一种利用玉米ZmDPS10-2基因选育早花玉米的方法及其应用与流程
本发明属于作物遗传育种领域,涉及一种利用玉米ZmDPS10-2基因选育早花玉米的方法及其应用。
背景技术:
目前,我国玉米育种和生产用的种质来源主要集中在来自美国的瑞德、兰卡斯特和国内唐四平头、旅大红骨四种传统类型以及一种较新种质类型 P 群。尤其是郑单 958 和先玉 335 取得成功以后,国内的育种又进一步局限于改良这两个杂交种的亲本,使育种的种质资源更加狭窄。狭窄的遗传基础不但成为玉米产量提高、品质改善的主要障碍,而且增加了玉米杂交种遭受病虫侵害的可能性,使玉米生产陷入遗传脆弱性的危险。
外来种质特别是来自玉米多样性中心的热带、亚热带种质,具有我国乃至美国玉米带种质不同的遗传变异,经过长期的自然和人工选择,形成了众多的种质群体,具有许多温带种质所不具备的优良性状,如耐旱性、耐湿性、气生根发达、抗倒伏、抗病性强、叶色深绿、叶绿素含量高、叶片功能期长,是拓宽温带玉米种质的优良素材,可以极大的丰富温带玉米育种的遗传基础。热带、亚热带玉米种质具有丰富的遗传变异性,且与温带种质的交流很少,是温带地区玉米育种不可多得的异源种质(Holland and Goodman 1995)。热带、亚热带玉米种质的利用,不仅可以拓宽温带玉米种质的基础、提高品种的生态适应性,对创制新的杂种优势模式也有重要意义。然而,热带、亚热带玉米种质长期处于低纬度地区,具有很强的光周期敏感性,在温带地区长日照环境下表现出营养生长旺盛,生殖生长受到抑制,抽雄期和吐丝期延迟,晚熟,雌雄不协调,甚至不能开花结实。光周期敏感性严重限制了热带玉米在温带地区的利用 (Holland and Goodman 1995)。因此,全面了解玉米光周期敏感性的生理生化机制和遗传机理,对钝化玉米的光周期敏感性,从战略上改良我国玉米种质有重要意义,同时对于进一步解析光周期影响高等植物开花机理的研究具有参考价值。
光周期现象是自然界普遍存在的现象,光周期影响高等植物开花的分子机制是目前发育生物学研究的热点和难点。近年来生物学家利用模式植物拟南芥做了大量的遗传研究,发现许多基因参与调控植物的开花时间,并且确定至少存在 4 条主要的调控植物开花时间的遗传途径:即光周期途径、春化途径、自主途径和赤霉素途径。其中,光周期途径是拟南芥开花途径中最重要、研究最为详细的一个途径,是由PhyA PhyB Cry1 Cry2等光受体直接感受光信号后把信号传递给生物节律钟,诱导CCA1、LHY等生物钟成分和一类伪应答调节蛋白 APRRs 小家族基因的表达,产生生物节律,并将节律输出给下游的节律调节基因GI等。节律调节基因通过调控下游开花时间基因FT、LFY和SOC1等成花途径整合因子的表达,进一步激活或抑制花分生组织基因和花器官基因的表达,从而控制拟南芥的开花过程(Fornara, et al.,2010)。尽管该路径中大部分关键基因的结构具有高度的保守性(Nemoto et al., 2003;Kevin et al., 2010;wang et al., 2011)。但各物种中调控光周期的路径不尽相同,并具有独特的基因成分(水稻的Ghd7和它在玉米中的同源基因ZmCCT、Hd1和EHd1,拟南芥没同源物);部分同源基因在不同物种中的作用不完全相同甚至相反。例如:在短日照条件下Hd3a和RFT1基因高效表达,促进水稻开花;在长日照条件下,Hd1 蛋白抑制Hd3a转录,使Hd3a表达量降低进而推迟水稻开花,EHd1 蛋白使RFT1表达量增加并促进开花,而EHd1基因受光敏色素PHYB及Ghd7的负向调控和MAD50的正向调控(Reina et al., 2009)。