抑制酶组合物的AA9溶解性多糖单加氧酶催化的失活的方法与流程
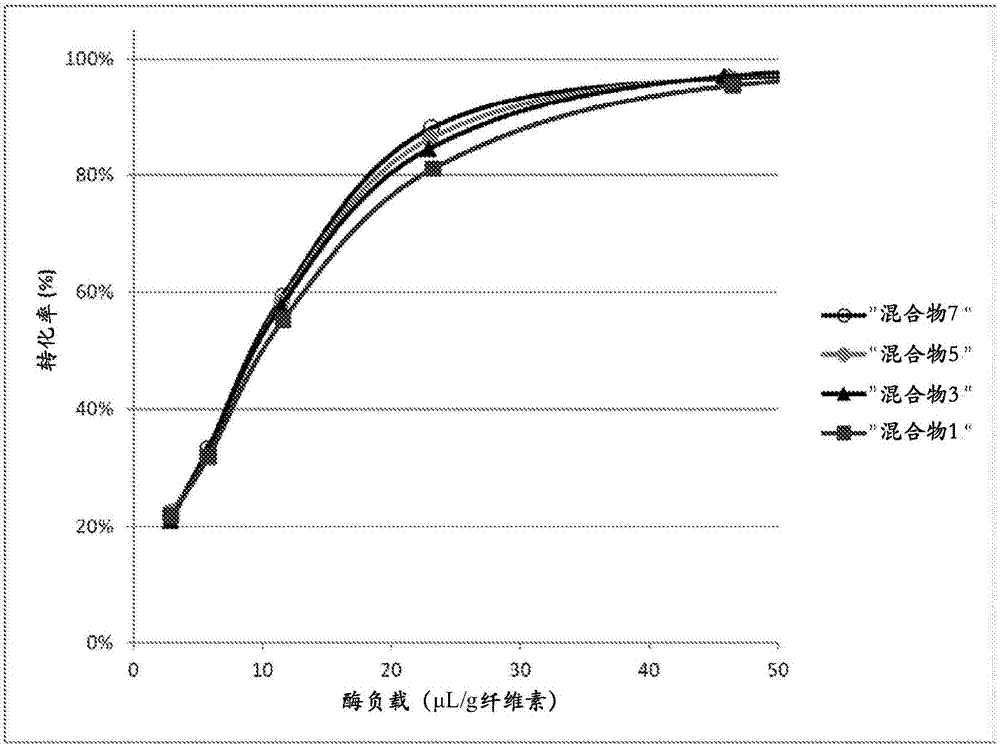
序列表的引用本申请含有一个计算机可读形式的序列表,将其通过引用结合在此。发明背景本发明涉及抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的失活的方法、用于增加酶组合物的产生的方法、以及用于稳定化酶组合物的方法。相关
技术领域:
的说明木质纤维素材料为产生化石燃料的可替代能源提供了有吸引力的平台。将木质纤维素材料(例如来自木质纤维素原料)转化成生物燃料具有以下优点:易于获得大量原料、避免燃烧或填埋材料的合意性、以及该生物燃料(例如乙醇)的清洁性。木材、农业残留物、草本作物以及城市固体废物已被认为是用于生物燃料产生的原料。一旦将木质纤维素材料糖化并转化成可发酵的糖,例如葡萄糖,那么这些可发酵的糖就可以被酵母发酵成生物燃料(如乙醇)。在过去十年里已经研发了新的且改进的酶和酶组合物并且已经使得预处理的纤维素材料的糖化变得更有效。但是,在本领域中对于进一步改进酶组合物存在需求。本发明提供抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的失活的方法、用于增加酶组合物的产生的方法、以及用于稳定酶组合物的方法。技术实现要素:本发明涉及抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的失活的方法,所述方法包括:将选自下组的一种或多种氧化还原酶添加至酶组合物中,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,其中该一种或多种添加的氧化还原酶抑制酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活。本发明还涉及用于增加酶组合物的产生的方法,所述方法包括:(a)在选自下组的一种或多种添加的氧化还原酶的存在下,发酵宿主细胞以产生该酶组合物,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活,并且其中与在该一种或多种氧化还原酶不存在下产生的酶组合物的量相比,在该一种或多种添加的氧化还原酶的存在下产生的酶组合物的量更高;并且任选地(b)回收该酶组合物。本发明还涉及用于稳定酶组合物的方法,该方法包括将选自下组的一种或多种氧化还原酶添加至该酶组合物中,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,并且其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活。附图说明图1显示用ph4.5的发酵液滤液1、3、5、和7(实例1)以及用ph3.5的发酵液滤液2、4、6、和8(实例2),在50℃和ph5.0下预处理的玉米穗轴和秸秆(pccs)水解测定(20g)5天的结果。图2显示在50℃和ph5.0下孵育6天之后,混合物1、3、5和7(ph4.5发酵,实例1)的荧光纤维素衰变(fcd)测定的结果。图3显示在ph5.0和50℃下孵育6天之后,混合物2、4、6和8(ph3.5发酵,实例2)的fcd测定的结果。图4a显示在4℃、25℃、40℃和50℃无菌储存4周之后,混合物1、3、5、和7的fcd测定的结果,并且图4b显示在4℃、25℃、40℃和50℃无菌储存4周之后,混合物2、4、6、和8的fcd测定的结果。图5显示,在发酵(混合物11和12)过程中添加过氧化氢酶,以及在发酵(混合物9和10)中不添加过氧化氢酶对在4℃、25℃、和40℃下储存4周之后性能的影响,在ph5.0和55℃持续5天通过fcd测定进行测量。图6显示通过以0%、0.1%、0.5%、1%和2%w/w过氧化氢酶蛋白质进行酶置换,在发酵之后,添加supreme过氧化氢酶对混合物13的影响,在ph5.0和55℃持续5天通过fcd测定进行测量。图7显示过滤的发酵液1-8(泳道1-8)的蛋白质印迹分析。泳道11-16分别代表发酵1(0%过氧化氢酶过量表达种子b)的从第2天至第7天每天样品的bca微板测定蛋白质标准化(1μg)的负载,而泳道17-22代表发酵5的等同样品(10%过氧化氢酶过表达种子b)。图8显示过滤的发酵液9(泳道1)、10(泳道2)、11(泳道3)和12(泳道4)的蛋白质印迹分析。未编号的泳道是以千道尔顿计的分子量标准品。定义乙酰木聚糖酯酶:术语“乙酰木聚糖酯酶”意指羧酸酯酶(ec3.1.1.72),其催化乙酰基自聚合木聚糖、乙酰化木糖、乙酰化葡萄糖、α-萘基乙酸酯、和对硝基苯基乙酸酯的水解。可以在包含0.01%tweentm20(聚氧乙烯脱水山梨糖醇单月桂酸酯)的50mm乙酸钠(ph5.0)中使用0.5mm对硝基苯基乙酸酯作为底物来测定乙酰木聚糖酯酶活性。将一个单位的乙酰木聚糖酯酶定义为在ph5、25℃下每分钟能够释放1微摩尔对硝基酚根阴离子的酶的量。等位基因变体:术语“等位基因变体”意指占用同一染色体基因座的基因的两个或更多个替代形式中的任一者。等位基因变异由突变天然产生,并且可以导致群体内的多态性。基因突变可以是沉默的(在所编码的多肽中没有改变)或可编码具有改变的氨基酸序列的多肽。多肽的等位基因变体是由基因的等位基因变体编码的多肽。α-l-阿拉伯呋喃糖苷酶:术语“α-l-阿拉伯呋喃糖苷酶”意指一种α-l-阿拉伯呋喃糖苷阿拉伯呋喃水解酶(ec3.2.1.55),其催化α-l-阿拉伯糖苷中的末端非还原性α-l-阿拉伯呋喃糖苷残基的水解。该酶对α-l-阿拉伯呋喃糖苷、包含(1,3)-和/或(1,5)-键的α-l-阿拉伯聚糖、阿拉伯糖基木聚糖以及阿拉伯半乳聚糖起作用。α-l-阿拉伯呋喃糖苷酶还被称为阿拉伯糖苷酶、α-阿拉伯糖苷酶、α-l-阿拉伯糖苷酶、α-阿拉伯呋喃糖苷酶、多糖α-l-阿拉伯呋喃糖苷酶、α-l-阿拉伯呋喃糖苷水解酶、l-阿拉伯糖苷酶、或α-l-阿拉伯聚糖酶。可以使用每ml的100mm乙酸钠(ph5)中5mg的中等粘度小麦阿拉伯糖基木聚糖(麦格酶国际爱尔兰股份有限公司(megazymeinternationalireland,ltd.))以总体积200μl在40℃下持续30分钟,接着通过hpx-87h柱层析(伯乐实验室有限公司(bio-radlaboratories,inc.))进行阿拉伯糖分析来确定α-l-阿拉伯呋喃糖苷酶活性。α-葡糖醛酸糖苷酶:术语“α-葡糖醛酸糖苷酶”意指一种α-d-葡糖苷酸葡糖醛酸水解酶(ec3.2.1.139),其催化α-d-葡糖苷酸水解成d-葡糖醛酸酯和醇。可以根据devries,1998,j.bacteriol.[细菌学杂志]180:243-249来确定α-葡糖醛酸糖苷酶活性。一个单位的α-葡糖醛酸糖苷酶等于能够在ph5、40℃下每分钟释放1微摩尔的葡糖醛酸或4-o-甲基葡糖醛酸的酶的量。辅助活性9多肽:术语“辅助活性9多肽”或“aa9多肽”意指分类为溶解性多糖单加氧酶(lyticpolysaccharidemonooxygenase)(quinlan等人,2011,proc.natl.acad.sci.usa[美国科学院院刊]108:15079-15084;phillips等人,2011,acschem.biol.[acs化学生物学]6:1399-1406;lin等人,2012,structure[结构]20:1051-1061)的多肽。根据henrissat,1991,biochem.j.[生物化学杂志]280:309-316以及henrissat和bairoch,1996,biochem.j.[生物化学杂志]316:695-696,aa9多肽之前被分类为糖苷水解酶家族61(gh61)。此类多肽在此被称为“aa9溶解性多糖单加氧酶”。aa9溶解性多糖单加氧酶通过具有纤维素分解活性的酶增强纤维素材料的水解。可以通过测量在以下条件下与具有无纤维素分解增强活性的相等的总蛋白负载的对照水解(1-50mg的纤维素分解蛋白/g的pcs中的纤维素)相比,由纤维素分解酶水解纤维素材料的还原糖的增加或纤维二糖与葡萄糖总量的增加来测定纤维素分解增强活性:1-50mg的总蛋白/g于预处理的玉米秸秆(pcs)中的纤维素,其中总蛋白由50%-99.5%w/w纤维素分解酶蛋白和0.5%-50%w/waa9多肽蛋白组成,在适合的温度(如40℃-80℃,例如40℃、45℃、50℃、55℃、60℃、65℃、70℃、75℃、或80℃)和适合的ph(如4-9,例如4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、或9.0)下持续1-7天。可以使用celluclasttm1.5l(诺维信公司,巴格斯瓦德(bagsvaerd),丹麦)和β-葡糖苷酶的混合物作为纤维素分解活性的来源来确定纤维素分解增强活性,其中该β-葡糖苷酶是以纤维素酶蛋白负载的至少2%-5%蛋白的重量存在的。在一方面,该β-葡糖苷酶是米曲霉β-葡糖苷酶(例如,根据wo02/095014,在米曲霉中重组产生的)。在另一方面,该β-葡糖苷酶是烟曲霉β-葡糖苷酶(例如,如在wo02/095014中描述的,在米曲霉中重组产生的)。纤维素分解增强活性还可通过以下来确定:在40℃下将aa9多肽与0.5%磷酸溶胀纤维素(pasc)、100mm乙酸钠(ph5)、1mmmnso4、0.1%没食子酸、0.025mg/ml的烟曲霉β-葡糖苷酶、以及0.01%x-100(4-(1,1,3,3-四甲基丁基)苯基-聚乙二醇)一起孵育24-96小时,接着测定从pasc释放的葡萄糖。还可以根据wo2013/028928确定高温组合物的纤维素分解增强活性。aa9溶解性多糖单加氧酶通过将达到相同的水解程度所需要的纤维素分解酶的量降低优选至少1.01倍,例如,至少1.05倍、至少1.10倍、至少1.25倍、至少1.5倍、至少2倍、至少3倍、至少4倍、至少5倍、至少10倍、或至少20倍,来增强由具有纤维素分解活性的酶催化的纤维素材料的水解。根据wo2008/151043或wo2012/122518,aa9溶解性多糖单加氧酶可以在可溶性活化二价金属阳离子(例如锰或铜)的存在下使用。aa9溶解性多糖单加氧酶还可以在二氧化合物、二环化合物、杂环化合物、含氮化合物、醌化合物、含硫化合物、或从预处理的纤维素材料或半纤维素材料(如预处理的玉米秸秆)获得的液体的存在下使用(wo2012/021394、wo2012/021395、wo2012/021396、wo2012/021399、wo2012/021400、wo2012/021401、wo2012/021408、以及wo2012/021410)。β-葡糖苷酶:术语“β-葡糖苷酶”意指β-d-葡糖苷葡糖水解酶(beta-d-glucosideglucohydrolase)(e.c.3.2.1.21),其催化末端非还原β-d-葡萄糖残基的水解,并释放β-d-葡萄糖。可以根据venturi等人,2002,j.basicmicrobiol.[基础微生物学杂志]42:55-66的程序使用对硝基苯基-β-d-吡喃葡萄糖苷作为底物来确定β-葡糖苷酶活性。一个单位的β-葡糖苷酶定义为在25℃、ph4.8下,在含有0.01%20的50mm柠檬酸钠中从作为底物的1mm对硝基苯基-β-d-吡喃葡萄糖苷中每分钟产生1.0微摩尔的对硝基苯酚阴离子。β-木糖苷酶:术语“β-木糖苷酶”意指β-d-木糖苷木糖水解酶(β-d-xylosidexylohydrolase)(e.c.3.2.1.37),其催化短β(1→4)-低聚木糖的外切水解,以将连续的d-木糖残基从非还原端移除。可以在包含0.01%20的100mm柠檬酸钠中,在ph5、40℃下,使用1mm对硝基苯基-β-d-木糖苷作为底物测定β-木糖苷酶活性。一个单位的β-木糖苷酶定义为在40℃、ph5下,在包含0.01%20的100mm柠檬酸钠中从1mm对硝基苯基-β-d-木糖苷每分钟产生1.0微摩尔的对硝基酚根阴离子。cdna:术语“cdna”意指可以通过从获得自真核或原核细胞的成熟的、剪接的mrna分子进行反转录而制备的dna分子。cdna缺乏可存在于对应基因组dna中的内含子序列。早先的初始rna转录本是mrna的前体,其在呈现为成熟的剪接的mrna之前要经一系列的步骤进行加工,包括剪接。过氧化氢酶:术语“过氧化氢酶”意指一种过氧化氢:过氧化氢氧化还原酶(e.c.1.11.1.6或e.c.1.11.1.21),其催化两个过氧化氢转化为氧和两个水。可以基于以下反应通过在240nm下监测过氧化氢的降解而确定过氧化氢酶活性:2h2o2→2h2o+o2在25℃下,在具有10.3mm底物(h2o2)的50mm磷酸盐(ph7)中进行该反应。用分光光度计监测16-24秒内的吸光度,这应该对应于从0.45至0.4的吸光度降低。可以将一个过氧化氢酶活性单位表示为在ph7.0和25℃下每分钟降解一微摩尔的h2o2。纤维二糖水解酶:术语“纤维二糖水解酶”意指一种1,4-β-d-葡聚糖纤维二糖水解酶(e.c.3.2.1.91和e.c.3.2.1.176),其催化纤维素、纤维寡糖、或任何包含β-1,4-连接葡萄糖的聚合物中的1,4-β-d-糖苷键的水解,从而从链的还原性末端(纤维二糖水解酶i)或非还原性末端(纤维二糖水解酶ii)释放纤维二糖(泰里(teeri),1997,生物技术趋势(trendsinbiotechnology)15:160-167;泰里等人,1998,生物化学学会会刊(biochem.soc.trans.)26:173-178)。可以根据lever等人,1972,anal.biochem.[分析生物化学]47:273-279;vantilbeurgh等人,1982,febsletters[欧洲生化学会联合会快报]149:152-156;vantilbeurgh和claeyssens,1985,febsletters[欧洲生化学会联合会快报]187:283-288;以及tomme等人,1988,eur.j.biochem.[欧洲生物化学杂志]170:575-581所描述的程序来确定纤维二糖水解酶活性。纤维素分解酶或纤维素酶:术语“纤维素分解酶”或“纤维素酶”意指一种或多种(例如,若干种)水解纤维素材料的酶。此类酶包括一种或多种内切葡聚糖酶、一种或多种纤维二糖水解酶、一种或多种β-葡糖苷酶、或其组合。用于测量纤维素分解酶活性的两种基本方法包括:(1)测定总纤维素分解酶活性,以及(2)测定单独的纤维素分解酶活性(内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶),如在张(zhang)等人,2006,生物技术进展(biotechnologyadvances)24:452-481中所述的。可使用不溶性底物,包括沃特曼(whatman)№1滤纸、微晶纤维素、细菌纤维素、藻类纤维素、棉花、预处理的木质纤维素等,测量总纤维素分解酶活性。最常见的总纤维素分解活性测定是将沃特曼№1滤纸用作底物的滤纸测定。该测定是由国际纯粹与应用化学联合会(iupac)建立的(ghose,1987,pureappl.chem.[纯粹与应用化学]59:257-68)。可以通过测量在以下条件下与未添加纤维素分解酶蛋白的对照水解相比,在一种或多种纤维素分解酶对纤维素材料的水解过程中产生/释放的糖的增加来测定纤维素分解酶活性:1-50mg的纤维素分解酶蛋白/g于预处理的玉米秸秆(pcs)中的纤维素(或其他预处理的纤维素材料),在适合的温度(如40℃-80℃,例如40℃、45℃、50℃、55℃、60℃、65℃、70℃、75℃、或80℃)和适合的ph(如4-9,例如,4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、或9.0)下持续3-7天。典型条件为:1ml反应,洗涤或未洗涤的pcs,5%不溶性固体(干重),50mm乙酸钠(ph5),1mmmnso4,50℃、55℃、或60℃,72小时,通过hpx-87h柱层析(伯乐实验室有限公司(bio-radlaboratories,inc.))进行糖分析。纤维素材料:术语“纤维素材料”意指包含纤维素的任何材料。生物质的初生细胞壁中的主要多糖是纤维素,第二丰富的是半纤维素,而第三丰富的是果胶。细胞停止生长后产生的次生细胞壁也含有多糖,并且它通过与半纤维素共价交联的聚合木质素得到强化。纤维素是脱水纤维二糖的均聚物,因此是线性β-(1-4)-d-葡聚糖,而半纤维素包括多种化合物,如具有一系列取代基以复杂支链结构存在的木聚糖、木葡聚糖、阿拉伯糖基木聚糖、以及甘露聚糖。尽管纤维素一般为多形态的,但发现其在植物组织中主要作为平行葡聚糖链的不溶性晶体基质存在。半纤维素通常氢键结合至纤维素以及其他半纤维素,这有助于稳定细胞壁基质。纤维素通常见于例如植物的茎、叶、壳、皮和穗轴或树的叶、枝和木材(wood)中。纤维素材料可以是,但不限于:农业残留物、草本材料(包括能源作物)、城市固体废物、纸浆和造纸厂废弃物、废纸、和木材(包括林业残留物)(参见,例如wiselogel等人,1995,在:handbookonbioethanol[生物乙醇手册](charlese.wyman编辑)中,第105-118页,taylor&francis[泰勒-弗朗西斯出版集团],华盛顿特区;wyman,1994,bioresourcetechnology[生物资源技术]50:3-16;lynd,1990,appliedbiochemistryandbiotechnology[应用生物化学与生物技术]24/25:695-719;mosier等人,1999,recentprogressinbioconversionoflignocellulosics[木质纤维素的生物转化的最近进展],在:advancesinbiochemicalengineering/biotechnology[生物化学工程/生物技术进展],t.scheper总编辑,第65卷,第23-40页,springer-verlag[施普林格出版公司],纽约)。在此应该理解的是,纤维素可以处于木素纤维素,在混合基质中含有木质素、纤维素、和半纤维素的植物细胞壁材料的形式。在一方面,该纤维素材料是任何生物质材料。在另一方面,该纤维素材料是木质纤维素(木质纤维素材料),该木质纤维素包含纤维素、半纤维素、以及木质素。在一个实施例中,该纤维素材料是农业残留物、草本材料(包括能源作物)、城市固体废物、纸浆和造纸厂废弃物、废纸或木材(包括林业残留物)。在另一个实施例中,该纤维素材料是芦竹、甘蔗渣、竹子、玉米穗轴、玉米纤维、玉米秸秆、芒属、稻秸、甘蔗秸秆、柳枝稷或麦秸。在另一个实施例中,该纤维素材料是山杨、桉树、冷杉、松树、白杨、云杉或柳树。在另一个实施例中,该纤维素材料是海藻纤维素、细菌纤维素、棉短绒、滤纸、微晶纤维素(例如,)、或经磷酸处理的纤维素。在另一个实施例中,该纤维素材料是水生生物质。如在此使用,术语“水生生物质”意指在水生环境中通过光合作用过程产生的生物质。水生生物质可以是藻类、挺水植物、浮叶植物、或沉水植物。可以按原样使用纤维素材料或可使用本领域已知的常规方法将纤维素材料进行预处理。在一个优选方面,对该纤维素材料进行预处理。内切葡聚糖酶:术语“内切葡聚糖酶”意指一种4-(1,3;1,4)-β-d-葡聚糖4-葡聚糖水解酶(e.c.3.2.1.4),其催化纤维素、纤维素衍生物(如羧甲基纤维素和羟乙基纤维素)、地衣多糖中的1,4-β-d-糖苷键和混合β-1,3-1,4葡聚糖如谷类β-d-葡聚糖或木葡聚糖以及包含纤维素组分的其他植物材料中的β-1,4键的内切水解。可以通过测量底物粘度的降低或通过还原糖测定所确定的还原性末端的增加来确定内切葡聚糖酶活性(zhang等人,2006,biotechnologyadvances[生物技术进展]24:452-481)。还可以根据以下文献中所述的程序,在ph5、40℃下,使用羧甲基纤维素(cmc)作为底物来确定内切葡聚糖酶活性:ghose,1987,pureandappl.chem.[纯粹与应用化学]59:257-268。阿魏酸酯酶:术语“阿魏酸酯酶”意指4-羟基-3-甲氧基肉桂酰基-糖水解酶(ec3.1.1.73),其催化4-羟基-3-甲氧基肉桂酰基(阿魏酰基)基团从酯化的糖(其在天然生物质底物中通常为阿拉伯糖)的水解,以产生阿魏酸酯(4-羟基-3-甲氧基肉桂酸酯)。阿魏酸酯酶(fae)也被称为阿魏酸酯酶(ferulicacidesterase)、羟基肉桂酰基酯酶、fae-iii、肉桂酸酯水解酶、faea、cinnae、fae-i、或fae-ii。可以在50mm乙酸钠(ph5.0)中,使用0.5mm对硝基苯基阿魏酸酯作为底物测定对阿魏酰酯酶活性。一个单位的阿魏酸酯酶等于,在ph5,25℃下,每分钟能够释放1微摩尔的对硝基酚根阴离子的酶的量。片段:术语“片段”意指多肽,该多肽使一个或多个(例如若干个)氨基酸从其成熟多肽的氨基和/或羧基末端缺失,其中该片段具有纤维素分解增强活性。在一方面,片段包含aa9溶解性多糖单加氧酶的成熟多肽的至少85%的氨基酸残基,例如至少90%的氨基酸残基、或至少95%的氨基酸残基。半纤维素分解酶或半纤维素酶:术语“半纤维素分解酶”或“半纤维素酶”意指水解半纤维素材料的一种或多种(例如,若干种)酶。参见例如,沙鲁姆(shallom)和沙哈姆(shoham),2003,微生物学当前观点(currentopinioninmicrobiology)6(3):219-228)。半纤维素酶是植物生物质的降解中的关键组分。半纤维素酶的实例包括但不限于,乙酰基甘露聚糖酯酶、乙酰基木聚糖酯酶、阿拉伯聚糖酶、阿拉伯呋喃糖苷酶、香豆酸酯酶、阿魏酸酯酶、半乳糖苷酶、葡糖醛酸糖苷酶、葡糖醛酸酯酶、甘露聚糖酶、甘露糖苷酶、木聚糖酶以及木糖苷酶。这些酶的底物半纤维素是支链和直链多糖的异质性组,其可通过氢键与植物细胞壁中的纤维素微纤维相结合,交联成坚固的网络。半纤维素还共价附接至木质素,从而与纤维素一起形成高度复杂的结构。半纤维素的可变结构和组织要求许多酶的协同作用以使其完全降解。半纤维素酶的催化模块是水解糖苷键的糖苷水解酶(gh),或是水解乙酸或阿魏酸侧基团的酯键的碳水化合物酯酶(ce)。这些催化模块,基于其一级结构的同源性,可分配到gh和ce家族。一些家族,具有总体上类似的折叠,可进一步归类为宗族(clan),以字母标记(例如,gh-a)。在碳水化合物活性酶(cazy)数据库中可得到这些以及其他碳水化合物活性酶的最翔实和更新的分类。可以根据ghose和bisaria,1987,pure&appi.chem.[纯粹与应用化学]59:1739-1752,在适合的温度如40℃-80℃,例如40℃、45℃、50℃、55℃、60℃、65℃、70℃、75℃、或80℃,以及适合的ph如4-9,例如4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、或9.0下测量半纤维素分解酶活性。半纤维素材料:该术语“半纤维素材料”意指包含半纤维素的任何材料。半纤维素包括木聚糖、葡糖醛酸木聚糖、阿拉伯糖基木聚糖、葡甘露聚糖以及木葡聚糖。这些多糖含有许多不同的糖单体。在半纤维素中的糖单体可以包括木糖、甘露糖、半乳糖、鼠李糖、以及阿拉伯糖。半纤维素含有大部分的d-戊糖糖类。在大多数情况下,木糖是以最大的量存在的糖单体,尽管在软木中甘露糖可以是最丰富的糖。木聚糖含有β-(1-4)-连接的木糖残基的主链。陆生植物的木聚糖是具有β-(1-4)-d-吡喃木糖主链的杂聚物,其通过短的碳水化合物链分支。它们包含d-葡糖醛酸或其4-o-甲基醚、l-阿拉伯糖、和/或不同的低聚糖,这些低聚糖由d-木糖、l-阿拉伯糖、d-或l-半乳糖、以及d-葡萄糖构成。可以将木聚糖类型的多糖分成同源木聚糖(homoxylan)和异源木聚糖(heteroxylan),包括葡糖醛酸木聚糖、(阿拉伯糖)葡糖醛酸木聚糖、(葡糖醛酸)阿拉伯糖基木聚糖、阿拉伯糖基木聚糖、以及复杂的异源木聚糖。参见,例如,ebringerova等人,2005,adv.polym.sci.[聚合物科学进展]186:1-67。半纤维素材料在此也称为“含木聚糖的材料”。半纤维素材料的来源基本上与在此描述的用于纤维素材料的那些来源相同。在一个优选方面,该半纤维素材料是木素纤维素(木质纤维素材料)。漆酶:术语“漆酶”意指催化以下反应的苯二醇:氧气氧化还原酶(e.c.1.10.3.2):1,2-或1,4-苯二醇+o2=1,2-或1,4-苯并半醌+2h2o。可以通过由漆酶将丁香醛连氮(4,4′-[连氮基双(甲基亚基)]双(2,6-二甲氧基苯酚))氧化为对应的醌4,4′-[偶氮双(甲基亚基])双(2,6-二甲氧基环已-2,5-二烯-1-酮)来确定漆酶活性。通过在530nm下吸光度的增加来检测该反应(如下所示)。在30℃下,在具有19μm底物(丁香醛连氮)和1g/l聚乙二醇(peg)6000的23mmmes(ph5.5)中进行该反应。将样品置于分光光度计中,并且在530nm下每15秒测量吸光度的变化,直至90秒。一个漆酶单位是在指定的分析条件下每分钟催化1微摩尔丁香醛连氮的转化的酶量。成熟多肽:术语“成熟多肽”意指在翻译和任何翻译后修饰(如n-末端加工、c-末端截短、糖基化作用、磷酸化作用等)之后处于其最终形式的多肽。本领域内公知,宿主细胞可以产生由相同多核苷酸表达的两种或更多种不同的成熟多肽(即,具有不同的c-末端和/或n-末端氨基酸)的混合物。成熟多肽编码序列:术语“成熟多肽编码序列”意指编码具有酶或生物活性的成熟多肽的多核苷酸。术语“成熟多肽编码序列”在此应理解为包括基因组dna序列的cdna序列或cdna序列的基因组dna序列。过氧化物酶:术语“过氧化物酶”意指将过氧化物(例如,过氧化氢)转化为较少氧化的种类(例如,水)的酶。在此应该理解的是,过氧化物酶涵盖过氧化物分解酶。在此将术语“过氧化物分解酶”定义为供体:催化还原底物(2e-)+roor’→氧化底物+roh+r’oh反应的过氧化物氧化还原酶(e.c.编号1.11.1.x,其中x=1-3、5、7-19、或21);如催化苯酚+h2o→醌+h2o反应的辣根过氧化物酶,和催化h2o2+h2o2→o2+2h2o反应的过氧化氢酶。除过氧化氢之外,其他过氧化物也可以被这些酶分解。在如以下所示的过氧化氢的存在下,可以通过测量由过氧化物酶氧化2,2’-连氮基-双(3-乙基苯噻唑啉-6-磺酸(abts)来确定过氧化物酶活性。反应产物abts氧化形成了在418nm下可以量化的蓝-绿颜色。h2o2+2abts还原+2h+→2h2o+2abts氧化在30℃下,在具有1.67mm底物(abts)、1.5g/lx-405、0.88mm过氧化氢、和大约0.040个单位的酶/ml的0.1m磷酸盐(ph7)中进行该反应。