSM接触的抗体残基构成结合互补位。两个可变域的所有六个CDR参与OSM结合。接触的残基在表13中给出并示于图5中。长CDR-L1连同重链可变区H1的CDR形成山谷状的抗原结合位点,其中底部具有小脊(图4C,左图)。在结合时,山谷的两侧沿着螺旋A和C包围OSM四螺旋束,其中底部脊结合在两螺旋之间(图4C,右图)。抗体和抗原结合界面遮蔽的溶剂可触及表面(对于Ab为对于Ag为)。虽然界面处存在多个带电残基,但不存在电荷-电荷配对,表明vdw和H键合在抗体和抗原相互作用中发挥最重要的作用。表13:*对于氢键(以粗体突出显示)距离截止值为而对于vdw接触为因此H17/L180Fab在之前显示被gp130接触的残基;Q20和G120处,并沿着A和C螺旋接触OSM。实例8:药代动力学OSM是与炎性过程相关的可溶性靶标,与细胞上的细胞表面展示的靶标不同。因此可使用用赋予改变的FcR结合的突变工程化Fc的方法将包含如本文发现的结合区和另外具有Fc域的完整IgG的性质调整成与其用途相关的目的和治疗规格。在本发明组合物中,循环中的缓释活性和持久性是治疗单克隆IgG的有益规格。因此,Fc域对新生受体(FcRn)具有增强的亲和力。使用之前描述的突变M428L(MedImmune美国专利No.7670600)结合N434S(US7371826,WO2006/053301),使用标准重组技术构造突变的和野生型抗体。将这两个Mab即M71和M71L/S在标准活性测定中进行比较,并比较它们在非人的灵长类动物的循环中的持久性。测定在A375-S2增殖测定中并行比较M71和M71L/S。从起始浓度1μg/ml至0.00032μg/ml(以1∶5稀释)评价剂量-反应。在1μg/ml的浓度时,这些抗体的同种型对照即分别是CNTO3930和CNTO8852,对2ng/ml的人制瘤素M抑制增殖的能力没有影响。M71和M71L/S在1μg/ml时均完全中和制瘤素M的效应,并且IC50无可测量的差异。在小鼠全身施用模型中比较M71和M71L/S。简而言之,给小鼠皮下给药M71和M71L/S(20、10或5.0mg/kg)、CNTO3930(huIgG1同种型对照,20mg/kg)、CNTO8852(用于Fc突变变型的同种型对照,20mg/kg)或体积为10μl/g的PBS。24小时之后,给每只小鼠i.p.注射10μg的于PBS(SigmaD8357)中的自身产生的CHO细胞来源的重组人制瘤素Mw/0.1%小鼠血清白蛋白(SigmaA3559)或单独PBS-MSA媒介物对照(200μL总体积)。1小时之后,用CO2使小鼠麻醉并通过末端心脏穿刺收集血液。使各个血液样品在冰上凝结20分钟,然后以3500rpm旋转10-15分钟。将血清样品冷冻保存直至使用由对IL-6、MCP-1、嗜酸性粒细胞趋化因子、KC和IP-10特异的珠组成的定制Millipore鼠多路复用进行分析。同种型对照对由人制瘤素M诱导的细胞因子释放均没有任何影响。然而,M71和M71L/S均中和制瘤素M诱导的细胞因子释放,并且在效能或功效方面无明显差异。药代动力学分析在非末端食蟹猴药代动力学研究中比较M71和M71L/S的血清半衰期。该研究包括总共12只食蟹猴,并对每种抗体评价皮下(s.c.,n=3)和静脉内(i.v.,n=3)施用。抗体按3mg/kg剂量给药,并且该研究进行超过60天。在1小时(仅IV组)和6小时以及在第1、2、4、6、8、12、16、30、37、45和60天时抽取血样样品。将来自样品的血清在-80℃下冻存直至测试。使用在食蟹猴血清中针对MesoScaleDiscovery平台优化的ELISA确定血清中的抗体水平。生物素酰化捕获抗体为针对M71培育的抗独特型抗体(小鼠抗M71)。检测抗体为钌标记的抗人IgG,并且读出是MesoScaleDiscovery化学发光。该研究的结果示于图6A和B中。图6A示出来自i.v.给药的图线,其中M71的血清半衰期为15.21+/-3.0天,而M71L/S的半衰期为29.4+/-2.3天。类似的结果从s.c.给药获得(图6B),其中M71的血清半衰期为15.4+/-4天,而M71L/S的血清半衰期为32.0+/-5.9天。结果显示,相比于M71,M71L/St1/2增加约两倍。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 一株分泌抗平足蛋白单克隆抗体的杂交瘤细胞株及其应用的制作方法与工艺
- 一种分泌抗黄曲霉毒素B1单克隆抗体的杂交瘤细胞株及其应用的制造方法与工艺
- 可分泌抗C反应蛋白单克隆抗体的杂交瘤细胞、C反应蛋白检测试剂及其制备方法和应用与流程
- 鼠抗人CD61单克隆抗体杂交瘤细胞株、单克隆抗体及其制备方法和应用、流式检测试剂与流程
- 一种分泌抗总黄曲霉毒素单克隆抗体的杂交瘤细胞株及其应用的制造方法与工艺
- 一株抗苯乙醇胺A单克隆抗体杂交瘤细胞株K‑6及其应用的制造方法与工艺
- 一株分泌潮霉素B特异性单克隆抗体的杂交瘤细胞株ZXL‑1及其应用的制造方法与工艺
- 可分泌抗H‑FABP单克隆抗体的杂交瘤细胞、单克隆抗体及其制备方法和应用与流程
- 抗USP2a蛋白单克隆抗体杂交瘤细胞及其产生的抗USP2a单克隆抗体和应用的制造方法与工艺
- 便携式多种肿瘤标志物快速联检装置的制作方法