拟南芥在长、短日照下通过 CO 蛋白来正向调控FT基因,但 CO 蛋白的稳定性受光敏色素PHYB的负向调控和PHYA、隐花色素的正向调控(Sawa et al. 2007;Srikanth et al., 2011);同时,GI能直接调控FT的表达而促进开花,生物钟蛋白 CCA1 通过与GI、SOC1基因的启动子结合而抑制开花(Lu et al., 2012)。此外,拟南芥和小麦均为长日照作物,在低温调控过程中,小麦没有拟南芥FLC基因的同源物,而是通过VRN家族来实现的(Aaron et al., 2009)。这些结果表明植物在长期的进化过程中,结构相似的基因既保留了已有的功能,又为适应环境变化而可能出现不同的功能,同时各物种也可能产生了各自独特的调控基因。
技术实现要素:
本发明的目的在于提供一种调控玉米开花时间的基因ZmDPS10-2,该基因具有如 SEQ ID NO:1 所示的DNA片段。该基因随着mRNA积累量增加,导致玉米长日照条件下早开花,因此该基因的克隆有助于理解玉米光周期敏感的分子机制。
本发明利用过表达转基因技术,增加玉米内源ZmDPS10-2基因的表达,导致玉米在长日照条件下早开花(见实施例4)。具体的说就是将ZmDPS10-2基因与其它调控元件,如组成型启动子(如CaMV35S启动子)或器官特异性启动子融合构建基因过量表达载体,通过转基因技术创造长日照条件下早开花的玉米自交系,用于培育早熟玉米新品种。
为实现上述目的,本发明采用以下技术方案:
一种利用玉米ZmDPS10-2基因选育早花玉米的方法,步骤如下:
(1)以光周期敏感晚开花的自交系CML288为供体亲本,以光周期钝感早开花的自交系黄早4为受体亲本,采用回交转育结合分子标记的方法,辅助选择构建位于第十染色体的主效qDPS10-2的近等基因系黄早4-NIL,近等基因系的构建方法具体实施例中详细描述;
(2)采用图位克隆技术分离ZmDPS10-2基因,利用qRT-PCR技术分析ZmDPS10-2的表达规律,结果表明该基因不同时期不同部位长日照条件下在黄早4中的表达量,显著高于黄早4-NIL中的表达量,且在茎尖和叶片中高量表达,说明ZmDPS10-2高量表达导致玉米提前开花,利用转基因过表达技术增加玉米内源ZmDPS10-2基因的表达,选育在长日照条件下玉米早开花的自交系。
所述玉米ZmDPS10-2基因为控制玉米开花时间基因,玉米ZmDPS10-2基因的核苷酸序列如SEQ ID NO:1所示,玉米ZmDPS10-2基因编码的蛋白质序列如SEQ ID NO:2所示。
所述利用转基因过表达技术增加玉米内源ZmDPS10-2基因的表达的具体操作为:
a 在玉米ZmDPS10-2基因的编码区域设计上游引物T-F和下游引物T-R,其中T-F序列如SEQ ID NO:3所示,T-R序列如SEQ ID NO:4所示;
b 以玉米自交系黄早4的总cDNA为模板,以步骤a中的T-F和T-R为引物,扩增目的基因;
c 将步骤b中的目的基因重组到pBI1304载体质粒中得到重组质粒,然后把重组质粒转入农杆菌感受态细胞,用于转化玉米自交系A188,制备开花时间不同的转基因阳性株。
一种利用玉米ZmDPS10-2基因选育早花玉米的方法,作为培育早熟玉米新品种的应用。
本发明的有益效果在于:
1. 本发明采用正向遗传学方法(通过构建作图群体对玉米开花时间进行QTL初步定为、精细定位)准确无误的分离候选基因。
2. ZmDPS10-2基因的功能在玉米中没有任何报道,本发明提供了调控玉米长日照条件下开花时间的基因ZmDPS10-2及其编码蛋白,该基因对于解析玉米光周期敏感调控开花时间的分子机理等理论研究具有重要的价值。将含有SEQ ID NO:1所示序列的基因进行改造,导入玉米植株中,可以创建长日照条件下提前开花的玉米育种材料,对玉米育种具有重要的实际应用意义。
