用于延长蛋白质半衰期的方法与流程
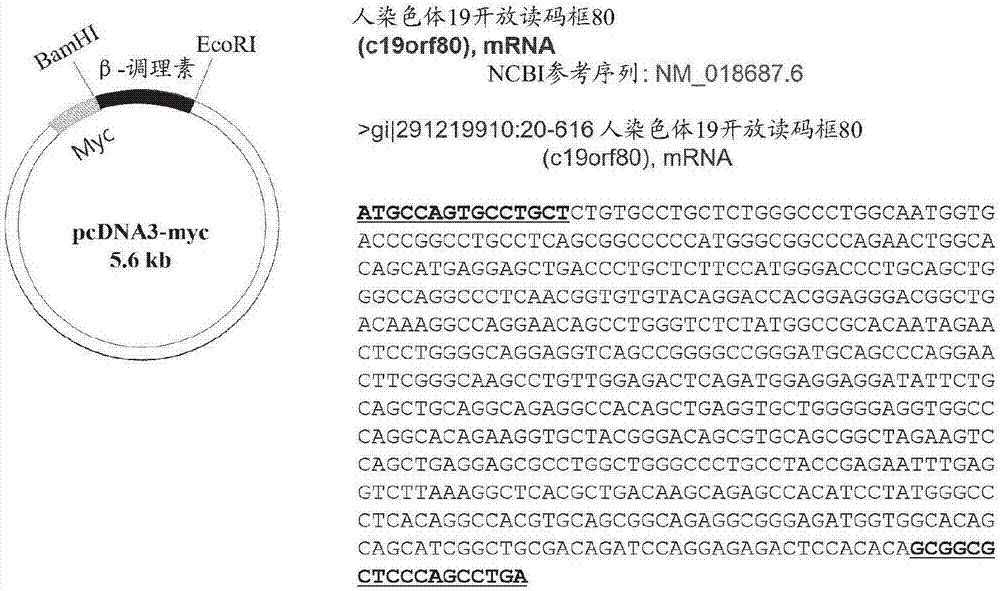
本发明涉及一种通过替换蛋白质的与泛素化相关的一个或更多个赖氨酸残基来延长蛋白质或(多)肽半衰期的方法,且涉及具有延长的半衰期的蛋白质。
背景技术:
:真核细胞中的蛋白质或(多)肽通过溶酶体系统和泛素-蛋白酶体系统两种不同途径被降解。分解10-20%的细胞蛋白的溶酶体系统,即没有底物特异性也没有精密时序可控性。也就是说,溶酶体系统是一种尤其分解大多数细胞外蛋白或膜蛋白的过程,例如表面蛋白通过内吞作用被细胞摄取,再被溶酶体降解。至于真核细胞中蛋白质的选择性降解,应涉及泛素-蛋白酶体途径(UPP),在这之中,目标蛋白首先结合泛素结合酶以形成多聚泛素链,再被蛋白酶体识别并分解。大约80-90%的真核细胞蛋白是通过UPP降解的,因此,UPP被认为调控真核细胞内大部分蛋白的降解,且负责体内的蛋白质转换和体内稳态。泛素是一个由76个高度保守的氨基酸组成的小蛋白,且它存在于所有真核细胞中。在泛素的所有氨基酸残基中,第6、11、27、29、33、48和63位残基是赖氨酸(Lysine,Lys,K),且已知第48位和63位的残基对多聚泛素链的形成起关键作用。通常称为E1、E2、E3的三种酶连续作用以促进泛素化,而被泛素标记的蛋白则被ATP依赖的蛋白质降解复合物中的26S蛋白酶体分解。如上所述,泛素蛋白酶体途径(UPP)由两个分离且连续的过程组成。一个过程是一定数目的泛素分子连接至底物蛋白的蛋白标记过程,而另外一个则是经标记的蛋白被26S蛋白酶体复合物降解的过程。泛素和底物蛋白之间的结合是通过泛素C末端的甘氨酸残基和底物的赖氨酸残基之间形成异肽键,然后再通过一系列酶(泛素激活酶E1、泛素结合酶E2、泛素连接酶E3)在泛素和底物蛋白之间形成硫脂键而实施的。已知E1(泛素激活酶)通过ATP依赖的反应机制激活泛素。被激活的泛素被转移至E2(泛素结合酶)的泛素结合域中的半胱氨酸残基上,然后E2运送被激活的的泛素到E3连接酶处或直接运送至底物蛋白处。E3同样催化底物蛋白的赖氨酸残基和泛素的甘氨酸残基之间形成异肽键。另外一个泛素能够被连接至与底物蛋白相连的泛素的C末端的赖氨酸残基上,额外的泛素部分的重复连接通过一定数目的泛素分子彼此相连而产生多聚泛素链。当多聚泛素链被生成之后,底物蛋白则被26S蛋白酶体选择性地识别并降解。与此同时,生物体内有着多种具有治疗效果的蛋白质。这些在体内具有治疗效果的蛋白质或(多)肽或生物活性多肽包括,但不限于,例如,生长激素释放激素(GHRH)、生长激素释放肽、干扰素(干扰素-α或干扰素-β)、干扰素受体、集落刺激因子(CSF)、胰高血糖素样肽、白介素、白介素受体、酵素、白介素结合蛋白、细胞因子结合蛋白、G蛋白偶联受体、人生长激素(hGH)、巨噬细胞活化因子、巨噬细胞肽、B细胞因子、T细胞因子、蛋白A、过敏抑制剂、细胞坏死糖蛋白、G蛋白偶联受体、免疫毒素、淋巴毒素、肿瘤坏死因子、肿瘤抑制因子、转移生长因子、α-1抗胰蛋白酶、白蛋白、α-乳白蛋白、载脂蛋白E、红细胞生成素、高度糖基化红细胞生成素、血管生成素、血红蛋白、凝血酶、凝血酶受体激活肽、血栓调节蛋白、第七因子、第七因子a、第八因子、第九因子、第十三因子、纤维蛋白溶酶原激活因子、尿激酶、链激酶、水蛭素、蛋白C、C反应蛋白、肾素抑制剂、胶原蛋白酶抑制剂、过氧化物歧化酶、瘦蛋白、血小板源生长因子、上皮生长因子、表皮生长因子、血管抑素、血管紧张肽、骨生长因子、骨刺激蛋白、降血钙素、胰岛素、心钠素、软骨诱导因子、纤维蛋白结合肽、依降钙素、结缔组织激活因子、组织因子通道抑制剂、促卵泡激素、促黄体激素、促黄体激素释放激素、神经生长因子、甲状旁腺素、松弛肽、分泌素、生长调节素、胰岛素样生长因子、肾上腺皮质激素、胰高血糖素、肠促胰酶肽、胰多肽、胃泌素释放肽、促皮质素释放因子、促甲状腺激素、自毒素、乳铁蛋白、肌生成抑制蛋白、受体、受体拮抗体、细胞表面抗原、病毒衍生的疫苗抗原、单克隆抗体、多克隆抗体和抗体片段。已知β-调理素促进分泌胰岛素的胰腺β细胞的增殖。因此,β-调理素能够以一年一次或两次施用于二型糖尿病患者,来维持胰腺β细胞的活性而控制血糖水平。施用β-调理素比施用胰岛素的副作用更小,这是因为被给予β-调理素治疗的患者能够为他们自己生产胰岛素。进一步地,已报道,在小鼠肝脏内β-调理素的暂时性表达促进胰腺β细胞增殖(Cell153,747758,2013)。生长激素(GH),一种多肽荷尔蒙,在脑下垂体的前叶被合成和分泌,且它不仅与骨和软骨的生长有关,还与脂肪的分解和蛋白的合成的刺激这一新陈代谢相关。因此,生长激素能被用于治疗侏儒症,该侏儒症由多种身体情况(包括,例如,先天性心脏病、慢性肺病、慢性肾病或者慢性消耗性疾病;由生长激素缺乏、甲状腺功能减退或糖尿病引起的激素异常分泌;和诸如特纳综合征的先天遗传性疾病)引起。进一步地,已知生长激素调控STAT蛋白的转录(信号转导和转录激活因子)(Oncogene,19,2585-2597,2000)。已知胰岛素在人体内调节血糖的水平。因此,胰岛素能被用于治疗由于胰腺的胰岛细胞的功能被削弱而遭受血糖水平上升的一型糖尿病患者。另外,胰岛素能被用于虽然胰岛素仍然能正常地被分泌,但由于体细胞的胰岛素受体抵抗而不能控制血糖的二型糖尿病患者。根据之前的研究,有报道称胰岛素在肝脏内刺激STAT磷酸化,从而控制肝脏内的葡萄糖稳态(CellMetabolism3,267275,2006)。干扰素是通过免疫系统的细胞产生和分泌的一群天然产生的蛋白质,包括白细胞、自然杀伤细胞、纤维细胞和上皮细胞。干扰素被分为3类,例如一型、二型和三型,所谓的分类是按照被相应蛋白运送至的受体区分的。尽管干扰素的功能机制复杂还并没有被完全理解,但是已知它调节免疫系统对病毒、癌症和一些其他外部(或传染性)物质的应答。与此同时,已知干扰素不会直接杀死病毒或者肿瘤细胞,但是会促进免疫系统应答,并控制调节多种细胞的蛋白质的分泌的基因的功能,从而起到抑制肿瘤细胞生长的功能。至于一型干扰素,已知干扰素-α能被用于治疗乙肝和丙肝,干扰素-β可以被用于治疗多发性硬化。进一步地,相关研究报道干扰素-α可以增强STAT-1,STAT-2和STAT-3(JImmunol,187,2578-2585,2011),以及它激活黑色素瘤细胞中的STAT3蛋白,有助于黑色素瘤的生成(EuroJCancer,45,1315-1323,2009)。进一步地,据报道,被干扰素-β处理过的细胞诱导包括AKT的信号通路的激活(Pharmaceuticals(Basel),3,994-1015,2010)。粒细胞集落刺激因子(G-CSF),一种糖蛋白,其产生骨髓干细胞和粒细胞,且刺激骨髓分泌骨髓干细胞和粒细胞到血管中。G-CSF是一种集落刺激因子,并且也行使激素和细胞因子的功能。进一步地,C-CSF还通过增加神经可塑性和抑制细胞凋亡而作为一种神经营养因子,除此之外还对造血作用产生影响。G-CSF受体在脑和脊柱中的神经元中被表达。在中枢神经系统中,G-CSF诱导神经元的生成和增加神经可塑性,从而与细胞凋亡相关。因此,G-CSF被研究用于治疗神经元疾病,例如脑梗死。G-CSF可以刺激粒细胞的生成,其是白细胞的一种。进一步地,重组G-CSF可以被用于加快在肿瘤学和血液学中的化学治疗引起的嗜中性白细胞减少症的恢复。研究报道称G-CSF激活神经胶质瘤细胞中的STAT3从而参与胶质瘤的生长(CancerBiolTher.,13(6),389-400,2012)。进一步地,据报道,G-CSF在卵巢上皮癌细胞中被表达,且通过调节JAK2/STAT3通路与女性子宫癌病理性相关(BrJCancer,110,133-145,2014)。红细胞生成素(EPO),一种糖蛋白激素,其与各种生长因子例如白细胞介素-3,白细胞介素-6,糖皮质激素和干细胞因子等)相互作用(。作为一种细胞因子,红细胞生成素存在于骨髓中作为一种红细胞的前体物质且与红细胞的生成相关。除此之外,红细胞生成素还与由于抑制铁调节激素中的铁调素的吸收从而上调铁离子的吸收所引起的血管收缩依赖性高血压相关。进一步地,红细胞生成素在脑部应答神经元损伤中的脑神经原的保护具有重要作用,例如心肌梗死或中风。另外,红细胞生成素被认为对记忆改善,疤痕恢复和抑郁症有治疗效果。进一步地,已报道在肺癌和血癌病人中,红细胞生成素的水平增加。进一步地,有研究报道,EPO通过Erk1/2磷酸化而调节细胞周期进程,因此对低氧有作用被报道(JHematolOncol.,6,65,2013)。纤维母细胞生长因子-1(FGF-1)是纤维母细胞生长因子的一种,其与胚胎发育、细胞生长、组织再生和癌症发展和转变相关。进一步地,据报道FGF-1在临床研究中诱导心血管的血管生成(BioDrugs.,11(5),301308,1999)。由于FGF-1可以促进细胞生长,因此它帮助维持表皮健康,从而增加皮肤弹性以使皮肤湿润。进一步地,FGF-1激活皮肤细胞,使皮肤外表亮泽,提供牛奶般的肌肤。另外,已知FGF-1帮助皮肤从受损或创伤中快速复原,并通过加强皮肤屏障以增强保护功能。进一步地,已知重组纤维母细胞生长因子-1(FGF-1)增强HEK293细胞的Erk1/2磷酸化(Nature,513(7518),436-439,2014)。血管内皮生长因子A(VEGFA)是一种在刺激血管新生和血管再生的细胞中产生的信号转导蛋白,且在低氧环境中,它还在组织中储存氧气(MolCellEndocrinol.,397,5157,2014)。在哮喘和糖尿病中,检测到血清中VEGF水平的升高(Diabetes,48(11),22292239,2013)。VEGF在胚胎发育、损伤后新血管的生成以及运动后渗透至肌肉和堵塞的血管的新血管生成中行使功能。与此同时,VEGF的过表达会导致疾病或紊乱。例如,如果血液流入被阻塞,实体肿瘤不会进一步生长,但是当VEGF表达时,肿瘤会持续性的生长并发生转移。进一步地,VEGF被认为是内皮细胞生长和增殖的重要因素且参与肿瘤细胞中的血管生成的发生。特别地,据报道在癌细胞中,P13K/Akt/HIF-1α信号转导途径通过肿瘤细胞中的VEGF与血管生成的发生有关(Carcinogenesis,34,426-435,2013)。进一步地,已知VEGF诱导AKT磷酸化(KidneyInt.,68,1648-1659,2005)。食欲抑制蛋白(瘦蛋白)和食欲刺激激素(胃饥饿素)在脂肪组织被分泌。瘦蛋白是一种循环激素(16kDa)(CellRes.,10,81-92,2000)并且在免疫、繁殖和造血中有着重要作用。胃饥饿素通过生长激素促分泌素受体(GHS-R)从脂肪组织中分泌并刺激食欲,是一种由28个氨基酸组成的胃肽,(JEndocrinol.,192,313323,2007;Nature,442,656-660,1999),且由前胃饥饿素原形成(PediatrRes.,65,3944,2009;JBiolChem.,281(50),3886738870,2006)。瘦蛋白是一种提供不能再摄取食物的饱腹信号的激素,并且瘦蛋白激素分泌的削弱被认为能刺激食欲。据报道,果糖在促进胃饥饿素的分泌而增加食欲的同时,干扰胰岛素的分泌和减少瘦蛋白的分泌(JBiolChem.,277(7),5667-5674,2002;I.J.S.N.,7(1),06-15,2016)。进一步地,食欲抑制蛋白被报道在乳腺癌细胞中增加AKT的磷酸化(CancerBiolTher.,16(8),1220-1230,2015),并且在子宫癌中通过PI3K/AKT信号转导刺激癌细胞生长(IntJOncol.,49(2),847,2016)。进一步地,已知瘦蛋白在子宫癌中通过PI3K/AKT信号转导通路刺激癌细胞生长(IntJOncol.,49(2),847,2016)。已知食欲刺激激素(胃饥饿素)通过生长激素促分泌素受体(GHS-R)而调节细胞生长,且在体内通过钙调节地方式而增强STAT3(MolCellEndocrinol.,285,19-25,2008)。胰高血糖素样肽-1(GLP-1),一种肠促胰岛素激素,由回肠和大肠的L细胞分泌,依赖葡萄糖浓度来增加胰岛素分泌,从而防止血糖过低。因此,GLP-1能够被用于治疗二型糖尿病(Pharmaceuticals(Basel),3(8),2554-2567,2010;Diabetologia,36(8),741-744,1993)。进一步地,GLP-1可以诱导上消化器官运动减退并抑制食欲,且能刺激现存胰岛β细胞的增殖(EndocrRev.,16(3),390-410,1995;Endocrinology,141(12),4600-4605,2000;DigDisSci.,38(4),665-673,1993;AmJPhysiol.,273(5Pt1),E981-988,1997)。然而,GLP-1在体内两分钟的短的半衰期,对于使用GLP-1的药物制剂的发展是一个缺点。胰高血糖素样肽-1(GLP-1)调节体内平衡并在胰岛素抵抗中起着关键性的作用,因此已经被用作一种糖尿病治疗药物。进一步地,据报道GLP-1诱导STAT3激活(BiochemBiophysResCommun.,425(2),304-308,2012)。BMP-2,是TGF-β超家族中的一员,有助于软骨和骨骼的形成,并在细胞生长、细胞死亡和细胞分化中具有关键作用(GenesDev.,10,1580-1594,1996;Development,122,3725-3734,1996;JBiolChem.,274,26503-26510,1999;JExpMed.,189,1139-1147,1999)。进一步地,已报道BMP-2能够用作一种用于治疗多发性硬化症的治疗剂(Blood,96(6),2005-2011,2000;LeukLymphoma.,43(3),635-639,2002)。免疫球蛋白G(IgG)是抗体的一类,且它是血液和细胞外液中最主要的一类抗体,以允许它控制身体组织的感染;并且免疫球蛋白以单体(也就是尺寸很小)分泌,这允许它容易地散布于组织中(BasicHistology,McGraw-Hill,ISBN0-8385-0590-2,2003)。IgG被用于治疗免疫缺陷、自身免疫性疾病和感染(ProcNatlAcadSciUSA.,107(46),19985-19990,2010)。与体内稳态有关的蛋白质治疗剂具有各种各样的副作用,诸如增加癌症诱发的风险。例如,已知肠促胰岛素降解酶(DPP-4)(二肽基肽酶-4)抑制剂家族治疗剂会增加甲状腺癌的诱发的可能,而已知甘精胰岛素会增加乳腺癌的风险。进一步地,向遭受生长激素分泌紊乱的疾病的病人持续性或过量性的施用生长激素会被卷入糖尿病、微血管疾病和病人的过早死亡。就这一点而言,有广泛的研究来减少治疗性蛋白的不利和副作用。延长蛋白的半衰期被认为是减少治疗性蛋白的不利和副作用风险的一种方法。为此,大量的方法已经被公开。就这一点,本发明人已经研究开发一种用于延长体内和/或体外蛋白的半衰期的新方法,并通过替换治疗性蛋白或(多)肽中与泛素化相关的一个或更多个赖氨酸残基以阻止蛋白或者(多)肽通过泛素-蛋白酶体系统降解而完成本发明。本文所引用的全部专利、出版物以及参考文献的教导通过引用的方式全部并入本申请中。技术实现要素:技术问题本发明的目的是增加蛋白质或(多)肽的半衰期。进一步地,本发明的另一个目的是提供一种具有延长半衰期的治疗性蛋白。进一步地,本发明的另一个目的是提供一种药物组合物,包括作为药物活性成分的具有延长半衰期的蛋白。问题解决为达到目的,本发明提供一种通过替换蛋白质的氨基酸的一个或多个赖氨酸残基而在体内和/或体外延长蛋白质半衰期的方法。在本发明中,赖氨酸残基能够被保守性氨基酸替换。术语“保守性氨基酸替换”是指一个氨基酸被另外一个与该氨基酸不同但是具有相似的化学性质的氨基酸替换,例如电荷和疏水性。一般说来,使用相应地保守性氨基酸的氨基酸替换,一个蛋白质的功能特点不会被本质地改变。例如,氨基酸根据侧链具有的相似的化学性质能够被分类,如下:①脂肪族侧链:甘氨酸、丙氨酸、缬氨酸、亮氨酸、和异亮氨酸;②脂肪族-羟基侧链:丝氨酸、苏氨酸;③酰胺侧链:天冬酰胺和谷氨酰胺;④芳香族侧链:苯丙氨酸、络氨酸、色氨酸;⑤碱性侧链:赖氨酸、精氨酸和组氨酸;⑥酸性侧链:天冬氨酸和谷氨酸;和⑦含硫侧链:半胱氨酸和甲硫氨酸。在本发明中,赖氨酸残基能够被具有碱性侧链的精氨酸和组氨酸替换。优选为,赖氨酸残基被精氨酸替换。本发明的有益效果根据本发明,被精氨酸替换一个或者多个赖氨酸残基的变体蛋白质具有显著延长的半衰期,因此能够维持一段很长的时间。附图说明图1为β-调理素的表达载体的结构;图2为β-调理素基因PCR克隆产物的结果;图3为HEK-293T细胞中β-调理素质粒基因的表达;图4为通过泛素化实验解释β-调理素的蛋白质水解途径;图5为相比野生型,用精氨酸替换赖氨酸残基的经取代的β-调理素的泛素化水平;图6为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的β-调理素的半衰期改变;图7为JAK-STAT信号转导样效应的结果;图8为生长激素表达载体的结构;图9为生长激素基因PCR克隆产物的结果;图10为HEK-293T细胞中生长激素质粒基因的表达;图11为通过泛素化实验解释生长激素的蛋白质水解途径;图12为相比野生型,用精氨酸替换赖氨酸残基的经取代的生长激素的泛素化水平;图13为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的生长激素的半衰期改变;图14为JAK-STAT信号转导样效应的结果;图15为胰岛素表达载体的结构;图16为胰岛素基因PCR克隆产物的结果;图17为HEK-293T细胞中胰岛素质粒基因的表达;图18为通过泛素化实验解释胰岛素的蛋白质水解途径;图19为相比野生型,用精氨酸替换赖氨酸残基的经取代的胰岛素的泛素化水平;图20为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的胰岛素的半衰期改变;图21为JAK-STAT信号转导样效应的结果;图22为干扰素-α表达载体的结构;图23为干扰素-α基因PCR克隆产物的结果;图24为HEK-293T细胞中干扰素-α质粒基因的表达;图25为通过泛素化实验解释干扰素-α的蛋白质水解途径;图26为相比野生型,用精氨酸替替换赖氨酸残基的经取代的干扰素-α的泛素化水平;图27为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的干扰素-α的半衰期改变;图28为JAK-STAT信号转导样效应的结果;图29为G-CSF表达载体的结构;图30为G-CSF基因PCR克隆产物的结果;图31为HEK-293T细胞中G-CSF质粒基因的表达;图32为通过泛素化实验解释G-CSF的蛋白质水解途径;图33为相比野生型,用精氨酸替替换赖氨酸残基的经取代的G-CSF的泛素化水平;图34为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的G-CSF的半衰期改变;图35为JAK-STAT信号转导效应样的结果;图36为干扰素-β表达载体的结构;图37为干扰素-β基因PCR克隆产物的结果;图38为HEK-293T细胞中干扰素-β质粒基因的表达;图39为通过泛素化实验解释干扰素-β的蛋白质水解途径;图40为相比野生型,用精氨酸替替换赖氨酸残基的经取代的干扰素-β的泛素化水平;图41为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的干扰素-β的半衰期改变;图42为JAK-STAT和PI3K/AKT信号转导样效应的结果;图43为红细胞生成素表达载体的结构;图44为红细胞生成素基因PCR克隆产物的结果;图45为HEK-293T细胞中红细胞生成素质粒基因的表达;图46为通过泛素化实验解释红细胞生成素的蛋白质水解途径;图47为相比野生型,用精氨酸替换赖氨酸残基的经取代的红细胞生成素的泛素化水平;图48为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的红细胞生成素的半衰期改变;图49为MAPK/ERK信号转导效应的结果;图50为BMP2表达载体的结构;图51为BMP2基因PCR克隆产物的结果;图52为HEK-293T细胞中BMP2质粒基因的表达;图53为通过泛素化实验解释BMP2的蛋白质水解途径;图54为相比野生型,用精氨酸替换赖氨酸残基的经取代的BMP2的泛素化水平;图55为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的BMP2的半衰期改变;图56为JAK-STAT信号转导样效应的结果;图57为纤维母细胞生长因子-1(FGF-1)表达载体的结构;图58为FGF-1基因PCR克隆产物的结果;图59为HEK-293T细胞中FGF-1质粒基因的表达;图60为通过泛素化实验解释FGF-1的蛋白质水解途径;图61为相比野生型,用精氨酸替换赖氨酸残基的经取代的FGF-1的泛素化水平;图62为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的FGF-1的半衰期改变;图63为MAPK/ERK信号转导效应的结果;图64为瘦蛋白表达载体的结构;图65为瘦蛋白基因PCR克隆产物的结果;图66为HEK-293T细胞中瘦蛋白质粒基因的表达;图67为通过泛素化实验解释瘦蛋白的蛋白质水解途径;图68为相比野生型,用精氨酸替换赖氨酸残基的经取代的瘦蛋白的泛素化水平;图69为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的瘦蛋白的半衰期改变;图70为PI3K/AKT信号转导效应的结果;图71为血管内皮细胞生长因子A(VEGFA)表达载体的结构;图72为VEGFA基因PCR克隆产物的结果;图73为HEK-293T细胞中VEGFA质粒基因的表达;图74为通过泛素化实验解释VEGFA的蛋白质水解途径;图75为相比野生型,用精氨酸替换赖氨酸残基的经取代的VEGFA的泛素化水平;图76为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的VEGFA的半衰期改变;图77为JAK-STAT和PI3K/AKT信号转导样效应的结果;图78为胃饥饿素/肥胖抑制素前多肽原(Prepro-GHRL)表达载体的结构;图79为Prepro-GHRL基因PCR克隆产物的结果;图80为HEK-293T细胞中Prepro-GHRL质粒基因的表达;图81为通过泛素化实验解释Prepro-GHRL的蛋白质水解途径;图82为相比野生型,用精氨酸替换赖氨酸残基的经取代的Prepro-GHRL的泛素化水平;图83为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的Prepro-GHRL的半衰期改变;图84为JAK-STAT信号转导样效应的结果;图85为GHRL表达载体的结构;图86为GHRL基因PCR克隆产物的结果;图87为HEK-293T细胞中GHRL质粒基因的表达;图88为通过泛素化实验解释GHRL的蛋白质水解途径;图89为相比野生型,用精氨酸替换中赖氨酸残基的经取代的GHRL的泛素化水平;图90为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的GHRL的半衰期改变;图91为JAK-STAT信号转导样效应的结果;图92为胰高血糖素样肽-1(GLP-1)表达载体的结构;图93为GLP-1基因PCR克隆产物的结果;图94为HEK-293T细胞中GLP-1质粒基因的表达;图95为通过泛素化实验解释GLP-1的蛋白质水解途径;图96为相比野生型,用精氨酸替换赖氨酸残基的经取代的GLP-1的泛素化水平;图97为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的GLP-1的半衰期改变;图98为JAK-STAT信号转导样效应的结果;图99为IgG重链的表达载体的结构;图100为IgG重链基因PCR克隆产物的结果;图101为HEK-293T细胞中IgG重链质粒基因的表达;图102为通过泛素化实验解释IgG重链的蛋白质水解途径;图103为相比野生型,用精氨酸替换赖氨酸残基的经取代的IgG重链的泛素化水平;图104为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的IgG重链的半衰期改变;图105为IgG轻链的表达载体的结构;图106为IgG轻链基因PCR克隆产物的结果;图107为HEK-293T细胞中IgG轻链质粒基因的表达;图108为通过泛素化实验解释IgG轻链的蛋白质水解途径;图109为相比野生型,用精氨酸替换赖氨酸残基的经取代的IgG轻链的泛素化水平;图110为经过蛋白合成抑制剂环十二碳三烯(CHX)处理之后的IgG轻链的半衰期改变;在下文中,将结合实施例对本发明进行更为详细的描述。