果实大小增大的植物的制作方法
本申请为分案申请,其原申请的申请日为2011年5月27日,申请号为201180036919.0,名称为“果实大小增大的植物”。
本发明涉及植物生物技术和植物育种的领域。本发明提供了果实大小增大的植物(特别是具有更大和更重的番茄果实的番茄(solanumlycopersicum)植物),以及用于制备果实大小增大的果实的遗传改良或突变植株的方法。本发明提供了基因slarf9(编码slarf9蛋白)的新用途,发现其为果实发育过程中细胞分裂的负调节因子。下调、敲除或沉默slarf9基因可导致植物在果实生长期结束时具有显著更大的果实。果实更大是由于果皮组织的细胞分裂增加,形成具有更多细胞(并因此含有更多纤维素、半纤维素、果胶等)的大果实。本发明还提供了植物、种子、果实和植物的部分,其基因组中含有突变型slarf9等位基因(本文中命名为slarf9),并且slarf9等位基因突变的结果是在果实生长期结束时具有显著更大的果实。在另一个实施方案中,本文提供了用于制造或鉴定其基因组中含有一个或多个突变型slarf9等位基因的植物的方法。
背景技术:
被子植物(显花植物)是陆生植物中最大的类群。在被子植物中,心皮是雌性生殖器官,其分化为柱头、花柱和子房(其包裹着胚珠)。传粉和受精成功完成后,胚珠发育成种子,子房发育成果实。从子房到快速生长的果实的转变包括必需紧密协调的分子变化、生化变化和结构变化。依据果实发育期,这些变化的时间和空间组织受植物激素例如生长素、赤霉素、细胞分裂素、脱落酸和乙烯的调节。20世纪早期的研究已经确定,生长素和赤霉素在果实发育起始阶段也是重要的因子。但是,到目前为止,对这些激素控制的复杂调节网络仍所知甚少。
“果实形成”(fruitset)定义为休眠的子房到快速生长的幼果的转变,其是显花植物有性生殖的重要过程。番茄(solanumlycopersicuml.)是研究最多的肉质果实之一,其是茄科(solanaceae)的代表,该科包括其他几种重要的果实作物例如茄子(solanummelongenal.)和辣椒属(capsicumspp.)。番茄的生物学是非常热门的。番茄具有相对短的生命周期,生长和维持的需要简单,并且虽然番茄是自花传粉植物,但其很容易杂交传粉。此外,其具有广泛的遗传资源例如表型趋异的栽培种、可相互杂交的野生近缘种、突变型、和基因组工具(muelleretal.,plantphysiology138,1310-1317;muelleretal.,comparativeandfunctionalgenomics6,153-158)例如bac文库和表达序列标签(est)。
番茄果实形成对环境条件,特别是会影响花粉发育和花药开裂(dehiscence)的太低或太高的温度,是非常敏感的。adamsetal.(2001,annalsofbotany88,869-877)表明,与22℃的方案相比,14℃或26℃的恒温方案显著减少番茄的果实形成。但是,栽培种不同,最适的生长温度也可能不同。这种温度敏感性的结果是,有效的番茄生产受限于具体的气候带。由于该原因,番茄种子公司在世界上不同的地方育种,以开发在当地气候条件下最适于果实生产的栽培种。但是,即使有所述最适宜的株系,在温带地区例如欧洲南部的夏季通常也不可能种植番茄。在更北方的地区,只可能在温暖的季节进行番茄生产,并且即使如此也只能在消耗大量能源进行加热的现代温室中。
果实形成依赖于传粉和受精的成功完成(gillaspyetal.,1993,theplantcell5,1439-1451)。当番茄花在开花期完全开放时,相容的花粉必须在雌蕊上萌发并形成花粉管。然后,所述花粉管生长穿过花柱和胚珠的珠孔以将两个精细胞递送到胚囊。在此发生双受精;两个精细胞中的一个与卵细胞受精,另一个与中央细胞中的两个单倍体极核融合。随后,胚胎和周围组织会产生刺激果实生长的信号。番茄子房由两个或多个心皮组成,其包裹含有胚珠的小室。成功受精后,经过持续10-14天的一段时间的细胞分裂,子房发育成初始果实。接下来的6-7周,果实生长主要依靠细胞膨大(mapellietal.,1978,plantandcellphysiology19,1281-1288;bünger-kiblerandbangerth,1982,plantgrowthregulation1,143-154;gillaspyetal.,1993,见上文)。心皮壁发育成果皮,胚珠附着的胎座发育成凝胶状物质,其由高度液泡化的大薄壁细胞组成。在细胞膨大期结束时,果实达到其最终的大小并且开始成熟(gillaspyetal.,1993,见上文)。共分为6个成熟阶段:不成熟期、成熟期、绿熟期、破色期、粉红期和红熟期。加工番茄通常在红熟期收获,而生鲜市场的番茄在更早的破色期(此时不需要用乙烯催熟至红熟期)或绿熟期(需要暴露到乙烯气体以催熟到红熟期)收获。