3. 本发明涉及的基因ZmDPS10-2是通过图位克隆技术发现的,与玉米开花时间有关的一个含有CCAAT-HAP5结构域的新基因,其分子生物学功能目前在玉米中尚未报道。本发明研究了该基因调控长日照条件下玉米开花时间的分子生物学功能,并为培育长日照条件下玉米早开花的优良高产玉米品种提供了一个潜在的新基因资源。
附图说明
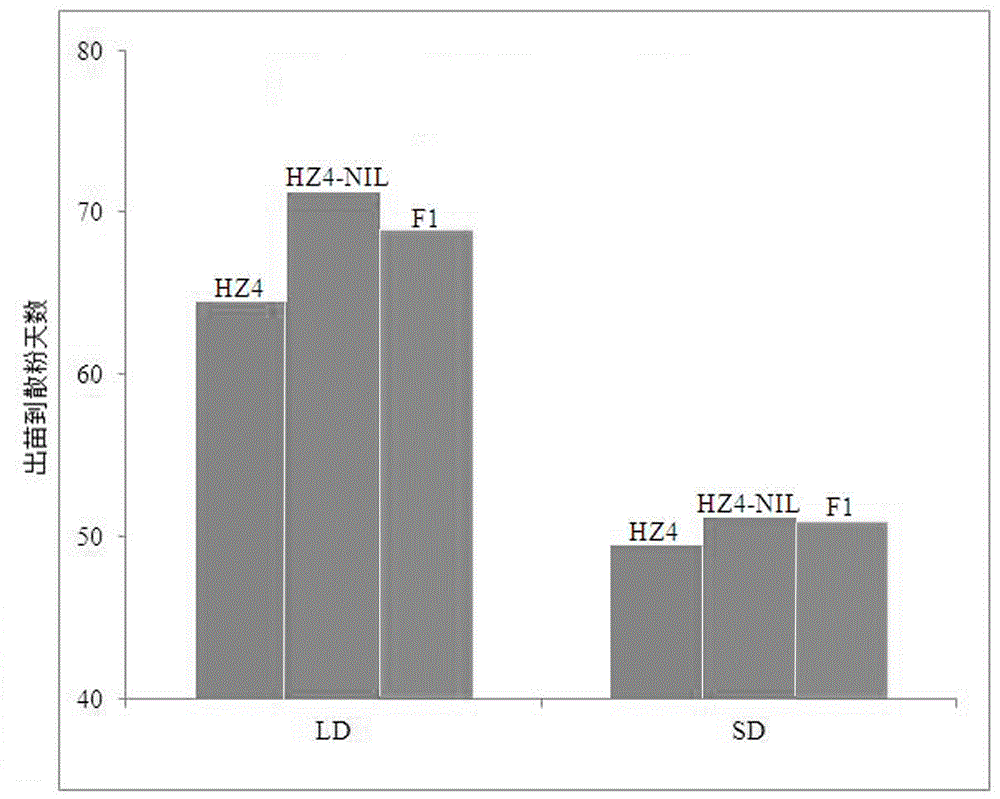
图1为黄早4和黄早4-NIL的出苗到开花散粉时间图,其中HZ4为黄早4的平均天数,HZ4-NIL为黄早482的平均天数,F1为杂交一代的平均天数。
图2为图位克隆ZmDPS10-2基因的分子鉴定结果,图中A:利用BC2F2、BC3F1、BC4F1和BC5F1群体14543个单株,筛选到72个交换单株,对72个交换单株进行精细定位将qDPS10-2 QTL缩小的标记SSR10-68和SSR10-88区间内;B:利用BC6F2群体共8324个单株,共筛选到18株交换单株,对18个交换单株进行精细定位将qDPS10-2 QTL缩小的标记SSR10-151 和 SSR10-230区间内;C:通过精细定位将qDPS10-2的区间缩小到86.81 Kb的区域内;D:该区域包含5个候选基因(其中GRMZM2G375585和GRMZM2G002320是专座子,基因不起作用;GRMZM2G489115不表达,可能也不起作用)。黑色线条和红色方框分别代表基因的内含子和外显子;E:2个表达的候选基因GRMZM2G311316(ZmDPS10-2,左)和GRMZM2G009681(右)在黄早4与黄早4-NIL中短日照(上)和长日照(下)条件下的表达量(mRNA积累量)。
图3为ZmDPS10-2基因的表达模式,A:qRT-PCR检测ZmDPS10-2基因从3-12叶期在黄早4和黄早4-NIL8在短(9小时光照,15小时黑暗)日照条件下叶片(SL)和茎尖(SS)中的表达量(mRNA积累量),18S基因作为内对照;B:qRT-PCR检测ZmDPS10-2基因从3-12叶期在黄早4和黄早4-NIL8在长(15小时光照,9小时黑暗)日照条件下叶片(LL)和茎尖(LS)中的表达量(mRNA积累量),18S基因作为内对照;C:qRT-PCR检测ZmDPS10-2基因6叶期在黄早4和黄早4-NIL8在短(9小时光照,15小时黑暗)日照条件下叶片(SL)和茎尖(SS)中的24小时节律性表达量(mRNA积累量),18S基因作为内对照;D:qRT-PCR检测ZmDPS10-2基因6叶期在黄早4和黄早4-NIL8在长(15小时光照,9小时黑暗)日照条件下叶片(LL)和茎尖(LS)中的24小时节律性表达量(mRNA积累量),18S基因作为内对照。
具体实施方式
一种利用玉米ZmDPS10-2基因选育早花玉米的方法,步骤如下:
(1)以光周期敏感晚开花的自交系CML288为供体亲本,以光周期钝感早开花的自交系黄早4为受体亲本,采用回交转育结合分子标记的方法,辅助选择构建位于第十染色体的主效qDPS10-2的近等基因系黄早4-NIL,近等基因系的构建方法具体实施例中详细描述;
(2)采用图位克隆技术分离ZmDPS10-2基因,利用qRT-PCR技术分析ZmDPS10-2的表达规律,结果表明该基因不同时期不同部位长日照条件下在黄早4中的表达量,显著高于黄早4-NIL中的表达量,且在茎尖和叶片中高量表达,说明ZmDPS10-2高量表达导致玉米提前开花,利用转基因过表达技术增加玉米内源ZmDPS10-2基因的表达,选育在长日照条件下玉米早开花的自交系。
所述玉米ZmDPS10-2基因为控制玉米开花时间基因,玉米ZmDPS10-2基因的核苷酸序列如SEQ ID NO:1所示,玉米ZmDPS10-2基因编码的蛋白质序列如SEQ ID NO:2所示。
