高效乙醇发酵菌的制作方法
本发明涉及在使用木质纤维素类生物质生产生物乙醇的过程中用于发酵糖化溶液的微生物。
特别涉及在使用木质纤维素类生物质生产生物乙醇的过程中能够由五碳糖(下文中也称为c5)以及六碳糖(下文中也称为c6)有效地生产乙醇的微生物。
背景技术:
生物乙醇作为由生物质产生的不会发生枯竭的可再生资源而倍受期待。另外,由于燃烧生物乙醇而产生的二氧化碳为碳中性的,因而据信通过进行生物乙醇的利用可抑制作为地球温室化的主要原因的二氧化碳的增加。
生物乙醇是将生物质发酵并进行蒸馏提纯乙醇制得的。为了提高生物乙醇的收率,需要由糖化溶液中生成大多数的醇。在生物乙醇生产的过程中通常使用的酵母无法将木糖、阿拉伯糖等五碳糖转换为醇,因而作为发酵原料,仅使用了六碳糖。
尽管根据原料而不同,但在代表性的生物质中被认为含有35%~45%的纤维素、25%~40%的半纤维素、15%~30%的木质素。因而,在不仅可利用六碳糖聚合而成的纤维素作为底物、而且还可利用主要含有作为五碳糖的木糖等的半纤维素作为底物时,可有效地产生乙醇。
木糖被认为是在生物质中大量含有的糖,其含量仅次于葡萄糖,有效地利用五碳糖是生物乙醇生产中的一大课题。
目前为止公开了少量利用木糖的技术,其是通过经基因重组而赋予木糖利用能力、或者利用木糖产生乙醇的微生物的利用等而实现的。
在专利文献1中公开了下述发明:将具有木糖转运体活性的基因导入到宿主细胞中,从而将木糖(c5)转换为木酮糖,整合在糖酵解系统的戊糖磷酸途径中,在发酵中加以利用。
在专利文献2中公开了利用被赋予了阿拉伯糖转运体的酵母来生成醇的技术。与专利文献1同样地将阿拉伯糖(c5)经阿拉伯糖醇、木酮糖而整合在糖酵解系统的戊糖磷酸途径中,在发酵中加以利用。
在非专利文献1中公开了通过将大肠杆菌来源的木糖同化基因重组到发酵单胞菌中而赋予木糖同化能力的内容。
在非专利文献2中记载了毕赤属酵母可利用木糖生产乙醇的内容。
现有技术文献
专利文献
专利文献1:日本特开2012-170422号公报
专利文献2:美国专利申请公开第2013/189788号说明书
非专利文献
非专利文献1:zhang,m.,etal.,science,1995.vol.267,pp.240-243.
非专利文献2:bicho,p.a.,etal.,appl.environ.microbiol.,1988,vol.54,pp.50-54.
技术实现要素:
发明所要解决的课题
但是,在专利文献1的发明中,是将季也蒙假丝酵母(candidaguilliermondii)来源的具有木糖转运体活性的蛋白质导入到作为宿主的酿酒酵母(saccharomycescerevisiae)中。即进行了外来基因的导入。
另外,在专利文献2的发明中,尽管转运基因不同,但也是向宿主中导入不同种基因的发明。
另外,非专利文献1所记载的技术是导入木糖同化基因的技术,与上述专利文献1和2的技术思想不同,但导入外来基因这一点不变。
因此,上述专利文献1和2、非专利文献1均需要采取用于实施联合国所采纳的“关于生物多样性公约的卡塔赫纳生物安全议定书”的封锁对策。从而需要用于保证生物安全的设施,因而利用该菌体生产乙醇从成本方面考虑是不利的。
另外,根据非专利文献2记载的技术,利用了毕赤属酵母,这由于野生型的毕赤属酵母的木糖利用性低,因而乙醇产生效率并不那么高。
本发明的课题在于在不导入外来基因的条件下得到乙醇产生效率高的发酵菌。
解决课题的手段
本发明涉及一种由五碳糖和六碳糖有效地产生乙醇的发酵菌,其特征在于,其在2014年11月19日(保藏日)以保藏编号nitebp-01964(下文中也称为bp-01964菌株)被保藏于专利微生物保藏中心(独立行政法人日本国家技术与评价研究所专利微生物保藏中心日本国〒292-0818千叶县木更津市上总鎌足2-5-8122号室)。
野生型季也蒙毕赤酵母具备木糖同化能力。但并不是说其具备对于生物乙醇生产为充分的木糖利用能力。bp-01964菌株是通过对季也蒙毕赤酵母进行菌株育种,选择出五碳糖的利用效率高的菌而得到的。其结果选择出了具备相对于亲株为约2倍的乙醇生产率的菌。
另外,本发明的特征在于,其是在上述季也蒙毕赤酵母(meyerozymaguilliermondii)株中导入了自克隆的转醛醇酶、醇脱氢酶、丙酮酸脱羧酶、木糖还原酶、木糖醇脱氢酶、转酮醇酶、甲酸脱氢酶基因而得到的。