虽然植物激素例如生长素和赤霉素对果实发育的影响在20世纪早期已经得到公认(gustafson,1937,americanjournalofbotany24,102-107;gustafson1939americanjournalofbotany26,135-138;wittweretal.,1957,plantphysiology32,39-41),但是果实形成的分子机制在很大程度上还是未知的,其正在被逐渐揭示。
通过影响许多基因的表达,生长素在植物生命周期的很多发育过程中都是重要的调节因子(theologis,1986,annualreviewsofplantphysiology37,407-438)。该生长素-介导的基因表达受到两个转录因子家族(生长素应答因子(arf)和生长素/吲哚-3-乙酸(aux/iaa))的控制,所述两个转录因子家族由植物种例如拟南芥和水稻中两个大基因家族代表(hagenandguilfoyle,2002,plantmolecularbiology49,373-385,plantmolecularbiology49,387-400;liscumandreed,2002,见下文;wangetal.,2007,gene394,13-24)。这些家族编码的蛋白质共有两个保守的c-末端结构域——作为aux/iaa和arf之间相互作用结构域的结构域iii和iv,其使得arf和aux/iaa之间的相互作用分别形成同源二聚体或异源二聚体(kimetal.,1997,proceedingsofthenationalacademyofsciences,usa94,11786-11791;ulmasovetal.,1997,science276,1865-1868;ulmasovetal.,1999,theplantjournal19,309-319)。此外,arf还含有n-末端b3-衍生的dna结合结构域(dbd),所述dbd可结合生长素-介导的基因的启动子区的生长素应答元件(auxre)(ulmasovetal.,1999,见上文)、以及依据其氨基酸组成作为转录激活结构域或抑制结构域发挥功能的中间区(mr)(ulmasovetal,1999,proceedingsofthenationalacademyofsciences,usa96,5844-5849;tiwarietal,2003,theplantcell15,533-543)。aux/iaa蛋白通过封闭arf的转录活性起到抑制因子的作用(liscumandreed,2002,plantmolecularbiology49,387-400)。aux/iaa的抑制活性由n-末端结构域i提供(tiwarietal,2004,theplantcell16,533-543)。最近,szemenyeietal.(2008,science319,1384-1386)表明,在许多aux/iaa中,该结构域含有erf-相关的两亲(amphiphilic)抑制(ear)基序,该基序可招募(recruit)转录共抑制因子topless(tpl)。此外,aux/iaa还含有第四个保守区——结构域ii(tiwarietal.,2001,theplantcell13,2809-2822)。生长素可增强该结构域和scftirl泛素连接酶复合体——含有f-box生长素受体蛋白tir1(transportinhibitorresistant1)——的相互作用,导致泛素-介导的aux/iaa降解(dharmasirietal,2005,nature435,441-445;kepinskiandleyser,2005,nature435,446-451;tanetal,2007,nature446,640-645;dossantosmaraschinetal.,2009,theplantjournal59,100-109)。结果,arf从抑制中释放,导致生长素应答基因的转录(woodwardandbartel,2005,annalsofbotany95,707-735)。但是,该模型只支持转录激活的arf的功能。
arf抑制因子调节生长素-依赖的基因表达的机制仍不清楚,因为它们与aux/iaa或与激活的arf的相互作用非常弱(tiwarietal,2003,见上文;hardtkeetal,2004,development131,1089-1100)。或者,arf抑制因子可能与所述arf激活因子竞争所述生长素应答基因启动子中的auxre结合位点,从而不依赖aux/iaa而抑制这些基因的表达并提供另一种基因调节机制(guilfoyleandhagen,2007,plantbiology10,453-460)。