所述利用转基因过表达技术增加玉米内源ZmDPS10-2基因的表达的具体操作为:
a 在玉米ZmDPS10-2基因的编码区域设计上游引物T-F和下游引物T-R,其中T-F序列如SEQ ID NO:3所示,T-R序列如SEQ ID NO:4所示;
b 以玉米自交系黄早4的总cDNA为模板,以步骤a中的T-F和T-R为引物,扩增目的基因;
c 将步骤b中的目的基因重组到pBI1304载体质粒中得到重组质粒,然后把重组质粒转入农杆菌感受态细胞,用于转化玉米自交系A188,制备开花时间不同的转基因阳性株。
一种利用玉米ZmDPS10-2基因选育早花玉米的方法作为培育早熟玉米新品种的应用。
本发明是利用北京市农林科学院选育的光周期钝感长日照条件下早开花散粉的玉米自交系黄早4为母本,以墨西哥国际玉米小麦中心引进的光周期敏感长日照条件下晚开花散粉的玉米自交系CML288为父本杂交获得F1,F1自交获得F2群体及其衍生的重组自交系群体为定位群体,通过长短日照2年2个地点的开花散粉时间的表型鉴定,对开花散粉时间进行了QTL定位,共定位到29个光周期敏感(开花时间)的QTLs,其中位于第十染色体上的主效qDPS10的效应值最大(Wang et al. 2008)。为此,以自交系CML288为供体亲本,自交系黄早4为受体亲本构建qDPS10的近等基因系。采用回交转育结合分子标记辅助选择方法构建而成,构建的近等基因系为黄早4-NIL(图1),为进一步理解本发明的内容与目的,下面的实施例将详细介绍本发明的具体技术实施步骤。
一、近等基因系的构建
以光周期敏感长日照条件下开花散粉晚的玉米自交系CML288为供体亲本,光周期钝感长日照条件下开花散粉早的玉米自交系黄早4为受体亲本通过回交转育结合分子标记辅助选择技术构建近等基因系,如图1所示,黄早4明显比黄早4-NIL开花早。2007年春在郑州两亲本组配获得F1;2007年冬在海南种植F1代及受体亲本黄早4,用黄早4回交F1获得BC1F1;将获得的BC1F1的于 2008年春在郑州种成穗行,继续与黄早4回交获得BC2F1;从中选择开花散粉晚的20个单株于2008年冬在海南三亚种成穗行,通过分子标记选择带有CML288目的片段的单株继续与黄早4回交获得BC3F1;2009年春,将BC3F1在郑州种成穗行,从BC3F1中根据选择开花散粉晚的单株25个交换单株继续与黄早4回交获得BC4F1;2010年春在郑州,将BC4F1种成穗行,根据BC4F1群体的表型结合基因型选择了20株交换单株继续与黄早4回交获得BC5F1;2011年春在郑州,将BC5F1种成穗行,根据BC5F1群体的表型结合基因型选择了14株交换单株继续与黄早4回交获得BC6F1;2012年春在郑州,将BC6F1种成穗行,根据BC6F1群体的表型结合基因型选择了16株交换单株继续与黄早4回交获得BC7F1;2013年春在郑州,将BC7F1种成穗行,根据BC4F1群体的表型结合基因型选择了13株交换单株自交获得BC7F2;2014春在郑州将获得BC7F2种植,根据BC7F2群体的表型结合基因型分析,获得13个交换单株,并对13个交换单株采用玉米3K MaizeSNP芯片进行遗传背景分析,从中选择遗传背景回复率高的5个交换单株自交,进而获得在该位点纯合、遗传稳定的近等基因系。
二、采用图位克隆技术分离ZmDPS10-2基因