将样品置于分光光度计中,并且在418nm下从15秒直至60秒测量吸光度的变化。一个过氧化物酶单位可以表示为在指定的分析条件下每分钟催化1微摩尔过氧化氢所需要酶的量。预处理的纤维素材料或半纤维素材料:术语“预处理的纤维素材料或半纤维素材料”意指通过热处理和稀硫酸处理、碱预处理、中性预处理、或本领域已知的任何预处理从生物质得到的纤维素材料或半纤维素材料。预处理的玉米穗轴和玉米秸杆:术语“预处理的玉米穗轴和玉米秸杆”或“pccs”意指通过热和稀硫酸处理、碱预处理、中性预处理、或本领域已知的任何预处理从玉米穗轴和玉米秸杆得到的纤维素材料。预处理的玉米秸杆:术语“预处理的玉米秸杆”或“pcs”意指通过热和稀硫酸处理、碱预处理、中性预处理、或本领域已知的任何预处理从玉米秸杆得到的纤维素材料。序列一致性:用参数“序列一致性”来描述两个氨基酸序列之间或两个核苷酸序列之间的相关性。出于本发明的目的,使用如在emboss包(emboss:欧洲分子生物学开放软件套件(theeuropeanmolecularbiologyopensoftwaresuite),rice等人,2000,trendsgenet.[遗传学趋势]16:276-277)(优选5.0.0版本或更新版本)的needle程序中所实施的尼德尔曼-翁施(needleman-wunsch)算法(needleman和wunsch,1970,j.mol.biol.[分子生物学杂志]48:443-453)来确定两个氨基酸序列之间的序列一致性。使用的参数是空位开放罚分10、空位扩展罚分0.5以及eblosum62(blosum62的emboss版本)取代矩阵。将标记为“最长一致性”的尼德尔输出(使用-非简化(nobrief选项)获得)用作百分比一致性并且如下进行计算:(一致的残基×100)/(比对长度-比对中的空位总数)出于本发明的目的,使用如在emboss包(emboss:欧洲分子生物学开放软件套件,赖斯等人,2000,同上)(优选5.0.0版或更新版本)的尼德尔程序中所实施的尼德尔曼-翁施算法(尼德尔曼和翁施,1970,同上)来确定两个脱氧核糖核苷酸序列之间的序列一致性。使用的参数是空位开放罚分10、空位扩展罚分0.5,以及ednafull(ncbinuc4.4的emboss版本)取代矩阵。将标记为“最长一致性”的尼德尔输出(使用-非简化(nobrief选项)获得)用作百分比一致性并且如下进行计算:(相同脱氧核糖核苷酸×100)/(比对的长度-在比对中的空位总数)。严格条件:术语“非常低严格条件”是指对于长度为至少100个核苷酸的探针而言,遵循标准dna印迹程序,在42℃下在5xsspe、0.3%sds、200微克/ml剪切并变性的鲑鱼精子dna和25%甲酰胺中预杂交和杂交12至24小时。载体材料最终使用0.2xssc、0.2%sds,在45℃下洗涤三次,每次15分钟。术语“低严格条件”意指对于长度为至少100个核苷酸的探针而言,遵循标准dna印迹程序,在42℃下在5xsspe、0.3%sds、200微克/ml剪切并变性的鲑鱼精子dna和25%甲酰胺中预杂交和杂交12至24小时。载体材料最终使用0.2xssc、0.2%sds,在50℃下洗涤三次,每次15分钟。术语“中严格条件”意指对于长度为至少100个核苷酸的探针而言,遵循标准dna印迹程序,在42℃下在5xsspe、0.3%sds、200微克/ml剪切并变性的鲑鱼精子dna和35%甲酰胺中预杂交和杂交12至24小时。载体材料最终使用0.2xssc、0.2%sds,在55℃下洗涤三次,每次15分钟。术语“中-高严格条件”意指对于长度为至少100个核苷酸的探针而言,遵循标准dna印迹程序,在42℃下在5xsspe、0.3%sds、200微克/ml剪切并变性的鲑鱼精子dna和35%甲酰胺中预杂交和杂交12至24小时。载体材料最终使用0.2xssc、0.2%sds,在60℃下洗涤三次,每次15分钟。术语“高严格条件”意指对于长度为至少100个核苷酸的探针而言,遵循标准dna印迹程序,在42℃下在5xsspe、0.3%sds、200微克/ml剪切并变性的鲑鱼精子dna和50%甲酰胺中预杂交和杂交12至24小时。载体材料最终使用0.2xssc、0.2%sds,在65℃下洗涤三次,每次15分钟。术语“非常高严格条件”意指对于长度为至少100个核苷酸的探针而言,遵循标准dna印迹程序,在42℃下在5xsspe、0.3%sds、200微克/ml剪切并变性的鲑鱼精子dna和50%甲酰胺中预杂交和杂交12至24小时。载体材料最终使用0.2xssc、0.2%sds,在70℃下洗涤三次,每次15分钟。子序列:术语“子序列”意指使一个或多个(例如,若干个)核苷酸从成熟多肽编码序列的5’端和/或3’端缺失的多核苷酸,其中该子序列编码具有纤维素分解增强活性的片段。在一方面,子序列含有aa9溶解性多糖单加氧酶的成熟多肽编码序列的至少85%的核苷酸,例如至少90%的核苷酸或至少95%的核苷酸。超氧化物歧化酶:术语“超氧化物歧化酶”意指如以下:可替代地催化超氧化物(o2-)基歧化(或分配)为普通分子氧(o2)或过氧化物(h2o2)的酶(e.c.1.15.1.1):cu2+-sod+o2-→cu+-sod+o2cu+-sod+o2-+2h+→cu2+-sod+h2o2可以根据beauchamp和fridovich,1971,anal.biochem.[分析生物化学]44:276-287确定超氧化物歧化酶活性。含有木聚糖的材料:术语“含有木聚糖的材料”意指包含含有β-(1-4)连接的木糖残基主链的植物细胞壁多糖的任何材料。陆生植物的木聚糖是具有β-(1-4)-d-吡喃木糖主链的杂聚物,其通过短的碳水化合物链分支。它们包含d-葡糖醛酸或其4-o-甲基醚、l-阿拉伯糖、和/或不同的低聚糖,这些低聚糖由d-木糖、l-阿拉伯糖、d-或l-半乳糖、以及d-葡萄糖构成。可以将木聚糖类型的多糖分成同源木聚糖(homoxylan)和异源木聚糖(heteroxylan),包括葡糖醛酸木聚糖、(阿拉伯糖)葡糖醛酸木聚糖、(葡糖醛酸)阿拉伯糖基木聚糖、阿拉伯糖基木聚糖、以及复杂的异源木聚糖。参见,例如,ebringerova等人,2005,adv.polym.sci.[聚合物科学进展]186:1-67。在优选方面中,含有木聚糖的材料是木质纤维素。木聚糖降解活性或木聚糖分解活性:术语“木聚糖降解活性”或“木聚糖分解活性”意指水解包含木聚糖的材料的生物活性。用于测量木聚糖分解活性的两种基本方法包括:(1)测量总木聚糖分解活性,和(2)测量单独的木聚糖分解活性(例如,内切木聚糖酶、β-木糖苷酶、阿拉伯呋喃糖苷酶、α-葡糖醛酸糖苷酶、乙酰木聚糖酯酶、阿魏酸酯酶、以及α-葡糖醛酸酯酶)。木聚糖分解酶的测定的最近进展总结于若干出版物中,这些出版物包括biely和puchard,2006,journalofthescienceoffoodandagriculture[食品与农业科学杂志]86(11):1636-1647;spanikova和biely,2006,febsletters[欧洲生化学会联合会快报]580(19):4597-4601;herrimann等人,1997,生物化学杂志321:375-381。可以通过确定由不同类型的木聚糖,包括例如燕麦木聚糖、山毛榉木材木聚糖、和落叶松木材木聚糖形成的还原糖,或者通过光度确定从不同共价染色的木聚糖释放的染色的木聚糖片段,测量总木聚糖降解活性。常见的总木聚糖分解活性测定是基于由聚合4-o-甲基葡糖醛酸木聚糖产生还原糖,如描述于bailey等人,1992,interlaboratorytestingofmethodsforassayofxylanaseactivity[用于木聚糖酶活性测定的多个实验室测试方法],journalofbiotechnology[生物技术杂志]23(3):257-270中。木聚糖酶活性还可以在37℃下在0.01%x-100和200mm磷酸钠(ph6)中用0.2%azcl-阿拉伯糖基木聚糖作为底物来测定。将一个单位的木聚糖酶活性定义为在37℃、ph6下,在200mm磷酸钠(ph6)中从作为底物的0.2%azcl-阿拉伯糖基木聚糖每分钟产生1.0微摩尔天青蛋白。木聚糖降解活性可以是通过测量由一种或多种木聚糖降解酶在以下典型条件下造成的桦木木聚糖(西格玛化学有限公司(sigmachemicalco.,inc.))水解的增加来测定:1ml反应、5mg/ml底物(总固体)、5mg木聚糖分解蛋白质/g底物、50mm乙酸钠(ph5)、50℃、24小时,如里弗(lever),1972,分析生物化学(anal.biochem)47:273-279所述使用对羟基苯甲酸酰肼(phbah)测定进行糖分析。木聚糖酶:术语“木聚糖酶”意指1,4-β-d-木聚糖-木糖水解酶(1,4-β-d-xylan-xylohydrolase)(e.c.3.2.1.8),其催化木聚糖中1,4-β-d-木糖苷键的内水解。可以在37℃下在0.01%x-100和200mm磷酸钠(ph6)中用0.2%azcl-阿拉伯糖基木聚糖作为底物来测定木聚糖酶活性。将一个单位的木聚糖酶活性定义为在37℃、ph6下,在200mm磷酸钠(ph6)中从作为底物的0.2%azcl-阿拉伯糖基木聚糖每分钟产生1.0微摩尔天青蛋白。在此提及“约”一个数值或参数包括指向那个数值或参数本身的方面。例如,提及“约x”的描述包括方面“x”。如在此和所附权利要求书中所使用,单数形式“一种/个”、“或”以及“该”包括复数指示物,除非上下文以另外的方式清楚表明。应理解的是,在此描述的本发明的这些方面包括“由方面组成”和/或“基本由方面组成”。除非另外定义或由背景清楚指示,否则在此所用的全部技术与科学术语具有如本发明所属领域的普通技术人员通常理解的相同含义。发明详述本发明涉及抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的失活的方法,所述方法包括:将选自下组的一种或多种氧化还原酶添加至酶组合物中,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,其中该一种或多种添加的氧化还原酶抑制酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活。本发明还涉及用于增加酶组合物的产生的方法,所述方法包括:(a)在选自下组的一种或多种添加的氧化还原酶的存在下,发酵宿主细胞以产生该酶组合物,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活,并且其中与在该一种或多种氧化还原酶不存在下产生的酶组合物的量相比,在该一种或多种添加的氧化还原酶的存在下产生的酶组合物的量更高;并且任选地(b)回收该酶组合物。在一方面,向发酵中添加该一种或多种添加的氧化还原酶。在另一方面,该一种或多种添加的氧化还原酶是由宿主细胞重组产生的。在另一方面,该一种或多种添加的氧化还原酶是通过重组细胞与第二宿主细胞的共培养重组产生的。在另一方面,将该一种或多种添加的氧化还原酶添加至发酵中,并且该一种或多种添加的氧化还原酶是由宿主细胞重组产生的。在另一方面,将该一种或多种添加的氧化还原酶添加至发酵中,并且该一种或多种添加的氧化还原酶是通过重组细胞与第二宿主细胞的共培养重组产生的。在另一方面,该一种或多种添加的氧化还原酶是由宿主细胞重组产生的,以及通过将重组细胞与第二宿主细胞共培养重组产生的。在另一方面,将该一种或多种添加的氧化还原酶添加至发酵中,该一种或多种添加的氧化还原酶是由宿主细胞重组产生的,以及通过重组细胞与第二宿主细胞的共培养重组产生的。本发明还涉及用于稳定酶组合物的方法,该方法包括将选自下组的一种或多种氧化还原酶添加至该酶组合物中,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,并且其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活。本发明允许大量产生aa9溶解性多糖单加氧酶,同时抑制酶组合物的组分的aa9溶解性多糖单加氧酶催化的失活。不受任何理论束缚,例如,过氧化氢酶将通过aa9酶产生的过氧化氢转化为水和氧,阻断可以修饰蛋白质(包括酶组合物的酶组分)的活性氧类的形成。然后可以去稳定或灭活这些通过活性氧类修饰的蛋白质。这些修饰的蛋白质也可以被可以存在于酶组合物中的蛋白酶降解。抑制酶组合物的组分的aa9溶解性多糖单加氧酶催化的失活导致在发酵和回收结束时更高质量的酶组合物。由于在较高ph(例如ph4.5)下可以用过氧化氢酶进行抑制,所以可以在产生更多蛋白质的条件下(而不是在较低ph下)进行发酵。而且,用过氧化氢酶进行抑制保证了更稳定的酶组合物,因为对于可以存在于酶组合物中的蛋白酶,未修饰的酶可能更稳定。在一方面,与不存在一种或多种添加的氧化还原酶相比,在存在一种或多种添加的氧化还原酶下,对该aa9溶解性多糖单加氧酶催化的失活的抑制更高。在一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶)抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的至少1%、至少2%、至少3%、至少4%、至少5%、至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、或至少100%的失活。抑制酶组合物的组分的aa9溶解性多糖单加氧酶催化的失活可以导致从纤维素材料糖化的可发酵糖(例如葡萄糖)的更高产率。可以根据wo2013/028928进行糖化。在一方面,可发酵糖(例如葡萄糖)的产量增加至少1%、至少2%、至少3%、至少4%、至少5%、至少10%、至少15%、或至少20%。在另一方面,氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶)的存在使活性酶组合物或其活性组分的产量增加至少1%、至少2%、至少3%、至少4%、至少5%、至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、或至少100%。在另一方面,与不含有一种或多种氧化还原酶的酶组合物相比,用一种或多种氧化还原酶稳定的酶组合物在25℃持续4周具有至少1%、至少2%、至少3%、至少5%、至少7%、至少9%、至少10%、至少15%、至少20%、至少40%、至少60%、至少80%、或至少100%的更高稳定性(酶活性保留)。在另一方面,与不含有一种或多种氧化还原酶的酶组合物相比,用一种或多种氧化还原酶稳定的酶组合物在40℃持续4周具有至少1%、至少2%、至少3%、至少5%、至少7%、至少9%、至少10%、至少12%、至少15%、至少20%、至少40%、至少60%、至少80%、或至少100%的更高稳定性。在另一方面,与不含有一种或多种氧化还原酶的酶组合物相比,用一种或多种氧化还原酶稳定的酶组合物在50℃持续4周具有至少1%、至少2%、至少3%、至少5%、至少7%、至少9%、至少10%、至少15%、至少20%、至少40%、至少60%、至少80%、或至少100%的更高稳定性。aa9溶解性多糖单加氧酶aa9溶解性多糖单加氧酶可以是任何aa9溶解性多糖单加氧酶。该aa9溶解性多糖单加氧酶对于从中衍生或分离酶组合物的菌株(如黑曲霉、米曲霉、卢克诺文思金孢子菌(chrysosporiumlucknowense)(嗜热毁丝霉)、廉孢霉、特异腐质酶、埃默森篮状菌、或里氏木霉的菌株)是天然的或外来的。在实施例中,该aa9溶解性多糖单加氧酶是重组aa9多肽。在另一个实施例中,该aa9溶解性多糖单加氧酶与酶组合物的宿主细胞来源不同,例如不是木霉属的来源(如不是里氏木霉来源)。在实施例中,该aa9溶解性多糖单加氧酶是作为酶组合物的一部分重组产生的,例如通过产生酶组合物的里氏木霉宿主细胞产生。aa9溶解性多糖单加氧酶的实例包括但不限于来自以下的aa9溶解性多糖单加氧酶:梭孢端梗霉(acrophialophorafusispora)(wo2013/043910)、棘孢曲霉(wo2012/030799)、烟曲霉(wo2010/138754)、aurantiporusalborubescens(wo2012/122477)、嗜热毛壳菌(wo2012/101206)、瘤孢棒囊菌(corynascussepedonium)(wo2013/043910)、特异腐质酶(wo2012/146171)、樟绒枝霉(malbrancheacinnamomea)(wo2012/101206)、嗜热毁丝酶(wo2009/085935、wo2009/085859、wo2009/085864、wo2009/085868、和wo2009/033071)、嗜松青霉(wo2011/005867)、青霉属物种(wo2011/041397和wo2012/000892)、托姆青霉(wo2012/122477)、埃默森篮状菌(wo2012/000892)、雷塞特纳斯篮状菌(talaromycesleycettanus,wo2012/101206)、柄篮状菌(wo2012/135659)、嗜热篮状菌(wo2012/129697和wo2012/130950)、橙色嗜热子囊菌(wo2005/074656和wo2010/065830)、甲壳嗜热子囊菌(wo2011/041504)、嗜热子囊菌属物种(wo2011/039319)、疏棉状嗜热丝孢菌(wo2012/113340、wo2012/129699、wo2012/130964、和wo2012/129699)、土生梭孢霉(wo2005/074647、wo2008/148131、和wo2011/035027)、里氏木霉(wo2007/089290和wo2012/149344)、和褐孢长毛盘菌(wo2012/122477)。aa9溶解性多糖单加氧酶的非限制性实例是来自以下的aa9溶解性多糖单加氧酶:梭孢端梗霉(acrophialophorafusispora)(geneseqp:bam80382);棘孢曲霉(geneseqp:azt94039,geneseqp:azt94041,geneseqp:azt94043,geneseqp:azt94045,geneseqp:azt94047,geneseqp:azt94049,geneseqp:azt94051);烟曲霉(geneseqp:aym96878);霉白曲霉(geneseqp:bbe80792);aurantiporusalborubescens(geneseqp:azz98498,geneseqp:azz98500);嗜热毛壳菌(geneseqp:azy42252);瘤孢棒囊菌(corynascussepedonium)(geneseqp:bam80384,geneseqp:bam80386);特异腐质酶(geneseqp:bae45292,geneseqp:bae45294,geneseqp:bae45296,geneseqp:bae45298,geneseqp:bae45300,geneseqp:bae45302,geneseqp:bae45304,geneseqp:bae45306,geneseqp:bae45308,geneseqp:bae45310,geneseqp:bae45312,geneseqp:bae45314,geneseqp:bae45316,geneseqp:bae45318,geneseqp:bae45320,geneseqp:bae45322,geneseqp:bae45324,geneseqp:bae45326,geneseqp:bae45328,geneseqp:bae45330,geneseqp:bae45332,geneseqp:bae45334,geneseqp:bae45336,geneseqp:bae45338,geneseqp:bae45340,geneseqp:bae45342,geneseqp:bae45344);樟绒枝霉(malbrancheacinnamomea)(geneseqp:azy42250);嗜热毁丝酶(geneseqp:axd75715,geneseqp:axd75717,geneseqp:axd58945,geneseqp:axd80944,geneseqp:axf00393);青霉属物种(geneseqp:azg65226);埃默森青霉菌(geneseqp:bam92736);樟绒枝霉(malbrancheacinnamomea)(geneseqp:bao18037,geneseqp:bao18039,geneseqp:bao18041,geneseqp:bao18043,geneseqp:bao18045,geneseqp:bao18047,geneseqp:bao18049,geneseqp:bao18051,geneseqp:bao18053);弗格斯毁丝酶(geneseqp:bao17567,geneseqp:bao17569,geneseqp:bao17571,geneseqp:bao17573,geneseqp:bao17575,geneseqp:bao17577,geneseqp:bao17579,geneseqp:bao17581,geneseqp:bao17583,geneseqp:bao17585,geneseqp:bao17587,geneseqp:bao17589,geneseqp:bao17591,geneseqp:bao17593,geneseqp:bao17595,geneseqp:bao17597);嗜松青霉(geneseqp:ayn30445);托姆青霉(geneseqp:azz98506);埃默森篮状菌(geneseqp:azr89286);雷塞特纳斯篮状菌(geneseqp:azy42258);柄篮状菌(geneseqp:bad71945);嗜热篮状菌(geneseqp:baa95296,geneseqp:baa22810);甲壳嗜热子囊菌(geneseqp:azg67666,geneseqp:azg67668,geneseqp:azg67670);嗜热子囊菌属物种(geneseqp:azg48808);橙色嗜热子囊菌(geneseqp:azj19467,geneseqp:ayd12322);里氏木霉(geneseqp:afy26868,geneseqp:baf28697);疏棉状嗜热丝孢菌(geneseqp:azz14902,geneseqp:azz14904,geneseqp:azz14906);土生梭孢霉(geneseqp:aeb90517,geneseqp:aeb90519,geneseqp:aeb90521,geneseqp:aeb90523,geneseqp:aeb90525,geneseqp:aum21652,geneseqp:azg26658,geneseqp:azg26660,geneseqp:azg26662,geneseqp:azg26664,geneseqp:azg26666,geneseqp:azg26668,geneseqp:azg26670,geneseqp:azg26672,geneseqp:azg26674,geneseqp:azg26676,geneseqp:azg26678);以及褐孢长毛盘菌(geneseqp:azz98502,geneseqp:azz98504)。将登录号以其全部内容结合在此。在一方面,该aa9溶解性多糖单加氧酶与在此披露的aa9溶解性多糖单加氧酶的成熟多肽具有至少60%,例如至少65%、至少70%、至少75%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、或100%的序列一致性,该成熟多肽具有aa9溶解性多糖单加氧酶活性。在另一方面,aa9溶解性多糖单加氧酶的氨基酸序列与在此披露的aa9溶解性多糖单加氧酶的成熟多肽相差多达10个氨基酸,例如1个、2个、3个、4个、5个、6个、7个、8个、9个、或10个。在另一方面,该aa9溶解性多糖单加氧酶包含在此披露的aa9溶解性多糖单加氧酶的氨基酸序列,或由其组成。在另一方面,该aa9溶解性多糖单加氧酶包含在此披露的aa9溶解性多糖单加氧酶的成熟多肽,或由其组成。在另一个实施例中,该aa9溶解性多糖单加氧酶是在此披露的aa9溶解性多糖单加氧酶的等位基因变体。在另一方面,该aa9溶解性多糖单加氧酶是含有在此披露的aa9溶解性多糖单加氧酶的成熟多肽的至少85%的氨基酸残基(例如至少90%的氨基酸残基或至少95%的氨基酸残基)的片段。在另一方面,该aa9溶解性多糖单加氧酶由以下多核苷酸编码,该多核苷酸在非常低、低、中、中-高、高或非常高严格条件下与在此披露的aa9溶解性多糖单加氧酶的成熟多肽编码序列或其全长补体杂交(sambrook等人,1989,见上文)。可以使用编码aa9溶解性多糖单加氧酶的多核苷酸、或其子序列,以及aa9溶解性多糖单加氧酶的多肽、或其片段来设计核酸探针以根据本领域熟知的方法鉴定并克隆编码来自不同属或种的菌株的aa9溶解性多糖单加氧酶的dna。具体而言,此类探针可以用于与感兴趣的细胞的基因组dna或cdna杂交,如在上文所述。出于本发明的目的,杂交是指在非常低至非常高严格条件下多核苷酸杂交到标记的核酸探针上。在这些条件下与该核酸探针杂交的分子可以使用例如x-射线胶片或本领域中已知的任何其他检测手段进行检测。在一方面,核酸探针是aa9溶解性多糖单加氧酶的成熟多肽编码序列。在另一方面,该核酸探针是编码全长aa9溶解性多糖单加氧酶;其成熟多肽;或其片段的多核苷酸。在另一方面,该aa9溶解性多糖单加氧酶由与在此披露的aa9溶解性多糖单加氧酶的成熟多肽编码序列具有至少60%,例如至少65%、至少70%、至少75%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、或100%序列一致性的多核苷酸编码。该aa9溶解性多糖单加氧酶可以是杂合多肽,其中一种多肽的区域融合在另一种多肽、或融合多肽或可裂解的融合多肽的区域的n-末端或c-末端,其中另一种多肽融合在aa9溶解性多糖单加氧酶的n-末端或c-末端,如在此所述。该aa9溶解性多糖单加氧酶可以从任何属的微生物获得。出于本发明的目的,如在此与给出的来源结合使用的,术语“从…获得”应当意指由多核苷酸编码的aa9溶解性多糖单加氧酶是由该来源或由其中已经插入来自该来源的多核苷酸的菌株产生的。在一个实施例中,该aa9溶解性多糖单加氧酶是胞外分泌的。该aa9溶解性多糖单加氧酶可以是细菌aa9溶解性多糖单加氧酶。