在番茄中,生长素在果实形成和果实发育中起重要作用。iwahori(1967,plantandcellphysiology8,15-22)和mapellietal.(1978,plantandcellphysiology19,1281-1288)表明,传粉后子房中的生长素浓度快速增加,其在7-8dap(传粉后天数)达到最大值。在30dap观测到生长素活性的第二峰值。生长素在番茄果实形成中的重要性通过如下得到证明:子房特异性表达农杆菌属(agrobacteriumspp.)的iaam或rolb基因,影响生长素合成或应答,导致形成无籽的(单性)番茄果实(ficcadentietal,1999,molecularbreeding5,463-470;carmietal,2003,planta217,726-735)。并且,在未传粉的子房上使用生长素可导致形成果实,而不需要传粉和受精(gustafson,1936,proceedingsofthenationalacademyofsciences,usa22,628-636;bünger-kiblerandbangerth,1982,见上文)。正常地,在发育的头10-14天(受精后),番茄果实的生长主要依靠细胞分裂。在接下来的6-7周,果实生长主要依靠细胞膨大(mapellietal,1978,见上文;bünger-kiblerandbangerth,1982,见上文;gillaspyetal.,1993,见上文)。但是,生长素吲哚-3-乙酸(iaa)诱导的果实的细胞分裂期较短(只持续10天),但与结籽的对照果实相比细胞分裂速率较快。但是,由于细胞膨大受到很大地损伤,这些iaa-诱导的果实仍然比对照果实小(bünger-kiblerandbangerth,1982,见上文)。用合成的生长素的处理刺激了细胞分裂持续延长的一段时间,导致形成具有更多果皮细胞的果实(bünger-kiblerandbangerth,1982,见上文;serranietal,2007,journalofplantgrowthregulation26,211-221)。这些发现显示,在番茄果实发育的早期,细胞分裂活性受生长素的紧密调节。
此前,使用基于转录谱的cdna-扩增片段长度多态性(cdna-aflp)以鉴定在果实形成中差异表达的基因(vriezenetal,2008,newphytologist177,60-76)。通过传粉在子房中诱导的874个基因中的一个与拟南芥arf9(atarf9)和genbank登录号bt013639.1(一个未知基因功能的番茄克隆)具有一些序列相似性。虽然所述基因与atarf9的氨基酸同一性不是很高(使用两两序列比对程序emboss“needle”,52%的序列同一性),该基因被命名为slarf9(番茄arf9)。
拟南芥有23个arf基因。由于拟南芥arf9突变株系——其转录本缺少3'端——在重力刺激后过度应答,所以atarf9被认为参与向重力性信号转导(robertsetal,2007,gravitationalandspacebiologybulletin20,103-104)。此外,还发现atarf9基因在拟南芥胚的胚柄中表达,并且arf9和arf13两者都被沉默的双敲除株系显示,atarf9是控制胚柄发育必需的(liuetal,2008,19thinternationalconferenceonarabidopsisresearch,montreal,canada)。迄今为止,已知参与果实发育过程的仅有的arf是fruitwithoutfertilization(fwf)/arf8,由于fwf/αrf8突变株系形成单性果实(goetzetal,2006,theplantcell18,1873-1886)。在番茄中,slarf7转录本水平减少的转基因株系也形成单性果实,这表明slarf7充当果实形成的负调节因子(dejongetal,2009,theplantjournal57,160-170)。迄今为止,经鉴定的番茄arf家族的仅有的其他成员是developmentallyregulatedgene12(dr12),其为atarf4的同源物。dr12的mrna水平在整个果实发育过程中都增加,并且在果实红熟期早期达到最高水平。通过反义方法下调该基因会影响红熟期的果实硬度(jonesetal,2002,theplantjournal32,603-613)。