应该明白的是这些实施例不能以任何方式被考虑为限定本发明。具体实施方式在本发明的一个实施方案中,所述蛋白是β-调理素。在β-调理素的氨基酸序列中(SEQNo.1),对应于N端第62,124,153和158位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的在体内和/或体外半衰期的β-调理素。进一步地,提供了一种用于预防和/或治疗糖尿病和肥胖的包含经取代的β-调理素的药物组合物(Cell,153(4),747758,2013;CellMetab.,18(1),5-6,2013;FrontEndocrinol(Lausanne),4,146,2013)。在本发明的另一个实施方案中,所述蛋白是生长激素。在生长激素的氨基酸序列中(SEQNo.10),对应于N端第64、67、96、141、166、171、184、194和198位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的在体内和/或体外半衰期的生长激素。进一步地,提供了一种用于预防和/或治疗侏儒症,歌舞伎综合征,卡恩斯-塞尔综合征的包含经替代的生长激素的药物组合物(JEndocrinolInvest.,39(6),667-677,2016;JPediatrEndocrinolMetab.,2016,[Epubaheadofprint];HormResPaediatr.2016,[Epubaheadofprint])。在本发明的另一个实施方案中,所述蛋白是胰岛素。在胰岛素的氨基酸序列中(SEQNo.17),对应于N端第53和88位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的半衰期的胰岛素。进一步地,提供了一种用于预防和/或治疗糖尿病的包含经取代的胰岛素的药物组合物。在本发明的另一个实施方案中,所述蛋白是干扰素-α。在干扰素-α的氨基酸序列中(SEQNo.22),对应于N端第17、54、72、93、106、135、144、154、156、157和187位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的在体内和/或体外半衰期的干扰素-α。进一步地,提供了一种用于预防和/或治疗免疫疾病的包含经取代的干扰素-α药物组合物,所述免疫疾病包括多发性硬化,自身免疫性疾病,类风湿性关节炎;和/或癌症,包括实体肿瘤和/或血癌;和/或感染性疾病包括病毒感染,艾滋病相关疾病和丙肝。需要用干扰素-α进行治疗的疾病或紊乱被提供(AnnRheumDis.,42(6),672-676,1983;Memo.,9,63-65,2016)。在本发明的另一个实施方案中,所述蛋白是G-CSF。在G-CSF的氨基酸序列中(SEQNo.31),对应于N端第11、46、53、64和73位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有延长的在体内和/或体外半衰期的G-CSF。进一步地,提供了一种用于预防和/或治疗嗜中性白细胞减少症的包含经取代的G-CSF而药物组合物(EMBOMolMed.2016,[Epubaheadofprint])。在本发明的一个实施方案中,所述蛋白是干扰素-β。在干扰素-β的氨基酸序列中(SEQNo.36),对应于N端的第4、40、54、66、73、120、126、129、136、144、155和157位的至少有一个赖氨酸被替换为精氨酸。因此,提供了具有延长的在体内和/或体外半衰期的干扰素-β。进一步地,提供了一种包含经取代的干扰素-β的药物组合物以用于预防和/或治疗免疫疾病,所述免疫疾病包括多发性硬化,自身免疫性疾病,类风湿性关节炎;和/或癌症,包括实体肿瘤和/或血癌;和/或感染性疾病包括病毒感染,艾滋病相关疾病和丙肝。在本发明的另一个实施方案中,所述蛋白是红细胞生成素。在红细胞生成素的氨基酸序列中(SEQNo.43),对应于N端第47、72、79、124、143、167、179和181位的至少有一个赖氨酸被替换为精氨酸。因此,提供了具有增加的在体内和/或体外半衰期延长的红细胞生成素。进一步地,提供了包含经取代的红细胞生成素的药物组合物以预防和/或治疗慢性肾衰竭,手术和癌症或癌症治疗等导致的贫血。在本发明的另一个实施方案中,所述蛋白是骨形态生成蛋白-2(BMP-2)。在BMP-2的氨基酸序列中(SEQNo.52),对应于N端第32、64、127、178、185、236、241、272、278、281、285、287、290、293、297、355、358、379和383位的至少有一个赖氨酸被替换为精氨酸。因此,提供了具有增加的半衰期的BMP-2。进一步地,提供了包含经取代的BMP-2的药物组合物以预防和/或治疗品贫血和骨病(CellJ.,17(2),193-200,2015;ClinOrthopRelatRes.,318,222-230,1995)。在本发明的另一个实施例中,所述蛋白是纤维母细胞生成因子-1(FGF-1)。在FGF-1的氨基酸序列中(SEQNo.61),对应于N端第15、24、25、27、72、115、116、120、127、128、133和143位的至少有一个赖氨酸被替换为精氨酸。因此,提供了具有增加的半衰期的FGF-1。进一步地,提供了包含经取代的FGF-1的药物组合物以预防和/或治疗神经元疾病。在本发明的另一个实施方案中,所述蛋白是食欲抑制激素(瘦蛋白)。在食欲抑制激素(瘦蛋白)的氨基酸序列中(SEQNo.66),对应于N端第26、32、36、54、56、74和115位的至少有一个至少有一个赖氨酸被替换为精氨酸。因此,提供了具有增加的半衰期的食欲抑制激素(瘦蛋白)。进一步地,提供了用于预防和/或治疗脑部疾病,心脏病和/或肥胖的包含经取代的食欲抑制激素(瘦蛋白)的药物组合物(AnnNYAcadSci.,1243,1529,2011;JNeurochem.,128(1),162-172,2014;ClinExpPharmacolPhysiol.,38(12),905-913,2011)。在本发明的另一个实施方案中,所述蛋白是VEGFA。在VEGFA的氨基酸序列中(SEQNo.75),对应于N端第22、42、74、110、127、133、134、141、142、147、149、152、154、156、157、169、180、184、191和206位的至少有一个赖氨酸被替换为精氨酸。因此,提供了具有增加的半衰期的VEGFA和包含其的药物组合物,以预防和/或治疗抗衰老,毛发生长,疤痕和/或血管生成相关疾病。在本发明的另一个实施方案中,所述蛋白是食欲刺激激素前体,肥胖抑制素前激素原(prepro-GHRL)。在食欲刺激激素前体的氨基酸序列中(SEQNo.80),对应于N端第39、42、43、47、85、100、111和117位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的半衰期的食欲刺激激素前体。进一步地,提供了一种包含替换的食欲刺激激素前体的药物组合物以预防和/或治疗肥胖,营养不良,和/或诸如神经性食欲缺乏的饮食失调。在本发明的另一个实施例中,所述蛋白是食欲刺激激素(胃饥饿素)。在胃饥饿素的氨基酸序列中(SEQNo.83),对应于N端第39、42、43和47位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的半衰期的食欲刺激激素(胃饥饿素)。进一步地,提供了一种包含替换的胃饥饿素的药物组合物以预防和/或治疗肥胖,营养不良,和/或诸如神经性食欲缺乏的饮食失调。在本发明的另一个实施方案中,所述蛋白是胰高血糖素样肽-1(GLP-1)。在GLP-1的氨基酸序列中(SEQNo.92),对应于N端第117和125位的至少有一个赖氨酸被替换为精氨酸。因此,提供了用于预防和/或治疗糖尿病的一种具有增加的半衰期的GLP-1和包含其的药物组合物。在本发明的另一个实施方案中,所述蛋白是IgG。在IgG重链的氨基酸序列中(SEQNo.97),对应于N端第49、62、84、95、143、155、169、227、232、235、236、240、244、268、270、296、310、312、339、342、344、348、356、360、362、382、392、414、431、436和461位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的半衰期的IgG和包含其的药物组合物以预防和/或治疗癌症。在本发明的另一个实施方案中,所述蛋白是IgG。在IgG轻链的氨基酸序列中(SEQNo.104),对应于N端第61、64、67、125、129、148、167、171、191、205、210、212和229位的至少有一个赖氨酸被替换为精氨酸。因此,提供了一种具有增加的半衰期的IgG和包含其的药物组合物以预防和/或治疗癌症。在本发明中,采用定点突变的方法将蛋白质氨基酸序列中的赖氨酸残基替换为精氨酸(R)。根据该方法,使用DNA序列设计引物而诱导定点突变,然后在特定条件下通过执行PCR而产生突变的质粒DNA。在本发明中,通过向细胞系转染目标蛋白和使用免疫沉淀反应测量泛素化程度。如果在使用MG132处理之后的被转染细胞系中泛素化水平升高,这就说明目标蛋白通过泛素-蛋白酶体途径被降解。本发明的药物组合物可以通过多种方式被施用进入身体,包括口服,经皮肤,经皮下,静脉内或肌肉施用,而更优选方案则是通过注射的形式制备。进一步地,本发明的药物组合物还可以通过本领域技术人员熟知的方法而被制作而在施用后提供活性物质的快速,保持或延迟地释放。制剂的形式可以为片剂、丸剂、粉剂、小袋、酏剂、悬浮剂、乳剂、溶液、糖浆、气雾剂、软和硬的胶囊、无菌注射液、无菌压缩粉末等。合适的载体、赋形剂和稀释剂的例子是乳糖、葡萄糖、蔗糖、甘露醇、木糖醇、赤藓醇、麦芽糖醇、淀粉、阿拉伯树胶、海藻酸盐、明胶、磷酸钙、硅酸钙、纤维素、甲基纤维素、微晶纤维素、聚乙烯吡咯烷酮、水、对羟基苯甲酸甲酯、对羟基苯甲酸丙酯、滑石、硬脂酸镁和矿物油。进一步地,药剂可以另外地包括填充料、抗粘剂、润滑剂、湿润剂、芳香剂、乳化剂、防腐剂等。合适的载体、赋形剂、稀释剂的例子是乳糖、葡萄糖、蔗糖、甘露醇、木糖醇、赤藓醇、麦芽糖醇、淀粉、阿拉伯树胶、海藻酸盐、明胶、磷酸钙、硅酸钙、纤维素、甲基纤维素、微晶纤维素、聚乙烯吡咯烷酮、水、对羟基苯甲酸甲酯、对羟基苯甲酸丙酯、滑石、硬脂酸镁、矿物油。进一步的,药剂可以另外地包括填充料、抗粘剂、润滑剂、湿润剂、芳香剂、乳化剂、防腐剂等。在此文中使用的,单数形式“a”,“an”,“the”也意图包括复数形式,除非本文中清楚地表明其他含义。此外,说明书和/或权利要求中使用的术语“包括(including)”,“包括(includes)”,“具有(having)”,“具有(has)”,“和(with)”,“例如”或它们的变体形式,并不起限定作用,也旨在是包括的方式类似于术语“包括(comprising)”。在本发明中,“生物活性的多肽或蛋白”是当被施用给哺乳动物包括人类时,表现出有用的生物学活性的(多)肽或蛋白。发明方式下述实施例提供了说明性的实施方案。根据本发明公开的内容和本领域技术人员的通常水平,本领域技术人员应理解下述实施例仅意图作为示例,且不脱离本发明请求保护的主题的范围能够做出很多改变、修饰和变化。实施例1β-调理素泛素化和半衰期延长以及细胞中的信号转导检测的分析1.β-调理素表达载体克隆和蛋白表达(1)β-调理素表达载体克隆使用苯酚和三氯甲烷从HepG2(ATCC,HB-8065)细胞中提取和纯化RNA以克隆β-调理素。于是,通过使用SuperScriptTMFirst-StrandcDNA合成系统(Invitrogen,GrandIsland,NY)合成单链DNA。再使用上述合成的cDNA作为模板扩增β-调理素。获得的β-调理素DNA扩增产物再经由BamHI和EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图1,β-调理素氨基酸序列:SEQNo.1)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图2)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃1分钟(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。图1中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图2)。为了分析蛋白表达,利用抗-myc抗体(9E10,sc-40)与图1图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示β-调理素被很好地表达。肌动蛋白作为标准确保蛋白质上样量是合适的(表3)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以产生经取代的质粒DNA。(β-调理素K62R)FP5'-AGGGACGGCTGACAAGGGCCAGGAA-3'(SEQNo.2),RP5'-CCAGGCTGTTCCTGGCCCTTGTCAGC-3'(SEQNo.3);(β-调理素K124R)FP5'-GGCACAGAGGGTGCTACGGGACAGC-3'(SEQNo.4),RP5'-CGTAGCACCCTCTGTGCCTGGGCCA-3'(SEQNo.5);(β-调理素K153R)FP5'-GAATTTGAGGTCTTAAGGGCTCACGC-3'(SEQNo.6),RP5'-CTTGTCAGCGTGAGCCCTTAAGACCTC-3'(SEQNo.7);和(β-调理素K158R)FP5'-GCTCACGCTGACAGGCAGAGCCACAT-3'(SEQNo.8),RP5'-CCATAGGATGTGGCTCTGCCTGTCAGC-3'(SEQNo.9)。通过使用pcDNA3-myc-β-调理素作为模板,制备了一个或更多个赖氨酸被取代为精氨酸(K→R)的4种质粒DNA(表1)。表1赖氨酸(K)残基构建的β-调理素,精氨酸替换赖氨酸62pcDNA3-myc-β-trophin(K62R)124pcDNA3-myc-β-trophin(K124R)153pcDNA3-myc-β-trophin(K153R)158pcDNA3-myc-β-trophin(K158R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-β-trophinWT和pMT123-HA-ubiquitin的质粒(JBiolChem.,279(4),2368-2376,2004;CellResearch,22,873885,2012;Oncogene,22,12731280,2003;Cell,78,787-798,1994)。为了分析泛素化的程度,pcDNA3-myc-β-trophinWT2μg和pMT123-HA-ubiquitin1μg被共转染入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,此后再进行免疫沉淀反应分析(图4)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-β-trophinWT,pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K124R),pcDNA3-myc-β-trophin变体(K153R)和pcDNA3-myc-β-trophin变体(K158R)的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与pcDNA3-myc-β-trophinWT,pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K124R),pcDNA3-myc-β-trophin变体(K153R),pcDNA3-myc-β-trophin变体(K158R)2μg共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图5)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。于是,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。接下来,分离的免疫沉淀物用缓冲液洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,之后使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc(9E10,sc-40),抗HA(SantaCruzBiotechnology,sc-7392)和抗β-actin抗体(SantaCruzBiotechnology,sc-47778)封闭液再通过ECL系统显影。因此,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-β-trophinWT的结合而形成。因此强烈的表明泛素存在的条带被产生(图4,泳道3和4)。进一步地,当细胞被MG132(蛋白酶体抑制剂,5μg/ml)处理六小时时,多聚泛素链的形成增加,因此更加强烈的代表泛素的条带被展示(图4,泳道4)。至于pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K153R)和pcDNA3-myc-β-trophin变体(K158R),条带不如野生型的强烈。这些结果表明,较少数量的泛素被检测到,因为,这些变体质粒不能与泛素结合(图5,泳道3,5和6)。这些结果说明β-调理素先结合泛素,再是多聚泛素链,再通过形成的多聚泛素链而被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估β-调理素半衰期HEK293T细胞被分别转入2μgpcDNA3-myc-β-trophinWT,pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K124R),pcDNA3-myc-β-trophin变体(K153R)和pcDNA3-myc-β-trophin变体(K158R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在蛋白合成抑制剂处理之后的20分钟,40分钟和60分钟时,分别检测每种蛋白的半衰期。结果,观察人β-调理素的降解(图6)。人β-调理素的半衰期少于1小时,而β-调理素变体(K62R)和β-调理素变体(K158R)的半衰期被延长至1小时或者更多,如图6所示。4.β-调理素和经取代的β-调理素在细胞中的信号转导据报道在小鼠肝脏内β-调理素的暂时性表达促进胰腺β细胞增殖(Cell,153,747-758,2013)。在这个实验中,我们检测了β-调理素和经取代的β-调理素在细胞中的信号转导。首先,PANC-1细胞用PBS清洗7次,再分别转染3μgpcDNA3-myc-β-trophinWT,pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K124R),pcDNA3-myc-β-trophin变体(K153R)和pcDNA3-myc-β-trophin变体(K158R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。为此,从分别转染pcDNA3-myc-β-trophinWT,pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K124R),pcDNA3-myc-β-trophin变体(K153R)和pcDNA3-myc-β-trophin变体(K158R)的PANC-1细胞分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,再使用以1:1000抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有稀释的抗myc(9E10,SantaCruzBiotechnology,sc-40),抗STAT3(SantaCruzBiotechnology,sc-21876),抗磷酸化-STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(SantaCruzBiotechnology,sc-47778)封闭液通过ECL系统对蛋白进行显影。因此,相比于野生型,pcDNA3-myc-β-trophin变体(K62R),pcDNA3-myc-β-trophin变体(K124R)和pcDNA3-myc-β-trophin变体(K153R)在PANC-1细胞中,显示出一样或者增加的磷酸化-STAT3信号转导。实施例2生长激素泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.GH表达载体克隆和蛋白表达(1)GH表达载体克隆用EcoRI处理通过PCR扩增的GHDNA,再连接到预先以同样的酶消化的pCS4-flag(4.3kb,Oncotarget.,7(12),14441-14457,2016)载体上(图8,生长激素氨基酸序列:SEQNo.10)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图9)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;60℃30秒;72℃30秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。图8中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图9)。为了分析蛋白表达,利用抗-myc抗体(9E10,sc-40)与图8图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示β-调理素被很好地表达。肌动蛋白作为标准确保蛋白质上样量是合适的(表10)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以产生经取代的质粒DNA。(GHK67R)FP5'-CCAAAGGAACAGAGGTATTCATTC-3'(SEQNo.11),RP5'-CAGGAATGAATACCTCTGTTCCTT-3'(SEQNo.12);(GHK141R)FP5'-GACCTCCTAAGGGACCTAGAG-3'(SEQNo.13),RP5'-CTCTAGGTCCCTTAGGAGGTC-3'(SEQNo.14);和(GHK166R)FP5'-CAGATCTTCAGGCAGACCTAC-3'(SEQNo.15),RP5'-GTAGGTCTGCCTGAAGATCTG-3'(SEQNo.16)通过使用pcDNA3-myc-β-生长激素作为模板,产生有一个或更多个赖氨酸被替换为精氨酸(K→R)的三种质粒DNA(表2)。表2赖氨酸(K)残基构建的生长激素,精氨酸替换赖氨酸67pCS4-flag-GH(K67R)141pCS4-flag-GH(K141R)166pCS4-flag-GH(K166R)2.体内泛素化分析HEK293T细胞被转染编码pCS4-flag-GHWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pCS4-flag-GHWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,此后再进行免疫沉淀反应分析(图11)。于是,HEK293T细胞被分别转入编码pCS4-flag-GHWT,pCS4-flag-GH变体(K67R),pCS4-flag-GH变体(K141R),pCS4-flag-GH变体(K166R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpCS4-flag-growthhormoneWT,pCS4-flag-growthhormone变体(K67R),pCS4-flag-growthhormone变体(K141R)和pCS4-flag-growthhormone变体(K166R)共转染入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图12)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗flag(Sigma-aldrich,F3165)一抗(SantaCruzBiotechnology,sc-40)混合,随后混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。