在导入季也蒙毕赤酵母(meyerozymaguilliermondii)自身的酶基因时,依据卡塔赫纳法的封锁对策是不必要的。因而,其不需要用于生物安全的特殊设施,能够使用现有的设备。
附图说明
图1是示出bp-01964菌株与亲株(n株)的乙醇生成量的图。
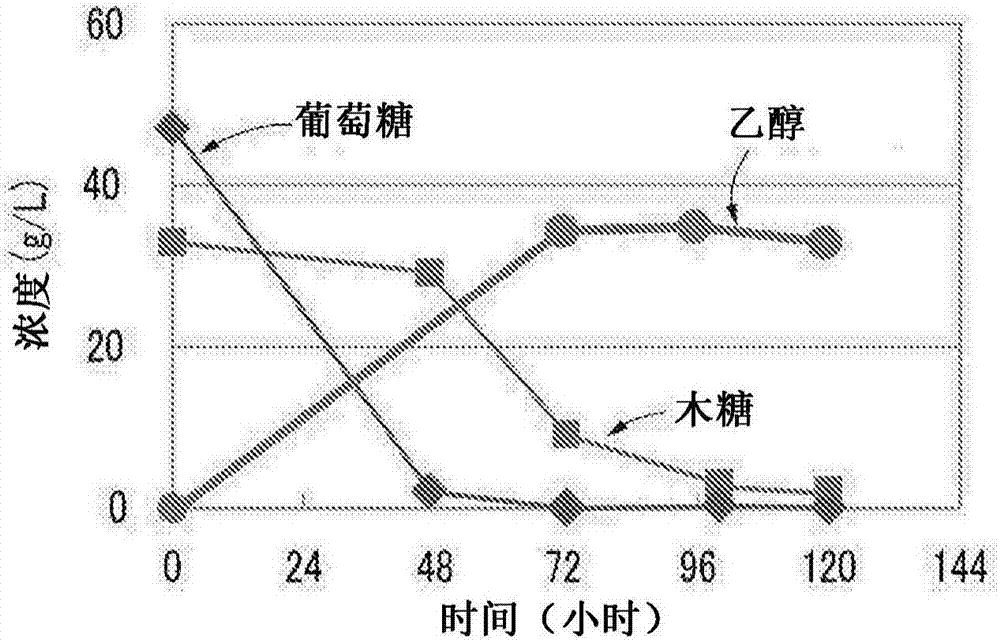
图2是示出浆液发酵中的葡萄糖、木糖同化能力的图。
图3是示出浆液发酵、澄清液发酵中的发酵收率的图。
图4是示出bp-01964菌株在玉米秸秆澄清液中的乙醇产生能力的图。
具体实施方式
作为子囊菌酵母的季也蒙毕赤酵母具备木糖同化能力。将季也蒙毕赤酵母的n株作为亲株,诱发变异并选择变异株进行育种,从而得到保藏编号nitebp-01964的菌。
通常,在变异的诱发中使用紫外线或放射线照射、n-亚硝基-n-乙基脲(enu)、甲磺酸乙酯(ems)等烷基化剂、brdu等碱类似化合物、硝基胺或亚硝基胍等亚硝基化合物等。在本发明中,通过uv照射或ems等化学药品的添加来诱发变异。
下面对菌株的获得方法进行说明。
实施例
1.菌株的分离
使用稻秆来源的糖液对季也蒙毕赤酵母的亲株进行培养。将熊谷产的稻秆浸入在等量的25%氨水中,于80℃浸入3小时后,将氨解吸出。将处理后的生物质利用10%naoh调整成ph4后,添加支顶孢属纤维素酶(アクレモニウムセルラーゼ,acremoniumcellulase)(meijiseikapharma公司制造),于50℃进行72小时酶糖化。将所制作的浆液通过压滤法进行固液分离,回收液体。使用该液体(下文中也称为澄清液),加入诱变剂进行19个月驯化培养,分选出发酵性能提高株。分选以一定时间后的乙醇量为基准来进行。将发酵性能高的菌株以保藏编号nitebp-01964保藏于独立行政法人日本国家技术与评价研究所专利微生物保藏中心。
2.菌株的性质
2.1乙醇产生能力
图1示出了bp-01964菌株相对于作为亲株的n株的乙醇产生量。对玉米秸秆进行稀硫酸处理,将所得到的糖化液利用naoh水溶液调整为ph6来使用,按照培养基的od600为2.0来添加同株的培养液,图1中示出了于30℃培养96小时后的培养液中的乙醇量。糖化溶液的葡萄糖为63.2g/l、木糖为34.5g/l。乙醇测定使用gc-fid(glsciences公司制造:gc390b)。
由图1可明确,得到了相对于野生型产生2倍以上乙醇的菌株。由于相对于野生株乙醇产生量提高,因而认为作为c5的木糖的同化能力提高。因此确认到了该菌株的葡萄糖和木糖同化能力。
其次,将稻秆利用氨水进行与上述氨处理同样的处理后,添加支顶孢属纤维素酶,于50℃进行72小时酶糖化,使用所制作的浆液进行发酵。
浆液发酵槽呈夹套结构,使温水在夹套部循环来进行温度调节。另外,在底部设置通气口,将规定量的通过了过滤器的空气持续由底部的通气口持续通入,在利用与马达联动的刮刀进行搅拌的同时进行发酵。
进行浆液中所含有的葡萄糖、木糖、乙醇的量的经时性变化的分析。关于葡萄糖、木糖,对浆液进行取样,对于通过离心得到的上清施以hplc测定来进行测定。关于乙醇,与上述同样地使用gc-fid(glsciences公司制造:gc390b)。将结果示于图2。
作为c6的葡萄糖先被消耗,其后,随着浆液中的葡萄糖减少,作为c5的木糖被消耗,产生乙醇。所得到的菌对c5、c6均具备同化能力,因而能够有效地产生乙醇。因此是在工业生产中也有用的菌株。
2.2浆液发酵能力、澄清液发酵能力
优选为在进行生物乙醇产生时不论使用浆液、澄清液中的哪一种的情况下均可有效地发酵的菌。因此,使用浆液、澄清液对发酵收率进行比较。发酵收率通过下式计算。
发酵收率=所得到的乙醇量(g/l)/发酵开始时的糖液中含有的葡萄糖+木糖量(g/l)/0.5114
如图3所示,得到了不论在浆液发酵还是在澄清液发酵中均能够发挥出同等性能的菌株。
2.3在玉米秸秆糖液中的乙醇产生能力
与稻秆同样地,玉米秸秆也是多用于生物乙醇产生的生物质。本菌株为使用由稻秆制造的澄清液进行驯化并分离出的菌株,但在玉米秸秆中也同样有效地产生生物乙醇。