例如,该aa9溶解性多糖单加氧酶可以是革兰氏阳性细菌多肽,如芽孢杆菌属、梭菌属、肠球菌属、土芽孢杆菌属、乳杆菌属、乳球菌属、海洋芽孢杆菌属、葡萄球菌属、链球菌属、或链霉菌属aa9溶解性多糖单加氧酶;或革兰氏阴性细菌多肽,如弯曲杆菌属、大肠杆菌、黄杆菌属、梭杆菌属、螺杆菌属、泥杆菌属、奈瑟氏菌属、假单胞菌属、沙门菌属或脲原体属aa9溶解性多糖单加氧酶。在一个实施例中,该aa9溶解性多糖单加氧酶是嗜碱芽孢杆菌(bacillusalkalophilus)、解淀粉芽孢杆菌(bacillusamyloliquefaciens)、短芽孢杆菌(bacillusbrevis)、环状芽孢杆菌(bacilluscirculans)、克劳氏芽孢杆菌(bacillusclausii)、凝结芽孢杆菌(bacilluscoagulans)、坚硬芽孢杆菌(bacillusfirmus)、灿烂芽孢杆菌(bacilluslautus)、迟缓芽孢杆菌(bacilluslentus)、地衣芽孢杆菌(bacilluslicheniformis)、巨大芽孢杆菌(bacillusmegaterium)、短小芽孢杆菌(bacilluspumilus)、嗜热脂肪芽孢杆菌(bacillusstearothermophilus)、枯草芽孢杆菌(bacillussubtilis)、或苏云金芽孢杆菌(bacillusthuringiensis)aa9溶解性多糖单加氧酶。该aa9溶解性多糖单加氧酶可以是真菌的aa9溶解性多糖单加氧酶。例如,该aa9溶解性多糖单加氧酶可以是酵母aa9溶解性多糖单加氧酶,如假丝酵母属、克鲁弗酵母属、毕赤酵母属、酵母属、裂殖酵母、或耶氏酵母属aa9溶解性多糖单加氧酶;或者丝状真菌aa9溶解性多糖单加氧酶,如枝顶孢霉属、端梗孢属、伞菌属、链格孢属、曲霉属、aurantiporus、短梗霉属、葡萄座腔菌属(botryospaeria)、保加利亚(bulgaria)、拟蜡菌属、毛喙壳属、金孢子菌属、麦角菌属、旋孢腔菌属、鬼伞属、乳白蚁属、棒囊壳属(corynascus)、隐丛赤壳菌属、隐球菌属、色二孢属、黑耳属、线黑粉酵母属、镰孢属、赤霉属、全鞭毛虫属、腐质霉属、耙齿菌属、香菇属、蘑燕属、小腔球菌属、梨孢菌属、黑果菌属(melanocarpus)、畸枝霉属(malbranchea)、多孔菌属、毛霉属、毁丝霉属、新美鞭菌属、脉孢菌属、拟青霉属、青霉菌属、平革菌属、瘤胃壶菌属、poitrasia、假黑盘菌属、假披发虫属、根毛霉菌属、裂褶菌属、柱顶孢属、荚孢腔菌属、踝节菌属、嗜热子囊菌属、嗜热真菌属、梭孢壳霉属、弯颈霉属、木霉属、长毛盘菌属、轮枝孢属、valsaria、小包脚菇属或炭角菌属aa9溶解性多糖单加氧酶。在另一个实施例中,该aa9溶解性多糖单加氧酶是卡尔酵母(saccharomycescarlsbergensis)、酿酒酵母(saccharomycescerevisiae)、糖化酵母(saccharomycesdiastaticus)、道格拉氏酵母(saccharomycesdouglasii)、克鲁弗酵母(saccharomyceskluyveri)、诺地酶母(saccharomycesnorbensis)、或卵形酵母(saccharomycesoviformis)aa9溶解性多糖单加氧酶。在另一个实施例中,该aa9溶解性多糖单加氧酶是解纤维枝顶孢霉、梭孢端梗霉(acrophialophorafusispora)、棘孢曲霉、泡盛曲霉、臭曲霉、烟曲霉、日本曲霉、迟缓曲霉(aspergilluslentulus)、构巢曲霉、黑曲霉、霉白曲霉、米曲霉、土曲霉、aurantiporusalborubescens、胶陀螺(bulgariainquinans)、嗜热毛壳菌、狭边金孢子菌、嗜角质金孢子菌、卢克诺文思金孢子菌(chrysosporiumlucknowense)、粪状金孢子菌、毡金孢子菌、昆士兰金孢子菌、热带金孢子菌、带纹金孢子菌、瘤孢棒囊菌(corynascussepedonium)、嗜热棒囊孢菌壳、fennellianivea、杆孢状镰孢、禾谷镰孢、库威镰孢、黄色镰孢菌、禾谷镰孢菌、禾赤镰孢、异孢镰孢、长柄镰孢(fusariumlongipes)、合欢木镰孢、尖孢镰孢菌、多枝镰孢、粉红镰孢菌、接骨木镰孢、肤色镰孢、拟枝孢镰孢菌、硫色镰孢菌、圆镰孢、拟丝孢镰孢菌、镶片镰孢菌、灰腐质霉、特异腐质霉、柔毛腐质霉、白囊耙齿菌、绒柄香菇(lentinussimilis)、樟绒枝霉(malbrancheacinnamomea)、米黑毛霉、嗜热毁丝霉、粗糙脉孢菌、胶囊青霉、埃默森青霉菌、绳状青霉菌、嗜松青霉、产紫青霉、暗边青霉(penicilliumsoppii)、托姆青霉、黄孢原毛平革菌、sporormiafimetaria、丝衣霉状篮状菌(talaromycesbyssochlamydoides)、埃默森篮状菌、雷塞特纳斯篮状菌、柄篮状菌、嗜热篮状菌、橙色嗜热子囊菌、甲壳嗜热子囊菌、疏棉状嗜热丝孢菌、无色梭孢壳、成层梭孢壳菌、白毛梭孢壳、澳洲梭孢壳、粪梭孢壳、小孢梭孢壳、卵孢梭孢壳、秘鲁梭孢壳、毛梭孢壳、瘤孢梭孢壳、耐热梭孢壳、土生梭孢壳、深绿木霉、哈茨木霉、康宁木霉、长枝木霉、里氏木霉、土星孢木霉(trichodermasaturnisporum)、绿色木霉、或valsariarubricosaaa9溶解性多糖单加氧酶。将理解的是,对于以上提到的物种,本发明涵盖完全状态和不完全状态(perfectandimperfectstates)二者、以及其他分类学等效物,例如无性型,而不管它们已知的物种名称。本领域的技术人员将容易地识别适当等同物的身份。这些物种的菌株可容易地在许多培养物保藏中心为公众所获得,如美国典型培养物保藏中心(atcc)、德国微生物和细胞培养物保藏中心(deutschesammlungvonmikroorganismenundzellkulturengmbh,dsmz)、荷兰菌种保藏中心(centraalbureauvoorschimmelcultures,cbs)以及美国农业研究服务专利培养物保藏中心北方地区研究中心(agriculturalresearchservicepatentculturecollection,northernregionalresearchcenter,nrrl)。可以使用以上提到的探针从其他来源,包括从自然界(例如,土壤、堆肥、水等等)分离的微生物或直接从自然材料(例如,土壤、堆肥、水等等)获得的dna样品鉴定和获得该aa9溶解性多糖单加氧酶。用于直接地从自然生活环境分离微生物和dna的技术在本领域是熟知的。然后可通过在另一种微生物或混合dna样本的基因组dna或cdna文库中类似地进行筛选来获得编码aa9溶解性多糖单加氧酶的多核苷酸。一旦用一种或多种探针检测到编码aa9溶解性多糖单加氧酶的多核苷酸,就可以通过使用本领域普通技术人员已知的技术分离或克隆该多核苷酸(参见例如,sambrook等人,1989,见上文)。在实施例中,aa9溶解性多糖单加氧酶构成该酶组合物的从0.1%-25%,如0.5%-20%、0.5%-15%、0.5%-10%或0.5%-7%。在另一个实施例中,酶组合物中的aa9溶解性多糖单加氧酶的量是约1g至约1000g,如约1g至约200g、约1g至约100g、约1g至约50g、约1g至约20g、约1g至约15g、约1g至约10g、约1g至约7g或约1g至约4g/g的酶组合物。氧化还原酶在本发明的方法中,氧化还原酶可以是过氧化氢酶、漆酶、过氧化物酶、超氧化物歧化酶、或其组合。在一方面,该一种或多种添加的氧化还原酶是过氧化氢酶。在另一方面,该一种或多种添加的氧化还原酶是漆酶。在另一方面,该一种或多种添加的氧化还原酶是过氧化物酶。在另一方面,该一种或多种添加的氧化还原酶是超氧化物歧化酶。在另一方面,该一种或多种添加的氧化还原酶是选自下组的两种或更多种氧化还原酶的组合,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶。过氧化氢酶可以是有用于本发明的方法中的任何过氧化氢酶。该过氧化氢酶可以包括但不限于e.c.1.11.1.6或e.c.1.11.1.21过氧化氢酶。有用的过氧化氢酶的实例包括但不限于来自以下的过氧化氢酶:海水产碱杆菌(alcaligenesaquamarinus)(wo98/00526)、兰图鲁斯曲霉(aspergilluslentilus)、烟曲霉(paris等人,2003,infectimmun.[感染与免疫]71(6):3551-3562)、黑曲霉(美国专利号5,360,901)、米曲霉(jp2002223772a;美国专利号6,022,721)、嗜热葡糖苷酶芽孢杆菌(jp11243961a)、特异腐质霉(wo2009/104622,wo2012/130120)、樟绒枝霉(us2014/0335572)、变黑微颤菌(wo98/00526)、粗糙脉孢菌(dominguez等人,2010,arch.biochem.biophys.[生物化学和生物物理学档案]500:82-91)、埃默森青霉菌(wo2012/130120)、嗜松青霉(ep2256192)、微小根毛霉(us2014/0335572)、巴斯德酵母(wo2007/105350)、嗜热柱顶孢霉(sutaykocabas等人,2009,actacrystallogr.sect.f[结晶学部分学报]65:486-488)、柄篮状菌(wo2012/130120)、橙色嗜热子囊菌(wo2012/130120)、布氏栖热菌(thermusbrockianus)(wo2005/044994)、和土生梭孢壳霉(wo2010/074972)。在本发明中有用的过氧化氢酶的非限制性实例是来自以下的过氧化氢酶:专性嗜碱芽孢杆菌(bacilluspseudofirmus)(uniprot:p30266)、枯草芽孢杆菌(uniprot:p42234)、灰腐质霉(geneseqp:axq55105)、费希新萨托菌(uniprot:a1dju9)、粗糙脉孢菌(uniprot:q9c168)、埃默森青霉(geneseqp:bac10987)、嗜松青霉(geneseqp:bac10995)、嗜热柱顶孢霉(geneseqp:aaw06109或geneseqp:adt89624)、柄状踝节菌(geneseqp:bac10983或geneseqp:bac11039;uniprot:b8mt74)以及橙色嗜热子囊菌(geneseqp:bac11005;seqidno:8)。将登录号以其全部内容结合在此。该漆酶可以是在本发明的方法中有用的任何漆酶。该漆酶可以包括但不限于e.c.1.10.3.2漆酶。有用的漆酶的实例包括但不限于来自以下的漆酶:灰盖鬼伞(coprinuscinereus)(wo97/008325;schneider等人,1999;enzymeandmicrobialtechnology[酶和微生物技术]25:502-508)、嗜热棒囊孢菌壳(wo2013/087027)、白毛黑果菌(melanocarpusalbomyces)(kiiskinen等人,2004,microbiology[微生物学]150:3065-3074)、嗜热毁丝酶(wo95/033836、wo2006/012902)、polyporuspinsitus(wo96/000290、wo2014/028833)、变色多孔菌(polyporusversicolor)(等人,1998,appl.microbiol.biotechnol.[应用微生物学和生物技术]49:691-697)、朱红密孔菌(pycnoporuscinnabarinus)、稻瘟霉(pyriculariaoryzae)(muralikrishna等人,1995,appl.environ.microbiol.[应用和环境微生物学]61(12):4374-4377)、立枯丝核菌(rhizoctoniasolani)(wo95/007988;wo97/009431;waleithner等人,1996,curr.genet.[当前遗传学]29:395-403)、漆树漆酶(rhusvernicifera)(yoshida,1983,chemistryoflacquer(urushi)part1[漆化学(urushi)部分1]j.chem.soc.[化学学报]43:472-486)、嗜热色串孢(scytalidiumthermophilum)(wo95/033837、wo97/019999)、天蓝色链霉菌(streptomycescoelicolor)(machczynski等人,2004,inproteinscience[蛋白质科学]13:2388-2397)、和云芝(trametesversicolor)(wo96/000290)。在本发明中有用的漆酶的非限制性实例是来自以下的漆酶:灰盖鬼伞(coprinuscinereus)(geneseqp:aaw17974,geneseqp:aaw17975)、嗜热棒囊孢菌壳(geneseqp:bap78725)、嗜热毁丝酶(geneseqp:aar88500,geneseqp:aef76888)、polyporuspinsitus(geneseqp:bbd26012,geneseqp:aar90721)、立枯丝核菌(rhizoctoniasolani)(geneseqp:aar72328,geneseqp:aaw16301)、嗜热色串孢(scytalidiumthermophilum)(geneseqp:aar88500,geneseqp:aaw19855)和云芝(trametesversicolor)(geneseqp:aar90722)。将登录号以其全部内容结合在此。该过氧化物酶可以是在本发明的方法中有用的任何过氧化物酶。该过氧化物酶可以包括但不限于e.c.1.11.1.x过氧化物酶,例如e.c.1.11.1.1nadh过氧化物酶、e.c.1.11.1.2nadph过氧化物酶、e.c.1.11.1.3脂肪酸过氧化物酶、e.c.1.11.1.5二血红素细胞色素c过氧化物酶、e.c.1.11.1.5细胞色素c过氧化物酶、e.c.1.11.1.6过氧化氢酶、e.c.1.11.1.6锰过氧化氢酶、e.c.1.11.1.7无脊椎动物的细胞黏附蛋白、e.c.1.11.1.7嗜曙红细胞过氧化物酶、e.c.1.11.1.7乳过氧化物酶、e.c.1.11.1.7绿过氧物酶、e.c.1.11.1.8甲状腺过氧化物酶、e.c.1.11.1.9谷胱甘肽过氧化物酶、e.c.1.11.1.10氯化物过氧化物酶、e.c.1.11.1.11抗坏血酸过氧化物酶、e.c.1.11.1.12其他谷胱甘肽过氧化物酶、e.c.1.11.1.13锰过氧化物酶、e.c.1.11.1.14木质素过氧化物酶、e.c.1.11.1.15半胱氨酸过氧化物氧还蛋白、e.c.1.11.1.16通用过氧化物酶、e.c.1.11.1.17谷胱甘肽酰胺依赖性过氧化物酶、e.c.1.11.1.18溴过氧化物酶、e.c.1.11.1.19染料脱色过氧化物酶、e.c.1.11.1.b2氯过氧化物酶、e.c.1.11.1.b4卤素过氧化物酶、e.c.1.11.1.b4无血红素钒卤素过氧化物酶、e.c.1.11.1.b6碘过氧化物酶、e.c.1.11.1.b7溴过氧化物酶、和e.c.1.11.1.b8碘化物过氧化物酶。有用的过氧化物酶的实例包括但不限于灰盖鬼伞过氧化物酶(baunsgaard等人,1993,eur.j.biochem.[欧洲生物化学杂志]213(1):605-611;wo92/016634);辣根过氧化物酶(fujiyama等人,1988,eur.j.biochem.[欧洲生物化学杂志]173(3):681-687);过氧化物氧还蛋白(singh和shichi,1998,j.biol.chem.[生物化学杂志]273(40):26171-26178);乳过氧化物酶(dull等人,1990,dnacellbiol.[dna细胞生物学]9(7):499-509);嗜酸性粒细胞过氧化物酶(fornhem等人,1996,int.arch.allergyimmunol.[过敏与免疫学国际档案]110(2):132-142);通用过氧化物酶(ruiz-duenas等人,1999,mol.microbiol.[分子微生物学]31(1):223-235);芜菁过氧化物酶(mazza和welinder,1980,eur.j.biochem.[欧洲生物化学杂志]108(2):481-489);髓过氧化物酶(morishita等人,1987,j.biol.chem.[生物化学杂志]262:15208-15213);过氧蛋白(peroxidasin)和过氧蛋白同系物(horikoshi等人,1999,biochem.biophys.res.commun.[生物化学与生物物理研究通讯]261(3):864-869);木质素过氧化物酶(tien和tu,1987,nature[自然]326(6112):520-523);和锰过氧化物酶(orth等人,1994,gene[基因]148(1):161-165)。有用于本发明的过氧化物酶的非限制性实例是来自以下的过氧化物酶:灰盖鬼伞(uniprot:p28314)、家牛(bostaurus)(uniprot:o77834、uniprot:p80025)芜菁亚种rapa(uniprot:p00434)、智人(uniprot:p05164、uniprot:q92616)、辣根过氧化物酶(uniprot:p15232)、杏鲍菇(uniprot:o94753)、黄孢原毛平革菌(uniprot:p06181、uniprot:p78733)和野猪(susscrofa)(uniprot:p80550)。将登录号以其全部内容结合在此。该超氧化物歧化酶可以是本发明的方法中有用的任何超氧化物歧化酶。超氧化物歧化酶可以包括但不限于e.c.1.15.1.1超氧化物歧化酶。有用的超氧化物歧化酶的实例包括但不限于来自以下的超氧化物歧化酶:黄曲霉(holdom等人,1996,infect.immun.[感染与免疫]64:3326-3332)、构巢曲霉(holdom等人,1996,infect.immun.[感染与免疫]64:3326-3332)、黑曲霉(dolashki等人,2008,spectrochim.actaa.mol.biomol.spectrosc.[光谱化学学报a分子和生物分子光谱学]71,975-983)、土曲霉(holdom等人,1996,infect.immun.[感染与免疫]64:3326-3332)、蜡样芽孢杆菌(wang等人,2007,femsmicrobiol.lett.[fems微生物快报]272:206-213)、嗜热毛壳菌(zhang等人,2011,biotechnol.lett.[生物技术快报]33:1127-1132)、马克斯克鲁维酵母(nedeva等人,2009,chromatogr.b[色谱b杂志]877:3529-3536)、嗜热毁丝酶(wo2012/068236)rasamsoniaemersonii(wo2014/002616)、酿酒酵母(borders等人,1998,biochemistry[生物化学]37,11323-11331)、马尔尼菲篮状菌(talaromycesmarneffei)(thirach等人,2007,med.mycol.[医学真菌学]45:409-417)、橙色嗜热子囊菌(shijin等人,2007,biosci.biotechnol.biochem.[生物科学、生物技术、以及生物化学]71:1090-1093;song等人,2009,j.microbiol.[微生物学杂志]47:123-130)和土生梭孢霉(berka等人,2011,nat.biotechnol.[自然生物技术]29:922-927)。有用于本发明的超氧化物歧化酶的非限制性实例是来自以下的超氧化物歧化酶:蜡样芽孢杆菌(uniprot:q6qht3)、嗜热毛壳菌(uniprot:q1heq0)、马克斯克鲁维酵母(uniprot:bob552)、嗜热毁丝酶(geneseqp:azw56690)、rasamsoniaemersonii(geneseqp:bbt31699)、马尔尼菲篮状菌(talaromycesmarneffei)(uniprot:b6qeb3)、橙色嗜热子囊菌(uniprot:q1hdv5,uniprot:q1hdv5)、和土生梭孢霉(uniprot:g2r3v2)。将登录号以其全部内容结合在此。在一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)与在此披露的氧化还原酶的成熟多肽具有至少60%,例如至少65%、至少70%、至少75%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、或100%的序列一致性,该成熟多肽具有氧化还原酶活性。在另一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)的氨基酸序列与在此披露的氧化还原酶的成熟多肽相差多达10个(例如1个、2个、3个、4个、5个、6个、7个、8个、9个、或10个)氨基酸。在另一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)包含在此披露的氧化还原酶的氨基酸序列,或由其组成。在另一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)包含在此披露的氧化还原酶的成熟多肽,或由其组成。在另一个实施例中,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)是在此披露的氧化还原酶的等位基因变体。在另一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)是含有在此披露的氧化还原酶的成熟多肽的至少85%氨基酸残基(例如至少90%氨基酸残基、或至少95%氨基酸残基)的片段。在另一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)由以下多核苷酸编码,该多核苷酸在非常低、低、中、中-高、高或非常高严格条件下与在此披露的氧化还原酶的成熟多肽编码序列或其全长补体杂交(sambrook等人,1989,见上文)。可以使用编码氧化还原酶的多核苷酸、或其子序列,以及氧化还原酶的多肽、或其片段来涉及核酸探针以根据本领域熟知的方法来鉴定并克隆编码来自不同属或种的菌株的氧化还原酶的dna。具体而言,此类探针可以用于与感兴趣的细胞的基因组dna或cdna杂交,如在上文所述。出于本发明的目的,杂交是指在非常低至非常高严格条件下多核苷酸杂交到标记的核酸探针上。在这些条件下与该核酸探针杂交的分子可以使用例如x-射线胶片或本领域中已知的任何其他检测手段进行检测。在一方面,该核酸探针是氧化还原酶的成熟多肽编码序列。在另一方面,该核酸探针是编码全长氧化还原酶的多核苷酸;其成熟多肽;或其片段。在另一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)由多核苷酸编码,该多核苷酸与在此披露的氧化还原酶的成熟多肽编码序列具有至少60%,例如至少65%、至少70%、至少75%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、或100%的序列一致性。该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)可以是杂合多肽,其中一种多肽的一个区域融合在另一种多肽、或融合多肽或可裂解的融合多肽的区域的n-末端或c-末端,其中另一种多肽融合在氧化还原酶的n-末端或c-末端,如在此所述。在酶组合物中,添加的氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)的蛋白质含量处于约0.1%至约10%,例如,约0.1%至约7%、约0.1%至约5%、约0.1%至约4%、约0.1%至约3%、约0.1%至约2%以及约0.1%至约1%的总酶蛋白的范围内。在实施例中,添加的氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、或超氧化物歧化酶)与aa9溶解性多糖单加氧酶的蛋白质比率处于约1:250至约1:10,例如约1:200至约1:10、约1:150至约1:15、约1:100至约1:15、约1:75至约1:20、或约1:50至约1:25的范围内。宿主细胞在本发明的方法中,该宿主细胞可以是野生型宿主细胞或重组宿主细胞。术语“宿主细胞”涵盖由于复制期间发生的突变而与亲本细胞不相同亲本细胞的任何后代。该宿主细胞可以是在酶组合物的产生中有用的任何细胞。在一方面,该宿主细胞是原核细胞。在另一方面,该宿主细胞是真核细胞。原核宿主细胞可为任何革兰氏阳性或革兰氏阴性细菌。革兰氏阳性细菌包括但不限于:芽孢杆菌属、梭菌属、肠球菌属、土芽孢杆菌属、乳杆菌属、乳球菌属、海洋芽孢杆菌属、葡萄球菌属、链球菌属、和链霉菌属。革兰氏阴性细菌包括但不限于:弯曲杆菌属、大肠杆菌、黄杆菌属、梭杆菌属、螺杆菌属、泥杆菌属、奈瑟氏菌属、假单胞菌属、沙门氏菌属、以及脲原体属。细菌宿主细胞可以是任何芽孢杆菌属细胞,包括但不限于:嗜碱芽孢杆菌、解淀粉芽孢杆菌、短芽孢杆菌、环状芽孢杆菌、克劳氏芽孢杆菌、凝结芽孢杆菌、坚强芽孢杆菌、灿烂芽孢杆菌、迟缓芽孢杆菌、地衣芽孢杆菌、巨大芽孢杆菌、短小芽孢杆菌、嗜热脂肪芽孢杆菌、枯草芽孢杆菌以及苏云金芽孢杆菌细胞。细菌宿主细胞还可以是任何链霉菌属细胞,包括但不限于:不产色链霉菌、除虫链霉菌、天蓝链霉菌、灰色链霉菌、以及浅青紫链霉菌细胞。将dna引入芽孢杆菌属细胞中可通过以下来实现:原生质体转化(参见,例如,chang和cohen,1979,mol.gen.genet.[分子遗传学与基因组学]168:111-115)、感受态细胞转化(参见,例如,young和spizizen,1961,j.bacteriol.[细菌学杂志]81:823-829;或dubnauh和davidoff-abelson,1971,j.mol.biol.[分子生物学杂志]56:209-221)、电穿孔(参见,例如,shigekawa和dower,1988,biotechniques[生物技术]6:742-751)、或者接合(参见,例如,koehler和thorne,1987,j.bacteriol.[细菌学杂志]169:5271-5278)。将dna引入大肠杆菌细胞中可通过以下来实现:原生质体转化(参见,例如,hanahan,1983,j.mol.biol.