于是,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗flag(Sigma-aldrich,F3165),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液再通过ECL系统显影。因此,当使用抗flag(Sigma-aldrich,F3165)抗体进行免疫沉淀反应实验时,多聚泛素链通过泛素与pCS4-flag-growthhormoneWT的结合而形成。因此强烈的表明泛素的条带被产生(图11,泳道2和3)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,因此更加强烈的代表泛素的条带被展示(图11,泳道3)。进一步地,至于pCS4-flag-growthhormone变体(K67R),pCS4-flag-growthhormone变体(K141R)和pCS4-flag-growthhormone变体(K166R),条带不如野生型的强烈。这些结果表明较少数量的泛素被检测到,因为,变体质粒不能与泛素结合(图12,泳道3-5)。这些结果说明β-调理素先结合泛素,再形成多聚泛素链,再通过形成的多聚泛素链而被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估生长激素的半衰期HEK293T细胞被分别的转入2μgpCS4-flag-GHWT,pCS4-flag-GH变体(K67R),pCS4-flag-GH变体(K141R)和pCS4-flag-GH变体(K166R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在所述抑制剂处理之后的1小时,2小时,4小时和8小时,分别检测每种蛋白的半衰期。结果,观察人生长激素的降解(图13)。人生长激素的半衰期少于2小时,而pCS4-flag-生长激素变体(K141R)的半衰期被延长至8小时或者更多,如图13所示。4.生长激素和经取代的生长激素在细胞中的信号转导有研究报道生长激素控制STAT(信号转导和转录激活因子)蛋白的转录(Oncogene,19,2585-2597,2000)。在这个实验中,我们检测了生长激素和经取代的生长激素在细胞中的信号转导。首先,HEK293T细胞被分别的转入3μgpCS4-flag-growthhormoneWT,pCS4-flag-growthhormone变体(K67R),pCS4-flag-growthhormone变体(K141R)和pCS4-flag-growthhormone变体(K166R)。转染1天后,蛋白通过声波裂解法裂解细胞而获取。PANC-1细胞(ATCC,CRL-1469)用PBS洗七次,再转染3μg上述获得的蛋白。蛋白质印记法被用于分析细胞中的信号转导。为此,从分别转染pCS4-flag-growthhormoneWT,pCS4-flag-growthhormone变体(K67R),pCS4-flag-growthhormone变体(K141R)和pCS4-flag-growthhormone变体(K166R)的PANC-1细胞分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。接下来,再使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗STAT3(sc-21876),抗磷酸化-STAT3(Y705,CellSignaling9131S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。因此,相比于pCS4-flag-growthhormoneWT,pCS4-flag-GH变体(K141R)在PANC-1细胞中,显示一样或者增加的磷酸化-STAT3信号转导水平,和相比于对照,pCS4-flag-growthhormone变体(K67R)显示增加的磷酸化-STAT3信号转导(图14)。实施例3胰岛素泛素化和半衰期延长分析,以及细胞中的信号转导的分析1.胰岛素表达载体克隆和蛋白表达(1)胰岛素表达载体克隆通过PCR的胰岛素DNA扩增产物被BamHI和EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图15,胰岛素氨基酸序列:SEQNo.17)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图16)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;60℃30秒;72℃30秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。图15中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图16)。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图15图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示胰岛素被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图17)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸。下述引物设定被用于PCR以制备经取代的质粒DNA。(insulinK53R)FP5'-GGCTTCTTCTACACACCCAGGACCC-3'(SEQNo.18),RP5'-CTCCCGGCGGGTCCTGGGTGTGTA-3'(SEQNo.19);和(insulinK88R)FP5'-TCCCTGCAGAGGCGTGGCATTGT-3'(SEQNo.20),RP5'-TTGTTCCACAATGCCACGCCTCTGCAG-3'(SEQNo.21)通过使用pcDNA3-myc-胰岛素作为模板,产生有一个或更多个赖氨酸被替换为精氨酸(K→R)的2种质粒DNA(表3)。表3赖氨酸(K)残基构建的胰岛素,精氨酸替换赖氨酸53pcDNA3-myc-insulin(K53R)88pcDNA3-myc-insulin(K88R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-insulinWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-insulinWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应(图18)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-insulinWT,pcDNA3-myc-insulin变体(K53R),pcDNA3-myc-insulin变体(K88R)和pMT123-HA-ubiquitin的质粒。进一步地,为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-insulinWT,pcDNA3-myc-insulin变体(K53R)和pcDNA3-myc-insulin变体(K88R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图19)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc抗体(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后将混合物孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,之后使用抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有以1:1000稀释的抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液再通过ECL系统显影。因此,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链可以通过泛素与pcDNA3-myc-insulinWT的结合而形成。因此强烈的表明泛素存在的条带被产生(图18,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,因此更加强烈的表明泛素的条带被展示(图18,泳道4)。进一步地,至于pcDNA3-myc-insulin变体(K53R),条带不如野生型的强烈,且较少数量的泛素被检测,因为,pcDNA3-myc-insulin变体(K53R)不能与泛素相结合(图19,泳道3)。这些结果说明胰岛素先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估胰岛素的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-insulinWT,pcDNA3-myc-insulin变体(K53R)和pcDNA3-myc-insulin变体(K88R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在蛋白合成抑制剂处理之后的2小时,4小时和8小时,分别检测每种蛋白的半衰期。再观察人胰岛素的降解(图20)。人胰岛素的半衰期少于30分钟,而人胰岛素变体(K53R)的半衰期被延长至1小时或者更多,如图20所示。4.胰岛素和经取代的胰岛素在细胞中的信号转导有研究报道胰岛素刺激肝脏中STAT磷酸化,从而控制肝脏中的葡萄糖稳态(CellMetab.,3,267275,2006)。在这个实验中,我们检测了胰岛素和经取代的胰岛素在细胞中的信号转导。首先,PANC-1和HepG2细胞用PBS洗七次,再分别转入3μgpcDNA3-myc-insulinWT,pcDNA3-myc-insulin变体(K53R)和pcDNA3-myc-insulin变体(K88R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。为此,从分别转染pcDNA3-myc-insulinWT,pcDNA3-myc-insulin变体(K53R)和pcDNA3-myc-insulin变体(K88R)的PANC-1细胞和HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗STAT3(sc-21876),抗磷酸化-STAT3(Y705,CellSignaling9131S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于pcDNA3-myc-insulinWT,pcDNA3-myc-insulin变体(K53R)在PANC-1细胞和HepG2细胞中,显示一样或者增加的磷酸化-STAT3信号转导(图21)。实施例4干扰素-α泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.干扰素-α表达载体克隆和蛋白表达(1)干扰素-α表达载体克隆通过PCR扩增的干扰素-αDNA被EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图22,干扰素-α氨基酸序列:SEQNo.22)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图23)。图22中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图23)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃1分钟(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图22图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示结合myc的干扰素-α的被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图24)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(IFN-αK93R)FP5'-CTTCAGCACAAGGGACTCATC-3'(SEQNo.23),RP5'-CAGATGAGTCCCTTGTGCTGA-3'(SEQNo.24);(IFN-αK106R)FP5'-CTCCTAGACAGATTCTACACT-3'(SEQNo.25),RP5'-AGTGTAGAATCTGTCTAGGAG-3'(SEQNo.26);(IFN-αK144R)FP5'-GCTGTGAGGAGATACTTCCAA-3'(SEQNo.27),RP5'-TTGGAAGTATCTCCTCACAGC-3'(SEQNo.28);和(IFN-αK154R)P5'-CTCTATCTGAGAGAGAAGAAA-3'(SEQNo.29),RP5'-TTTCTTCTCTCTCAGATAGAG-3'(SEQNo.30)通过使用pcDNA3-myc-干扰素-α作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的4种质粒DNA(表4)。表4赖氨酸(K)残基构建的干扰素-α,精氨酸替换赖氨酸93pcDNA3-myc-IFN-α(K93R)106pcDNA3-myc-IFN-α(K106R)144pcDNA3-myc-IFN-α(K144R)154pcDNA3-myc-IFN-α(K154R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-interferon-αWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-interferon-αWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图25)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-interferon-αWT,pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R),pcDNA3-myc-interferon-α(K154R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-interferon-αWT,pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R)和pcDNA3-myc-interferon-α(K154R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图26)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。接下来,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。因此,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链可以通过泛素与pcDNA3-myc-interferon-αWT的结合而形成。所以强烈的条带表明泛素的存在被检测到(图25,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,所以更强烈的表明泛素的条带被产生(图25,泳道4)。进一步地,至于pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R)和pcDNA3-myc-interferon-α(K154R),条带不如野生型的强烈,且较少数量的泛素被检测到,因为,变体质粒不能与泛素相结合(图26,泳道3-6)。这些结果说明干扰素-α先连接泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估干扰素-α的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-interferon-αWT,pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R)和pcDNA3-myc-interferon-α(K154R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在蛋白酶合成抑制剂处理之后的一天和两天,分别检测每种蛋白的半衰期。结果,观察人干扰素-α的降解(图27)。人干扰素-α的半衰期少于一天,而人干扰素-α变体(K93R),人干扰素-α变体(K144R)和人干扰素-α变体(K154R)的半衰期被延长至两天或者更多,如图27所示。4。干扰素-α和经取代的干扰素-α在细胞中的信号转导有研究报道在干扰素-α增强STAT-1,STAT-2,STAT-3(JImmunol.,187,2578-2585,2011),且干扰素-α激活STAT3蛋白有助于黑色素瘤的发生(EurJCancer,45,1315-1323,2009)。在这个实验中,我们检测了干扰素-α和经取代的干扰素-α在细胞中的信号转导。首先,THP-1细胞(ATCC,TIB-202)用PBS洗七次,再分别转入3μgpcDNA3-myc-interferon-αWT,pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R)和pcDNA3-myc-interferon-α(K154R)。转染1和2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pcDNA3-myc-interferon-αWT,pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R),pcDNA3-myc-interferon-α(K154R)的THP-1细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗STAT3(sc-21876),抗磷酸化-STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(sc-47778)的封闭液通过ECL系统对蛋白进行显影。因此,相比于pcDNA3-myc-interferon-αWT,pcDNA3-myc-interferon-α变体(K93R),pcDNA3-myc-interferon-α变体(K106R),pcDNA3-myc-interferon-α(K144R)和pcDNA3-myc-interferon-α变体(K154R)在THP-1细胞中,显示一样或者增加的磷酸化-STAT3信号转导(图28)。实施例5G-CSF泛素化和半衰期延长分析,以及细胞中的信号转导的分析1.G-CSF表达载体克隆和蛋白表达(1)G-CSF表达载体克隆通过PCR扩增G-CSFDNA被EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图29,G-CSF氨基酸序列:SEQNo.31)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图30)。图29中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图30)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃1分钟(25次循环);第三步:72℃10分钟(循环1次),之后再保存于4℃。为了评估克隆DNA编码的蛋白的表达,利用抗-myc抗体(9E10,sc-40)与图29图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示结合myc的G-CSF被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图31)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(G-CSFK46R)FP5'-AGCTTCCTGCTCAGGTGCTTAGAG-3'(SEQNo.32),RP5'-TTGCTCTAAGCACCTGAGCAGGAA-3'(SEQNo.33);和(G-CSFK73R)FP5'-TGTGCCACCTACAGGCTGTGCCAC-3'(SEQNo.34),RP5'-GGGGTGGCACAGCCTGTAGGTGGC-3'(SEQNo.35)通过使用pcDNA3-myc-G-CSF作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的2种质粒DNA(表5)。表5赖氨酸(K)残基构建的G-CSF,精氨酸替换赖氨酸46pcDNA3-myc-G-CSF(K46R)73pcDNA3-myc-G-CSF(K73R)2体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-G-CSFWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-G-CSFWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图32)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-G-CSFWT,pcDNA3-myc-G-CSF变体(K46R),pcDNA3-myc-G-CSF变体(K73R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-G-CSFWT,pcDNA3-myc-G-CSF变体(K46R),pcDNA3-myc-G-CSF变体(K73R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图33)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。因此,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-G-CSFWT的结合而形成。所以强烈的表明泛素存在的条带被检测到(图32,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,所以更加强烈的代表泛素的条带被产生(图32,泳道4)。进一步地,至于pcDNA3-myc-G-CSF变体(K73R),条带不如野生型的强烈。且较少数量的泛素被检测,因为,pcDNA3-myc-G-CSF变体不能与泛素相结合(K73R)(图33,泳道4)。这些结果说明G-CSF先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估G-CSF的半衰期HEK293T细胞被分别地转入2μgpcDNA3-myc-G-CSFWT,pcDNA3-myc-G-CSF变体(K46R),pcDNA3-myc-G-CSF变体(K73R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在蛋白酶合成抑制剂处理之后的4小时,8小时和16小时,分别检测每种蛋白的半衰期。结果,观察人G-CSF的降解(图34)。人G-CSF的半衰期少于大约4小时,而人G-CSF变体(K73R)的半衰期被延长至16小时或者更多,如图34所示。4.G-CSF和经取代的G-CSF在细胞中的信号转导有研究报道G-CSF激活神经胶质瘤细胞中STAT-3,从而参与神经胶质瘤生长(CancerBiolTher.,13(6),389-400,2012)。