将美国爱荷华州产的玉米秸秆浸在2倍量的3.7重量%硫酸水溶液中,于170℃处理10分钟。回复常温后,利用4m氢氧化钠水溶液制备成ph4,添加生物质糖化用酶(支顶孢属纤维素酶等、meijiseikapharma公司制造)于50℃进行72小时酶糖化。将所制作的浆液利用离心分离法进行固液分离,回收液体(下文中称为玉米秸秆澄清液)。
使用将玉米秸秆澄清液利用naoh水溶液调整成ph6而得到的液体,使用本菌株进行发酵,经时性地测定乙醇产生量。将结果示于图4。野性型(n株)在48小时后乙醇产生大致达到顶点,与此相对,本菌株直到72小时后仍继续产生乙醇,最终能够以约1.5倍左右的高产率产生乙醇。
另外,为了使本发明中得到的菌株更容易利用木糖,也可以导入下述酶的基因:作为戊糖磷酸途径酶的转醛醇酶、作为由乙醛生成乙醇的酶的醇脱氢酶、由丙酮酸生成作为醇脱氢酶底物的乙醛的丙酮酸脱羧酶。
例如,在基因导入时,可以进行下述过程。对于希望导入的基因及其终止子部(下文中称为基因+终止子部)进行pcr扩增。对导入中想要使用的启动子部进行pcr扩增。它们均由本发明中使用的菌株即季也蒙毕赤酵母的染色体进行pcr扩增。
将pcr扩增后的dna片段按照启动子、基因+终止子部的顺序使用融合(in-fusion)法克隆到大肠杆菌用的市售载体中。将克隆后的载体转化到大肠杆菌中,进行载体的扩增。利用限制酶由扩增后的载体切出启动子以及基因+终止子部,或者由扩增后的载体进行pcr扩增,从而得到同源重组用的dna片段。
将所得到的dna片段同源重组到菌株中,得到所期望的菌株。同源重组中使用电穿孔法。通过该方法导入基因时,能够在染色体上导入多个拷贝,因而能够增强所导入的酶的活性。
作为同源重组用dna片段,例如可以使用木糖还原酶的启动子、转醛醇酶+终止子。这是由于,通过使用在木糖同化时发挥功能的木糖还原酶的启动子,转醛醇酶被认为可有效地发挥出作用。
木糖还原酶的启动子具体地说使用下述序列编号1和序列编号2的引物进行扩增,转醛醇酶基因和终止子部分使用下述序列编号3和4的引物进行扩增。
序列编号1:aaggcttgggaactttcttt
序列编号2:agcaattgatgattaatttt
序列编号3:atgaccaattctcttgaaca
序列编号4:aaattgtgccgtgtcaaact
另外,也可以使用gapdh的启动子、醇脱氢酶+终止子。gapdh是存在于糖酵解系统中的强力的启动子,因而通过将其用作醇脱氢酶(其为糖酵解系统的酶)的启动子,据信可有效地发挥出作用。醇脱氢酶具有将乙醛转换为乙醇的作用,同时在nadh依赖性的情况下生成nad+,因而具有增强nad+依赖性木糖醇脱氢酶作用的作用。
gapdh的启动子具体地说使用下述序列编号5和序列编号6的引物进行扩增,醇脱氢酶基因和终止子部分使用下述序列编号7和8的引物进行扩增。
序列编号5:gttgtagcggaggctcaatt
序列编号6:tgtataatttaaatgtgggt
序列编号7:atgtcaattccagaatccat
序列编号8:caccttggctggaagtgctg
此外,为了使所得到的菌株对于制作糖液时所含有的乙酸等有机酸或糠醛等醛具有耐性,除了转醛醇酶、醇脱氢酶以外,还可以在上述启动子中的任一者的下游克隆导入丙酮酸脱羧酶、木糖还原酶、木糖醇脱氢酶、转酮醇酶、甲酸脱氢酶等酶。
丙酮酸脱羧酶、木糖还原酶、木糖醇脱氢酶、转酮醇酶、甲酸脱氢酶可以通过下述引物进行扩增。
丙酮酸脱羧酶
序列编号9:atgacagaaattactttggg
序列编号10:acaaacaaatgctgaaaac
木糖还原酶(xr)
序列编号11:atgtctattactttgaactc
序列编号12:cacaaaagttggaatcttgt
木糖醇脱氢酶(xdh)
序列编号13:atgactcccaacccatcttt
序列编号14:ctcgggaccatctataataa
转酮醇酶(tlk)
序列编号15:atgaccaccgacgactacga
序列编号16:aacagctagcaagtcctga
甲酸脱氢酶(fdh)
序列编号17:atgagtccagcaacaaaagg
序列编号18:tttcatcttgtgtctttcac
另外,通过该方法得到的菌株进行了基因导入,但为自克隆导入,因而在卡塔赫纳法上属于被看作非重组菌的范畴。
如上所示,在bp-01964菌株中,野生型季也蒙毕赤酵母的木糖同化能力通过育种而得到增强,即使在使用作为生物质的稻秆、玉米秸秆中的任一种的情况下也能够有效地产生乙醇。