利用BC2F2、BC3F1、BC4F1和BC5F1群体14543株作为精细定位群体,从中选择晚开花的单株876,用qDPS10-2双侧的标记bnlg210和bnlg1079分析其基因型,从中选择在bnlg210或bnlg1079处发生重组交换单株72株。利用新开发并具有多态性强的19对引物SSR10-8, SSR10-18, SSR10-19, SSR10-21, SSR10-22, SSR10-28, SSR10-31, SSR10-32, SSR10-33, SSR10-44, SSR10-67, SSR10-68, SSR10-72, SSR10-73, SSR10-77, SSR10-78, SSR10-85, SSR10-86, SSR10-88(标记开发方法参照张君,玉米叶夹角有关的ZmCLA4基因图位克隆与功能分析,河南农业大学博士学位论文,2014)分析交换单株基因型。根据交换单株目标区段的标记基因型,可以把qDPS10-2限定在标记SSR10-68和SSR10-88之间(图2A,B)。
为了进一步确定qDPS10-2为1个或多个候选基因,利用BC6F2群体8324株作为精细定位群体,从中选择晚开花散粉的单株有1605株,用qDPS10-2双侧的标记SSR10-68和SSR10-88分析其基因型,从中选择在SR10-68或SSR10-88处发生重组交换单株18株。再利用具有多态性强的21对新开发的标记(SSR10-90, SSR10-96, SSR10-103, SSR10-108, SSR10-122, SSR10-131, SSR10-137, SSR10-146, SSR10-151, SSR10-161, SSR101-164, SSR10-172, SSR10-179, SSR10-188, SSR10193, SSR10-205, SSR10-218, SSR10-223, SSR10-225, SSR10-230)分析交换单株基因型。根据交换单株目标区段的标记基因型,可以把qDPS10-2限定在标记SSR10-151和 SSR10-230之间(图2B,C)。
为更进一步确定qDPS10-2为1个或多个候选基因,利用BC7F2群体6425株作为精细定位群体,从中选择晚开花散粉的单株有1254株,用qDPS10-2双侧的标记SSR10-151和 SSR10-230分析其基因型,从中选择在SSR10-151或SSR10-230处发生重组交换单株11株。再利用具有多态性强的11对新开发的标记(SSR10-161, SSR101-164, SSR10-172, SSR10-179, SSR10-188, SSR10193, SSR10-205, SSR10-212, SSR10-218, SSR10-223, SSR10-225)分析交换单株基因型。根据交换单株目标区段的标记基因型,可以把qDPS10-2限定在标记SSR10-188 and SSR10-212之间(图2C,D)。该区间大约86.81-kb,包含5个预测基因(图2D),因此将标记SSR10-188和SSR10-212之间的86.81-kb区域作为qDPS10-2的基因所在区域。基因功能注释结果表明GRMZM2G375585和GRMZM2G002320是专座子,这两个基因自身没有什么功能;GRMZM2G489115不表达,其余2个基因(GRMZM2G311316,GRMZM2G009681)高效表达。以黄早4和黄早4-NIL的mRNA反转录成cDNA为模板,分离高效表达的2个候选基因,同时以上述2个材料的DNA为模板,分离2个候选基因的启动序列。通过序列差异性分析结果显示,GRMZM2G311316编码序列和3’-UTR序列在黄早4和黄早4-NIL中存在差异,另一个表达基因所有序列都不存在差异。说明黄早4和黄早4-NIL的长日照条件下开花散粉时间存在差异是由于GRMZM2G311316在编码和3’-UTR序列的差异引起的。为进一步验证此结论,采用qTR-PCR技术(参照张君,玉米叶夹角有关的ZmCLA4基因图位克隆与功能分析,河南农业大学博士学位论文,2014)以5、6、7叶期的黄早4和黄早4-NILs为叶片材料,提取mRNA并反转录成cDNA;以这些cDNA为模板分析上述2个高效表达的候选基因的表达量,结果表明,GRMZM2G311316在2个长日照条件下材料中存在显著差异,GRMZM2G009681在2个不同日照条件下材料中虽然表达但不存在显著差异(图2E)。这些结果表明黄早4和黄早4-NIL长日照条件下开花散粉时间存在差异是由GRMZM2G311316的在长日照条件下表达量不同引起的,由此说明GRMZM2G311316就是qDPS10-2区域内调控长日照条件下玉米开花散粉早晚的的候选基因,命名为ZmDPS10-2。