[分子生物学杂志]166:557-580)或电穿孔(参见,例如dower等人,1988,nucleicacidsres.[核酸研究]16:6127-6145)。将dna引入链霉菌属细胞中可通过以下来实现:原生质体转化、电穿孔(参见,例如,gong等人,2004,foliamicrobiol.[叶线形微生物学](布拉格(praha))49:399-405)、接合(参见例如,mazodier等人,1989,j.bacteriol.[细菌学杂志]171:3583-3585)、或转导(参见例如,burke等人,2001,proc.natl.acad.sci.usa[美国国家科学院院刊]98:6289-6294)。将dna引入假单孢菌属细胞中可通过以下来实现:电穿孔(参见,例如,choi等人,2006,j.microbiol.methods[微生物学方法杂志]64:391-397)或接合(参见,例如,pinedo和smets,2005,appl.environ.microbiol.[应用与环境微生物学]71:51-57)。将dna引入链球菌属细胞中可通过以下来实现:天然感受态(参见例如,perry和kuramitsu,1981,infect.immun.[感染与免疫]32:1295-1297)、原生质体转化(参见,例如,catt和jollick,1991,microbios[微生物学]68:189-207)、电穿孔(参见,例如,buckley等人,1999,appl.environ.microbiol.[应用与环境微生物学]65:3800-3804)、或者接合(参见,例如,clewell,1981,microbiol.rev.[微生物学评论]45:409-436)。然而,可使用本领域已知的将dna引入宿主细胞的任何方法。宿主细胞还可为真核生物,如哺乳动物、昆虫、植物或真菌细胞。宿主细胞可为真菌细胞。如在此使用的“真菌”包括子囊菌门(ascomycota)、担子菌门(basidiomycota)、壶菌门(chytridiomycota)和接合菌门(zygomycota)以及卵菌门(oomycota)和所有有丝分裂孢子真菌(如由hawksworth等人所定义的,在:ainsworthandbisby’sdictionaryofthefungi[ainsworth和bisby的真菌大词典],第8版,1995,国际cab,大学出版社,剑桥,英国)。真菌宿主细胞可以是酵母细胞。如在此所用的“酵母”包括产子囊酵母(ascosporogenousyeast)(内孢霉目(endomycetales))、产担子酵母(basidiosporogenousyeast)和属于半知菌类(fungiimperfecti)(芽孢纲(blastomycetes))的酵母。由于酵母的分类可能在将来变化,出于本发明的目的,酵母应当如酵母的生物学与活性(skinner、passmore和davenport编辑,soc.app.bacteriol.symposiumseriesno.9[应用细菌学学会专题论文集系列9],1980)所描述那样定义。酵母宿主细胞可以是假丝酵母属细胞、汉逊酵母属细胞、克鲁维酵母属细胞、毕赤酵母属细胞、酵母菌属细胞、裂殖酵母或耶罗维亚酵母属细胞、如乳酸克鲁维酵母细胞、卡氏酵母细胞、酿酒酵母细胞、糖化酵母细胞、道格拉斯酵母(saccharomycesdouglasii)细胞、克鲁弗酵母细胞、诺地酵母细胞、卵形酵母细胞或解脂耶罗维亚酵母细胞。真菌宿主细胞可为丝状真菌细胞。“丝状真菌”包括真菌门(eumycota)和卵菌门(oomycota)的亚门的所有丝状形式(如由hawksworth等人,1995,见上文)。丝状真菌通常的特征在于由几丁质、纤维素、葡聚糖、壳多糖、甘露聚糖、以及其他复杂多糖构成的菌丝体壁。营养生长是通过菌丝延长,而碳分解代谢是专性需氧的。相反,酵母(如酿酒酵母)的营养生长是通过单细胞菌体的出芽(budding),且碳分解代谢可以是发酵性的。丝状真菌宿主细胞可以是枝顶孢属、曲霉属、短梗霉属、烟管霉属(bjerkandera)、拟腊菌属、金孢子菌属、鬼伞属、革盖菌属(coriolus)、隐球菌属、线黑粉菌科(filibasidium)、镰孢属、腐质霉属、梨孢菌属、毛霉属、毁丝霉属、新美鞭菌属、链孢菌属、拟青霉属、青霉属、平革菌属、射脉菌属(phlebia)、瘤胃壶菌属、侧耳属(pleurotus)、裂褶菌属、篮状菌属、嗜热子囊菌属、梭孢壳属、弯颈霉属、栓菌属(trametes)或木霉属细胞。例如,丝状真菌宿主细胞可以是泡盛曲霉、臭曲霉、烟曲霉、日本曲霉、构巢曲霉、黑曲霉、米曲霉、黑刺烟管菌(bjerkanderaadusta)、干拟蜡菌(ceriporiopsisaneirina)、卡内基拟蜡菌(ceriporiopsiscaregiea)、浅黄拟蜡孔菌(ceriporiopsisgilvescens)、潘诺希塔拟蜡菌(ceriporiopsispannocinta)、环带拟蜡菌(ceriporiopsisrivulosa)、微红拟蜡菌(ceriporiopsissubrufa)、虫拟蜡菌(ceriporiopsissubvermispora)、狭边金孢子菌(chrysosporiuminops)、嗜角质金孢子菌、卢克诺文思金孢子菌(chrysosporiumlucknowense)、粪状金孢子菌(chrysosporiummerdarium)、租金孢子菌、女王杜香金孢子菌(chrysosporiumqueenslandicum)、热带金孢子菌、褐薄金孢子菌(chrysosporiumzonatum)、灰盖鬼伞(coprinuscinereus)、毛革盖菌(coriolushirsutus)、杆孢状镰孢、谷类镰孢、库威镰孢、大刀镰孢、禾谷镰孢、禾赤镰孢、异孢镰孢、合欢木镰孢、尖镰孢、多枝镰孢、粉红镰孢、接骨木镰孢、肤色镰孢、拟分枝孢镰孢、硫色镰孢、圆镰孢、拟丝孢镰孢、镶片镰孢、特异腐质霉、柔毛腐质霉、米黑毛霉、嗜热毁丝霉、粗糙链孢菌、产紫青霉、黄孢平革菌(phanerochaetechrysosporium)、射脉菌(phlebiaradiata)、刺芹侧耳(pleurotuseryngii)、埃默森篮状菌、土生梭孢霉、长域毛栓菌(trametesvillosa)、变色栓菌(trametesversicolor)、哈茨木霉、康宁木霉、长枝木霉、里氏木霉或绿色木霉细胞。真菌细胞能通过涉及原生质体形成、原生质体的转化和细胞壁的再生的过程以本身已知的方式进行转化。用于转化曲霉属和木霉属宿主细胞的适合程序描述于以下文献中:ep238023,yelton等人,1984,proc.natl.acad.sci.usa[美国国家科学院院刊]81:1470-1474以及christensen等人,1988,bio/technology[生物/技术]6:1419-1422。用于转化镰孢属菌种的合适方法在malardier等,1989,gene78:147-156和wo96/00787中描述。可以使用由如以下文献描述的程序转化酵母:becker和guarente,在abelson,j.n.和simon,m.i.编,guidetoyeastgeneticsandmolecularbiology[酵母遗传学与分子生物学指南],methodsinenzymology[酶学方法],第194卷,第182-187页,学术出版社有限公司(academicpress,inc.),纽约;ito等人,1983,j.bacteriol.[细菌学杂志]153:163;以及hinnen等人,1978,proc.natl.acad.sci.usa[美国国家科学院院刊]75:1920。酶组合物该酶组合物可以包含选自下组的一种或多种(例如,若干种)酶,该组由以下组成:水解酶、异构酶、连接酶、裂解酶、氧化还原酶、或转移酶。在一方面,该酶组合物可以包含选自下组的一种或多种(例如,若干种)酶,该组由以下组成:α-半乳糖苷酶、α-葡糖苷酶、氨肽酶、淀粉酶、β-半乳糖苷酶、β-葡糖苷酶、β-木糖苷酶、糖酶、羧肽酶、过氧化氢酶、纤维二糖水解酶、纤维素酶、壳多糖酶、角质酶、环糊精糖基转移酶、脱氧核糖核酸酶、内切葡聚糖酶、酯酶、葡糖淀粉酶、转化酶、漆酶、脂肪酶、甘露糖苷酶、变位酶、氧化酶、果胶分解酶、过氧化物酶、植酸酶、多酚氧化酶、蛋白水解酶、核糖核酸酶、转谷氨酰胺酶、和木聚糖酶。在另一方面,该酶组合物可以包含在降解木质纤维素材料(例如纤维素或半纤维素材料)中有用的任何蛋白质。在另一方面,该酶组合物包含或进一步包含选自下组的一种或多种(例如,若干种)蛋白质,该组由以下组成:纤维素酶、aa9多肽、半纤维素酶、纤维素诱导蛋白(cip)、酯酶、棒曲霉素、木质素分解酶、果胶酶、蛋白酶以及膨胀素。在另一方面,纤维素酶优选是选自下组的一种或多种(例如,若干种)酶,该组由以下组成:内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶。在另一方面,半纤维素酶优选是选自下组的一种或多种(例如,若干种)酶,该组由以下组成:乙酰甘露聚糖酯酶、乙酰木聚糖酯酶、阿拉伯聚糖酶、阿拉伯呋喃糖苷酶、香豆酸酯酶、阿魏酸酯酶、半乳糖苷酶、葡糖醛酸糖苷酶、葡萄糖醛酸酯酶、甘露聚糖酶、甘露糖苷酶、木聚糖酶、以及木糖苷酶。在另一方面,该酶组合物包含一种或多种(例如,若干种)纤维素分解酶。在另一方面,该酶组合物包含或进一步包含一种或多种(例如,若干种)半纤维素分解酶。在另一方面中,该酶组合物包含一种或多种(例如,若干种)纤维素分解酶和一种或多种(例如,若干种)半纤维素分解酶。在另一方面中,该酶组合物包含选自纤维素分解酶和半纤维素分解酶的组的一种或多种(例如,若干种)酶。在另一方面中,该酶组合物包含内切葡聚糖酶。在另一方面,该酶组合物包含纤维二糖水解酶。在另一方面,该酶组合物包含β-葡糖苷酶。在另一方面,该酶组合物包含aa9多肽。在另一方面,该酶组合物包含内切葡聚糖酶和aa9多肽。在另一方面,该酶组合物包含纤维二糖水解酶和aa9多肽。在另一方面,该酶组合物包含β-葡糖苷酶和aa9多肽。在另一方面,该酶组合物包含内切葡聚糖酶和纤维二糖水解酶。在另一方面,该酶组合物包含内切葡聚糖酶i、内切葡聚糖酶ii、或内切葡聚糖酶i和内切葡聚糖酶ii的组合、以及纤维二糖水解酶i、纤维二糖水解酶ii、或纤维二糖水解酶i和纤维二糖水解酶ii的组合。在另一方面,该酶组合物包含内切葡聚糖酶和β-葡糖苷酶。在另一方面,该酶组合物包含内切葡聚糖酶i、内切葡聚糖酶ii、或内切葡聚糖酶i和内切葡聚糖酶ii的组合、以及β-葡糖苷酶。在另一方面,该酶组合物包含β-葡糖苷酶和纤维二糖水解酶。在另一方面,该酶组合物包含β-葡糖苷酶和纤维二糖水解酶i、纤维二糖水解酶ii、或纤维二糖水解酶i和纤维二糖水解酶ii的组合。在另一方面,该酶组合物包含内切葡聚糖酶、aa9多肽、以及纤维二糖水解酶。在另一方面,该酶组合物包含内切葡聚糖酶i、内切葡聚糖酶ii、或内切葡聚糖酶i和内切葡聚糖酶ii的组合、aa9多肽、以及纤维二糖水解酶i、纤维二糖水解酶ii、或纤维二糖水解酶i和纤维二糖水解酶ii的组合。在另一方面,该酶组合物包含内切葡聚糖酶、β-葡糖苷酶、和aa9多肽。在另一方面,该酶组合物包含β-葡糖苷酶、aa9多肽、和纤维二糖水解酶。在另一方面,该酶组合物包含β-葡糖苷酶、aa9多肽、以及纤维二糖水解酶i、纤维二糖水解酶ii、或纤维二糖水解酶i和纤维二糖水解酶ii的组合。在另一方面,该酶组合物包含内切葡聚糖酶、β-葡糖苷酶和纤维二糖水解酶。在另一方面,该酶组合物包含内切葡聚糖酶i、内切葡聚糖酶ii、或内切葡聚糖酶i和内切葡聚糖酶ii的组合、β-葡糖苷酶、以及纤维二糖水解酶i、纤维二糖水解酶ii、或纤维二糖水解酶i和纤维二糖水解酶ii的组合。在另一方面,该酶组合物包含内切葡聚糖酶、纤维二糖水解酶、β-葡糖苷酶、和aa9多肽。在另一方面,该酶组合物包含内切葡聚糖酶i、内切葡聚糖酶ii、或内切葡聚糖酶i和内切葡聚糖酶ii的组合、β-葡糖苷酶、aa9多肽、以及纤维二糖水解酶i、纤维二糖水解酶ii、或纤维二糖水解酶i和纤维二糖水解酶ii的组合。在另一方面,该酶组合物包含乙酰甘露聚糖酯酶。在另一方面,该酶组合物包含乙酰木聚糖酯酶。在另一方面,该酶组合物包含阿拉伯聚糖酶(例如,α-l-阿拉伯聚糖酶)。在另一方面,该酶组合物包含阿拉伯呋喃糖苷酶(例如,α-l-阿拉伯呋喃糖苷酶)。在另一方面,该酶组合物包含香豆酸酯酶。在另一方面,该酶组合物包含阿魏酸酯酶。在另一方面,该酶组合物包含半乳糖苷酶(例如,α-半乳糖苷酶和/或β-半乳糖苷酶)。在另一方面,该酶组合物包含葡糖醛酸糖苷酶(例如,α-d-葡糖醛酸糖苷酶)。在另一方面,该酶组合物包含葡糖醛酸酯酶。在另一方面,该酶组合物包含甘露聚糖酶。在另一方面,该酶组合物包含甘露糖苷酶(例如,β-甘露糖苷酶)。在另一方面,该酶组合物包含木聚糖酶。在一个实施例中,木聚糖酶是家族10的木聚糖酶。在另一个实施例中,木聚糖酶是家族11的木聚糖酶。在另一方面,该酶组合物包含木糖苷酶(例如β-木糖苷酶)。在另一方面,该酶组合物包含酯酶。在另一方面,该酶组合物包含棒曲霉素。在另一方面,该酶组合物包含木质素分解酶。在一个实施例中,木质素分解酶是锰过氧化物酶。在另一个实施例中,木质素分解酶是木质素过氧化物酶。在另一个实施例中,木质素分解酶是h2o2产生酶。在另一方面,该酶组合物包含果胶酶。在另一方面,该酶组合物包含氧化还原酶。在另一方面,该酶组合物包含蛋白酶。在另一方面,该酶组合物包含膨胀素。该酶组合物的一种或多种(例如,若干种)组分可以是天然蛋白、重组蛋白或天然蛋白与重组蛋白的组合。例如,一种或多种(例如,若干种)组分可以是用作宿主细胞以重组表达该酶组合物的一种或多种(例如,若干种)其他组分的细胞的天然蛋白。在此应理解的是,重组蛋白对于宿主细胞可以是异源的(例如,外源的)和/或原生的。可以作为单组分生成酶组合物的一种或多种(例如,若干种)组分,然后将它们组合以形成酶组合物。酶组合物可以是多组分和单组分蛋白制剂的组合。具有纤维素分解酶活性或半纤维素分解酶活性的多肽以及有用于降解纤维素材料或半纤维素材料的其他蛋白/多肽(例如,aa9多肽)可以从任何合适的来源衍生或获得,包括古细菌、细菌、真菌、酵母、植物或哺乳动物来源。术语“获得的”在此还意指该酶可能已在宿主生物中采用在此所描的方法重组产生,其中重组产生的酶对于宿主生物是原生的或外源的,或具有修饰的氨基酸序列,例如,具有一个或多个(例如,若干个)缺失、插入和/或取代的氨基酸,即重组产生的酶为天然氨基酸序列的突变体和/或片段,或通过本领域已知的核酸改组方法产生的酶。原生酶的含义中涵盖天然变体,而外源酶的含义中涵盖如通过定点诱变或改组获得的变体。每种多肽都可以是细菌多肽。例如,每种多肽可以是具有酶活性的革兰氏阳性细菌多肽,或具有酶活性的革兰氏阴性细菌多肽。每种多肽还可以是真菌多肽(例如,酵母菌多肽或丝状真菌多肽)。还可以使用多肽的化学修饰的或蛋白工程化的突变体。该酶组合物的一种或多种(例如,若干种)组分可以是重组组分,即,通过克隆编码该单一组分的dna序列并且随后用该dna序列转化细胞并且在宿主中表达产生(参见,例如,wo91/17243和wo91/17244)。该宿主可以是异源宿主(酶对于宿主来说是外源的),但该宿主在某些条件下也可以是同源宿主(酶对于宿主来说是原生的)。还可以通过纯化来自发酵液的这样一种蛋白来制备单组分纤维素分解蛋白。在一方面,该一种或多种(例如,若干种)纤维素分解酶包括商业纤维素分解酶制剂。适用于本发明的商业纤维素分解酶制剂的实例包括例如:ctec(诺维信公司)、ctec2(诺维信公司)、ctec3(诺维信公司)、celluclasttm(诺维信公司)、novozymtm188(诺维信公司)、spezymetmcp(杰能科国际(genencorint.))、accellerasetmtrio(杜邦公司(dupont))、nl(dsm公司);s/l100(dsm公司)、rohamenttm7069w(罗姆公司(gmbh))、或cmax3tm(dyadic国际有限公司(dyadicinternational,inc.))。以从约0.001wt.%至约5.0wt.%的固体,例如0.025wt.%至约4.0wt.%的固体、或约0.005wt.%至约2.0wt.%的固体的有效量添加纤维素分解酶制剂。细菌内切葡聚糖酶的实例包括但不限于:解纤维热酸菌(acidothermuscellulolyticus)内切葡聚糖酶(wo91/05039;wo93/15186;美国专利申请号5,275,944;wo96/02551;美国专利申请号5,536,655,wo00/70031,wo05/093050)、胡萝卜软腐欧文氏菌(erwiniacarotovara)内切葡聚糖酶(saarilahti等人,1990,gene[基因]90:9-14)、嗜热裂孢菌(thermobifidafusca)内切葡聚糖酶iii(wo05/093050)、以及嗜热裂孢菌(thermobifidafusca)内切葡聚糖酶v(wo05/093050)。可以用于本发明的真菌内切葡聚糖酶的实例包括但不限于:里氏木霉内切葡聚糖酶i(penttila等人,1986,基因45:253-263,里氏木霉cel7b内切葡聚糖酶i(genbank:m15665)、里氏木霉内切葡聚糖酶ii(saloheimo等人,1988,gene[基因]63:11-22),里氏木霉cel5a内切葡聚糖酶ii(genbank:m19373)、里氏木霉内切葡聚糖酶iii(okada等人,1988,appl.environ.microbiol.[应用与环境微生物学]64:555-563,genbank:ab003694)、里氏木霉内切葡聚糖酶v(saloheimo等人,1994,molecularmicrobiology[分子微生物学]13:219-228,genbank:z33381)、棘孢曲霉内切葡聚糖酶(ooi等人,1990,nucleicacidsresearch[核酸研究]18:5884)、白曲霉内切葡聚糖酶(sakamoto等人,1995,currentgenetics[当代遗传学]27:435-439)、尖镰孢内切葡聚糖酶(genbank:l29381)、灰腐质霉高温变种(humicolagriseavar.thermoidea)内切葡聚糖酶(genbank:ab003107)、热白丝菌(melanocarpusalbomyces)内切葡聚糖酶(genbank:mal515703)、粗糙脉孢菌内切葡聚糖酶(genbank:xm_324477)、特异腐质霉内切葡聚糖酶v、嗜热毁丝霉cbs117.65内切葡聚糖酶、金黄色嗜热子囊菌内切葡聚糖酶i(genbank:af487830)、里氏木霉菌株号vtt-d-80133内切葡聚糖酶(genbank:m15665)、以及嗜松青霉内切葡聚糖酶(wo2012/062220)。可用在本发明中的纤维二糖水解酶的实例包括但不限于:棘孢曲霉纤维二糖水解酶ii(wo2011/059740)、烟曲霉纤维二糖水解酶i(wo2013/028928)、烟曲霉纤维二糖水解酶ii(wo2013/028928)、嗜热毛壳菌纤维二糖水解酶i、嗜热毛壳菌纤维二糖水解酶ii、特异腐质霉纤维二糖水解酶i、嗜热毁丝霉纤维二糖水解酶ii(wo2009/042871)、奥斯塔尼青霉(penicilliumoccitanis)纤维二糖水解酶i(genbank:ay690482)、埃默森篮状菌纤维二糖水解酶i(genbank:af439936)、赫卡尼亚梭孢壳霉(thielaviahyrcanie)纤维二糖水解酶ii(wo2010/141325)、土生梭孢壳霉纤维二糖水解酶ii(cel6a,wo2006/074435)、里氏木霉纤维二糖水解酶i、里氏木霉纤维二糖水解酶ii、以及褐孢长毛盘菌纤维二糖水解酶ii(wo2010/057086)。适用于本发明的β-葡糖苷酶的实例包括但不限于来自以下的β-葡糖苷酶:棘孢曲霉(kawaguchi等人,1996,gene[基因]173:287-288)、烟曲霉(wo2005/047499)、黑曲霉(dan等人,2000,j.biol.chem.[生物化学杂志]275:4973-4980)、米曲霉(wo02/095014)、巴西青霉菌ibt20888(wo2007/019442和wo2010/088387)、土生梭孢壳霉(wo2011/035029)、以及褐孢长毛盘菌(wo2007/019442)。其他有用的内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶在采用根据下述文献的分类的很多糖基水解酶家族中披露:henrissat,1991,biochem.j.[生物化学杂志]280:309-316,以及henrissat和bairoch,1996,生物化学杂志316:695-696。在一方面,该一种或多种(例如,若干种)半纤维素分解酶包含商业半纤维素分解酶制剂。适于在本发明中使用的商业化半纤维素分解酶制剂的实例包括例如shearzymetm(诺维信公司)、htec(诺维信公司)、htec2(诺维信公司)、htec3(诺维信公司)、(诺维信公司)、(诺维信公司)、hc(诺维信公司)、木聚糖酶(杰能科公司)、xy(杰能科公司)、xc(杰能科公司)、tx-200a(ab酶公司(abenzymes))、hsp6000木聚糖酶(dsm)、depoltm333p(生物催化剂有限公司(biocatalystslimit),威尔士,英国)、depoltm740l(生物催化剂有限公司,威尔士,英国)以及depoltm762p(生物催化剂有限公司,威尔士,英国)、alternafuel100p(dyadic公司)和alternafuel200p(dyadic公司)。木聚糖酶的实例包括但不限于来自以下的木聚糖酶:棘孢曲霉(geneseqp:aar63790;wo94/21785)、烟曲霉(wo2006/078256)、嗜松青霉(wo2011/041405)、青霉属物种(wo2010/126772)、疏棉状嗜热丝孢菌(geneseqp:baa22485)、嗜热篮状菌(geneseqp:baa22834)、土生梭孢霉nrrl8126(wo2009/079210)、以及褐孢长毛盘菌(wo2011/057083)。β-木糖苷酶的实例包括但不限于来自以下的β-木糖苷酶:粗糙脉孢菌(swissprot:q7sow4)、里氏木霉(uniprotkb/trembl:q92458)、埃默森篮状菌(swissprot:q8x212)、以及嗜热篮状菌(geneseqp:baa22816)。乙酰木聚糖酯酶的实例包括但不限于来自以下的乙酰木聚糖酯酶:棘孢曲霉(wo2010/108918),球毛壳菌(uniprot:q2gwx4),细丽毛壳菌(chaetomiumgracile)(geneseqp:aab82124),特异腐质霉dsm1800(wo2009/073709),红褐肉座菌(wo2005/001036),嗜热毁丝菌(myceliophterathermophila)(wo2010/014880),粗糙脉孢菌(uniprot:q7s259),颖枯壳针孢(phaeosphaerianodorum)(uniprot:q0uhj1),以及土生梭孢壳霉nrrl8126(wo2009/042846)。阿魏酸酯酶(feruloylesterase,ferulicacidesterase)的实例包括但不限于来自以下的阿魏酸酯酶:特异腐质霉dsm1800(wo2009/076122),费希新萨托菌(neosartoryafischeri)(uniprot:a1d9t4),粗糙脉孢菌(uniprot:q9hgr3),黄灰青霉(penicilliumaurantiogriseum)(wo2009/127729),以及土生梭孢壳霉(wo2010/053838和wo2010/065448)。阿拉伯呋喃糖苷酶的实例包括但不限于来自以下的阿拉伯呋喃糖苷酶:黑曲霉(geneseqp:aar94170)、特异腐质霉dsm1800(wo2006/114094和wo2009/073383)、以及大型亚灰树花菌(m.giganteus)(wo2006/114094)。α-葡糖醛酸糖苷酶的实例包括但不限于来自以下的α-葡糖醛酸糖苷酶:棒曲霉(uniprot:alcc12)、烟曲霉(swissprot:q4ww45)、黑曲霉(uniprot:q96wx9)、土曲霉(swissprot:q0cjp9)、特异腐质霉(wo2010/014706)、黄灰青霉(wo2009/068565)、埃默森篮状菌(uniprot:q8x211)、以及里氏木霉(uniprot:q99024)。在一方面,该氧化还原酶(例如过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶)抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的失活。在一方面,该酶组分是纤维素酶。在另一方面,该酶组分是半纤维素酶。在另一方面,该酶组分是纤维素诱导蛋白(cip)。在另一方面,该酶组分是酯酶。在另一方面,该酶组分是棒曲霉素。在另一方面,该酶组分是木质素分解酶。在另一方面,该酶组分是果胶酶。在另一方面,该酶组分是蛋白酶。在另一方面,该酶组分是膨胀素。在另一方面,该酶组分是纤维二糖水解酶。在另一方面,该酶组分是纤维二糖水解酶i。在另一方面,该酶组分是纤维二糖水解酶ii。在另一方面,该酶组分是内切葡聚糖酶。在另一方面,该酶组分是β-葡糖苷酶。在另一方面,该酶组分是木聚糖酶。在另一方面,该酶组分是β-木糖苷酶。组合物组分可以通过在含有适合碳源和氮源以及无机盐的营养培养基上,使用本领域中已知的程序发酵上述宿主细胞来产生(参见,例如,bennett,j.w.和lasure,l(编辑),moregenemanipulationsinfungi[真菌中的更多基因操纵],学术出版社(academicpress),加州,1991)。适合的培养基可从商业供应商获得或可以根据公开的组成(例如,在美国典型培养物保藏中心(americantypeculturecollection)的目录中)制备。适合于生长和酶产生的温度范围和其他条件在本领域中是已知的(参见,例如,贝利·j.e.(bailey,j.e.)和奥利斯·d.f.(ollis,d.f.),生物化学工程基础(biochemicalengineeringfundamentals),麦格劳-希尔图书公司(mcgraw-hillbookcompany),纽约,1986)。发酵可以是导致酶或蛋白质的表达或分离的培养细胞的任何方法。