虽然有slarf9与atarf9的序列相似性可能显示这些基因可能是直系同源基因(尽管序列相似度低)的事实,但发明人发现slarf9具有与atarf9完全不同的功能,并且与atarf9在不同的组织中表达。发明人发现,slarf9基因编码在果实生长过程中负调节细胞分裂的转录因子蛋白。通过rnai(基因沉默)使slarf9mrna转录本水平降低的转基因番茄植株形成了比野生型对照株系显著更大且更重的果实。相比而言,slarf9过表达株系具有比野生型显著更小且更轻的果实。此外,slarf9-rnai果实的果皮具有比野生型果实的果皮显著更多的细胞数目和细胞层数目。具有更多、更小的细胞的较大果实不仅具有产量相对增加的益处,还具有固体组分(细胞壁,其含有果胶、半纤维素和纤维素)相对增加的益处。
一般性定义
术语“核酸序列”(或核酸分子)是指单链或双链形式的dna或rna分子,特别是编码本发明的蛋白质或蛋白质片段的dna。“分离的核酸序列”是指不再处于将其从中分离的天然环境中的核酸序列,例如细菌宿主细胞或者植物细胞核或质体基因组中的核酸序列。
术语“蛋白质”或“多肽”可互换使用,是指由氨基酸链构成的分子,不管具体的作用模式、大小、三维结构或来源。因此,slarf9蛋白的“片段”或“部分”也可称为“蛋白质”。“分离的蛋白质”用于指不再处于其天然环境中(例如在体外或者在重组的细菌或植物宿主细胞中)的蛋白质。
术语“基因”是指含有可操作地连接到适合的调节区(例如启动子)的区(转录区)——其在细胞中转录成为rna分子(例如mrna或rnai分子)——的dna序列。因此,基因可包含几个可操作地连接的序列例如启动子、含有例如参与转录起始的序列的5'前导序列、(蛋白质)编码区(cdna或基因组dna)和含有例如转录终止点的3'非翻译序列。
“嵌合基因”(或重组基因)是指正常不天然存在于物种中的任何基因,尤其是其中的一部分或多部分核酸序列在天然条件下彼此不连接(associate)的基因。例如,启动子在天然条件下不与所述转录区的部分或全部连接,不与另外的调节区连接。术语“嵌合基因”应理解为包括表达构建体,其中启动子或转录调节序列可操作地连接到一个或多个编码序列或连接到反义(正义链的反向互补链)或反向重复序列(正义和反义序列,以此转录后的rna转录本形成双链rna)。“顺式-基因”是嵌合基因,其中优选全部基因序列(但至少所述转录序列)来自于与将所述基因导入其中的物种性相容的植物物种。
“基因表达”是指可操作地连接到适合的调节区(尤其是启动子)的dna区,转录成有生物活性即能够翻译成生物活性蛋白质或肽(或活性肽段)或者自身有活性(例如在转录后基因沉默中或rnai)的rna的过程。所述编码序列可以是正义方向,并编码需要的生物活性蛋白质或肽或者活性肽段。在基因沉默方法中,所述dna序列优选以反义dna或反向重复dna的形式存在,所述反义dna或反向重复dna含有反义方向的短靶基因序列或者含有正义和反义方向(反向重复)的短靶基因序列。“异位表达”是指在通常不表达所述基因的组织中表达。
“活性蛋白质”或者“功能性蛋白质”是指具有在体外(例如通过体外活性测定)和/或体内(例如通过所述蛋白赋予的表型)可测量的蛋白质活性的蛋白质。“野生型”蛋白质是指野生型植物中存在的全功能性蛋白质。本文中的“突变蛋白质”是在编码该蛋白质的核酸序列中含有一个或多个突变的蛋白质,由此所述突变导致(所述突变型核酸分子编码的)“功能下降”或“功能丧失”的蛋白质,这例如可在体内测量,例如通过突变型等位基因产生的表型。
编码蛋白质的核酸分子中的“突变”是例如通过置换、缺失或插入一个或多个核苷酸造成的与野生型序列相比一个或多个核苷酸的改变。“点突变”是单个核苷酸的置换或者单个核苷酸的插入或缺失。
“无义”突变是编码蛋白质的核酸序列中的(点)突变,由此一个密码子变为终止密码子。这导致在mrna中过早地出现终止密码子,并产生截短的蛋白质。截短的蛋白质可能具有降低的功能或失去功能。
“错义”突变是编码蛋白质的核酸序列中的(点)突变,由此一个密码子变为不同氨基酸的密码子。所产生的蛋白可能具有降低的功能或失去功能。
“剪接位点”突变是编码蛋白质的核酸序列中的突变,由此mrna前体的rna剪接被改变,产生具有与野生型不同的核苷酸序列的mrna和与野生型不同的氨基酸序列的蛋白质。所产生的蛋白质可具有降低的功能或失去功能。
“移码”突变是编码蛋白质的核酸序列的突变,由此mrna的阅读框被改变,产生不同的氨基酸序列。所产生的蛋白质可具有降低的功能或失去功能。
调节序列中例如基因启动子中的突变是通过一个或多个核苷酸的置换、缺失或插入造成的与野生型序列相比一个或多个核苷酸的改变,这导致所述基因的mrna转录本减少或者无mrna转录本。
“沉默”是指下调或完全抑制靶基因或基因家族的基因表达。