进一步地,报道G-CSF在卵巢上皮癌细胞中表达,并通过调节JAK2/STAT3通路与女性子宫癌病理性相关(BrJCancer,110,133-145,2014)。在这个实验中,我们检测了G-CSF和经取代的G-CSF在细胞中的信号转导。首先,THP-1细胞(ATCC,TIB-202)用PBS洗七次,再分别转入3μgpcDNA3-myc-G-CSFWT,pcDNA3-myc-G-CSF变体(K46R)和pcDNA3-myc-G-CSF变体(K73R)。转染1天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pcDNA3-myc-G-CSFWT,pcDNA3-myc-G-CSF变体(K46R)和pcDNA3-myc-G-CSF变体(K73R)的THP-1细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,再使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗STAT3(sc-21876),抗磷酸化-STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于野生型,pcDNA3-myc-G-CSF变体(K46R)和pcDNA3-myc-G-CSF变体(K73R)在THP-1细胞中,显示一样或者增加的磷酸化-STAT3信号转导(图35)。实施例6干扰素-β泛素化和半衰期延长分析,以及细胞中的信号转导的分析1.干扰素-β表达载体克隆和蛋白表达(1)干扰素-β表达载体克隆通过PCR扩增的干扰素-βDNA被EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图36,干扰素-β氨基酸序列:SEQNo.36)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图37)。图36中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图37)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃50秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了分析克隆DNA编码的蛋白的表达,利用抗-myc抗体(9E10,sc-40)与图36图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的干扰素-β被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图38)。进一步地,至于干扰素-β,两种表达条带会在细胞中通过糖基化而产生。在通过500单位的PNGaseF(NewEnglandBiolabsInc.,P0704S)处理细胞,阻断通路后,仅一条带被检测到(图38)(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(IFN-βK40R)FP5'-CAGTGTCAGAGGCTCCTGTGG-3'(SEQNo.37),RP5'-CCACAGGAGCCTCTGACACTG-3'(SEQNo.38);(IFN-βK126R)FP5'-CTGGAAGAAAGACTGGAGAAA-3'(SEQNo.39),RP5'-TTTCTCCAGTCTTTCTTCCAG-3'(SEQNo.40);和(IFN-βK155R)FP5'-CATTACCTGAGGGCCAAGGAG-3'(SEQNo.41),RP5'-CTCCTTGGCCCTCAGGTAATG-3'(SEQNo.42)通过使用pcDNA3-myc-干扰素-β素作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的3种质粒DNA(表3)。表62体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-interferon-βWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-interferon-βWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图39)。进一步地,HEK293T细胞被分别转入pcDNA3-myc-interferon-βWT,pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R),pcDNA3-myc-IFN-β变体(K155R)和pMT123-HA-ubiquitin。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-interferon-βWT,pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图40)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-interferon-βWT的结合而形成,且强烈的表明泛素存在的条带被检测到(图39,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,所以出现更加强烈的代表泛素的条带(图39,泳道4)。进一步地,至于pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R),条带不如野生型的强烈且较少数量的泛素被检测到,因为,变体质粒不能与泛素相结合(图40,泳道3-5)。这些结果说明干扰素-β先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估干扰素-β的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-interferon-βWT,pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的4小时和8小时,分别检测每种蛋白的半衰期。结果,观察人干扰素-β的降解(图41)。人干扰素-α的半衰期少于4小时,而pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R)的半衰期被延长至8小时或者更多,如图41所示。4.干扰素-β和经取代的干扰素-β在细胞中的信号转导有研究报道通过干扰素-β处理细胞,诱导包括AKT的信号通路的激活(Pharmaceuticals(Basel),3,994-1015,2010)。在这个实验中,我们检测了干扰素-β和经取代的干扰素-β在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入3μgpcDNA3-myc-interferon-βWT,pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R)。转染1天后,蛋白从通过声波裂解法裂解的HepG2细胞中获得。蛋白被转染进用PBS清洗7次的HepG2细胞中。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pcDNA3-myc-interferon-βWT,pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗STAT3(sc-21876),抗磷酸化-STAT3(Y705,cellsignaling9131S),抗-AKT(H-136,sc-8312),抗磷酸化-AKT(S473,cellsignaling9271S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于野生型,pcDNA3-myc-IFN-β变体(K40R),pcDNA3-myc-IFN-β变体(K126R)和pcDNA3-myc-IFN-β变体(K155R)在HepG2细胞中显示出一样或者增加的磷酸化-AKT信号转导(图42)。实施例7红细胞生成素(EPO)泛素化和半衰期延长分析,以细胞中的信号转导的分析1.红细胞生成素(EPO)表达载体克隆和蛋白表达(1)红细胞生成素(EPO)表达载体克隆通过PCR扩增的红细胞生成素(EPO)DNA被EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图43,红细胞生成素氨基酸序列:SEQNo.43)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图44)。图43中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图44)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃1分钟(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图43图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的红细胞生成素(EPO)被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图45)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以产生经取代的质粒DNA。(EPOK124R)FP5'-GCATGTGGATAGAGCCGTCAGTGC-3'(SEQNo.44),RP5'-GCACTGACGGCTCTATCCACATGC-3'(SEQNo.45);(EPOK167R)FP5'-TGACACTTTCCGCAGACTCTTCCGAGTCTAC-3'(SEQNo.46),RP5'-GTAGACTCGGAAGAGTCTGCGGAAAGTGTCA-3'(SEQNo.47);(EPOK179R)FP5'-CTCCGGGGAAGGCTGAAGCTG-3'(SEQNo.48),RP5'-CAGCTTCAGCCTTCCCCGGAG-3'(SEQNo.49);和(EPOK181R)FP5'-GGAAAGCTGAGGCTGTACACAGG-3'(SEQNo.50),RP5'-CCTGTGTACAGCCTCAGCTTTCC-3'(SEQNo.51)通过使用pcDNA3-myc-EPO作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的4种质粒DNA(表7)。表7赖氨酸(K)残基构建的β-调理素,精氨酸替换赖氨酸124pcDNA3-myc-EPO(K124R)167pcDNA3-myc-EPO(K167R)179pcDNA3-myc-EPO(K179R)181pcDNA3-myc-EPO(K181R)2体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-EPOWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-EPOWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图46)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-EPOWT,pcDNA3-myc-EPO变体(K124R),pcDNA3-myc-EPO变体(K167R),pcDNA3-myc-EPO变体(K179R),pcDNA3-myc-EPO变体(K181R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-EPOWT,pcDNA3-myc-EPO变体(K124R),pcDNA3-myc-EPO变体(K167R),pcDNA3-myc-EPO变体(K179R)和pcDNA3-myc-EPO变体(K181R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图47)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(SantaCruzBiotechnology,sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-EPOWT的结合而形成,且强烈的表明泛素存在的条带被检测到(图46,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,因此出现更加强烈的代表泛素的条带(图46,泳道4)。进一步地,针对pcDNA3-myc-EPO变体(K181R),更少量的泛素被检测到,因为,pcDNA3-myc-EPO变体(K181R)不能与泛素相结合(图46,泳道4)。这些结果说明胰岛素先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估红细胞生成素的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-EPOWT,pcDNA3-myc-EPO变体(K124R),pcDNA3-myc-EPO变体(K167R),pcDNA3-myc-EPO变体(K179R)和pcDNA3-myc-EPO变体(K181R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2小时,4小时和8小时,分别检测每种蛋白的半衰期。结果,观察人红细胞生成素的降解(图48)。人红细胞生成素的半衰期少于4小时,而人红细胞生成素变体(K181R)的半衰期被延长至8小时或者更多,如图48所示。4.EPO和经取代的EPO在细胞中的信号转导有研究报道如果EPO被施用,其通过Erk1/2磷酸化来调控细胞周期进程,因此在缺氧中起作用(JHematolOncol.,6,65,2013)。在这个实验中,我们检测了EPO和经取代的EPO在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入3μgpcDNA3-myc-EPOWT,pcDNA3-myc-EPO变体(K124R),pcDNA3-myc-EPO变体(K167R),pcDNA3-myc-EPO变体(K179R)和pcDNA3-myc-EPO变体(K181R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pcDNA3-myc-EPOWT,pcDNA3-myc-EPO变体(K124R),pcDNA3-myc-EPO变体(K167R),pcDNA3-myc-EPO变体(K179R)和pcDNA3-myc-EPO变体(K181R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗-Erk1/2(9B3,AbfrontierLF-MA0134),抗磷酸化Erk1/2(Thr202/Tyr204,AbfrontierLF-PA0090)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于pcDNA3-myc-EPOWT,pcDNA3-myc-EPO变体(K124R),pcDNA3-myc-EPO变体(K167R),pcDNA3-myc-EPO变体(K179R)和pcDNA3-myc-EPO变体(K181R)在HepG2细胞中,显示出一样或者增加的磷酸化Erk1/2信号转导(图49)。实施例8骨形态生成蛋白2(BMP2)泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.骨形态生成蛋白2(BMP2)表达载体克隆和蛋白表达(1)骨形态生成蛋白2(BMP2)表达载体克隆通过PCR扩增的骨形态生成蛋白2(BMP2)DNA被EcoRI和XhoI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图50,BMP2氨基酸序列:SEQNo.52)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图51)。图50中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图51)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃1分钟30秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图50图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示结合myc的BMP2被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图52)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(BMP2K293R)FP5'-GAAACGCCTTAGGTCCAGCTGTAAGAGAC-3'(SEQNo.53),RP5'-GTCTCTTACAGCTGGACCTAAGGCGTTTC3'(SEQNo.54);(BMP2K297R)FP5'-TTAAGTCCAGCTGTAGGAGACACCCTTTGT-3'(SEQNo.55),RP5'-ACAAAGGGTGTCTCCTACAGCTGGACTTAA-3'(SEQNo.56);(BMP2K355R)FP5'-GTTAACTCTAGGATTCCTAAGGC-3'(SEQNo.57),RP5'-GCCTTAGGAATCCTAGAGTTAAC-3'(SEQNo.58);and(BMP2K383R)FP5'-GGTTGTATTAAGGAACTATCAGGAC-3'(SEQNo.59),RP5'-GTCCTGATAGTTCCTTAATACAACC-3'(SEQNo.60)通过使用pcDNA3-myc-BMP2作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的5种质粒DNA。表8赖氨酸(K)残基构建的BMP2,精氨酸替换赖氨酸293pcDNA3-myc-BMP2(K293R)297pcDNA3-myc-BMP2(K297R)355pcDNA3-myc-BMP2(K355R)383pcDNA3-myc-BMP2(K383R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-BMP2WT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-BMP2WT2μg和pMT123-HA-ubiquitin1μgDNA被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图53)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-BMP2WT,pcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R),pcDNA3-myc-BMP2变体(K383R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-BMP2WT,pcDNA3-myc-BMP2变体(K62R),pcDNA3-myc-BMP2变体(K124R),pcDNA3-myc-BMP2变体(K153R)和pcDNA3-myc-BMP2变体(K158R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图54)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应时,多聚泛素链可以通过泛素与pcDNA3-myc-BMP2WT的结合而形成,因此强烈的代表泛素存在的条带被检测到(图53,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,因此出现更加强烈的代表泛素的条带(图53,泳道4)。进一步地,至于pcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R)和pcDNA3-myc-BMP2变体(K355R),条带不如野生型的强烈,且更少量的泛素被检测到,因为pcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R)不能与泛素相结合(图54,泳道3-5)。这些结果说明BMP2先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估BMP2的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R)和pcDNA3-myc-BMP2变体(K383R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的4小时和8小时,分别检测每种蛋白的半衰期。结果,观察人BMP2的降解(图55)。人BMP2的半衰期少于2小时,而pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R)的半衰期被延长至4小时或者更多,如图55所示。4.BMP2和经取代的BMP2在细胞中的信号转导已经知道在多种骨髓瘤细胞中,骨形态生成蛋白可以抑制STAT3,因此诱导细胞凋亡(Blood,96,2005-2011,2000)。在这个实验中,我们检测了BMP2和经取代的BMP2在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入3μgpcDNA3-myc-BMP2WT,pcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R)和pcDNA3-myc-BMP2变体(K383R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pcDNA3-myc-BMP2WT,pcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R)和pcDNA3-myc-BMP2变体(K383R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,使用以1:1000稀释的抗兔和抗鼠的二抗和包含有抗-STAT3(sc-21876),抗磷酸化STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白质进行显影。结果,相比于野生型,pcDNA3-myc-BMP2变体(K293R),pcDNA3-myc-BMP2变体(K297R),pcDNA3-myc-BMP2变体(K355R),pcDNA3-myc-BMP2变体(K383R)在HepG2细胞中,显示出一样或者增加的磷酸化STAT3信号转导(图56)。实施例9纤维母细胞生长因子-1(FGF-1)泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.纤维母细胞生长因子-1(FGF-1)表达载体克隆和蛋白表达(1)纤维母细胞生长因子-1(FGF-1)表达载体克隆通过PCR扩增的纤维母细胞生长因子-1(FGF-1)DNA被KpnI和XbaI处理,再连接到预先以同样的酶进行消化的pCMV3-C-myc(6.1kb)载体上(图57,FGF-1氨基酸序列:SEQNo.61)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图58)。图57中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图58)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃30秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码蛋白的表达的评估,利用抗-myc抗体(9E10,sc-40)与图57图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的FGF-1被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图59)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(FGF-1K27R)FP5'-AAGAAGCCCAGACTCCTCTAC-3'(SEQNo.62)。