序列表
<110>本田技研工业株式会社(hondamotorco.,ltd.)
<120>高效乙醇发酵菌(highefficiencyethanol-zymocyte)
<130>pct140400
<160>18
<170>patentinversion3.5
<210>1
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>1
aaggcttgggaactttcttt20
<210>2
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>2
agcaattgatgattaatttt20
<210>3
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>3
atgaccaattctcttgaaca20
<210>4
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>4
aaattgtgccgtgtcaaact20
<210>5
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>5
gttgtagcggaggctcaatt20
<210>6
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>6
tgtataatttaaatgtgggt20
<210>7
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>7
atgtcaattccagaatccat20
<210>8
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>8
caccttggctggaagtgctg20
<210>9
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>9
atgacagaaattactttggg20
<210>10
<211>19
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>10
acaaacaaatgctgaaaac19
<210>11
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>11
atgtctattactttgaactc20
<210>12
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>12
cacaaaagttggaatcttgt20
<210>13
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>13
atgactcccaacccatcttt20
<210>14
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>14
ctcgggaccatctataataa20
<210>15
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>15
atgaccaccgacgactacga20
<210>16
<211>19
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>16
aacagctagcaagtcctga19
<210>17
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>17
atgagtccagcaacaaaagg20
<210>18
<211>20
<212>dna
<213>人工序列
<220>
<223>季也蒙毕赤酵母(meyerozymaguilliermondii)
<400>18
tttcatcttgtgtctttcac20
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!