三、ZmDPS10-2正向调控玉米长日照条件下开花散粉时间
ZmDPZ10-2在黄早4中使玉米在长日照条件下提前开花散粉,在黄早4-NIL中使玉米在长日照条件下晚开花散粉,因此,检测了ZmDPS10-2基因的表达情况。qRT-PCR检测结果表明,从玉米生长发育的3到12叶期的叶片和茎尖(图3A,B)中均高效表达。在玉米的不同发育阶段,黄早4和黄早4-NIL的表达量具有一定的相似性,都是从3叶期开始表达,6-7和10-11叶期表达量较高。就不同材料而言,短日照条件下ZmDPS10-2在黄早4和黄早4-NIL(图3A)中表达量差异不明显;长日照条件下ZmDPS10-2在黄早4的表达量显著高于黄早4-NIL(图3B)。这些结果表明ZmDPS10-2的表达量高玉米在长日照条件下提前开花散粉,由此提出ZmDPS10-2作为正向调控因子调控玉米长日照条件下早开花散粉。
四、ZmCLA1过表达转基因验证其基因功能
1、过表达载体的构建
根据ZmDPS10-2基因的编码区序列设计一对引物T-F与T-R,引物5’端分别加上Nco I 和SpeI酶切位点。用该引物扩增黄早4材料中ZmDPS10-2的cDNA序列,PCR产物琼脂糖凝胶电泳检测,将回收目的PCR产物连接pMD18-T载体,转化、挑选单克隆、PCR检测、测序、并将测序正确的菌液提取质粒,将该质粒与pBI1304载体质粒分别用Nco I和SpeI酶切位点进行双酶切。分别回收酶切后的目的片段,利用T4连接酶连接载体片段和目的片段,连接产物转化大肠杆菌,鉴定阳性克隆,提取重组质粒,命名为pBI1304-ZmDPS10-2。将重组质粒转化农杆菌感受态细胞,用于转化玉米自交系A188。
引物T-F如SEQ ID NO:3所示,其中'5-CCATGG-3'为酶切位点Nco I;引物T-R如SEQ ID NO:4所示,其中5'-GGACTA-3'为酶切位点SpeI。
2、遗传转化
采用农杆菌介导的玉米幼胚遗传转化方法(参照王平安,基于RNA干扰的抗粗缩病转基因玉米材料创制,河南农业大学硕士学位论文,2011)。经过幼胚准备、农杆菌准备、农杆菌侵染幼胚、幼胚和农杆菌的共培养、静息培养、抗性愈伤组织的筛选、植株的再生和再生植株的环境适应等环节,再生的植株适应环境后移栽到大田。等移栽的植株长到4-5叶期时,喷洒除草剂进行抗性筛选,剔除对除草剂敏感的植株,对除草剂钝感的植株提取叶片总DNA,进行标记基因的PCR检测,自交获得T1代种子,完成对转基因T1代的鉴定过程。
本实施例共获得17株独立的转基因阳性株,其在长日照条件下玉米开花散粉时间表现不同程度的提前,而7株空载体pBI1304的对照株则没有明显的变化。
由此可见,通过过表达转基因技术增强玉米中内源ZmDPS10-2基因的表达后,导致玉米在长日照条件下开花散粉时间明显提前,证明控制ZmDPS10-2基因的表达可应用培育长日照条件下玉米提前开花散粉的自交系作为育种的基础材料,可用于培育早熟的育杂交种在生产上推广应用。
<110>河南农业大学
<120>一种利用玉米ZmDPS10-2基因选育早花玉米的方法及其应用
<160>4
<210> 1
<211> 1399
<212> DNA
<213> 玉米(Zea mays L.)