RP5'-GTAGAGGAGTCTGGGCTTCTT-3'(SEQNo.63);和(FGF-1K120R)FP5'-CATGCAGAGAGGAATTGGTTT-3'(SEQNo.64)。RP5'-AAACCAATTCCTCTCTGCATG-3'(SEQNo.65)通过使用pCMV3-C-myc-FGF-1作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的2种质粒DNA(表9)。表9赖氨酸(K)残基构建的FGF-1,精氨酸替换赖氨酸27pCMV3-C-myc-FGF-1(K27R)120pCMV3-C-mycFGF-1(K120R)2.体内泛素化分析HEK293T细胞被转染编码pCMV3-C-myc-FGF-1WT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pCMV3-C-myc-FGF-1WT2μg和pMT123-HA-ubiquitin1μgDNA被共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图60)。于是,HEK293T细胞被分别转入编码pCMV3-C-myc-FGF-1WT,pCMV3-C-myc-FGF-1变体(K27R),pCMV3-C-myc-FGF-1变体(K120R),和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpCMV3-C-myc-FGF-1WT,pCMV3-C-myc-FGF-1变体(K27R),pCMV3-C-myc-FGF-1变体(K120R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图61)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应时,多聚泛素链通过泛素与pCMV3-C-myc-FGF-1WT的结合而形成,因此强烈的表明泛素存在的条带被检测到(图60,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,因此出现更加强烈的代表泛素的条带(图60,泳道4)。进一步地,至于pCMV3-C-myc-FGF-1变体(K27R)和pCMV3-C-myc-FGF-1变体(K120R),条带不如野生型的强烈且更少量的泛素被检测到,因为,pCMV3-C-myc-FGF-1变体(K27R)和pCMV3-C-myc-FGF-1变体(K120R)不能与泛素相结合(图61,泳道3和4)。这些结果说明FGF-1先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估FGF-1的半衰期HEK293T细胞被分别的转入2μgpCMV3-C-myc-FGF-1WT,pCMV3-C-myc-FGF-1变体(K27R),pCMV3-C-myc-FGF-1变体(K120R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,于是在抑制剂处理之后的24小时和36小时,分别检测每种蛋白的半衰期。结果,观察人FGF-1的降解(图62)。人FGF-1的半衰期少于1天,而pCMV3-C-myc-FGF-1变体(K27R)和pCMV3-C-myc-FGF-1变体(K120R)的半衰期被延长至1天或者更多,如图62所示。4.FGF-1和经取代的FGF-1在细胞中的信号转导有研究报道当HEK293细胞被重组FGF-1处理时,Erk1/2的磷酸化增加(Nature,513(7518),436-439,2014)。在这个实验中,我们检测了FGF-1和经取代的FGF-1在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入3μgpCMV3-C-myc-FGF-1WT,pCMV3-C-myc-FGF-1变体(K27R)和pCMV3-C-myc-FGF-1变体(K120R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pCMV3-C-myc-FGF-1WT,pCMV3-C-myc-FGF-1变体(K27R)和pCMV3-C-myc-FGF-1变体(K120R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。于是,再使用)以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗-Erk1/2(9B3,AbfrontierLF-MA0134),抗磷酸化Erk1/2(Thr202/Tyr204,AbfrontierLF-PA0090)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于野生型,pCMV3-C-myc-FGF-1变体(K27R)和pCMV3-C-myc-FGF-1变体(K120R)在HepG2细胞中,显示出一样或者增加的磷酸化Erk1/2信号转导(图63)。实施例10瘦蛋白泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.瘦蛋白表达载体克隆和蛋白表达(1)瘦蛋白表达载体克隆通过PCR扩增的瘦蛋白DNA被KpnI和XbaI处理,再连接到预先以同样的酶进行消化的pCMV3-C-myc(6.1kb)载体上(图64,瘦蛋白氨基酸序列:SEQNo.66)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图65)。图64中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图65)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃45秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图64图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的瘦蛋白被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图66)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(LeptinK26R)FP5'-CCCATCCAAAAGGTCCAAGAT-3'(SEQNo.67),RP5'-ATCTTGGACCTTTTGGATGGG-3'(SEQNo.68);(LeptinK32R)FP5'-GATGACACCAAGACCCTCATC-3'(SEQNo.69),RP5'-GATGAGGGTCTTGGTGTCATC-3'(SEQNo.70);(LeptinK36R)FP5'-ACCCTCATCAGGACAATTGTC-3'(SEQNo.71),RP5'-GACAATTGTCCTGATGAGGGT-3'(SEQNo.72);和(LeptinK74R)FP5'-ACCTTATCCAGGATGGACCAG-3'(SEQNo.73),RP5'-CTGGTCCATCCTGGATAAGGT-3'(SEQNo.74)通过使用pCMV3-C-myc-Leptin作为模板,产生有一个或更多个赖氨酸被替换为精氨酸(K→R)的4种质粒DNA(表10)。表102.体内泛素化分析HEK293T细胞被转染编码pCMV3-C-myc-LeptinWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pCMV3-C-myc-LeptinWT6μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,再对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,此后进行免疫沉淀反应分析(图67)。于是,HEK293T细胞被分别转入编码pCMV3-C-myc-LeptinWT,pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K32R),pCMV3-C-myc-Leptin变体(K36R),pCMV3-C-myc-Leptin变体(K74R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与6μgpCMV3-C-myc-LeptinWT,pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K32R),pCMV3-C-myc-Leptin变体(K36R),pCMV3-C-myc-Leptin变体(K74R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图68)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。因此,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pCMV3-C-myc-LeptinWT的结合而形成,因此强烈的表明泛素存在的条带被检测到(图67,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加了,所以出现更加强烈的表明泛素的条带被产生(图67,泳道4)。进一步地,至于pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K36R)和pCMV3-C-myc-Leptin变体(K74R),条带不如野生型的强烈且更少量的泛素被检测到,因为,这些变体不能与泛素相连(图68,泳道3,5和6)。这些结果说明胰岛素先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统被降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估瘦蛋白的半衰期HEK293T细胞被分别的转入6μgpCMV3-C-myc-LeptinWT,pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K32R),pCMV3-C-myc-Leptin变体(K36R)和pCMV3-C-myc-Leptin变体(K74R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2,4和8小时,分别检测每种蛋白的半衰期。结果,观察人瘦蛋白的降解(图69)。人瘦蛋白的半衰期大约为4小时,而pCMV3-C-myc-Leptin变体(K26R)和pCMV3-C-myc-Leptin变体(K36R)的半衰期被延长至8小时或者更多,如图69所示。4.瘦蛋白和经取代的瘦蛋白在细胞中的信号转导有研究报道瘦蛋白增强乳腺癌细胞的AKT磷酸化(CancerBiolTher.,16(8),1220-1230,2015),并且还有报道通过PI3K/AKT信号转导,会刺激子宫癌中癌细胞的生长(IntJOncol.,49(2),847,2016)。在这个实验中,我们检测了瘦蛋白和经取代的瘦蛋白在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入6μgpCMV3-C-myc-LeptinWT,pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K32R),pCMV3-C-myc-Leptin变体(K36R)和pCMV3-C-myc-Leptin变体(K74R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pCMV3-C-myc-LeptinWT,pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K32R),pCMV3-C-myc-Leptin变体(K36R)和pCMV3-C-myc-Leptin变体(K74R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。使用以1:1000稀释的抗兔和抗鼠的二抗和包含有抗myc(9E10,sc-40),抗AKT(H-136,sc-8312),抗磷酸化AKT(S473,CellSignaling9271S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果。相比于对照,pCMV3-C-myc-Leptin变体(K26R),pCMV3-C-myc-Leptin变体(K32R),pCMV3-C-myc-Leptin变体(K36R)和pCMV3-C-myc-Leptin变体(K74R)在HepG2细胞中,显示出一样或者增加的磷酸化AKT信号转导水平(图70)。实施例11血管内皮细胞生长因子A(VEGFA)的泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.血管内皮细胞生长因子A(VEGFA)表达载体克隆和蛋白表达(1)血管内皮细胞生长因子A(VEGFA)表达载体克隆通过PCR扩增的血管内皮细胞生长因子A(VEGFA)DNA被KpnI和XbaI处理,再连接到预先以同样的酶进行消化的pCMV3-C-myc(6.1kb)载体上(图71,VEGFA氨基酸序列:SEQNo.75)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图72)。图71中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图72)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃1分钟(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码蛋白的表达的评估,利用抗-myc抗体(9E10,sc-40)与图71图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的VEGFA被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图73)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(VEGFAK127R)FP5'-TACAGCACAACAGATGTGAATGCAGACC-3'(SEQNo.76),RP5'-GGTCTGCATTCACATCTGTTGTGCTGTA-3'(SEQNo.77);和(VEGFAK180R)FP5'-ATCCGCAGACGTGTAGATGTTCCTGCA-3'(SEQNo.78),RP5'-TGCAGGAACATCTACACGTCTGCGGAT-3'(SEQNo.79)通过使用pCMV3-C-myc-VEGFA作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的2种质粒DNA(表11)。表11赖氨酸(K)残基构建的VEGFA,精氨酸替换赖氨酸127pCMV3-C-myc-VEGFA(K127R)180pCMV3-C-myc-VEGFA(K180R)2.体内泛素化分析HEK293T细胞被转染编码pCMV3-C-myc-VEGFAWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pCMV3-C-myc-VEGFAWT6μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图74)。于是,HEK293T细胞被分别转入编码pCMV3-C-myc-VEGFAWT,pCMV3-C-myc-VEGFA变体(K127R),pCMV3-C-myc-VEGFA变体(K180R),和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与6μgpCMV3-C-myc-VEGFAWT,pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应实验(图75)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应时,多聚泛素链通过泛素与pCMV3-C-myc-VEGFAWT的结合而形成。所以强烈的表明泛素存在的条带被检测到(图74,泳道3和4)。进一步地,当细胞被MG132(蛋白酶体抑制剂,5μg/ml)处理六小时时,多聚泛素链的形成增加,所以出现更加强烈的代表泛素的条带(图74,泳道4)。进一步地,至于pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R),条带不如野生型的强烈且更少量的泛素被检测,因为,这些变体不能与泛素相结合(图75,泳道3和4)。这些结果表明VEGFA先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估VEGFA的半衰期HEK293T细胞被分别的转入6μgpCMV3-C-myc-VEGFAWT,pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2,4和8小时,分别检测每种蛋白的半衰期。结果,观察人VEGFA的降解(图76)。人VEGFA的半衰期少于2小时,而pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R)的半衰期被延长至4小时或者更多,如图76所示。4.VEGFA和经取代的VEGFA在细胞中的信号转导VEGFA与内皮细胞的生长和增殖相关,通过PI3K/Akt/HIF-α通路在肿瘤细胞血管生成中起着功能(Carcinogenesis,34,426-435,2013)。进一步地,VEGF诱导AKT磷酸化(KidneyInt.,68,1648-1659,2005)。在这个实验中,我们检测了VEGFA和经取代的VEGFA在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入6μgpCMV3-C-myc-VEGFAWT,pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pCMV3-C-myc-VEGFAWT,pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗myc(9E10,SantaCruzBiotechnology,sc-40),抗STAT3(sc-21876),抗磷酸化STAT3(Y705,cellsignaling9131S),抗AKT(H-136,sc-8312),抗磷酸化AKT(S473,CellSignaling9271S)和抗β-actin抗体(SantaCruzBiotechnology,sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于野生型,pCMV3-C-myc-VEGFA变体(K127R)和pCMV3-C-myc-VEGFA变体(K180R)在HepG2细胞中,显示出一样或者增加的磷酸化STAT3和磷酸化AKT信号转导(图77)。实施例12食欲刺激激素前体(胃饥饿素/肥胖抑制素前激素原;prepro-GHRL)泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.prepro-GHRL表达载体克隆和蛋白表达(1)prepro-GHRL表达载体克隆通过PCR扩增的prepro-GHRLDNA被KpnI和XbaI处理,再连接到预先以同样的酶进行消化的pCMV3-C-myc(6.1kb)载体上(图78,prepro-GHRL氨基酸序列:SEQNo.80)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图79)。图78中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定(图79)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃30秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图78图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示结合myc的食欲刺激激素前体蛋白被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图80)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(prepro-GHRLK100R)FP5'-GCCCTGGGGAGGTTTCTTCAG-3'(SEQNo.81),RP5'-CTGAAGAAACCTCCCCAGGGC-3'(SEQNo.82)通过使用pCMV3-C-myc-prepro-GHRL作为模板,制备1个赖氨酸被替换为精氨酸(K→R)的质粒DNA(表12)。表12赖氨酸(K)残基构建的prepro-GHRL,精氨酸替换赖氨酸100pCMV3-C-myc-prepro-GHRL(K100R)2.体内泛素化分析HEK293T细胞被转染pCMV3-C-myc-prepro-GHRLWT和pMT123-HA-ubiquitin质粒。为了泛素化水平的分析,pCMV3-C-myc-VEGFAWT6μg和pMT123-HA-ubiquitin1μg共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图81)。于是,HEK293T细胞被分别转入编码pCMV3-C-myc-prepro-GHRLWT,pCMV3-C-myc-prepro-GHRL变体(K100R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与6μgpCMV3-C-myc-prepro-GHRLWT和pCMV3-C-myc-prepro-GHRL变体(K100R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图82)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8and1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链可以通过泛素与pCMV3-C-myc-prepro-GHRLWT的结合而形成。所以强烈的表明泛素存在的条带被检测到(图81,泳道4)。进一步地,当细胞被MG132(蛋白酶体抑制剂,5μg/ml)处理六小时时,多聚泛素链的形成增加,所以出现更加强烈的代表泛素的条带(图81,泳道4)。进一步地,至于pCMV3-C-myc-prepro-GHRL变体(K100R),条带不如野生型的强烈且更少量的泛素被检测到,因为,pCMV3-C-myc-prepro-GHRL变体(K100R)不能与泛素相结合(图82,泳道3)。这些结果说明prepro-GHRL先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估prepro-GHRL的半衰期HEK293T细胞被分别的转入2μgpCMV3-C-myc-prepro-GHRLWT,pCMV3-C-myc-prepro-GHRL变体(K100R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2,4和8小时,再分别检测每种蛋白的半衰期。结果,观察人prepro-GHRL的降解(图76)。人prepro-GHRL的半衰期少于2小时,而pCMV3-C-myc-prepro-GHRL变体(K100R)的半衰期被延长至2小时或者更多,如图83所示。