所以,可以将发酵理解为包含摇瓶培养,或者在一种适合的培养基中并且在允许表达或分离该酶的条件下在实验室或工业发酵罐中进行小规模或大规模发酵(包括连续发酵、分批发酵、分批补料发酵或固态发酵)。通过上述方法产生的所得酶可以从发酵培养基回收并且通过常规程序纯化。酶组合物可以处于任何适于使用的形式,例如像发酵液配制品或细胞组合物、具有或不具有细胞碎片的细胞裂解物、半纯化的或纯化的酶制剂、或作为酶的来源的宿主细胞。该酶组合物可为干粉或颗粒,无粉尘的颗粒,液体,稳定化液体或稳定化受保护的酶。可以根据已建立的方法例如通过添加稳定剂(如糖、糖醇或其他多元醇)、和/或乳酸或另一种有机酸,对液体酶制剂进行稳定化。酶组合物可以是包含本发明的多肽的发酵液配制品或细胞组合物。发酵液产物进一步包含在发酵过程中使用的另外的成分,例如像,细胞(包括含有编码本发明的多肽的基因的宿主细胞,这些宿主细胞被用于产生多肽)、细胞碎片、生物质、发酵培养基和/或发酵产物。在一些实施例中,该组合物是含有一种或多种有机酸、杀灭的细胞和/或细胞碎片以及培养基的细胞杀灭的全培养液。术语“发酵液”是指由细胞发酵产生、不经历或经历最低限的回收和/或纯化的制品。例如,当微生物培养株在允许蛋白质合成(例如,由宿主细胞的酶表达)并且将蛋白质分泌到细胞培养基中的碳受限的条件下孵育生长到饱和时,产生发酵液。发酵液可以含有在发酵结束时得到的发酵材料的未分级的或分级的内容物。典型地,发酵液是未分级的并且包含用过的培养基以及例如通过离心去除微生物细胞(例如,丝状真菌细胞)之后存在的细胞碎片。在一些实施例中,发酵液含有用过的细胞培养基、胞外酶以及有活力的和/或无活力的微生物细胞。在实施例中,该发酵液配制品和细胞组合物包含第一有机酸组分(包含至少一种1-5碳的有机酸和/或其盐)以及第二有机酸组分(包含至少一种6碳或更多碳的有机酸和/或其盐)。在具体实施例中,该第一有机酸组分是乙酸、甲酸、丙酸、其盐,或前述两种或更多种的混合物;并且该第二有机酸组分是苯甲酸、环己烷羧酸、4-甲基戊酸、苯乙酸、其盐,或前述两种或更多种的混合物。在一方面,该组合物包括一种或多种有机酸,并且任选地进一步含有杀灭的细胞和/或细胞碎片。在一个实施例中,从细胞杀灭的全培养液中去除这些杀灭的细胞和/或细胞碎片,以提供不含这些组分的组合物。这些发酵液配制品或细胞组合物可以进一步包括防腐剂和/或抗微生物(例如,抑菌)剂,包括但不限于山梨醇、氯化钠、山梨酸钾、以及本领域已知的其他试剂。这些发酵液配制品或细胞组合物可以进一步包含多种酶活性,如一种或多种(例如,若干种)选自下组的酶,该组由以下组成:纤维素酶、半纤维素酶、aa9多肽、纤维素诱导蛋白(cip)、过氧化氢酶、酯酶、棒曲霉素、漆酶、木质素分解酶、果胶酶、过氧化物酶、蛋白酶以及膨胀素。这些发酵液配制品或细胞组合物还可以包含选自下组的一种或多种(例如,若干种)酶,该组由以下组成:水解酶、异构酶、连接酶、裂解酶、氧化还原酶或转移酶,例如,α-半乳糖苷酶、α-葡糖苷酶、氨肽酶、淀粉酶、β-半乳糖苷酶、β-葡糖苷酶、β-木糖苷酶、糖酶、羧肽酶、过氧化氢酶、纤维二糖水解酶、纤维素酶、壳多糖酶、角质酶、环糊精糖基转移酶、脱氧核糖核酸酶、内切葡聚糖酶、酯酶、葡糖淀粉酶、转化酶、漆酶、脂肪酶、甘露糖苷酶、变位酶、氧化酶、果胶分解酶、过氧化物酶、植酸酶、多酚氧化酶、蛋白分解酶、核糖核酸酶、转谷氨酰胺酶或木聚糖酶。该细胞杀灭的全培养液或组合物可以含有在发酵结束时得到的发酵材料的未分级的内容物。典型地,该细胞杀灭的全培养液或组合物含有用过的培养基以及在微生物细胞(例如,丝状真菌细胞)生长至饱和、在碳限制条件下孵育以允许蛋白质合成(例如,纤维素酶和/或一种或多种葡糖苷酶的表达)之后存在的细胞碎片。在一些实施例中,该细胞杀灭的全培养液或组合物包括用过的细胞培养基、胞外酶和杀灭的丝状真菌细胞。在一些实施例中,可以使用本领域已知的方法来使细胞杀灭的全培养液或组合物中存在的微生物细胞透性化和/或裂解。如在此描述的,全培养液或细胞组合物典型地是液体,但是可以含有不溶性组分,如杀灭的细胞、细胞碎片、培养基组分和/或一种或多种不溶性酶。在一些实施例中,可除去不溶性组分以提供澄清的液体组合物。可以通过wo90/15861或wo2010/096673所述的该方法产生本发明的全培养液配制品和细胞组合物。本发明还涉及组合物,该组合物包含aa9溶解性多糖单加氧酶和选自下组的一种或多种添加的氧化还原酶,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该添加的氧化还原酶与该aa9溶解性多糖单加氧酶的蛋白质比率处于约1:250至约1:10,例如约1:200至约1:10、约1:150至约1:15、约1:100至约1:15、约1:75至约1:20、或约1:50至约1:25的范围内。通过以下实例进一步描述本发明,所述实例不应理解为对本发明范围的限制。实例菌株里氏木霉菌株rutc30是最初分离株qm6a(montenecourt和eveleigh,1979,adv.chem.ser.[化学进展丛书]181:289-301)的诱变的里氏木霉菌株。里氏木霉菌株btr213(o326pt)是里氏木霉rutc30的诱变菌株。里氏木霉菌株981-o8-d4是里氏木霉rutc30的诱变菌株。里氏木霉菌株btr-tl12-10是里氏木霉菌株btr213,其包含用seqidno:2的纤维二糖水解酶i的编码序列替换天然纤维二糖水解酶i编码序列,并用seqidno:4的纤维二糖水解酶ii的编码序列替换纤维二糖水解酶ii编码序列。里氏木霉菌株jfys99-19b4是里氏木霉菌株981-o8-d4,其包含用seqidno:2的纤维二糖水解酶i的编码序列替换天然纤维二糖水解酶i编码序列,并用seqidno:4的纤维二糖水解酶ii的编码序列替换天然纤维二糖水解酶ii编码序列。菌株a(里氏木霉q2b-1,o62j7z)是包含seqidno:6的aa9多肽的编码序列的里氏木霉btr-tl12-10菌株。菌株b(里氏木霉agjg005-35a,o622qv)是包含seqidno:6的aa9多肽和seqidno:8的过氧化氢酶的编码序列的里氏木霉菌株btr213-tl12-10。菌株c(里氏木霉qmji051-8b-4,o428dh)是包含seqidno:6的aa9多肽的编码序列的里氏木霉菌株jfys99-19b4。菌株d(里氏木霉agjg004-202a4,o422w5)是包含seqidno:6的aa9多肽和seqidno:8的过氧化氢酶的编码序列的里氏木霉菌株jfys99-19b4。培养基每升分批发酵培养基(fermentationbatchmedium)由以下构成:24g的右旋糖、40g的大豆粉、8g的(nh4)2so4、3g的k2hpo4、8g的k2so4、3g的caco3、8g的mgso4·7h2o、1g的柠檬酸、8.8ml的85%磷酸、1ml的消泡剂、和14.7ml的微量金属溶液。pda板由以下各项构成:39g的马铃薯右旋糖琼脂(difco)和补足至1升的去离子水。每升摇瓶培养基由以下组成:20g的甘油、10g的大豆粉、1.5g的(nh4)2so4、2g的kh2po4、0.2g的cacl2、0.4g的mgso4·7h2o、和0.2ml的微量金属溶液。每升微量金属溶液由以下组成:26.1g的feso4·7h2o、5.5g的znso4·7h2o、6.6g的mnso4·h2o、2.6g的cuso4·5h2o,和2g的柠檬酸。实例1:在ph4.5下共培养发酵菌株a和b在28℃,将菌株a和b各自在pda板生长4-7天。对于每种菌株,将各自含有100ml摇瓶培养基的三个500ml摇瓶用来自各pda板的两个塞子接种。在28℃,将摇瓶在定轨摇床上在200rpm下孵育48小时。将这些培养物用作种子,以用于更大规模发酵。使用总计150ml的种子培养物接种3升玻璃夹套发酵罐(applikonbiotechnology),根据下表1,每个发酵罐含有1.5升的发酵分批培养基。表1:共培养中若干个水平的过氧化氢酶表达菌株在ph4.5发酵。发酵发酵ph种子a种子b14.5100%0%34.595%5%54.590%10%74.575%25%将发酵罐维持在28℃的温度下,并且使用1030biocontroller(applikon生物技术公司(applikonbiotechnology))将ph控制在4.5+/-0.1的设定值。将空气以2.5l/min的速率添加到容器中,并用以1100rpm旋转的rushton叶轮搅拌培养液。将由右旋糖和磷酸构成的发酵补料培养基以0至10g/l/小时的速率给予165小时。每日取1ml样品并离心,并将上清液储存在-20℃直至蛋白质印迹分析(参见实例10)。在发酵结束时,从发酵罐中收获全发酵液,并以3000xg离心去除生物质。使用0.22μm滤器(密理博公司(millipore))过滤上清液。将过滤的上清液(“滤液”)储存在5℃-10℃。使用微板bcatm蛋白测定试剂盒(赛默飞世尔科技公司(thermofischerscientific))确定这些滤液的蛋白浓度,在该试剂盒中将牛血清白蛋白用作蛋白标准品。在测定之前,通过用纯化的seqidno:10的β-葡糖苷酶、seqidno:12的gh10木聚糖酶、和seqidno:14的β-木糖苷酶(分别为5%、5%、和3%的总蛋白质)替换滤液蛋白质来补充滤液的组合物,这产生混合物1、3、5、和7。实例2:在ph3.5下共培养发酵菌株a和b重复实例1,除了将ph控制在3.5+/-0.1的设定值,并且根据下表2将发酵物接种到菌株a和b的种子培养物中。表2:共培养中若干个水平的过氧化氢酶表达菌株在ph3.5发酵。发酵发酵ph种子a种子b23.5100%0%43.595%5%63.590%10%83.575%25%每日取1ml样品并离心,并将上清液储存在-20℃。在发酵结束时,从发酵罐中收获全发酵液,并以3000xg离心去除生物质。使用0.22μm滤器过滤上清液。将过滤的上清液(“滤液”)储存在5℃至10℃。使用微板bcatm蛋白测定试剂盒确定这些滤液的蛋白浓度,在该试剂盒中将牛血清白蛋白用作蛋白标准品。在测定之前,通过纯化的seqidno:10的β-葡糖苷酶、seqidno:12的gh10木聚糖酶、和seqidno:14的β-木糖苷酶(分别为5%、5%、和3%的总蛋白质)来补充这些滤液的组合物,这生产混合物2、4、6、和8。实例3:大剂量过氧化氢酶(bolus)的制备将supreme(诺维信公司(novozymesa/s),丹麦;lot#odn00025)(含有seqidno:8的过氧化氢酶的产物)在550mlsephadexg-25(gelifesciences)柱上在水中以100ml的两个等份脱盐。合并在280nm通过吸光度检测的所得洗脱蛋白质峰,使用0.22μm滤器无菌过滤,并在4℃下储存直至使用。使用10dg柱(伯乐实验室有限公司(bio-radlaboratories,inc.))将过滤池中的样品脱盐。使用其中使用牛血清白蛋白作为蛋白标准品的微板bcatm蛋白质测定试剂盒确定蛋白质浓度为8.7mg蛋白质/ml(至少60%是过氧化氢酶)。过氧化氢酶在此被称为“ts过氧化氢酶”。实例4:在ph5.0发酵菌株d与实例1中的发酵相似(但是在2.5立方米的发酵罐中,具有缩放量的分批和补料培养基)在ph5.0发酵菌株d。将所得培养液离心,过滤,蒸发浓缩,并与苯甲酸钠、山梨酸酯和葡萄糖混合。通过使用具有10,000mwco(德国赛多利斯公司(sartoriusag))的vivaflow200柱筒,通过与水的切向流动将该材料脱盐以除去苯甲酸钠、山梨酸盐和葡萄糖。基于280nm处的吸光度合并所得的脱盐浓缩物。对脱盐池中残余葡萄糖的hplc分析显示葡萄糖浓度为2.3mg/ml。将该池使用0.22μm滤器无菌过滤并在4℃下储存直至使用。使用econo-pac10dg柱将等分试样脱盐。使用其中使用牛血清白蛋白作为蛋白标准品的微板bcatm蛋白质测定试剂盒确定蛋白质浓度为177mg蛋白质/ml。过氧化氢酶在此被称为“trire过氧化氢酶”。实例5:在ph3.5和4.5发酵菌株c在28℃,将菌株c在pda板上生长4-7天。将3个500ml摇瓶(各含100ml摇瓶培养基)用来自固体板培养物的两个塞子接种,并在定轨摇床上以200rpm在28℃下孵育48小时。重复此步骤以产生用于5个发酵罐的足够的种子培养物(发酵9-13)。将这些培养物用作种子,以用于更大规模发酵。根据下表3,使用总共150ml的菌株c种子培养物接种3升玻璃夹套式发酵罐(applikon生物技术公司(applikonbiotechnology)),每个发酵罐含有1.5升补充有过氧化氢酶蛋白的发酵分批培养基(实例3和4)。表3将发酵罐维持在28℃的温度下,并且使用1030biocontroller(applikon生物技术公司(applikonbiotechnology))将ph控制在4.5或3.5+/-0.1的设定值。将空气以2.5l/min的速率添加到发酵罐中,并用以1100rpm旋转的rushton叶轮搅拌培养液。将由右旋糖和磷酸构成的发酵补料培养基以0至10g/l/小时的速率给予165小时。在发酵结束时,从发酵罐中收获全发酵液,并以3000xg离心去除生物质。使用0.22μm滤器过滤上清液。将过滤的上清液(滤液)储存在5℃-10℃。使用微板bcatm蛋白测定试剂盒(其中将牛血清白蛋白用作蛋白标准品)确定蛋白浓度。通过用纯化的seqidno:10的β-葡糖苷酶、seqidno:12的gh10木聚糖酶、和seqidno:14的β-木糖苷酶(分别为5%、5%、和3%的总蛋白质)替换滤液蛋白质来补充滤液的组合物,这生产混合物9、10、11、12、和13。实例6:预处理的玉米秸杆的活性测定针对发酵液滤液1-8水解经预处理的玉米穗轴和秸秆(pccs)以产生糖的能力,或针对它们水解纤维素的能力,使用荧光纤维素衰变(fcd)测定(wo2011/008785)通过降低的荧光测量发酵液滤液1-8的活性。将由稀释的酸预处理的玉米穗轴和玉米秸杆(pccs)组成的预处理的生物质混合物用水稀释,并在添加0.1ml的来自实例1和2的发酵液滤液1-8加0.5mg的纯化的seqidno:10的β-葡糖苷酶、0.5mg的纯化的seqidno:12的gh10木聚糖酶、和0.3mg的纯化的seqidno:14的β-木糖苷酶之前调节至ph5.0。最终的组合物为20g总重量,具有来自生物质的大约17%干重的固体。在通过96孔hv0.45μm膜板(密理博公司(millipore))上以3000rpm离心10分钟(使用rt7平板离心机(赛默飞世尔科技公司(thermofisherscientific))离心过滤水解产物浆液之后,在通过测量所得葡萄糖测量酶活性之前,将所得酶/生物质浆液在50℃下伴随12rpm不断搅拌孵育5天。在不立即使用时,将过滤的含糖等分试样冰冻于-20℃。通过以下方式测量稀释于0.005mh2so4中的样品的糖浓度:在65℃通过0.005mh2so4以0.6ml/分钟从4.6×250mmhpx-87h柱(伯乐实验室有限公司(bio-radlaboratories,inc.))洗脱,通过整合来自折射率检测(使用1100hplc(安捷伦技术公司(agilenttechnologies))通过纯糖样品校准)葡萄糖信号进行定量。图1显示了20g测定中pccs水解反应的结果。发酵液滤液1和2缺乏过氧化氢酶。尽管所有发酵液滤液都以相同的体积剂量(0.1ml过滤的发酵液)添加,并补充有相同量的纯化的seqidno:10的β-葡糖苷酶、seqidno:12的gh10木聚糖酶、和seqidno:14的β-木糖苷酶,结果表明,由于具有较高的单位体积水解活性或者每生产单位更大的活性,为产生过氧化氢酶的共培养的结果的酶组合物具有较高的葡萄糖产量。当用10%或25%种子共培养物在ph4.5发酵时,葡萄糖的这种改善大约为4%,而当用5%、10%或25%种子共培养物在ph3.5发酵时,大约为4%。根据wischman等人,2012,methodsenzymol.[酶方法]510:19-36,混合物1-8(实例1和2)的活性的测量通过以下实现:将适当的酶稀释物添加至生物质浆液中,在50℃孵育24小时至144小时,并测量由纤维素水解(由荧光增白剂(fb-28,西格玛公司(sigma))与纤维素的结合减少引起)引起的荧光信号的下降。以上描述的pccs通过在cosmos湿研磨机(essemm基团(essemmcorp))中湿磨6小时进一步改性,用as200筛选机(德国莱驰公司(retsch))通过425μm筛过筛,用水稀释,用60mm乙酸盐、180μmfb-28缓冲,调节ph,并且在121℃高压灭菌45分钟以产生为6.25%总干重固体(ph5.0)的物质。底物被称为fcd-gs-pccs;将200μl的fcd-gs-pccs放置在costar3364板(康宁公司(corning))中。将混合物1-8稀释25xv/v,并然后在96孔深孔板(axygen)中在milliq水中连续稀释两倍,得到8种酶稀释液,每种混合物为从25xv/v至3200xv/v。然后将混合物的五十μl来自板的每种稀释液添加到含有fcd-gs-pccs的板的每个相应孔中,相当于大约2μl至0.04μl原始发酵。使用alps300tm自动化实验室板密封机(abgeneinc.)将这些板热封。在水解开始时并在每个取样时间点之前反转并摇动96孔板,将反应混合物混合。在50mm乙酸钠(ph5.0)中,最终pccs浓度为50g/l,具有150μmfb-28。pccs水解是在50℃和55℃孵育下进行的,没有另外的搅拌,除了如上所述的取样过程。每个反应一式三份进行,绘制的值是重复的平均值。使用无酶和高酶对照(>5倍半最大消化)的荧光来确定0%(f最小)和100%(f最大)转化。如以下从测量的荧光(f样品)计算任何剂量的转化,其中在365处激发和在465处发射:转化率%=(f最大-f样品)/(f最大-f最小)。(方程1)图2显示混合物1、3、5和7(ph4.5发酵)在50℃和ph5.0下持续6天的剂量反应曲线图,这表明在共培养中增加表达过氧化氢酶的种子的百分比产生更高的纤维素水解。由于纤维素水解与葡萄糖的酶促释放相关,所以结果表明,当给予等体积的发酵液滤液时,较高的过氧化氢酶表达与更多的葡萄糖释放相关(参见wischmann等,2012,同上)。图3显示混合物2、4、6和8(ph3.5发酵)在50℃和ph5.0下持续6天的剂量反应曲线图,这表明在共培养中增加表达过氧化氢酶的种子的百分比产生更高的纤维素水解。由于纤维素水解与葡萄糖的酶促释放相关,所以结果表明,当给予等体积的发酵液滤液时,较高的过氧化氢酶表达与更多的葡萄糖释放相关(参见wischmann等,2012,同上)。实例7:共发酵液的储存稳定性将实例1和2中描述的发酵液1-8无菌过滤,等分到无菌96孔深孔板(axygen)中,使用alps300tm自动化实验室板密封器(abgeneinc.)密封,并在4℃、25℃、40℃和50℃无菌条件下储存4周。如实例1和2中描述,将所得样品补充到等同于混合物1至8的具有β-葡糖苷酶、gh10木聚糖酶、和β-木糖苷酶的混合物中,并使用如实例6中描述的cd测定进行测定,孵育7天。图4a显示通过混合物1、3、5和7(ph4.5发酵)所获得的转化率,针对每个储存温度与储存在4℃(100%的4℃样品)的样品获得的值以比率进行比较。从发酵1产生混合物1,发酵1不含表达过氧化氢酶的共培养种子菌株。在升高的温度下储存之后,所有含有过氧化氢酶的混合物3、5和7比混合物1显示出更高的稳定性(活性保留)。图4b显示通过混合物4、6和8(ph3.5发酵)所获得的转化率,针对每个储存温度与储存在4℃(100%的4℃样品)的样品获得的值以比率进行比较。从发酵2产生混合物2,发酵2无含表达过氧化氢酶的共培养种子菌株。在升高的温度下储存之后,所有含有过氧化氢酶的混合物4、6和8比混合物2显示出更高的稳定性(活性保留)。更确切地说,表达过氧化氢酶的共培养液比对照混合物在25℃显示5%至9%更高的稳定性、在40℃储存显示1%至12%更高的稳定性、并且在50℃储存显示3%至7%更高的稳定性。实例8:发酵中添加有大剂量过氧化氢酶的发酵液的储存稳定性如实例7中描述的将实例5中描述的过滤的发酵液在无菌条件下在4℃、25℃、和40℃下储存4周,然后等同地补充到来自实例5的混合物9、10、11和12(如先前描述的具有纯化的seqidno:10的β-葡糖苷酶、seqidno:12的gh10木聚糖酶、和seqidno:14的β-木糖苷酶)中。如实例6中描述的测量这些混合物的连续稀释物的水解活性,在55℃下孵育5天,产生与图2和3中显示的相似的水解曲线。基于该方程生成接近水解曲线的曲线其中对于每个样品稀释曲线的常数p(幂函数)和k(水解的半最大值)通过excel插件solver(微软)优化以最小化误差的平方和以拟合酶负载x(以培养基中的mg蛋白质计,或以培养液中的u计)并计算转化率%。然后可使用这些常数来插值达到所需的转化率(例如80%的转化率)目标(t)必需的酶负载:达到作为目标(t)的恒定水解百分比的酶负载的计算允许比较不同酶样品的效率,例如达到80%转化率所需的μl发酵液/g纤维素。图5显示了从实例3或实例4衍生的过氧化氢酶蛋白质添加对发酵液11和12的储存性能的益处,因为在储存材料的所有温度下,发酵中添加有过氧化氢酶的混合物11和12优于缺乏过氧化氢酶的混合物9(ph3.5)和10(ph4.5)(由于达到80%转化率的目标需要较少μl)。储存性能的改善导致在4℃和25℃储存后需要的μl减少15%至18%,并且在40℃储存后需要的μl减少9%至15%。实例9:在发酵后向混合物13添加supreme的影响如先有实例,测量来自实例5的菌株c(不过表达过氧化氢酶的木霉属菌株)的过滤的发酵培养液13的蛋白质含量,并通过如下制作混合物:用分别以5%、5%和3%的纯化的seqidno:10的β-葡糖苷酶、seqidno:12的gh10木聚糖酶、和seqidno:14的β-木糖苷酶替换发酵液蛋白质进行补充、并且用原样使用的supreme替换,使用微板bcatm蛋白质测定试剂盒测量为13.5mg/ml(其中牛血清白蛋白作为蛋白标准品),最终混合物具有0%、0.1%、0.5%、1%和2%w/w蛋白质的supreme蛋白质。如实例6中描述,在ph5和55℃下,持续5天,通过fcd测量混合物13在水解中的活性,并且通过如实例8中的拟合曲线的插值计算达到80%转化率所需的μl/g纤维素负载。图6显示,在发酵后添加supreme(过氧化氢酶来源)不会显著改善性能(具有2%supreme蛋白质的最好的混合物比0%supreme混合物好2%,但标准偏差为3%-6%)。这种益处不及在发酵期间添加过氧化氢酶时所观察到的,如图5中的混合物11或12、实例8中。实例10:共培养的蛋白质印迹在兔中产生抗体作为针对合成肽kqafgdtddfskhg(seqidno:15)的多克隆应答,代表seqidno:2的纤维二糖水解酶i(残基371-384)的部分序列。该抗体被称为αcbh1第一抗体。将来自实例1和2的过滤的发酵液1-8稀释为在5μl水中大约1μg蛋白质,然后用2xlaemlli缓冲液(伯乐实验室有限公司(bio-radlaboratories,inc.))和1xtcep(赛默科技公司(thermoscientific))进一步1:1稀释,并在95℃加热5分钟、冷却、离心,并加载到26孔10%tgxstainfreesds-page凝胶(伯乐实验室有限公司(bio-radlaboratories,inc.))上。将凝胶在300伏特运行20分钟。使用半干turbotm印迹系统(伯乐实验室有限公司(bio-radlaboratories,inc.))将凝胶转移至immune-blotpvdf膜(伯乐实验室有限公司(bio-radlaboratories,inc.))上。在摇臂上在室温下,将该膜在ph7.5的tris缓冲盐水(tbs;20mmtris-500mmnacl)中洗涤两次持续5分钟,并在tbst(tbs+0.05%20)中用1%bsa封闭缓冲液孵育1小时。随后的所有步骤包括用tbst进行三次洗涤步骤持续5分钟。将印迹与用tbst以1/10,000稀释的αcbh1第一抗体(科文斯公司(covance))孵育1小时,随后与用tbst以1/10,000稀释的第二抗体山羊抗兔hrp(杰克逊免疫研究实验室(jacksonimmunoresearchlaboratories))孵育1小时。在使用chemidocmp(伯乐实验室有限公司(bio-radlaboratories,inc.))上的化学发光设置进行印迹检测之前,将蛋白质印迹用supersignalwestpicosubstrate(赛默科技公司(thermoscientific))在tbs中进行最后洗涤。通过imagelab(伯乐实验室有限公司(bio-radlaboratories,inc.))的默认设置定量印迹强度。图7显示得到的蛋白质印迹图像,其中泳道1-8代表根据实例1和2生产的过滤的发酵液1-8,与表达过氧化氢酶的菌株的共培养如表1中概述的。大约37,000道尔顿的条带代表纤维二糖水解酶i的断裂,其在具有aa9多肽表达的样品中发生,但是在ph4.5发酵时无过氧化氢酶表达。表达过氧化氢酶的共培养样品(泳道3-8)不显示该带。泳道11-16分别代表发酵1(0%过氧化氢酶过量表达种子b)的从第2天至第7天每天样品的bca微板测定蛋白质标准化(1μg)的负载,而泳道17-22代表发酵5的等同样品(10%过氧化氢酶过表达种子b)。在无过氧化氢酶共培养的发酵中可见大约37,000道尔顿的片段的显影,而与来自产过氧化氢酶的菌株b的10%种子的共培养中不存在片段,表明在发酵过程中发生断裂,并且过氧化氢酶表达将这种断裂减少到肉眼不可见的水平。实例11:在发酵过程中添加过氧化氢酶蛋白质的蛋白质印迹如实例10中描述,对来自实例5发酵液滤液(发酵9至12,参见表3,分别代表图8中泳道1至4)的5μl水中大约1μg发酵液蛋白进行处理。图8显示来自发酵10的高含量的37,000道尔顿片段,如泳道2显示。与过氧化氢酶未与种子一起添加的泳道2相比,在发酵(发酵11和12)开始时过氧化氢酶蛋白质与种子一起添加在泳道3和4中分别显示37,000道尔顿片段的大量减少,表明在用过氧化氢酶发酵之后这种蛋白质更高的完整性。泳道1显示在ph3.5下生长的发酵9,其中观察到较少量的37,000道尔顿片段。本发明通过以下编号的段落来进一步说明:段落[1]:一种抑制酶组合物或其组分的aa9溶解性多糖单加氧酶催化的失活的方法,所述方法包括:将选自下组的一种或多种氧化还原酶添加至该酶组合物中,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活。