<220>
<221> CDS
<222> (1)...(756)
<400> 1
atg gaa cca tcc tca cag cct caa cca gtg atg ggt gtt ggg tca 45
Met Glu Pro Ser Ser Gln Pro Gln Pro Val Met Gly Val Gly Ser
1 5 10 15
caa cct tat cct gct gct gct ccc tat gtg cca cta cct gga gct 90
Gln Pro Tyr Pro Ala Ala Ala Pro Tyr Val Pro Leu Pro Gly Ala
20 25 30
ctt gct gtt cct cct ggc tca cag cca gca gca gca ttc ccc aat 135
Leu Ala Val Pro Pro Gly Ser Gln Pro Ala Ala Ala Phe Pro Asn
35 40 45
cca act caa ctc agt gct cag cac cag atg gtg tac cag cag gct 180
Pro Thr Gln Leu Ser Ala Gln His Gln Met Val Tyr Gln Gln Ala
50 55 60
cag caa ttt cat caa caa ctt cag caa cag cag gaa caa cag ctc 225
Gln Gln Phe His Gln Gln Leu Gln Gln Gln Gln Glu Gln Gln Leu
65 70 75
agg gag ttc tgg act acc cag atg gat gag atc aag caa gca aat 270
Arg Glu Phe Trp Thr Thr Gln Met Asp Glu Ile Lys Gln Ala Asn
80 85 90
gac ttc aag atc cac acc ttg cca ctt gca agg ata aag aag ata 315
Asp Phe Lys Ile His Thr Leu Pro Leu Ala Arg Ile Lys Lys Ile
95 100 105
atg aag gct gat gag gat gtg cgg atg atc tct gca gaa gct cct 360
Met Lys Ala Asp Glu Asp Val Arg Met Ile Ser Ala Glu Ala Pro
110 115 120
gtt gtg ttt gcg aag gca tgc gag gta ttc ata tta gag ctg aca 405
Val Val Phe Ala Lys Ala Cys Glu Val Phe Ile Leu Glu Leu Thr
125 130 135
ttgagg tca tgg atg cac aca gag gag aac aag cgc cgg acc ttg 450
Leu Arg Ser Trp Met His Thr Glu Glu Asn Lys Arg Arg Thr Leu
140 145 150
cag aac aac gac att gca gct gcc atc acc agg act gat ata tat 495
Gln Asn Asn Asp Ile Ala Ala Ala Ile Thr Arg Thr Asp Ile Tyr
155 160 165
gac ttc ctg gtg gac ata atc ccg agg gat gaa atg aaa gag gag 540
Asp Phe Leu Val Asp Ile Ile Pro Arg Asp Glu Met Lys Glu Glu
170 175 180
ggg ctc gga cat aat ccc ata gtt ggc ctg ccg cct gct atg ggg 585
Gly Leu Gly His Asn Pro Ile Val Gly Leu Pro Pro Ala Met Gly
185 190 195
gct cca gct gat cat ggt tct cat cca tat tac tat gtg cca gct 630
Ala Pro Ala Asp His Gly Ser His Pro Tyr Tyr Tyr Val Pro Ala
200 205 210
cag cag gtg caa caa gca ggt atg ttg tat ggt ggc cag cag ggt 675
Gln Gln Val Gln Gln Ala Gly Met Leu Tyr Gly Gly Gln Gln Gly
215 220 225
cac cca gtg act tat atg tgg cag acg ccg caa ggg caa gcg gaa 720
His Pro Val Thr Tyr Met Trp Gln Thr Pro Gln Gly Gln Ala Glu
230 235 240
gag cct ccc gaa gag gag cag cag cag tct gcc tga aagtagcaaa 766
Glu Pro Pro Glu Glu Glu Gln Gln Gln Ser Ala