4.prepro-GHRL和经取代的prepro-GHRL在细胞中的信号转导有研究报道食欲刺激激素前体通过生长激素促分泌素受体(GHS-R)而调节细胞生长,且在体内还可以通过钙调控而增强STAT3(MolCellEndocrinol.,285,19-25,2008)。在这个实验中,我们检测了prepro-GHRL和经取代的prepro-GHRL在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入6μgpCMV3-C-myc-prepro-GHRLWT和pCMV3-C-myc-prepro-GHRL变体(K100R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pCMV3-C-myc-prepro-GHRLWT和pCMV3-C-myc-prepro-GHRL变体(K100R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。再使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗myc(9E10,sc-40),抗STAT3(sc-21876),抗磷酸化STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显影。结果,相比于野生型,pCMV3-C-myc-prepro-GHRL变体(K100R)在HepG2细胞中,显示出一样或者增加的磷酸化STAT3信号转导(图84)。实施例13胃饥饿素泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.胃饥饿素表达载体克隆和蛋白表达(1)胃饥饿素表达载体克隆通过PCR扩增的食欲刺激激素(胃饥饿素)DNA被BamHI和XhoII处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图85,胃饥饿素氨基酸序列:SEQNo.83)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图86)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃20秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。图85中下划线标黑的核苷酸序列表明用于PCR并确定克隆位点的引物设定。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图85图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的食欲刺激激素(胃饥饿素)pcDNA3-myc被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图87)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(GhrelinK39RFP)5'-AGTCCAGCAGAGAAGGGAGTCGAAGAAGCCA-3'(SEQNo.84),RP5'-TGGCTTCTTCGACTCCCTTCTCTGCTGGACT-3'(SEQNo.85);(GhrelinK42RFP)5'-AGAAAGGAGTCGAGGAAGCCACCAGCCAAGC-3'(SEQNo.86),RP5'-GCTTGGCTGGTGGCTTCCTCGACTCCTTTCT-3'(SEQNo.87);(GhrelinK43RFP)5'-AGAAAGGAGTCGAAGAGGCCACCAGCCAAGC-3'(SEQNo.88),RP5'-GCTTGGCTGGTGGCCTCTTCGACTCCTTTCT-3'(SEQNo.89);和(GhrelinK47R)FP5'-AAGAAGCCACCAGCCAGGCTGCAGCCCCGA-3'(SEQNo.90),RP5'-TCGGGGCTGCAGCCTGGCTGGTGGCTTCTT-3'(SEQNo.91)通过使用pcDNA3-myc-Ghrelin作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的4种质粒DNA(表13)。表132.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-GhrelinWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-GhrelinWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图88)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-GhrelinWT,pcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R),pcDNA3-myc-Ghrelin变体(K47R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R)和pcDNA3-myc-Ghrelin变体(K47R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图89)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc抗体(9E10,SantaCruzBiotechnology,sc-40)混合,此后,将混合物孵育在4℃并过夜。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-GhrelinWT的结合而形成。所以强烈的表明泛素存在的条带被检测到(图88,泳道3和4)。进一步地,当细胞被MG132(蛋白酶体抑制剂,5μg/ml)处理六小时时,多聚泛素链的形成增加,因此出现更加明强烈的代表泛素的条带(图88,泳道4)。进一步地,至于pcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R)和pcDNA3-myc-Ghrelin变体(K47R),条带不如野生型的明亮,且较少量的泛素被检测到,因为,上述变体不能与泛素相结合(图89,泳道3-6)。这些结果说明prepro-GHRL先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估胃饥饿素的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R)和pcDNA3-myc-Ghrelin变体(K47R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的12,24和36小时,分别检测每种蛋白的半衰期。结果,观察人胃饥饿素的降解(图90)。人胃饥饿素的半衰期少于15小时,而pcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R)和pcDNA3-myc-Ghrelin变体(K47R)的半衰期被延长至36小时或者更多,如图90所示。4.胃饥饿素和经取代的胃饥饿素在细胞中的信号转导有研究报道食欲刺激激素可以通过生长激素促分泌素受体而调节细胞生长,在体内通过钙调控而增加STAT3(MolCellEndocrinol.,285,19-25,2008)。在这个实验中,我们检测了胃饥饿素和经取代的胃饥饿素在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入3μgpcDNA3-myc-GhrelinWT,pcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R)和pcDNA3-myc-Ghrelin变体(K47R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。从分别转染pcDNA3-myc-GhrelinWT,pcDNA3-myc-Ghrelin变体(K39R),pcDNA3-myc-Ghrelin变体(K42R),pcDNA3-myc-Ghrelin变体(K43R)和pcDNA3-myc-Ghrelin变体(K47R)的HepG2细胞中分离得到的蛋白被移至聚偏氟乙烯(PVDF)膜上。再使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗myc(9E10,SantaCruzBiotechnology,sc-40),抗STAT3(sc-21876),抗磷酸化STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(sc-47778)封闭液通过ECL系统对蛋白进行显色。结果,相比于野生型,pcDNA3-myc-Ghrelin变体(K39R)在HepG2细胞中,显示出一样或者增加的磷酸化STAT3信号转导(图91)。实施例14胰高血糖素样肽-1(GLP-1)泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.胰高血糖素样肽-1(GLP-1)表达载体克隆和蛋白表达(1)胰高血糖素样肽-1(GLP-1)表达载体克隆通过PCR扩增的胰高血糖素样肽-1(GLP-1)被EcoRI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图92,GLP-1氨基酸序列:SEQNo.92)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图93)。PCR的条件如下所示:第一步:94℃3分钟(1次循环);第二步:94℃30秒;58℃30秒;72℃20秒(25次循环);第三步:72℃10分钟(1次循环),之后再保存于4℃。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图92图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示结合myc的GLP-1被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图94)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(GLP-1K117R)FP5'-AAGCTGCCAGGGAATTCA-3'(SEQNo.93),RP5'-TGAATTCCCTGGCAGCTT-3'(SEQNo.94);和(GLP-1K125R)FP5'-TTGGCTGGTGAGAGGCC-3'(SEQNo.95),RP5'-GGCCTCTCACCAGCCAA-3'(SEQNo.96)通过使用pcDNA3-myc-GLP-1作为模板,产生有一个或更多个赖氨酸被替换为精氨酸(K→R)的2种质粒DNA(表15)。表15赖氨酸(K)残基构建的GLP-1,精氨酸替换赖氨酸117pcDNA3-myc-GLP-1(K117R)125pcDNA3-myc-GLP-1(K125R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-GLP-1WT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-GLP-1WT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图95)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-GLP-1WT,pcDNA3-myc-GLP-1变体(K117R),pcDNA3-myc-GLP-1变体(K125R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-GLP-1WT,pcDNA3-myc-GLP-1变体(K117R)和pcDNA3-myc-GLP-1变体(K125R)被共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图96)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc抗体(9E10,SantaCruzBiotechnology,sc-40)混合。此后,将混合物孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显色。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-GLP-1WT的结合而形成。所以强烈的表明泛素存在的条带被检测到(图95,泳道3和4)。进一步地,当细胞被MG132(蛋白酶体抑制剂,5μg/ml)处理六小时时,多聚泛素链的形成增加,因此出现更加强烈的代表泛素的条带(图95,泳道4)。进一步地,至于pcDNA3-myc-GLP-1变体(K117R)和pcDNA3-myc-GLP-1变体(K125R),条带不如野生型的明亮,且较少量的泛素被检测到,因为,上述变体不能与泛素相结合(图96,泳道3和4)。这些结果说明GLP-1先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估GLP-1的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-GLP-1WT,pcDNA3-myc-GLP-1变体(K117R)和pcDNA3-myc-GLP-1变体(K125R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2,4和8小时,分别检测每种蛋白的半衰期。结果,观察人GLP-1的降解(图97)。人GLP-1的半衰期大约为2小时,而pcDNA3-myc-GLP-1变体(K117R)和pcDNA3-myc-GLP-1变体(K125R)的半衰期被延长至4小时或者更多,结果如图97所示。4.GLP-1和经取代的GLP-1在细胞中的信号转导GLP-1调控葡萄糖平衡和改善胰岛素的敏感性,因此其可以被用于治疗糖尿病和诱导STAT3活性(BiochemBiophysResCommun.,425(2),304-308,2012)。在这个实验中,我们检测了GLP-1和经取代GLP-1在细胞中的信号转导。首先,HepG2细胞饥饿8小时,再分别转入6μgpcDNA3-myc-GLP-1WT,pcDNA3-myc-GLP-1变体(K117R)和pcDNA3-myc-GLP-1变体(K125R)。转染2天后,细胞内的蛋白被提取并定量。蛋白质印记法被用于分析细胞中的信号转导。为此,从分别转染pcDNA3-myc-GLP-1WT,pcDNA3-myc-GLP-1变体(K117R)和pcDNA3-myc-GLP-1变体(K125R)的HepG2细胞中分离的蛋白被移至聚偏氟乙烯(PVDF)膜上。使用以1:1000稀释的抗兔(辣根过氧化物酶标记的山羊抗兔IgG(H+L),SantaCruzBiotechnology,sc-2004)和抗鼠(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)的二抗和包含有抗myc(9E10,SantaCruzBiotechnology,sc-40),抗STAT3(sc-21876),抗磷酸化STAT3(Y705,cellsignaling9131S)和抗β-actin抗体(sc-47778)封闭液。结果,相比于野生型,pcDNA3-myc-GLP-1变体(K117R)在HepG2细胞中,显示出一样或者增加的磷酸化STAT3信号转导(图98)。实施例15IgG重链泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.IgG重链表达载体克隆和蛋白表达(1)IgG重链表达载体克隆IgG重链(HC)的DNA序列通过Roche的EP1308455B9(一种包含抗HER2抗体的组合物,p.24)中的描述而合成,且进一步地优化使其在哺乳动物细胞中很好地表达。于是,通过PCR扩增的IgG重链(HC)DNA被EcoRI和XhoI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图99,IgG重链氨基酸序列:SEQNo.97)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图100)。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图99图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的IgG重链(HC)被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图101)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(IgGHCK235R)FP5'-ACAAAGGTGGACAGGAAGGTGGAGCCCAAG-3'(SEQNo.98),RP5'-CTTGGGCTCCACCTTCCTGTCCACCTTTGT-3'(SEQNo.99);(IgGHCK344R)FP5'-GAGTATAAGTGCAGGGTGTCCAATAAGGCCCTGC-3'(SEQNo.100),RP5'-GCAGGGCCTTATTGGACACCCTGCACTTATACTC-3'(SEQNo.101);和(IgGHCK431R)FP5'-CTTTCTGTATAGCAGGCTGACCGTGGATAAGTCC-3'(SEQNo.102),RP5'-GGACTTATCCACGGTCAGCCTGCTATACAGAAAG-3'(SEQNo.103)通过使用pcDNA3-myc-IgGHC作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的3种质粒DNA(表14)。表14赖氨酸(K)残基构建的IgG重链,精氨酸替换赖氨酸235pcDNA3-myc-IgGHC(K235R)344pcDNA3-myc-IgGHC(K344R)431pcDNA3-myc-IgGHC(K431R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3-myc-IgGHCWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-IgGHCWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应分析(图102)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-IgGHCWT,pcDNA3-myc-IgGHC变体(K235R),pcDNA3-myc-IgGHC变体(K344R),pcDNA3-myc-IgGHC变体(K431R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-IgGHCWT,pcDNA3-myc-IgGHC变体(K235R),pcDNA3-myc-IgGHC变体(K344R)和pcDNA3-myc-IgGHC变体(K431R)被共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图103)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合。此后,将混合物孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。因此,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-IgGHCWT结合而形成,因此强烈的表明泛素存在的条带被检测到(图102,泳道3和4)。进一步地,当细胞被MG132(蛋白酶体抑制剂,5μg/ml)处理六小时时,多聚泛素链的形成增加,因此出现更加强烈的代表泛素的条带(图102,泳道4)。进一步地,至于pcDNA3-myc-IgGHC变体(K431R),条带不如野生型的明亮且较少量的泛素被检测到,因为变体不能与泛素相结合(图103,泳道5)。这些结果说明IgG-HC先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估IgG-HC的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-IgGHCWT,pcDNA3-myc-IgGHC变体(K235R),pcDNA3-myc-IgGHC变体(K344R)和pcDNA3-myc-IgGHC变体(K431R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2,4和8小时,分别检测每种蛋白的半衰期。结果,观察人IgGHC的降解(图104)。人IgGHC的半衰期少于2小时,而pcDNA3-myc-IgGHC变体(K431R)的半衰期被延长至4小时或者更多,如图104所示。实施例16IgG轻链(LC)泛素化和半衰期延长的分析,以及细胞中的信号转导的分析1.IgG轻链(LC)表达载体克隆和蛋白表达(1)IgG轻链(LC)表达载体克隆IgG轻链(LC)的DNA序列通过Roche的EP1308455B9(一种包含抗HER2抗体的组合物,p.23)中的描述而合成,且进一步地优化使其在哺乳动物细胞中很好地表达。通过PCR扩增的IgG轻链(LC)DNA被EcoRI和XhoI处理,再连接到预先以同样的酶进行消化的pcDNA3-myc(5.6kb)载体上(图105,IgG轻链氨基酸序列:SEQNo.104)。于是,在克隆载体的限制性内切酶消化之后,通过琼脂糖凝胶电泳确认DNA插入片段的存在(图106)。为了克隆DNA编码的蛋白表达的评估,利用抗-myc抗体(9E10,sc-40)与图105图谱的pcDNA3-myc载体上的myc的结合而进行蛋白质印迹法。蛋白质印记的结果显示与myc结合的IgG轻链(LC)被很好地表达。肌动蛋白作为标准确保蛋白质上样量的是合适的(图107)。(2)赖氨酸(Lysine,K)残基的取代使用定点突变的方法将赖氨酸残基替换为精氨酸(Arginine,R)。下述引物设定被用于PCR以制备经取代的质粒DNA。(IgGLCK67R)FP5'-CCTGGCAAGGCCCCAAGGCTGCTGATCTAC-3'(SEQNo.105),RP5'-GTAGATCAGCAGCCTTGGGGCCTTGCCAGG-3'(SEQNo.106);(IgGLCK129R)FP5'-ACAAAGGTGGAGATCAGGAGGACCGTGGCC-3'(SEQNo.107),RP5'-GGCCACGGTCCTCCTGATCTCCACCTTTGT-3'(SEQNo.108);and(IgGLCK171R)FP5'-GCCAAGGTGCAGTGGAGGGTGGATAACGCC-3'(SEQNo.109),RP5'-GGCGTTATCCACCCTCCACTGCACCTTGGC-3'(SEQNo.110)通过使用pcDNA3-myc-IgGLC作为模板,制备有一个或更多个赖氨酸被替换为精氨酸(K→R)的3种质粒DNA(表16)。表16赖氨酸(K)残基构建的IgG轻链,精氨酸替换赖氨酸67pcDNA3-myc-IgGLC(K67R)129pcDNA3-myc-IgGLC(K129R)171pcDNA3-myc-IgGLC(K171R)2.体内泛素化分析HEK293T细胞被转染编码pcDNA3.1-6myc-IgGLCWT和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pcDNA3-myc-IgGLCWT2μg和pMT123-HA-ubiquitin1μg被共转入细胞中。转染后24小时,对细胞进行MG132(蛋白酶体抑制剂,5μg/ml)处理6小时,再进行免疫沉淀反应(图108)。于是,HEK293T细胞被分别转入编码pcDNA3-myc-IgGLCWT,pcDNA3-myc-IgGLC变体(K67R),pcDNA3-myc-IgGLC变体(K129R),pcDNA3-myc-IgGLC变体(K171R)和pMT123-HA-ubiquitin的质粒。