段落[2]:如段落1所述的方法,其中该一种或多种氧化还原酶是过氧化氢酶。段落[3]:如段落1所述的方法,其中该一种或多种氧化还原酶是漆酶。段落[4]:如段落1所述的方法,其中该一种或多种氧化还原酶是过氧化物酶。段落[5]:如段落1所述的方法,其中该一种或多种氧化还原酶是超氧化物歧化酶。段落[6]:如段落1所述的方法,其中该一种或多种氧化还原酶是选自下组的两种或更多种氧化还原酶的组合,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶。段落[7]:如段落1-6中任一项所述的方法,其中该酶组合物包含选自下组的一种或多种组分,该组由以下组成:水解酶、异构酶、连接酶、裂解酶、氧化还原酶、或转移酶。段落[8]:如段落1-6中任一项所述的方法,其中该酶组合物包含选自下组的一种或多种组分,该组由以下组成:纤维素酶、aa9多肽、半纤维素酶、纤维素诱导蛋白、酯酶、棒曲霉素、木质素分解酶、果胶酶、蛋白酶、和膨胀素。段落[9]:如段落8所述的方法,其中该纤维素酶是选自下组的一种或多种酶,该组由以下组成:内切葡聚糖酶、纤维二糖水解酶、以及β-葡糖苷酶。段落[10]:如段落8所述的方法,其中该半纤维素酶是选自下组的一种或多种酶,该组由以下组成:木聚糖酶、乙酰木聚糖酯酶、阿魏酸酯酶、阿拉伯呋喃糖苷酶、木糖苷酶、以及葡糖醛酸糖苷酶。段落[11]:如段落1-10中任一项所述的方法,其中该添加的氧化还原酶与该aa9溶解性多糖单加氧酶的蛋白质比率处于约1:250至约1:10,例如约1:200至约1:10、约1:150至约1:15、约1:100至约1:15、约1:75至约1:20、或约1:50至约1:25的范围内。段落[12]:如段落1-11中任一项所述的方法,其中与不存在一种或多种添加的氧化还原酶相比,在存在一种或多种添加的氧化还原酶下,对该aa9溶解性多糖单加氧酶催化的失活的抑制的量更高。段落[13]:一种用于增加酶组合物的产生的方法,所述方法包括:(a)在选自下组的一种或多种添加的氧化还原酶的存在下,发酵宿主细胞以产生该酶组合物,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活,并且其中与在该一种或多种氧化还原酶不存在下产生的酶组合物的量相比,在该一种或多种添加的氧化还原酶的存在下产生的酶组合物的量更高;并且任选地(b)回收该酶组合物。段落[14]:如段落13所述的方法,其中该一种或多种添加的氧化还原酶是过氧化氢酶。段落[15]:如段落13所述的方法,其中该一种或多种添加的氧化还原酶是漆酶。段落[16]:如段落13所述的方法,其中该一种或多种添加的氧化还原酶是过氧化物酶。段落[17]:如段落13所述的方法,其中该一种或多种添加的氧化还原酶是超氧化物歧化酶。段落[18]:如段落13所述的方法,其中该一种或多种添加的氧化还原酶是选自下组的两种或更多种氧化还原酶的组合,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶。段落[19]:如段落13-18中任一项所述的方法,其中该宿主细胞包含相对于该宿主细胞是天然的aa9溶解性多糖单加氧酶。段落[20]:如段落13-18中任一项所述的方法,其中该宿主细胞包含相对于该宿主细胞是异源的aa9溶解性多糖单加氧酶。段落[21]:如段落13-18中任一项所述的方法,其中该宿主细胞包含相对于该宿主细胞是天然的aa9溶解性多糖单加氧酶以及相对于该宿主细胞是异源的aa9溶解性多糖单加氧酶。段落[22]:如段落13-21中任一项所述的方法,其中该酶组合物包含选自下组的一种或多种组分,该组由以下组成:水解酶、异构酶、连接酶、裂解酶、氧化还原酶、或转移酶。段落[23]:如段落13-21中任一项所述的方法,其中该酶组合物包含选自下组的一种或多种组分,该组由以下组成:纤维素酶、aa9多肽、半纤维素酶、纤维素诱导蛋白、酯酶、棒曲霉素、木质素分解酶、果胶酶、蛋白酶、和膨胀素。段落[24]:如段落23所述的方法,其中该纤维素酶是选自下组的一种或多种酶,该组由以下组成:内切葡聚糖酶、纤维二糖水解酶、以及β-葡糖苷酶。段落[25]:如段落23所述的方法,其中该半纤维素酶是选自下组的一种或多种酶,该组由以下组成:木聚糖酶、乙酰木聚糖酯酶、阿魏酸酯酶、阿拉伯呋喃糖苷酶、木糖苷酶、以及葡糖醛酸糖苷酶。段落[26]:如段落13-25中任一项所述的方法,其中将该一种或多种添加的氧化还原酶添加至发酵中。段落[27]:如段落13-25中任一项所述的方法,其中该一种或多种添加的氧化还原酶是由宿主细胞重组产生的。段落[28]:如段落13-25中任一项所述的方法,其中该一种或多种添加的氧化还原酶是通过重组细胞与第二宿主细胞的共培养重组产生的。段落[29]:如段落13-25中任一项所述的方法,其中将该一种或多种添加的氧化还原酶添加至发酵中,并且该一种或多种添加的氧化还原酶是由宿主细胞重组产生的。段落[30]:如段落13-25中任一项所述的方法,其中将该一种或多种添加的氧化还原酶添加至发酵中,并且该一种或多种添加的氧化还原酶是通过重组细胞与第二宿主细胞的共培养重组产生的。段落[31]:如段落13-25中任一项所述的方法,其中该一种或多种添加的氧化还原酶是由宿主细胞重组产生的,以及通过重组细胞与第二宿主细胞的共培养重组产生的。段落[32]:如段落13-25中任一项所述的方法,其中将该一种或多种添加的氧化还原酶添加至发酵中,该一种或多种添加的氧化还原酶是由宿主细胞重组产生的,以及通过重组细胞与第二宿主细胞的共培养重组产生的。段落[33]:如段落13-32中任一项所述的方法,其中该添加的氧化还原酶与该aa9溶解性多糖单加氧酶的蛋白质比率处于约1:250至约1:10,例如约1:200至约1:10、约1:150至约1:15、约1:100至约1:15、约1:75至约1:20、或约1:50至约1:25的范围内。段落[34]:如段落13-33中任一项所述的方法,其中与不存在一种或多种添加的氧化还原酶相比,在存在一种或多种添加的氧化还原酶下,对该aa9溶解性多糖单加氧酶催化的失活的抑制更高。段落[35]:一种用于稳定酶组合物的方法,该方法包括将选自下组的一种或多种氧化还原酶添加至该酶组合物中,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该酶组合物包含aa9溶解性多糖单加氧酶和一种或多种酶组分,并且其中该一种或多种添加的氧化还原酶抑制该酶组合物的一种或多种酶组分的aa9溶解性多糖单加氧酶催化的失活。段落[36]:如段落35所述的方法,其中该一种或多种氧化还原酶是过氧化氢酶。段落[37]:如段落35所述的方法,其中该一种或多种氧化还原酶是漆酶。段落[38]:如段落35所述的方法,其中该一种或多种氧化还原酶是过氧化物酶。段落[39]:如段落35所述的方法,其中该一种或多种氧化还原酶是超氧化物歧化酶。段落[40]:如段落35所述的方法,其中该一种或多种氧化还原酶是选自下组的两种或更多种氧化还原酶的组合,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶。段落[41]:如段落35-40中任一项所述的方法,其中该酶组合物包含选自下组的一种或多种组分,该组由以下组成:水解酶、异构酶、连接酶、裂解酶、氧化还原酶、或转移酶。段落[42]:如段落35-40中任一项所述的方法,其中该酶组合物包含选自下组的一种或多种组分,该组由以下组成:纤维素酶、aa9多肽、半纤维素酶、纤维素诱导蛋白、酯酶、棒曲霉素、木质素分解酶、果胶酶、蛋白酶、和膨胀素。段落[43]:如段落42所述的方法,其中该纤维素酶是选自下组的一种或多种酶,该组由以下组成:内切葡聚糖酶、纤维二糖水解酶、以及β-葡糖苷酶。段落[44]:如段落42所述的方法,其中该半纤维素酶是选自下组的一种或多种酶,该组由以下组成:木聚糖酶、乙酰木聚糖酯酶、阿魏酸酯酶、阿拉伯呋喃糖苷酶、木糖苷酶、以及葡糖醛酸糖苷酶。段落[45]:如段落35-44中任一项所述的方法,其中该添加的氧化还原酶与该aa9溶解性多糖单加氧酶的蛋白质比率处于约1:250至约1:10,例如约1:200至约1:10、约1:150至约1:15、约1:100至约1:15、约1:75至约1:20、或约1:50至约1:25的范围内。段落[46]:如段落35-45中任一项所述的方法,其中与不存在一种或多种添加的氧化还原酶相比,在存在一种或多种添加的氧化还原酶下,对该aa9溶解性多糖单加氧酶催化的失活的抑制的量更高。段落[47]:一种组合物,该组合物包含aa9溶解性多糖单加氧酶和选自下组的一种或多种添加的氧化还原酶,该组由以下组成:过氧化氢酶、漆酶、过氧化物酶、和超氧化物歧化酶,其中该添加的氧化还原酶与该aa9溶解性多糖单加氧酶的蛋白质比率处于约1:250至约1:10,例如约1:200至约1:10、约1:150至约1:15、约1:100至约1:15、约1:75至约1:20、或约1:50至约1:25的范围内。本申请中描述并且要求保护的本发明不限于在此披露的特定方面的范围,因为这些方面旨在说明本发明的若干方面。预期任何等效方面都处于本发明的范围内。实际上,除本申请中所示和描述的那些之外,对于本领域的技术人员而言本发明的多种修改将从前述的描述变得显而易见。这样的修改也旨在落入所附权利要求书的范围内。在有冲突的情况下,以包括定义的本披露为准。序列表<110>诺维信公司(novozymesa/s)k·麦克法兰(mcfarland,keith)a·特吉瑞安(tejirian,ani)d·阿克尔赫尔姆(akerhielm,derek)<120>抑制酶组合物的aa9溶解性多糖单加氧酶催化的失活的方法<130>13277-wo-pct<150>us62/214,373<151>2015-09-04<160>14<170>patentin版本3.5<210>1<211>1730<212>dna<213>雷塞特纳斯篮状菌(talaromycesleycettanus)<400>1atggcgtccctcttctctttcaagatgtacaaggctgctctcgtcctgtcttctctcctg60gccgctacgcaggctcagcaggccggcactctcacgacggagacccatccgtccctgaca120tggcagcaatgctcggccggtggcagctgcaccacccagaacggcaaggtcgtcatcgat180gcgaactggcgttgggtgcacagcacgagcggaagcaacaactgctacaccggcaatacc240tgggacgctaccctatgccctgacgatgtgacctgcgccgccaactgtgcgctggacggt300gccgactactcgggcacctacggagtgaccaccagcggcaactccctccgcctcaacttc360gtcacccaggcgtcacagaagaacgtcggctcccgtctttacctgatggagaatgacaca420acctaccagatcttcaagctgctgaaccaggagttcacctttgatgtcgatgtgtccaac480ctgccgtaagtgacttaccatgaacccctgacgctatcttcttgttggctcccagctgac540tggccaattcaagctgcggcttgaacggtgctctctacctggtggccatggacgccgatg600gtggcatggccaagtaccccaccaacaaggctggtgccaagtacggtaccgggtactgcg660actcccagtgtccccgcgacctcaagttcatcaatggcgaggccaacgtcgagggctggc720agccgtcgtccaacgatcccaactctggcattggcaaccacggatcctgctgcgcggaga780tggatatctgggaggccaacagcatctccaatgctgtcactccccacccgtgcgacactc840ccggccaggtgatgtgcaccggtaacaactgcggtggcacatacagcactactcgctatg900cgggcacttgcgatcccgacggctgcgacttcaacccctaccgcatgggcaaccacagct960tctacggccctaaacagatcgtcgataccagctcgaagttcaccgtcgtgacgcagttcc1020tcacggatgacggcacctccaccggcaccctctctgaaatccgccgcttctatgtccaga1080acggccaggtgatcccgaactcggtgtcgaccatcagtggcgtgagcggcaactccatca1140ccaccgagttctgcactgcccagaagcaggccttcggcgacacggacgacttctcaaagc1200acggcggcctgtccggcatgagcgctgccctctctcagggtatggttctggtcatgagtc1260tgtgggatgatgtgagtttgatggacaaacatgcgcgttgacaaagagtcaagcagctga1320ctgagatgttacagcacgccgccaacatgctctggctcgacagcacctacccgaccaacg1380cgacctcctccacccccggtgccgcccgtggaacctgcgacatctcgtccggtgtccctg1440cggatgtcgaatccaacgaccccaacgcctacgtggtctactcgaacatcaaggttggtc1500ccatcggctcgaccttcagcagcagcggctctggatcttcttcctctagctccaccacta1560ccacgaccaccgcttccccaaccaccacgacctcctccgcatcgagcaccggcactggag1620tggcacagcactggggccagtgtggtggacagggctggaccggccccacaacctgcgtca1680gcccttatacttgccaggagctgaacccttactactaccagtgtctgtaa1730<210>2<211>532<212>prt<213>雷塞特纳斯篮状菌<400>2metalaserleupheserphelysmettyrlysalaalaleuvalleu151015serserleuleualaalathrglnalaglnglnalaglythrleuthr202530thrgluthrhisproserleuthrtrpglnglncysseralaglygly354045sercysthrthrglnasnglylysvalvalileaspalaasntrparg505560trpvalhisserthrserglyserasnasncystyrthrglyasnthr65707580trpaspalathrleucysproaspaspvalthrcysalaalaasncys859095alaleuaspglyalaasptyrserglythrtyrglyvalthrthrser100105110glyasnserleuargleuasnphevalthrglnalaserglnlysasn115120125valglyserargleutyrleumetgluasnaspthrthrtyrglnile130135140phelysleuleuasnglngluphethrpheaspvalaspvalserasn145150155160leuprocysglyleuasnglyalaleutyrleuvalalametaspala165170175aspglyglymetalalystyrprothrasnlysalaglyalalystyr180185190glythrglytyrcysaspserglncysproargaspleulyspheile195200205asnglyglualaasnvalgluglytrpglnproserserasnasppro210215220asnserglyileglyasnhisglysercyscysalaglumetaspile225230235240trpglualaasnserileserasnalavalthrprohisprocysasp245250255thrproglyglnvalmetcysthrglyasnasncysglyglythrtyr260265270serthrthrargtyralaglythrcysaspproaspglycysaspphe275280285asnprotyrargmetglyasnhisserphetyrglyprolysglnile290295300valaspthrserserlysphethrvalvalthrglnpheleuthrasp305310315320aspglythrserthrglythrleusergluileargargphetyrval325330335glnasnglyglnvalileproasnservalserthrileserglyval340345350serglyasnserilethrthrgluphecysthralaglnlysglnala355360365pheglyaspthraspasppheserlyshisglyglyleuserglymet370375380seralaalaleuserglnglymetvalleuvalmetserleutrpasp385390395400asphisalaalaasnmetleutrpleuaspserthrtyrprothrasn405410415alathrserserthrproglyalaalaargglythrcysaspileser420425430serglyvalproalaaspvalgluserasnaspproasnalatyrval435440445valtyrserasnilelysvalglyproileglyserthrpheserser450455460serglyserglyserserserserserserthrthrthrthrthrthr465470475480alaserprothrthrthrthrserseralaserserthrglythrgly485490495valalaglnhistrpglyglncysglyglyglnglytrpthrglypro500505510thrthrcysvalserprotyrthrcysglngluleuasnprotyrtyr515520525tyrglncysleu530<210>3<211>1898<212>dna<213>雷塞特纳斯篮状菌<400>3atgcggtctctcctggctcttgcccctaccctgctcgcgcctgttgttcaggctcagcaa60accatgtggggtcaatgtaagttcttttcactgcttaccatgtataatctttgatatcaa120gcatcatatctgactcacgttttaggcggtggtcagggctggaccggacctaccatctgt180gtagcaggcgcgacatgcagcacacagaacccttgtaagtcgggccttcatcaaaacttc240aacatcaccacctcgatggagcaggagttgacctgatctttacccttagggtatgcgcag300tgcaccccagcacctaccgcgccgacgaccttgcaaacaacaactacgacgagctcgaaa360tcgtccacgaccacgagctcgaagtcgtccacgaccacaggtggaagtggcggtggaact420acgacctcaacgtcagccaccatcaccgcggctccatctggtaacccatactccggatac480cagctctatgtgaaccaggaatactcgtccgaggtgtacgcgtctgctattccttccctt540accggcactctggtcgcgaaggcaagcgccgcggcagaggtgccatctttcctgtggctg600taagtttttttgaccttgaatgaacgccctgtcctctacgagtggccgcaggagctaatt660gagatgccaatgaacagggacactgcctccaaggtgccactgatgggcacttacttgcag720gatatccaggcgaagaacgctgctggcgccaaccccccatatgccggtcaattcgtggtt780tacgacttgccggatcgtgattgcgctgcattggccagcaatggagagtactccattgct840aacaatggtgttgccaactacaaggcttacatcgactccatccgcgcgcttcttgttcaa900tactcgaacgtccatgtcatccttgtgatcggtgagctattgcagtctcgctttaaagca960tttgactagatcaatgtcgctaatggtacctaccgcacagagcccgacagcttggccaac1020cttgtcaccaacctgaatgttcagaagtgtgctaatgctcagagtgcttacctggagtgc1080atcaactatgccctcactcagttgaacctcaagaacgttgctatgtacatcgatgctggt1140gcgtgaaccttccctagtcagcccaaaataactgaaataaagagacggagtgtactgatt1200gtcatgcaggtcatgctggatggctcggctggcccgccaaccttagcccggccgctcaac1260tctttgcttccgtataccagaatgcaagctccccagctgccgttcgcggcctggcaacca1320acgtggccaactataatgcctggtcgatcgccacttgcccatcttacacccaaggcgacc1380ccaactgcgacgagcagaaatacatcaacgctctggctccattgcttcagcaacagggat1440ggtcatcagttcactttatcaccgataccggtaagtctgcctgtcctgccaaccatgcgt1500tcaagagcgttgcaatcctaaccatgctggtatcttccaggccgtaacggtgtccagcct1560accaagcagaatgcctggggtgactggtgcaacgttatcggaaccggcttcggtgtccgt1620cccaccaccaacactggcgatccattggaggatgctttcgtctgggtcaagcctggtggt1680gagagtgatggtacttccaactccacttcgcctcgctacgacgcccactgcggttacagt1740gatgctcttcagcctgctcctgaggctggtacctggttcgaggtaagcttctgcatactg1800agatcgagaatcctgaaagggttaacctgctaatgcttcggtgtttgatataggcttact1860ttgagcaactccttaccaacgccaacccctctttctaa1898<210>4<211>464<212>prt<213>雷塞特纳斯篮状菌<400>4metargserleuleualaleualaprothrleuleualaprovalval151015glnalaglnglnthrmettrpglyglncysglyglyglnglytrpthr202530glyprothrilecysvalalaglyalathrcysserthrglnasnpro354045trptyralaglncysthrproalaprothralaprothrthrleugln505560thrthrthrthrthrserserlysserserthrthrthrserserlys65707580serserthrthrthrglyglyserglyglyglythrthrthrserthr859095seralathrilethralaalaproserglyasnprotyrserglytyr100105110glnleutyrvalasnglnglutyrsersergluvaltyralaserala115120125ileproserleuthrglythrleuvalalalysalaseralaalaala130135140gluvalproserpheleutrpleuaspthralaserlysvalproleu145150155160metglythrtyrleuglnaspileglnalalysasnalaalaglyala165170175asnproprotyralaglyglnphevalvaltyraspleuproasparg180185190aspcysalaalaleualaserasnglyglutyrserilealaasnasn195200205glyvalalaasntyrlysalatyrileaspserileargalaleuleu210215220valglntyrserasnvalhisvalileleuvalilegluproaspser225230235240leualaasnleuvalthrasnleuasnvalglnlyscysalaasnala245250255glnseralatyrleuglucysileasntyralaleuthrglnleuasn260265270leulysasnvalalamettyrileaspalaglyhisalaglytrpleu275280285glytrpproalaasnleuserproalaalaglnleuphealaserval290295300tyrglnasnalaserserproalaalavalargglyleualathrasn305310315320valalaasntyrasnalatrpserilealathrcysprosertyrthr325330335glnglyaspproasncysaspgluglnlystyrileasnalaleuala340345350proleuleuglnglnglnglytrpserservalhispheilethrasp355360365thrglyargasnglyvalglnprothrlysglnasnalatrpglyasp370375380trpcysasnvalileglythrglypheglyvalargprothrthrasn385390395400thrglyaspproleugluaspalaphevaltrpvallysproglygly405410415gluseraspglythrserasnserthrserproargtyraspalahis420425430cysglytyrseraspalaleuglnproalaproglualaglythrtrp435440445pheglualatyrphegluglnleuleuthrasnalaasnproserphe450455460<210>5<211>835<212>dna<213>青霉属物种<400>5atgctgtcttcgacgactcgcaccctcgcctttacaggccttgcgggccttctgtccgct60cccctggtcaaggcccatggctttgtccagggcattgtcatcggtgaccaattgtaagtc120cctctcttgcagttctgtcgattaactgctggactgcttgcttgactccctgctgactcc180caacagctacagcgggtacatcgtcaactcgttcccctacgaatccaacccaccccccgt240catcggctgggccacgaccgccaccgacctgggcttcgtcgacggcacaggataccaagg300cccggacatcatctgccaccggaatgcgacgcccgcgccgctgacagcccccgtggccgc360cggcggcaccgtcgagctgcagtggacgccgtggccggacagccaccacggacccgtcat420cacctacctggcgccgtgcaacggcaactgctcgaccgtcgacaagacgacgctggagtt480cttcaagatcgaccagcagggcctgatcgacgacacgagcccgccgggcacctgggcgtc540ggacaacctcatcgccaacaacaatagctggaccgtcaccattcccaacagcgtcgcccc600cggcaactacgtcctgcgccacgagatcatcgccctgcactcggccaacaacaaggacgg660cgcccagaactacccccagtgcatcaacatcgaggtcacgggcggcggctccgacgcgcc720tgagggtactctgggcgaggatctctaccatgacaccgacccgggcattctggtcgacat780ttacgagcccattgcgacgtataccattccggggccgcctgagccgacgttctag835<210>6<211>253<212>prt<213>青霉属物种<400>6metleuserserthrthrargthrleualaphethrglyleualagly151015leuleuseralaproleuvallysalahisglyphevalglnglyile202530valileglyaspglnphetyrserglytyrilevalasnserphepro354045tyrgluserasnproproprovalileglytrpalathrthralathr505560aspleuglyphevalaspglythrglytyrglnglyproaspileile65707580cyshisargasnalathrproalaproleuthralaprovalalaala859095glyglythrvalgluleuglntrpthrprotrpproaspserhishis100105110glyprovalilethrtyrleualaprocysasnglyasncysserthr115120125valasplysthrthrleugluphephelysileaspglnglnglyleu130135140ileaspaspthrserproproglythrtrpalaseraspasnleuile145150155160alaasnasnasnsertrpthrvalthrileproasnservalalapro165170175glyasntyrvalleuarghisgluileilealaleuhisseralaasn180185190asnlysaspglyalaglnasntyrproglncysileasnilegluval195200205thrglyglyglyseraspalaprogluglythrleuglygluaspleu210215220tyrhisaspthraspproglyileleuvalaspiletyrgluproile225230235240alathrtyrthrileproglyproprogluprothrphe245250<210>7<211>2502<212>dna<213>橙色嗜热子囊菌<400>7atgcgcgcaattggacttctgccaggcatcatcggcattgctggtgctgcctgtccttac60atgacaggcgagctgccgcgctccttcgccgagaaccctcatgctatcaaccgtcgtgct120gagggtggtggtggtgccgctgccgagacggagaagttcctgtctcagttctacctgaac180gacaacgacaccttcatgaccaccgatgttggcggtccaattgaggatcagaacagtctc240agcgctggtgacagaggtcctaccctgctggaggacttcatcctccgtcaaaagatccag300cgctttgaccatgagcgggtaggttgatctttactttcggccttcttcgagcggggtgat360attaaaacaggtaataggtgcccgagcgtgctgtccatgcccgaggagcgggagcgcatg420gcgtgttcacatcctacgcagactggtccaacatcactgccgcttccttcctgtctgctg480caggaaaggagacacctgtctttgtccggttctccactgtagcaggaagcagaggaagcg540cagacacggcgcgtgacgtgcacggtttcgcgacgaggttctacacggatgaagggaact600tcggtaggcaactatcatgctctctttaaatgttctcgatctgacagccagcagacattg660tcggcaacaacatccctgtcttcttcattcaagatgcgatccagttccccgacctgatcc720atgctgtcaagcccagcccgaacaacgagatccctcaggccgcaaccgcccatgactctg780cctgggactttttcagccagcagccgagctctttgcatactctgttctgggctatggccg840gtcatggcattcctcgttcctacaggaacatggatggcttcggcatccacaccttccgct900ttgtgacggacgatggagcttccaagctcgtcaagttccactggacgtcgctgcagggca960aggcgagccttgtgtgggaagaggcacaggccgtggctggaaagaacgcggactatcacc1020gccaggacttgtgggacgcaatcgaggctggaaggtaccctgagtgggaggtaggctctc1080cctgctatgtatggatgtgccagaagcttaataatggcctagctcggcgtgcaaatcatg1140gatgaggaagaccagctgcgctttggcttcgatctgttggacccgaccaagatcgttccc1200gaggaatacgtgcccatcacgaagctcggaaagatgcagctcaaccgcaacccgctgaac1260tacttcgccgagactgaacagatcatggtcagttcgccaccgtgttcggttgctcgttgc1320tgaagtgctaacttgcaacagttccaaccgggtcacgttgtccgtggcattgatttcacc1380gaggaccctctgctccagggacgtctcttctcttacctcgacacccagctcaaccgccac1440ggaggtccgaacttcgagcagatccccatcaaccggccacgcactccaattcacaacaac1500aaccgtgacggagccggtatgctagcccatgtattcctttctttatgcatttttatatga1560tgcgttctaacggcaacagcgcaaatgtacatccccctgaacaaggcggcgtacaccccc1620aacactctgaacaacggctcccccaagcaggccaaccagacggtcggaaagggcttcttc1680acgactccaggccggacggcaagcggcaggcttgtgcgcgccgtcagctcaaccttcgcc1740gacgtctggtcgcagcctcgtctgttctacaactccctcgtgccggcggagcagcagttc1800ctgatcaacgcgatccgctttgagacggcccacatcacgagcgacgtcgtgaagaacaac1860gtcatcatccagctgaaccgcgtgagcaacaacctcgccaagagagtcgcccgggccatc1920ggtgtcgcggagcccgagccagacccaaccttgtaccacaacaacaagaccgccaacgtc1980ggggtgttcggcaagccgctcgccagactcgacggcctgcaggtcggggtcctcgccacc2040gtcaacaagcccgactcgatcaagcaggccgccagcctgaaggccagcttcgcggcggac2100aacgtcgacgtcaaggtcgtcgcggagcgcctcgccgacggcgtcgacgagacctactcg2160gccgccgacgcggtcaacttcgacgccatcctggtcgccaacggcgctgagggcctcttc2220gcgcgcgacagcttcaccgccaggccggccaactcgaccaccgcgacgctctaccccgcg2280ggccgcccgctccagatcctggtcgacgggttccgctacggcaagccggtcggggcgctc2340ggcagcggcgccaaggcgctcgacgcagcggagatttcgacgacccgggccggcgtgtac2400gtcgccaactcgacgaccgacagcttcatcaatggcgtcagggacggtctgcggacgttc2460aagttcctggaccggttcgcgattgacgaggatgctgagtga2502<210>8<211>740<212>prt<213>橙色嗜热子囊菌<400>8metargalaileglyleuleuproglyileileglyilealaglyala151015alacysprotyrmetthrglygluleuproargserphealagluasn202530prohisalaileasnargargalagluglyglyglyglyalaalaala354045gluthrglulyspheleuserglnphetyrleuasnaspasnaspthr505560phemetthrthraspvalglyglyproilegluaspglnasnserleu65707580seralaglyaspargglyprothrleuleugluasppheileleuarg859095glnlysileglnargpheasphisgluargvalprogluargalaval100105110hisalaargglyalaglyalahisglyvalphethrsertyralaasp115120125trpserasnilethralaalaserpheleuseralaalaglylysglu130135140thrprovalphevalargpheserthrvalalaglyserargglyser145150155160alaaspthralaargaspvalhisglyphealathrargphetyrthr165170175aspgluglyasnpheaspilevalglyasnasnileprovalphephe180185190ileglnaspalaileglnpheproaspleuilehisalavallyspro195200205serproasnasngluileproglnalaalathralahisaspserala210215220trpaspphepheserglnglnproserserleuhisthrleuphetrp225230235240alametalaglyhisglyileproargsertyrargasnmetaspgly245250255pheglyilehisthrpheargphevalthraspaspglyalaserlys260265270leuvallysphehistrpthrserleuglnglylysalaserleuval275280285trpgluglualaglnalavalalaglylysasnalaasptyrhisarg290295300glnaspleutrpaspalaileglualaglyargtyrproglutrpglu305310315320leuglyvalglnilemetaspglugluaspglnleuargpheglyphe325330335aspleuleuaspprothrlysilevalprogluglutyrvalproile340345350thrlysleuglylysmetglnleuasnargasnproleuasntyrphe355360365alagluthrgluglnilemetpheglnproglyhisvalvalarggly370375380ileaspphethrgluaspproleuleuglnglyargleuphesertyr385390395400leuaspthrglnleuasnarghisglyglyproasnphegluglnile405410415proileasnargproargthrproilehisasnasnasnargaspgly420425430alaalaglnmettyrileproleuasnlysalaalatyrthrproasn435440445thrleuasnasnglyserprolysglnalaasnglnthrvalglylys450455460glyphephethrthrproglyargthralaserglyargleuvalarg465470475480alavalserserthrphealaaspvaltrpserglnproargleuphe485490495tyrasnserleuvalproalagluglnglnpheleuileasnalaile500505510argphegluthralahisilethrseraspvalvallysasnasnval515520525ileileglnleuasnargvalserasnasnleualalysargvalala530535540argalaileglyvalalagluprogluproaspprothrleutyrhis545550555560asnasnlysthralaasnvalglyvalpheglylysproleualaarg565570575leuaspglyleuglnvalglyvalleualathrvalasnlysproasp580585590serilelysglnalaalaserleulysalaserphealaalaaspasn595600605valaspvallysvalvalalagluargleualaaspglyvalaspglu610615620thrtyrseralaalaaspalavalasnpheaspalaileleuvalala625630635640asnglyalagluglyleuphealaargaspserphethralaargpro645650655alaasnserthrthralathrleutyrproalaglyargproleugln660665670ileleuvalaspglypheargtyrglylysprovalglyalaleugly675680685serglyalalysalaleuaspalaalagluileserthrthrargala690695700glyvaltyrvalalaasnserthrthraspserpheileasnglyval705710715720argaspglyleuargthrphelyspheleuaspargphealaileasp725730735gluaspalaglu740<210>9<211>3060<212>dna<213>烟曲霉<400>9atgagattcggttggctcgaggtggccgctctgacggccgcttctgtagccaatgcccag60gtttgtgatgctttcccgtcattgtttcggatatagttgacaatagtcatggaaataatc120aggaattggctttctctccaccattctacccttcgccttgggctgatggccagggagagt180gggcagatgcccatcgacgcgccgtcgagatcgtttctcagatgacactggcggagaagg240ttaaccttacaacgggtactgggtgggttgcgacttttttgttgacagtgagctttcttc300actgaccatctacacagatgggaaatggaccgatgcgtcggtcaaaccggcagcgttccc360aggtaagcttgcaattctgcaacaacgtgcaagtgtagttgctaaaacgcggtggtgcag420acttggtatcaactggggtctttgtggccaggattcccctttgggtatccgtgactgtga480gctatacccgcggagtctttcagtccttgtattatgtgctgatgattgtctctgtatagc540tgacctcaactccgccttccctgctggtactaatgtcgccgcgacatgggacaagacact600cgcctaccttcgtggcaaggccatgggtgaggaattcaacgacaagggcgtggacatttt660gctggggcctgctgctggtcctctcggcaaatacccggacggcggcagaatctgggaagg720cttctctcctgatccggttctcactggtgtacttttcgccgaaactatcaagggtatcca780agacgcgggtgtgattgctactgccaagcattacattctgaatgaacaggagcatttccg840acaggttggcgaggcccagggatatggttacaacatcacggagacgatcagctccaacgt900ggatgacaagaccatgcacgagttgtacctttggtgagtagttgacactgcaaatgagga960ccttgattgatttgactgacctggaatgcaggccctttgcagatgctgtgcgcggtaaga1020ttttccgtagacttgacctcgcgacgaagaaatcgctgacgaaccatcgtagctggcgtt1080ggcgctgtcatgtgttcctacaatcaaatcaacaacagctacggttgtcaaaacagtcaa1140actctcaacaagctcctcaaggctgagctgggcttccaaggcttcgtcatgagtgactgg1200ggcgctcaccacagcggtgtcggcgctgccctcgctgggttggatatgtcgatgcctgga1260gacatttccttcgacgacggactctccttctggggcacgaacctaactgtcagtgttctt1320aacggcaccgttccagcctggcgtgtcgatgacatggctgttcgtatcatgaccgcgtac1380tacaaggttggtcgtgaccgtcttcgtattccccctaacttcagctcctggacccgggat1440gagtacggctgggagcattctgctgtctccgagggagcctggaccaaggtgaacgacttc1500gtcaatgtgcagcgcagtcactctcagatcatccgtgagattggtgccgctagtacagtg1560ctcttgaagaacacgggtgctcttcctttgaccggcaaggaggttaaagtgggtgttctc1620ggtgaagacgctggttccaacccgtggggtgctaacggctgccccgaccgcggctgtgat1680aacggcactcttgctatggcctggggtagtggtactgccgagttcccttaccttgtcacc1740cccgagcaggctatccagcgagaggtcatcagcaacggcggcaatgtctttgctgtgact1800gataacggggctctcagccagatggcagatgttgcatctcaatccaggtgagtgcgggct1860cttagaaaaagaacgttctctgaatgaagttttttaaccattgcgaacagcgtgtctttg1920gtgtttgtcaacgccgactctggagagggttacatcagtgtcgacggcaacgagggtgac1980cgcaaaaatctcactctgtggaagaacggcgaggccgtcattgacactgttgtcagccac2040tgcaacaacacgattgtggttattcacagtgttgggcccgtcttgatcgaccggtggtat2100gataaccccaacgtcactgccatcatctgggccggcttgcccggtcaggagagtggcaac2160tccctggtcgacgtgctctatggccgcgtcaaccccagcgccaagaccccgttcacctgg2220ggcaagactcgggagtcttacggggctcccttgctcaccgagcctaacaatggcaatggt2280gctccccaggatgatttcaacgagggcgtcttcattgactaccgtcactttgacaagcgc2340aatgagacccccatttatgagtttggccatggcttgagctacaccacctttggttactct2400caccttcgggttcaggccctcaatagttcgagttcggcatatgtcccgactagcggagag2460accaagcctgcgccaacctatggtgagatcggtagtgccgccgactacctgtatcccgag2520ggtctcaaaagaattaccaagtttatttacccttggctcaactcgaccgacctcgaggat2580tcttctgacgacccgaactacggctgggaggactcggagtacattcccgaaggcgctagg2640gatgggtctcctcaacccctcctgaaggctggcggcgctcctggtggtaaccctaccctt2700tatcaggatcttgttagggtgtcggccaccataaccaacactggtaacgtcgccggttat2760gaagtccctcaattggtgagtgacccgcatgttccttgcgttgcaatttggctaactcgc2820ttctagtatgtttcactgggcggaccgaacgagcctcgggtcgttctgcgcaagttcgac2880cgaatcttcctggctcctggggagcaaaaggtttggaccacgactcttaaccgtcgtgat2940ctcgccaattgggatgtggaggctcaggactgggtcatcacaaagtaccccaagaaagtg3000cacgtcggcagctcctcgcgtaagctgcctctgagagcgcctctgccccgtgtctactag3060<210>10<211>863<212>prt<213>烟曲霉<400>10metargpheglytrpleugluvalalaalaleuthralaalaserval151015alaasnalaglngluleualapheserproprophetyrproserpro202530trpalaaspglyglnglyglutrpalaaspalahisargargalaval354045gluilevalserglnmetthrleualaglulysvalasnleuthrthr505560glythrglytrpglumetaspargcysvalglyglnthrglyserval65707580proargleuglyileasntrpglyleucysglyglnaspserproleu859095glyileargaspseraspleuasnseralapheproalaglythrasn100105110valalaalathrtrpasplysthrleualatyrleuargglylysala115120125metglygluglupheasnasplysglyvalaspileleuleuglypro130135140alaalaglyproleuglylystyrproaspglyglyargiletrpglu145150155160glypheserproaspprovalleuthrglyvalleuphealagluthr165170175ilelysglyileglnaspalaglyvalilealathralalyshistyr180185190ileleuasngluglngluhispheargglnvalglyglualaglngly195200205tyrglytyrasnilethrgluthrileserserasnvalaspasplys210215220thrmethisgluleutyrleutrpprophealaaspalavalargala225230235240glyvalglyalavalmetcyssertyrasnglnileasnasnsertyr245250255glycysglnasnserglnthrleuasnlysleuleulysalagluleu260265270glypheglnglyphevalmetserasptrpglyalahishissergly275280285valglyalaalaleualaglyleuaspmetsermetproglyaspile290295300serpheaspaspglyleuserphetrpglythrasnleuthrvalser305310315320valleuasnglythrvalproalatrpargvalaspaspmetalaval325330335argilemetthralatyrtyrlysvalglyargaspargleuargile340345350proproasnphesersertrpthrargaspglutyrglytrpgluhis355360365seralavalsergluglyalatrpthrlysvalasnaspphevalasn370375380valglnargserhisserglnileilearggluileglyalaalaser385390395400thrvalleuleulysasnthrglyalaleuproleuthrglylysglu405410415vallysvalglyvalleuglygluaspalaglyserasnprotrpgly420425430alaasnglycysproaspargglycysaspasnglythrleualamet435440445alatrpglyserglythralaglupheprotyrleuvalthrproglu450455460glnalaileglnarggluvalileserasnglyglyasnvalpheala465470475480valthraspasnglyalaleuserglnmetalaaspvalalasergln485490495serservalserleuvalphevalasnalaaspserglygluglytyr500505510ileservalaspglyasngluglyasparglysasnleuthrleutrp515520525lysasnglyglualavalileaspthrvalvalserhiscysasnasn530535540thrilevalvalilehisservalglyprovalleuileaspargtrp545550555560tyraspasnproasnvalthralaileiletrpalaglyleuprogly565570575glngluserglyasnserleuvalaspvalleutyrglyargvalasn580585590proseralalysthrprophethrtrpglylysthrargglusertyr595600605glyalaproleuleuthrgluproasnasnglyasnglyalaprogln610615620aspasppheasngluglyvalpheileasptyrarghispheasplys625630635640argasngluthrproiletyrglupheglyhisglyleusertyrthr645650655thrpheglytyrserhisleuargvalglnalaleuasnserserser660665670seralatyrvalprothrserglygluthrlysproalaprothrtyr675680685glygluileglyseralaalaasptyrleutyrprogluglyleulys690695700argilethrlyspheiletyrprotrpleuasnserthraspleuglu705710715720aspserseraspaspproasntyrglytrpgluaspserglutyrile725730735progluglyalaargaspglyserproglnproleuleulysalagly740745750glyalaproglyglyasnprothrleutyrglnaspleuvalargval755760765seralathrilethrasnthrglyasnvalalaglytyrgluvalpro770775780glnleutyrvalserleuglyglyproasngluproargvalvalleu785790795800arglyspheaspargilepheleualaproglygluglnlysvaltrp805810815thrthrthrleuasnargargaspleualaasntrpaspvalgluala820825830glnasptrpvalilethrlystyrprolyslysvalhisvalglyser835840845serserarglysleuproleuargalaproleuproargvaltyr850855860<210>11<211>1520<212>dna<213>雷塞特纳斯篮状菌<400>11atggtccatctttcttccctggccctggctttggccgccggctcgcagctgtatgtgatc60catgccatgactcgagaagtgctcccaaaactgactccaagtctcaatcttagtgcccaa120gctgcaggtcttaacactgctgccaaagcgattggaaagctctatttcggtaccgcaacc180gacaacccggagctgtccgacagcacatacatgcaggagacggataacaccgatgatttc240ggccaactcaccccagctaactccatgaaggttcgctgacatcttagttccccccccctt300ttgggaatctgcgcggagatatgctgagccttcaaaactagtgggatgccaccgagccct360ctcagaacaccttcaccttcaccaacggtgatcagatcgcaaaccttgctaagagcaacg420gtcagatgctgagatgccacaacctggtgtggtacaaccagttgcccagctggggtaagc480aaccggttctgttaatatcatcagcgtgaccgcatcgatcgtattgcgcggagattggaa540agatttgcaagctaatgtcactacagtcaccagcggatcttggaccaatgccacgcttct600tgcggccatgaagaaccacatcaccaacgttgtgacccactacaagggacagtgctacgc660ttgggatgttgtcaacgaaggtacgtttcgattcggcttccctcggaccgtatctgcagg720caaaaaggtcaatcaattgacaatcgtgatccccagctctcaacgatgatggcacctacc780gatccaatgtcttctatcagtacatcggcgaggcatacattcccattgcctttgcgaccg840ctgccgccgccgatccaaacgcgaagctctactacaacgactacaacattgagtaccccg900gcgccaaggccaccgccgcccagaacatcgtcaagatggtcaaggcttacggcgcgaaaa960tcgacggtgtcggtctgcaatctcacttcatcgttggcagcacccctagccagagctccc1020agcagagcaacatggctgctttcaccgcgctcggcgtcgaggtcgccatcaccgaactgg1080atatccgcatgacgttgccttccaccagtgctctcttggcccagcaatccaccgattacc1140agagcactgtgtcggcttgcgtgaacactccgaagtgcattggtatcaccctctgggact1200ggaccgacaagtactcctgggttcccaacaccttctccggccaaggtgacgcctgcccct1260gggattctaactaccagaagaagcctgcctactacggtatcttgactgcgctcggaggca1320gcgcttccacctccaccaccaccactctggtgacctccaccaggacttcgactacgacca1380gcacttcggccacctccacgtctactggcgttgctcagcactggggccagtgcggtggta1440tcggctggacagggccgactacctgcgctagcccctacacctgccaggaactgaatccct1500actactaccagtgcctgtaa1520<210>12<211>405<212>prt<213>雷塞特纳斯篮状菌<400>12metvalhisleuserserleualaleualaleualaalaglysergln151015leualaglnalaalaglyleuasnthralaalalysalaileglylys202530leutyrpheglythralathraspasnprogluleuseraspserthr354045tyrmetglngluthraspasnthraspasppheglyglnleuthrpro505560alaasnsermetlystrpaspalathrgluproserglnasnthrphe65707580thrphethrasnglyaspglnilealaasnleualalysserasngly859095glnmetleuargcyshisasnleuvaltrptyrasnglnleuproser100105110trpvalthrserglysertrpthrasnalathrleuleualaalamet115120125lysasnhisilethrasnvalvalthrhistyrlysglyglncystyr130135140alatrpaspvalvalasnglualaleuasnaspaspglythrtyrarg145150155160serasnvalphetyrglntyrileglyglualatyrileproileala165170175phealathralaalaalaalaaspproasnalalysleutyrtyrasn180185190asptyrasnileglutyrproglyalalysalathralaalaglnasn195200205ilevallysmetvallysalatyrglyalalysileaspglyvalgly210215220leuglnserhispheilevalglyserthrproserglnsersergln225230235240glnserasnmetalaalaphethralaleuglyvalgluvalalaile245250255thrgluleuaspileargmetthrleuproserthrseralaleuleu260265270alaglnglnserthrasptyrglnserthrvalseralacysvalasn275280285thrprolyscysileglyilethrleutrpasptrpthrasplystyr290295300sertrpvalproasnthrpheserglyglnglyaspalacysprotrp305310315320aspserasntyrglnlyslysproalatyrtyrglyileleuthrala325330335leuglyglyseralaserthrserthrthrthrthrleuvalthrser340345350thrargthrserthrthrthrserthrseralathrserthrserthr355360365glyvalalaglnhistrpglyglncysglyglyileglytrpthrgly370375380prothrthrcysalaserprotyrthrcysglngluleuasnprotyr385390395400tyrtyrglncysleu405<210>13<211>2391<212>dna<213>埃默森篮状菌<400>13atgatgactcccacggcgattctcaccgcagtggcggcgctcctgcccaccgcgacatgg60gcacaggataaccaaacctatgccaattactcgtcgcagtctcagccggacctgtttccc120cggaccgtcgcgaccatcgacctgtccttccccgactgtgagaatggcccgctcagcacg180aacctggtgtgcaacaaatcggccgatccctgggcccgagctgaggccctcatctcgctc240tttaccctcgaagagctgattaacaacacccagaacaccgctcctggcgtgccccgtttg300ggtctgccccagtatcaggtgtggaatgaagctctgcacggactggaccgcgccaatttc360tcccattcgggcgaatacagctgggccacgtccttccccatgcccatcctgtcgatggcg420tccttcaaccggaccctcatcaaccagattgcctccatcattgcaacgcaagcccgtgcc480ttcaacaacgccggccgttacggccttgacagctatgcgcccaacatcaatggcttccgc540agtcccctctggggccgtggacaggagacgcctggtgaggatgcgttcttcttgagttcc600acctatgcgtacgagtacatcacaggcctgcagggcggtgtcgacccagagcatgtcaag660atcgtcgcgacggcgaagcacttcgccggctatgatctggagaactggggcaacgtctct720cggctggggttcaatgctatcatcacgcagcaggatctctccgagtactacacccctcag780ttcctggcgtctgctcgatacgccaagacgcgcagcatcatgtgctcctacaatgcagtg840aatggagtcccaagctgtgccaactccttcttcctccagacgcttctccgagaaaacttt900gacttcgttgacgacgggtacgtctcgtcggattgcgacgccgtctacaacgtcttcaac960ccacacggttacgcccttaaccagtcgggagccgctgcggactcgctcctagcaggtacc1020gatatcgactgtggtcagaccttgccgtggcacctgaatgagtccttcgtagaaggatac1080gtctcccgcggtgatatcgagaaatccctcacccgtctctactcaaacctggtgcgtctc1140ggctactttgacggcaacaacagcgagtaccgcaacctcaactggaacgacgtcgtgact1200acggacgcctggaacatctcgtacgaggccgcggtggaaggtatcaccctgctcaagaac1260gacggaacgctgccgctgtccaagaaggtccgcagcattgcgctcatcggtccttgggcc1320aatgccacggtgcagatgcagggtaactactatggaacgccaccgtatctgatcagtccg1380ctggaagccgccaaggccagtgggttcacggtcaactatgcattcggtaccaacatctcg1440accgattctacccagtggttcgcggaagccatcgcggcggcgaagaagtcggacgtgatc1500atctacgccggtggtattgacaacacgatcgaggcagagggacaggaccgcacggatctc1560aagtggccggggaaccagctggatctgatcgagcagctcagccaggtgggcaagcccttg1620gtcgtcctgcagatgggcggtggccaggtggattcgtcgtcactcaaggccaacaagaat1680gtcaacgctctggtgtggggtggctatcccggacagtcgggtggtgcggccctgtttgac1740atccttacgggcaagcgtgcgccggccggtcgtctggtgagcacgcagtacccggccgag1800tatgcgacgcagttcccggccaacgacatgaacctgcgtccgaacggcagcaacccggga1860cagacatacatctggtacacgggcacgcccgtgtatgagttcggccacggtctgttctac1920acggagttccaggagtcggctgcggcgggcacgaacaagacgtcgactttcgacattctg1980gaccttttctccacccctcatccgggatacgagtacatcgagcaggttccgttcatcaac2040gtgactgtggacgtgaagaacgtcggccacacgccatcgccgtacacgggtctgttgttc2100gcgaacacgacagccgggcccaagccgtacccgaacaaatggctcgtcgggttcgactgg2160ctgccgacgatccagccgggcgagactgccaagttgacgatcccggtgccgttgggcgcg2220attgcgtgggcggacgagaacggcaacaaggtggtcttcccgggcaactacgaattggca2280ctgaacaatgagcgatcggtagtggtgtcgttcacgctgacgggcgatgcggcgactcta2340gagaaatggcctttgtgggagcaggcggttccgggggtgctgcagcaataa2391<210>14<211>796<212>prt<213>埃默森篮状菌<400>14metmetthrprothralaileleuthralavalalaalaleuleupro151015thralathrtrpalaglnaspasnglnthrtyralaasntyrserser202530glnserglnproaspleupheproargthrvalalathrileaspleu354045serpheproaspcysgluasnglyproleuserthrasnleuvalcys505560asnlysseralaaspprotrpalaargalaglualaleuileserleu65707580phethrleuglugluleuileasnasnthrglnasnthralaprogly859095valproargleuglyleuproglntyrglnvaltrpasnglualaleu100105110hisglyleuaspargalaasnpheserhisserglyglutyrsertrp115120125alathrserpheprometproileleusermetalaserpheasnarg130135140thrleuileasnglnilealaserileilealathrglnalaargala145150155160pheasnasnalaglyargtyrglyleuaspsertyralaproasnile165170175asnglypheargserproleutrpglyargglyglngluthrprogly180185190gluaspalaphepheleuserserthrtyralatyrglutyrilethr195200205glyleuglnglyglyvalaspprogluhisvallysilevalalathr210215220alalyshisphealaglytyraspleugluasntrpglyasnvalser225230235240argleuglypheasnalaileilethrglnglnaspleuserglutyr245250255tyrthrproglnpheleualaseralaargtyralalysthrargser260265270ilemetcyssertyrasnalavalasnglyvalprosercysalaasn275280285serphepheleuglnthrleuleuarggluasnpheaspphevalasp290295300aspglytyrvalserseraspcysaspalavaltyrasnvalpheasn305310315320prohisglytyralaleuasnglnserglyalaalaalaaspserleu325330335leualaglythraspileaspcysglyglnthrleuprotrphisleu340345350asngluserphevalgluglytyrvalserargglyaspileglulys355360365serleuthrargleutyrserasnleuvalargleuglytyrpheasp370375380glyasnasnserglutyrargasnleuasntrpasnaspvalvalthr385390395400thraspalatrpasnilesertyrglualaalavalgluglyilethr405410415leuleulysasnaspglythrleuproleuserlyslysvalargser420425430ilealaleuileglyprotrpalaasnalathrvalglnmetglngly435440445asntyrtyrglythrproprotyrleuileserproleuglualaala450455460lysalaserglyphethrvalasntyralapheglythrasnileser465470475480thraspserthrglntrpphealaglualailealaalaalalyslys485490495seraspvalileiletyralaglyglyileaspasnthrilegluala500505510gluglyglnaspargthraspleulystrpproglyasnglnleuasp515520525leuilegluglnleuserglnvalglylysproleuvalvalleugln530535540metglyglyglyglnvalaspserserserleulysalaasnlysasn545550555560valasnalaleuvaltrpglyglytyrproglyglnserglyglyala565570575alaleupheaspileleuthrglylysargalaproalaglyargleu580585590valserthrglntyrproalaglutyralathrglnpheproalaasn595600605aspmetasnleuargproasnglyserasnproglyglnthrtyrile610615620trptyrthrglythrprovaltyrglupheglyhisglyleuphetyr625630635640thrglupheglngluseralaalaalaglythrasnlysthrserthr645650655pheaspileleuaspleupheserthrprohisproglytyrglutyr660665670ilegluglnvalpropheileasnvalthrvalaspvallysasnval675680685glyhisthrproserprotyrthrglyleuleuphealaasnthrthr690695700alaglyprolysprotyrproasnlystrpleuvalglypheasptrp705710715720leuprothrileglnproglygluthralalysleuthrileproval725730735proleuglyalailealatrpalaaspgluasnglyasnlysvalval740745750pheproglyasntyrgluleualaleuasnasngluargservalval755760765valserphethrleuthrglyaspalaalathrleuglulystrppro770775780leutrpgluglnalavalproglyvalleuglngln785790795当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1