245 250
tagacgataa cagcaaagca attacatctt gatctgaagt tctgaactta aggcctgttt 826
ggtgtggcgc cgaaaccgcc gcaccgcgtc taagtcgcgg cgctcgattt ggtcgcgcca 886
actgcggcac aaaatctgtg gcgcgtgtgg gcgtggacgg cggtcaacca aacatgctct 946
tagttctcta aacaaacaaa tatgatttgg ttatgttaat tgtgcagatg attgctggct 1006
gaaacaattg tcgctaatta ggtgtcatag ggtttcctaa tggtagtttc gactcccgac 1066
gctgccatgg cagcagtaga aaatgattgt gatttgcaga ctaatgtagt aagattagcc 1126
ttttctgtct aatccaatat acaattaagg ccctcgtatg aactgagtgg tggtgggact 1186
tctttcgatg cctgcgtttg tttgccttat tttttttaac cattgtgctc tgtgcttggc 1246
tagaaattgc cggtcaccaa agtttcgtag ttaattcaag tgcttatttc aggctgaact 1306
gaattttttt ttcttttgta tggcaagttt taacacataa atactagcag taataactaa 1366
ataaaatttg gccgaccata aatccgcccg tat 1399
<210> 1
<211> 251
<212> PRT
<213> 玉米(Zea mays L.)
<400> 2
Met Glu Pro Ser Ser Gln Pro Gln Pro Val Met Gly Val Gly Ser
1 5 10 15
Gln Pro Tyr Pro Ala Ala Ala Pro Tyr Val Pro Leu Pro Gly Ala
20 25 30
Leu Ala Val Pro Pro Gly Ser Gln Pro Ala Ala Ala Phe Pro Asn
35 40 45
Pro Thr Gln Leu Ser Ala Gln His Gln Met Val Tyr Gln Gln Ala
50 55 60
Gln Gln Phe His Gln Gln Leu Gln Gln Gln Gln Glu Gln Gln Leu
65 70 75
Arg Glu Phe Trp Thr Thr Gln Met Asp Glu Ile Lys Gln Ala Asn
80 85 90
Asp Phe Lys Ile His Thr Leu Pro Leu Ala Arg Ile Lys Lys Ile
95 100 105
Met Lys Ala Asp Glu Asp Val Arg Met Ile Ser Ala Glu Ala Pro
110 115 120
Val Val Phe Ala Lys Ala Cys Glu Val Phe Ile Leu Glu Leu Thr
125 130 135
Leu Arg Ser Trp Met His Thr Glu Glu Asn Lys Arg Arg Thr Leu
140 145 150
Gln Asn Asn Asp Ile Ala Ala Ala Ile Thr Arg Thr Asp Ile Tyr
155 160 165
Asp Phe Leu Val Asp Ile Ile Pro Arg Asp Glu Met Lys Glu Glu
170 175 180
Gly Leu Gly His Asn Pro Ile Val Gly Leu Pro Pro Ala Met Gly
185 190 195
Ala Pro Ala Asp His Gly Ser His Pro Tyr Tyr Tyr Val Pro Ala
200 205 210
Gln Gln Val Gln Gln Ala Gly Met Leu Tyr Gly Gly Gln Gln Gly
215 220 225
His Pro Val Thr Tyr Met Trp Gln Thr Pro Gln Gly Gln Ala Glu
230 235 240
Glu Pro Pro Glu Glu Glu Gln Gln Gln Ser Ala
245 250
<210> 3
<211> 24
<212> DNA
<213> 人工序列
<220>
<221> misc_difference
<222> (1)...(24)
<400> 3
ccatggatgg aaccatcctc acag 24
<210> 4
<211> 24
<212> DNA
<213> 人工序列
<220>
<221> misc_difference
<222> (1)...(24)
<400> 4
ggactatcag gcagactgct gctg 24
- 还没有人留言评论。精彩留言会获得点赞!