为了泛素化水平的分析,pMT123-HA-ubiquitin1μg分别与2μgpcDNA3-myc-IgGLCWT,pcDNA3-myc-IgGLC变体(K67R),pcDNA3-myc-IgGLC变体(K129R)和pcDNA3-myc-IgGLC变体(K171R)共转入细胞中。接下来,转染后24小时,进行免疫沉淀反应(图109)。通过免疫沉淀反应获得的蛋白样品被溶于缓冲液中,其包含(1%TritonX,150mMNaCl,50mMTris-HCl,pH8和1mMPMSF(苯甲基硫磺酰)),再将其与抗myc(9E10)一抗(SantaCruzBiotechnology,sc-40)混合,此后,混合物被孵育在4℃并过夜。免疫沉淀物可以通过与A/G珠在4℃反应2小时而分离。随后,分离的免疫沉淀物用缓冲液清洗两次。在与2X的SDS缓冲液混合并加热至100℃,7分钟之后,蛋白样品通过SDS凝胶电泳分离。分离的蛋白被转移到聚偏氟乙烯(PVDF)膜上,再使用以1:1000稀释的抗小鼠的二抗(过氧化物酶标记的小鼠IgG(H+L),KPL,074-1806)和包含有抗myc抗体(9E10,sc-40),抗HA(sc-7392)和抗β-actin抗体(sc-47778)封闭液通过ECL系统进行显影。结果,当使用抗myc抗体(9E10,sc-40)进行免疫沉淀反应实验时,多聚泛素链通过泛素与pcDNA3-myc-IgGLCWT的结合而形成,且强烈的表明泛素存在的条带被检测到(图108,泳道3和4)。进一步地,当细胞被MG132处理六小时时,多聚泛素链的形成增加,因此出现更加强烈的代表泛素的条带(图108,泳道4)。进一步地,至于pcDNA3-myc-IgGLC变体(K171R),条带不如野生型的明亮且更少量的泛素被检测到,因为上述变体不能与泛素相结合(图109,泳道5)。这些结果说明IgG-LC先结合泛素,再通过形成的多聚泛素链被泛素-蛋白酶体系统降解。3.使用蛋白酶合成抑制剂环十二碳三烯(CHX)评估IgG-LC的半衰期HEK293T细胞被分别的转入2μgpcDNA3-myc-IgGLCWT,pcDNA3-myc-IgGLC变体(K67R),pcDNA3-myc-IgGLC变体(K129R),pcDNA3-myc-IgGLC变体(K171R)。转染48小时后,细胞用蛋白酶合成抑制剂环十二碳三烯(CHX)(Sigma-Aldrich,100μg/ml)处理,在抑制剂处理之后的2,4和8小时,分别检测每种蛋白的半衰期。结果,观察人IgGLC的降解(图110)。人IgGLC的半衰期少于1小时,而pcDNA3-myc-IgGLC变体(K171R)的半衰期被被延长至2小时或者更多,如图110所示。实用性因为本发明的变体蛋白具有延长的半衰期,本发明可以被用于开发蛋白或者(多)肽治疗性试剂。序列表<110>UBI蛋白公司<120>用于延长蛋白质半衰期的方法<130>UBPRN16P01WO<150>KR10-2015-0160728<151>2015-11-16<160>110<170>KoPatentIn3.0<210>1<211>198<212>PRT<213>人工序列<220><223>人β-调理素<400>1MetProValProAlaLeuCysLeuLeuTrpAlaLeuAlaMetValThr151015ArgProAlaSerAlaAlaProMetGlyGlyProGluLeuAlaGlnHis202530GluGluLeuThrLeuLeuPheHisGlyThrLeuGlnLeuGlyGlnAla354045LeuAsnGlyValTyrArgThrThrGluGlyArgLeuThrLysAlaArg505560AsnSerLeuGlyLeuTyrGlyArgThrIleGluLeuLeuGlyGlnGlu65707580ValSerArgGlyArgAspAlaAlaGlnGluLeuArgAlaSerLeuLeu859095GluThrGlnMetGluGluAspIleLeuGlnLeuGlnAlaGluAlaThr100105110AlaGluValLeuGlyGluValAlaGlnAlaGlnLysValLeuArgAsp115120125SerValGlnArgLeuGluValGlnLeuArgSerAlaTrpLeuGlyPro130135140AlaTyrArgGluPheGluValLeuLysAlaHisAlaAspLysGlnSer145150155160HisIleLeuTrpAlaLeuThrGlyHisValGlnArgGlnArgArgGlu165170175MetValAlaGlnGlnHisArgLeuArgGlnIleGlnGluArgLeuHis180185190ThrAlaAlaLeuProAla195<210>2<211>25<212>DNA<213>人工序列<220><223>人β-调理素<400>2agggacggctgacaagggccaggaa25<210>3<211>26<212>DNA<213>人工序列<220><223>人β-调理素<400>3ccaggctgttcctggcccttgtcagc26<210>4<211>25<212>DNA<213>人工序列<220><223>人β-调理素<400>4ggcacagagggtgctacgggacagc25<210>5<211>25<212>DNA<213>人工序列<220><223>人β-调理素<400>5cgtagcaccctctgtgcctgggcca25<210>6<211>26<212>DNA<213>人工序列<220><223>人β-调理素<400>6gaatttgaggtcttaagggctcacgc26<210>7<211>27<212>DNA<213>人工序列<220><223>人β-调理素<400>7cttgtcagcgtgagcccttaagacctc27<210>8<211>26<212>DNA<213>人工序列<220><223>人β-调理素<400>8gctcacgctgacaggcagagccacat26<210>9<211>27<212>DNA<213>人工序列<220><223>人β-调理素<400>9ccataggatgtggctctgcctgtcagc27<210>10<211>217<212>PRT<213>人工序列<220><223>人生长激素<400>10MetAlaThrGlySerArgThrSerLeuLeuLeuAlaPheGlyLeuLeu151015CysLeuProTrpLeuGlnGluGlySerAlaPheProThrIleProLeu202530SerArgLeuPheAspAsnAlaMetLeuArgAlaHisArgLeuHisGln354045LeuAlaPheAspThrTyrGlnGluPheGluGluAlaTyrIleProLys505560GluGlnLysTyrSerPheLeuGlnAsnProGlnThrSerLeuCysPhe65707580SerGluSerIleProThrProSerAsnArgGluGluThrGlnGlnLys859095SerAsnLeuGluLeuLeuArgIleSerLeuLeuLeuIleGlnSerTrp100105110LeuGluProValGlnPheLeuArgSerValPheAlaAsnSerLeuVal115120125TyrGlyAlaSerAspSerAsnValTyrAspLeuLeuLysAspLeuGlu130135140GluGlyIleGlnThrLeuMetGlyArgLeuGluAspGlySerProArg145150155160ThrGlyGlnIlePheLysGlnThrTyrSerLysPheAspThrAsnSer165170175HisAsnAspAspAlaLeuLeuLysAsnTyrGlyLeuLeuTyrCysPhe180185190ArgLysAspMetAspLysValGluThrPheLeuArgIleValGlnCys195200205ArgSerValGluGlySerCysGlyPhe210215<210>11<211>24<212>DNA<213>人工序列<220><223>人生长激素<400>11ccaaaggaacagaggtattcattc24<210>12<211>24<212>DNA<213>人工序列<220><223>人生长激素<400>12caggaatgaatacctctgttcctt24<210>13<211>21<212>DNA<213>人工序列<220><223>人生长激素<400>13gacctcctaagggacctagag21<210>14<211>21<212>DNA<213>人工序列<220><223>人生长激素<400>14ctctaggtcccttaggaggtc21<210>15<211>21<212>DNA<213>人工序列<220><223>人生长激素<400>15cagatcttcaggcagacctac21<210>16<211>21<212>DNA<213>人工序列<220><223>人生长激素<400>16gtaggtctgcctgaagatctg21<210>17<211>110<212>PRT<213>人工序列<220><223>人胰岛素<400>17MetAlaLeuTrpMetArgLeuLeuProLeuLeuAlaLeuLeuAlaLeu151015TrpGlyProAspProAlaAlaAlaPheValAsnGlnHisLeuCysGly202530SerHisLeuValGluAlaLeuTyrLeuValCysGlyGluArgGlyPhe354045PheTyrThrProLysThrArgArgGluAlaGluAspLeuGlnValGly505560GlnValGluLeuGlyGlyGlyProGlyAlaGlySerLeuGlnProLeu65707580AlaLeuGluGlySerLeuGlnLysArgGlyIleValGluGlnCysCys859095ThrSerIleCysSerLeuTyrGlnLeuGluAsnTyrCysAsn100105110<210>18<211>25<212>DNA<213>人工序列<220><223>人胰岛素<400>18ggcttcttctacacacccaggaccc25<210>19<211>24<212>DNA<213>人工序列<220><223>人胰岛素<400>19ctcccggcgggtcctgggtgtgta24<210>20<211>23<212>DNA<213>人工序列<220><223>人胰岛素<400>20tccctgcagaggcgtggcattgt23<210>21<211>27<212>DNA<213>人工序列<220><223>人胰岛素<400>21ttgttccacaatgccacgcctctgcag27<210>22<211>188<212>PRT<213>人工序列<220><223>人干扰素α<400>22MetAlaLeuThrPheAlaLeuLeuValAlaLeuLeuValLeuSerCys151015LysSerSerCysSerValGlyCysAspLeuProGlnThrHisSerLeu202530GlySerArgArgThrLeuMetLeuLeuAlaGlnMetArgArgIleSer354045LeuPheSerCysLeuLysAspArgHisAspPheGlyPheProGlnGlu505560GluPheGlyAsnGlnPheGlnLysAlaGluThrIleProValLeuHis65707580GluMetIleGlnGlnIlePheAsnLeuPheSerThrLysAspSerSer859095AlaAlaTrpAspGluThrLeuLeuAspLysPheTyrThrGluLeuTyr100105110GlnGlnLeuAsnAspLeuGluAlaCysValIleGlnGlyValGlyVal115120125ThrGluThrProLeuMetLysGluAspSerIleLeuAlaValArgLys130135140TyrPheGlnArgIleThrLeuTyrLeuLysGluLysLysTyrSerPro145150155160CysAlaTrpGluValValArgAlaGluIleMetArgSerPheSerLeu165170175SerThrAsnLeuGlnGluSerLeuArgSerLysGlu180185<210>23<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>23cttcagcacaagggactcatc21<210>24<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>24cagatgagtcccttgtgctga21<210>25<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>25ctcctagacagattctacact21<210>26<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>26agtgtagaatctgtctaggag21<210>27<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>27gctgtgaggagatacttccaa21<210>28<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>28ttggaagtatctcctcacagc21<210>29<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>29ctctatctgagagagaagaaa21<210>30<211>21<212>DNA<213>人工序列<220><223>人干扰素α<400>30tttcttctctctcagatagag21<210>31<211>207<212>PRT<213>人工序列<220><223>人G-CSF<400>31MetAlaGlyProAlaThrGlnSerProMetLysLeuMetAlaLeuGln151015LeuLeuLeuTrpHisSerAlaLeuTrpThrValGlnGluAlaThrPro202530LeuGlyProAlaSerSerLeuProGlnSerPheLeuLeuLysCysLeu354045GluGlnValArgLysIleGlnGlyAspGlyAlaAlaLeuGlnGluLys505560LeuValSerGluCysAlaThrTyrLysLeuCysHisProGluGluLeu65707580ValLeuLeuGlyHisSerLeuGlyIleProTrpAlaProLeuSerSer859095CysProSerGlnAlaLeuGlnLeuAlaGlyCysLeuSerGlnLeuHis100105110SerGlyLeuPheLeuTyrGlnGlyLeuLeuGlnAlaLeuGluGlyIle115120125SerProGluLeuGlyProThrLeuAspThrLeuGlnLeuAspValAla130135140AspPheAlaThrThrIleTrpGlnGlnMetGluGluLeuGlyMetAla145150155160ProAlaLeuGlnProThrGlnGlyAlaMetProAlaPheAlaSerAla165170175PheGlnArgArgAlaGlyGlyValLeuValAlaSerHisLeuGlnSer180185190PheLeuGluValSerTyrArgValLeuArgHisLeuAlaGlnPro195200205<210>32<211>24<212>DNA<213>人工序列<220><223>人G-CSF<400>32agcttcctgctcaggtgcttagag24<210>33<211>24<212>DNA<213>人工序列<220><223>人G-CSF<400>33ttgctctaagcacctgagcaggaa24<210>34<211>24<212>DNA<213>人工序列<220><223>人G-CSF<400>34tgtgccacctacaggctgtgccac24<210>35<211>24<212>DNA<213>人工序列<220><223>人G-CSF<400>35ggggtggcacagcctgtaggtggc24<210>36<211>187<212>PRT<213>人工序列<220><223>人干扰素β<400>36MetThrAsnLysCysLeuLeuGlnIleAlaLeuLeuLeuCysPheSer151015ThrThrAlaLeuSerMetSerTyrAsnLeuLeuGlyPheLeuGlnArg202530SerSerAsnPheGlnCysGlnLysLeuLeuTrpGlnLeuAsnGlyArg354045LeuGluTyrCysLeuLysAspArgMetAsnPheAspIleProGluGlu505560IleLysGlnLeuGlnGlnPheGlnLysGluAspAlaAlaLeuThrIle65707580TyrGluMetLeuGlnAsnIlePheAlaIlePheArgGlnAspSerSer859095SerThrGlyTrpAsnGluThrIleValGluAsnLeuLeuAlaAsnVal100105110TyrHisGlnIleAsnHisLeuLysThrValLeuGluGluLysLeuGlu115120125LysGluAspPheThrArgGlyLysLeuMetSerSerLeuHisLeuLys130135140ArgTyrTyrGlyArgIleLeuHisTyrLeuLysAlaLysGluTyrSer145150155160HisCysAlaTrpThrIleValArgValGluIleLeuArgAsnPheTyr165170175PheIleAsnArgLeuThrGlyTyrLeuArgAsn180185<210>37<211>21<212>DNA<213>人工序列<220><223>人干扰素β<400>37cagtgtcagaggctcctgtgg21<210>38<211>21<212>DNA<213>人工序列<220><223>人干扰素β<400>38ccacaggagcctctgacactg21<210>39<211>21<212>DNA<213>人工序列<220><223>人干扰素β<400>39ctggaagaaagactggagaaa21<210>40<211>21<212>DNA<213>人工序列<220><223>人干扰素β<400>40tttctccagtctttcttccag21<210>41<211>21<212>DNA<213>人工序列<220><223>人干扰素β<400>41cattacctgagggccaaggag21<210>42<211>21<212>DNA<213>人工序列<220><223>人干扰素β<400>42ctccttggccctcaggtaatg21<210>43<211>193<212>PRT<213>人工序列<220><223>人红细胞生成素<400>43MetGlyValHisGluCysProAlaTrpLeuTrpLeuLeuLeuSerLeu151015LeuSerLeuProLeuGlyLeuProValLeuGlyAlaProProArgLeu202530IleCysAspSerArgValLeuGluArgTyrLeuLeuGluAlaLysGlu354045AlaGluAsnIleThrThrGlyCysAlaGluHisCysSerLeuAsnGlu505560AsnIleThrValProAspThrLysValAsnPheTyrAlaTrpLysArg65707580MetGluValGlyGlnGlnAlaValGluValTrpGlnGlyLeuAlaLeu859095LeuSerGluAlaValLeuArgGlyGlnAlaLeuLeuValAsnSerSer100105110GlnProTrpGluProLeuGlnLeuHisValAspLysAlaValSerGly115120125LeuArgSerLeuThrThrLeuLeuArgAlaLeuGlyAlaGlnLysGlu130135140AlaIleSerProProAspAlaAlaSerAlaAlaProLeuArgThrIle145150155160ThrAlaAspThrPheArgLysLeuPheArgValTyrSerAsnPheLeu165170175ArgGlyLysLeuLysLeuTyrThrGlyGluAlaCysArgThrGlyAsp180185190Arg<210>44<211>24<212>DNA<213>人工序列<220><223>人红细胞生成素<400>44gcatgtggatagagccgtcagtgc24<210>45<211>24<212>DNA<213>人工序列<220><223>人红细胞生成素<400>45gcactgacggctctatccacatgc24<210>46<211>31<212>DNA<213>人工序列<220><223>人红细胞生成素<400>46tgacactttccgcagactcttccgagtctac31<210>47<211>31<212>DNA<213>人工序列<220><223>人红细胞生成素<400>47gtagactcggaagagtctgcggaaagtgtca31<210>48<211>21<212>DNA<213>人工序列<220><223>人红细胞生成素<400>48ctccggggaaggctgaagctg21<210>49<211>21<212>DNA<213>人工序列<220><223>人红细胞生成素<400>49cagcttcagccttccccggag21<210>50<211>23<212>DNA<213>人工序列<220><223>人红细胞生成素<400>50ggaaagctgaggctgtacacagg23<210>51<211>23<212>DNA<213>人工序列<220><223>人红细胞生成素<400>51cctgtgtacagcctcagctttcc23<210>52<211>396<212>PRT<213>人工序列<220><223>人骨形态生成蛋白-2<400>52MetValAlaGlyThrArgCysLeuLeuAlaLeuLeuLeuProGlnVal151015LeuLeuGlyGlyAlaAlaGlyLeuValProGluLeuGlyArgArgLys202530PheAlaAlaAlaSerSerGlyArgProSerSerGlnProSerAspGlu354045ValLeuSerGluPheGluLeuArgLeuLeuSerMetPheGlyLeuLys505560GlnArgProThrProSerArgAspAlaValValProProTyrMetLeu65707580AspLeuTyrArgArgHisSerGlyGlnProGlySerProAlaProAsp859095HisArgLeuGluArgAlaAlaSerArgAlaAsnThrValArgSerPhe100105110HisHisGluGluSerLeuGluGluLeuProGluThrSerGlyLysThr115120125ThrArgArgPhePhePheAsnLeuSerSerIleProThrGluGluPhe130135140IleThrSerAlaGluLeuGlnValPheArgGluGlnMetGlnAspAla145150155160LeuGlyAsnAsnSerSerPheHisHisArgIleAsnIleTyrGluIle165170175IleLysProAlaThrAlaAsnSerLysPheProValThrArgLeuLeu180185190AspThrArgLeuValAsnGlnAsnAlaSerArgTrpGluSerPheAsp195200205ValThrProAlaValMetArgTrpThrAlaGlnGlyHisAlaAsnHis210215220GlyPheValValGluValAlaHisLeuGluGluLysGlnGlyValSer225230235240LysArgHisValArgIleSerArgSerLeuHisGlnAspGluHisSer245250255TrpSerGlnIleArgProLeuLeuValThrPheGlyHisAspGlyLys260265270GlyHisProLeuHisLysArgGluLysArgGlnAlaLysHisLysGln275280285ArgLysArgLeuLysSerSerCysLysArgHisProLeuTyrValAsp290295300PheSerAspValGlyTrpAsnAspTrpIleValAlaProProGlyTyr305310315320HisAlaPheTyrCysHisGlyGluCysProPheProLeuAlaAspHis325330335LeuAsnSerThrAsnHisAlaIleValGlnThrLeuValAsnSerVal340345350AsnSerLysIleProLysAlaCysCysValProThrGluLeuSerAla355360365IleSerMetLeuTyrLeuAspGluAsnGluLysValValLeuLysAsn370375380TyrGlnAspMetValValGluGlyCysGlyCysArg385390395<210>53<211>29<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>53gaaacgccttaggtccagctgtaagagac29<210>54<211>29<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>54gtctcttacagctggacctaaggcgtttc29<210>55<211>30<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>55ttaagtccagctgtaggagacaccctttgt30<210>56<211>30<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>56acaaagggtgtctcctacagctggacttaa30<210>57<211>23<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>57gttaactctaggattcctaaggc23<210>58<211>23<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>58gccttaggaatcctagagttaac23<210>59<211>25<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>59ggttgtattaaggaactatcaggac25<210>60<211>25<212>DNA<213>人工序列<220><223>人骨形态生成蛋白-2<400>60gtcctgatagttccttaatacaacc25<210>61<211>155<212>PRT<213>人工序列<220><223>人纤维母细胞生长因子-1<400>61MetAlaGluGlyGluIleThrThrPheThrAlaLeuThrGluLysPhe151015AsnLeuProProGlyAsnTyrLysLysProLysLeuLeuTyrCysSer202530AsnGlyGlyHisPheLeuArgIleLeuProAspGlyThrValAspGly354045ThrArgAspArgSerAspGlnHisIleGlnLeuGlnLeuSerAlaGlu505560SerValGlyGluValTyrIleLysSerThrGluThrGlyGlnTyrLeu65707580AlaMetAspThrAspGlyLeuLeuTyrGlySerGlnThrProAsnGlu859095GluCysLeuPheLeuGluArgLeuGluGluAsnHisTyrAsnThrTyr100105110IleSerLysLysHisAlaGluLysAsnTrpPheValGlyLeuLysLys115120125AsnGlySerCysLysArgGlyProArgThrHisTyrGlyGlnLysAla130135140IleLeuPheLeuProLeuProValSerSerAsp145150155<210>62<211>21<212>DNA<213>人工序列<220><223>人纤维母细胞生长因子-1<400>62aagaagcccagactcctctac21<210>63<211>21<212>DNA<213>人工序列<220><223>人纤维母细胞生长因子-1<400>63gtagaggagtctgggcttctt21<210>64<211>21<212>DNA<213>人工序列<220><223>人纤维母细胞生长因子-1<400>64catgcagagaggaattggttt21<210>65<211>21<212>DNA<213>人工序列<220><223>人纤维母细胞生长因子-1<400>65aaaccaattcctctctgcatg21<210>66<211>167<212>PRT<213>人工序列<220><223>人瘦蛋白<400>66MetHisTrpGlyThrLeuCysGlyPheLeuTrpLeuTrpProTyrLeu151015PheTyrValGlnAlaValProIleGlnLysValGlnAspAspThrLys202530ThrLeuIleLysThrIleValThrArgIleAsnAspIleSerHisThr354045GlnSerValSerSerLysGlnLysValThrGlyLeuAspPheIlePro505560GlyLeuHisProIleLeuThrLeuSerLysMetAspGlnThrLeuAla65707580ValTyrGlnGlnIleLeuThrSerMetProSerArgAsnValIleGln859095IleSerAsnAspLeuGluAsnLeuArgAspLeuLeuHisValLeuAla100105110PheSerLysSerCysHisLeuProTrpAlaSerGlyLeuGluThrLeu115120125AspSerLeuGlyGlyValLeuGluAlaSerGlyTyrSerThrGluVal130135140ValAlaLeuSerArgLeuGlnGlySerLeuGlnAspMetLeuTrpGln145150155160LeuAspLeuSerProGlyCys165<210>67<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>67cccatccaaaaggtccaagat21<210>68<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>68atcttggaccttttggatggg21<210>69<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>69gatgacaccaagaccctcatc21<210>70<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>70gatgagggtcttggtgtcatc21<210>71<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>71accctcatcaggacaattgtc21<210>72<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>72gacaattgtcctgatgagggt21<210>73<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>73accttatccaggatggaccag21<210>74<211>21<212>DNA<213>人工序列<220><223>人瘦蛋白<400>74ctggtccatcctggataaggt21<210>75<211>209<212>PRT<213>人工序列<220><223>人血管内皮生长因子A<400>75MetAsnPheLeuLeuSerTrpValHisTrpSerLeuAlaLeuLeuLeu151015TyrLeuHisHisAlaLysTrpSerGlnAlaAlaProMetAlaGluGly202530GlyGlyGlnAsnHisHisGluValValLysPheMetAspValTyrGln354045ArgSerTyrCysHisProIleGluThrLeuValAspIlePheGlnGlu505560TyrProAspGluIleGluTyrIlePheLysProSerCysValProLeu65707580MetArgCysGlyGlyCysCysAsnAspGluGlyLeuGluCysValPro859095ThrGluGluSerAsnIleThrMetGlnIleMetArgIleLysProHis100105110GlnGlyGlnHisIleGlyGluMetSerPheLeuGlnHisAsnLysCys115120125GluCysArgProLysLysAspArgAlaArgGlnGluLysLysSerVal130135140ArgGlyLysGlyLysGlyGlnLysArgLysArgLysLysSerArgPro145150155160CysGlyProCysSerGluArgArgLysHisLeuPheValGlnAspPro165170175GlnThrCysLysCysSerCysLysAsnThrAspSerArgCysLysAla180185190ArgGlnLeuGluLeuAsnGluArgThrCysArgCysAspLysProArg195200205Arg<210>76<211>28<212>DNA<213>人工序列<220><223>人血管内皮生长因子A<400>76tacagcacaacagatgtgaatgcagacc28<210>77<211>28<212>DNA<213>人工序列<220><223>人血管内皮生长因子A<400>77ggtctgcattcacatctgttgtgctgta28<210>78<211>27<212>DNA<213>人工序列<220><223>人血管内皮生长因子A<400>78atccgcagacgtgtagatgttcctgca27<210>79<211>27<212>DNA<213>人工序列<220><223>人血管内皮生长因子A<400>79tgcaggaacatctacacgtctgcggat27<210>80<211>117<212>PRT<213>人工序列<220><223>人prepro-GHRL<400>80MetProSerProGlyThrValCysSerLeuLeuLeuLeuGlyMetLeu151015TrpLeuAspLeuAlaMetAlaGlySerSerPheLeuSerProGluHis202530GlnArgValGlnGlnArgLysGluSerLysLysProProAlaLysLeu354045GlnProArgAlaLeuAlaGlyTrpLeuArgProGluAspGlyGlyGln505560AlaGluGlyAlaGluAspGluMetGluValArgPheAsnAlaProPhe65707580AspValGlyIleLysLeuSerGlyValGlnTyrGlnGlnHisSerGln859095AlaLeuGlyLysPheLeuGlnAspIleLeuTrpGluGluAlaLysGlu100105110AlaProAlaAspLys115<210>81<211>21<212>DNA<213>人工序列<220><223>人prepro-GHRL<400>81gccctggggaggtttcttcag21<210>82<211>21<212>DNA<213>人工序列<220><223>人prepro-GHRL<400>82ctgaagaaacctccccagggc21<210>83<211>28<212>PRT<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>83GlySerSerPheLeuSerProGluHisGlnArgValGlnGlnArgLys151015GluSerLysLysProProAlaLysLeuGlnProArg2025<210>84<211>31<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>84agtccagcagagaagggagtcgaagaagcca31<210>85<211>31<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>85tggcttcttcgactcccttctctgctggact31<210>86<211>31<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>86agaaaggagtcgaggaagccaccagccaagc31<210>87<211>31<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>87gcttggctggtggcttcctcgactcctttct31<210>88<211>31<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>88agaaaggagtcgaagaggccaccagccaagc31<210>89<211>31<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>89gcttggctggtggcctcttcgactcctttct31<210>90<211>30<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>90aagaagccaccagccaggctgcagccccga30<210>91<211>30<212>DNA<213>人工序列<220><223>人食欲刺激激素(胃饥饿素)<400>91tcggggctgcagcctggctggtggcttctt30<210>92<211>31<212>PRT<213>人工序列<220><223>人胰高血糖素样肽-1(GLP-1)<400>92HisAlaGluGlyThrPheThrSerAspValSerSerTyrLeuGluGly151015GlnAlaAlaLysGluPheIleAlaTrpLeuValLysGlyArgGly202530<210>93<211>18<212>DNA<213>人工序列<220><223>人胰高血糖素样肽-1(GLP-1)<400>93aagctgccagggaattca18<210>94<211>18<212>DNA<213>人工序列<220><223>人胰高血糖素样肽-1(GLP-1)<400>94tgaattccctggcagctt18<210>95<211>17<212>DNA<213>人工序列<220><223>人胰高血糖素样肽-1(GLP-1)<400>95ttggctggtgagaggcc17<210>96<211>17<212>DNA<213>人工序列<220><223>人胰高血糖素样肽-1(GLP-1)<400>96ggcctctcaccagccaa17<210>97<211>470<212>PRT<213>人工序列<220><223>人IgG重链<400>97MetAspTrpThrTrpArgPheLeuPheValValAlaAlaAlaThrGly151015ValGlnSerGluValGlnLeuValGluSerGlyGlyGlyLeuValGln202530ProGlyGlySerLeuArgLeuSerCysAlaAlaSerGlyPheAsnIle354045LysAspThrTyrIleHisTrpValArgGlnAlaProGlyLysGlyLeu505560GluTrpValAlaArgIleTyrProThrAsnGlyTyrThrArgTyrAla65707580AspSerValLysGlyArgPheThrIleSerAlaAspThrSerLysAsn859095ThrAlaTyrLeuGlnMetAsnSerLeuArgAlaGluAspThrAlaVal100105110TyrTyrCysSerArgTrpGlyGlyAspGlyPheTyrAlaMetAspTyr115120125TrpGlyGlnGlyThrLeuValThrValSerSerAlaSerThrLysGly130135140ProSerValPheProLeuAlaProSerSerLysSerThrSerGlyGly145150155160ThrAlaAlaLeuGlyCysLeuValLysAspTyrPheProGluProVal165170175ThrValSerTrpAsnSerGlyAlaLeuThrSerGlyValHisThrPhe180185190ProAlaValLeuGlnSerSerGlyLeuTyrSerLeuSerSerValVal195200205ThrValProSerSerSerLeuGlyThrGlnThrTyrIleCysAsnVal210215220AsnHisLysProSerAsnThrLysValAspLysLysValGluProLys225230235240SerCysAspLysThrHisThrCysProProCysProAlaProGluLeu245250255LeuGlyGlyProSerValPheLeuPheProProLysProLysAspThr260265270LeuMetIleSerArgThrProGluValThrCysValValValAspVal275280285SerHisGluAspProGluValLysPheAsnTrpTyrValAspGlyVal290295300GluValHisAsnAlaLysThrLysProArgGluGluGlnTyrAsnSer305310315320ThrTyrArgValValSerValLeuThrValLeuHisGlnAspTrpLeu325330335AsnGlyLysGluTyrLysCysLysValSerAsnLysAlaLeuProAla340345350ProIleGluLysThrIleSerLysAlaLysGlyGlnProArgGluPro355360365GlnValTyrThrLeuProProSerArgGluGluMetThrLysAsnGln370375380ValSerLeuThrCysLeuValLysGlyPheTyrProSerAspIleAla385390395400ValGluTrpGluSerAsnGlyGlnProGluAsnAsnTyrLysThrThr405410415ProProValLeuAspSerAspGlySerPhePheLeuTyrSerLysLeu420425430ThrValAspLysSerArgTrpGlnGlnGlyAsnValPheSerCysSer435440445ValMetHisGluAlaLeuHisAsnHisTyrThrGlnLysSerLeuSer450455460LeuSerProGlyLeuGlu465470<210>98<211>30<212>DNA<213>人工序列<220><223>人IgG重链<400>98acaaaggtggacaggaaggtggagcccaag30<210>99<211>30<212>DNA<213>人工序列<220><223>人IgG重链<400>99cttgggctccaccttcctgtccacctttgt30<210>100<211>34<212>DNA<213>人工序列<220><223>人IgG重链<400>100gagtataagtgcagggtgtccaataaggccctgc34<210>101<211>34<212>DNA<213>人工序列<220><223>人IgG重链<400>101gcagggccttattggacaccctgcacttatactc34<210>102<211>34<212>DNA<213>人工序列<220><223>人IgG重链<400>102ctttctgtatagcaggctgaccgtggataagtcc34<210>103<211>34<212>DNA<213>人工序列<220><223>人IgG重链<400>103ggacttatccacggtcagcctgctatacagaaag34<210>104<211>238<212>PRT<213>人工序列<220><223>人IgG轻链<400>104MetAspMetArgValProAlaGlnLeuLeuGlyLeuLeuLeuLeuTrp151015LeuSerGlyAlaArgCysAspIleGlnMetThrGlnSerProSerSer202530LeuSerAlaSerValGlyAspArgValThrIleThrCysArgAlaSer354045GlnAspValAsnThrAlaValAlaTrpTyrGlnGlnLysProGlyLys505560AlaProLysLeuLeuIleTyrSerAlaSerPheLeuTyrSerGlyVal65707580ProSerArgPheSerGlySerArgSerGlyThrAspPheThrLeuThr859095IleSerSerLeuGlnProGluAspPheAlaThrTyrTyrCysGlnGln100105110HisTyrThrThrProProThrPheGlyGlnGlyThrLysValGluIle115120125LysArgThrValAlaAlaProSerValPheIlePheProProSerAsp130135140GluGlnLeuLysSerGlyThrAlaSerValValCysLeuLeuAsnAsn145150155160PheTyrProArgGluAlaLysValGlnTrpLysValAspAsnAlaLeu165170175GlnSerGlyAsnSerGlnGluSerValThrGluGlnAspSerLysAsp180185190SerThrTyrSerLeuSerSerThrLeuThrLeuSerLysAlaAspTyr195200205GluLysHisLysValTyrAlaCysGluValThrHisGlnGlyLeuSer210215220SerProValThrLysSerPheAsnArgGlyGluCysLeuGlu225230235<210>105<211>30<212>DNA<213>人工序列<220><223>人IgG轻链<400>105cctggcaaggccccaaggctgctgatctac30<210>106<211>30<212>DNA<213>人工序列<220><223>人IgG轻链<400>106gtagatcagcagccttggggccttgccagg30<210>107<211>30<212>DNA<213>人工序列<220><223>人IgG轻链<400>107acaaaggtggagatcaggaggaccgtggcc30<210>108<211>30<212>DNA<213>人工序列<220><223>人IgG轻链<400>108ggccacggtcctcctgatctccacctttgt30<210>109<211>30<212>DNA<213>人工序列<220><223>人IgG轻链<400>109gccaaggtgcagtggagggtggataacgcc30<210>110<211>30<212>DNA<213>人工序列<220><223>人IgG轻链<400>110ggcgttatccaccctccactgcaccttggc30当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1