基因沉默方法中的“靶基因”是指,当嵌合沉默基因(或者“嵌合rnai基因”)表达并且例如产生沉默rna转录本(例如能够沉默内源靶基因表达的dsrna或发夹rna)时,所述内源基因表达下调或完全被抑制(沉默)的基因或基因家族(或者所述基因的一个或多个具体等位基因)。在诱变方法中,靶基因是要被突变以导致基因表达改变(下降或丧失)或者所编码的蛋白质功能改变(下降或缺失)的内源基因。“正义”rna转录本通常是通过将启动子可操作地连接到双链dna分子产生的,其中所述dna分子的正义链(编码链)为5'到3'方向,使得在转录时转录出正义rna,其具有与所述正义dna链相同的核苷酸序列(除了在rna中t被替换为u)。“反义”rna转录本通常是通过将启动子可操作地连接到所述正义dna的互补链(反义链)产生的,使得在转录时转录出反义rna。
本文将“转录调节序列”定义为能够调节可操作地连接到所述转录调节序列的(编码)序列的转录速率的核酸序列。因此,本文中转录调节序列包含用于起始转录的序列元件(启动子元件)、用于维持和用于调节转录的元件包括例如衰减子或增强子。虽然大多数情况下是指编码序列的上游(5')转录调节序列,但是该定义也涵盖在编码序列下游(3')发现的调节序列。
在本文中,术语“启动子”是指具有控制一个或多个基因转录的功能的核酸片段——其就转录方向而言位于所述基因转录起始位点的上游——并且通过以下在结构上被识别:dna-依赖的rna聚合酶结合位点、转录起始点以及包括但不限于如下的其他任意dna序列:转录因子结合位点、抑制子或激活子蛋白结合位点,以及本领域技术人员公知的能够直接或间接调节所述启动子的转录数量的其他任何核苷酸序列。“组成型”启动子是在大多数生理和发育条件下在大多数组织中有活性的启动子。“诱导型”启动子是受生理(例如通过外部应用某些化合物)或发育调节的启动子。“组织特异性”启动子只在具体类型的组织或细胞中有活性。“组织优选型”启动子主要在某些组织(例如发育中的果实组织)中有活性,但在其他组织中也有一些活性。
本文中,“可操作连接”是指多核苷酸元件以功能关系的连接。当将核酸被置于与另一核酸序列的功能关系中时,所述核酸即为“可操作连接”。例如,如果启动子(或者更确切地说是转录调节序列)影响编码序列的转录,那么该启动子就可操作连接到所述编码序列。可操作连接意味着所连接的dna序列通常是邻近的,并且在需要的时候将两个蛋白编码区连接为邻近且在开放阅读框内,以产生“嵌合蛋白”。“嵌合蛋白”或“杂合蛋白”是由多个蛋白“结构域”(或基序)组成的蛋白,其在天然条件下不存在,但是连接后形成了有功能的蛋白,显示出所连接的结构域的功能性。嵌合蛋白还可以是两个或多个天然存在的蛋白的融合蛋白。
本文中使用的术语“结构域”是指具有具体结构或功能的任何蛋白部分或区,其可被转移到另一蛋白以形成具有至少所述结构域的功能特征的新的杂合蛋白。具体结构域还可被用于识别属于slarf9蛋白家族的蛋白质成员,例如来自番茄植株的slarf9变体或来自其他植物物种的slarf9的直系同源物。slarf9蛋白中发现的结构域的实例有:含有seqidno:2的约氨基酸74-236的b3-衍生的dna-结合结构域或其变体、含有seqidno:2的约氨基酸237-564的中间区(mr)或其变体、含有seqidno:2的约氨基酸256-332的生长素应答区或其变体,或者分别含有seqidno:2的约氨基酸565-602或氨基酸609-651的二聚化结构域iii或iv或其变体。
术语“靶肽”是将蛋白质靶向至细胞内的细胞器例如质体(优选为叶绿体、线粒体)或至细胞外间隙(分泌信号肽)的氨基酸序列。可使编码靶肽的核酸序列融合(在阅读框内)到编码所述蛋白的氨基末端(n-末端)的核酸序列。
本文中“核酸构建体”或“载体”可理解为意指使用重组dna技术产生的、用于将外源dna递送到宿主细胞的人造核酸分子。例如,所述载体骨架可以是本技术领域公知的或者本文中他处描述的双元载体或超双元载体(见,例如us5591616、us2002138879和wo9506722)、共整合载体或t-dna载体,嵌合基因可整合到所述载体中,并且如果已经存在适合的转录调节序列,只需要在将需要的核酸序列(例如编码序列、反义序列或反向重复序列)整合到所述转录调节序列的下游。载体通常还包含方便其在分子克隆中使用的遗传元件例如选择标记、多克隆位点等(见下文)。
术语“宿主细胞”或“重组体宿主细胞”或“转化细胞”是指按如下得到的新个体细胞(或生物体):将至少一个核酸分子(特别是含有编码需要的蛋白的嵌合基因的核酸分子)或者转录时产生反义rna或反向重复rna(或发夹rna)用于沉默靶基因/基因家族的核酸序列导入所述细胞。所述宿主细胞优选为植物细胞或细菌细胞。所述宿主细胞可包含所述核酸构建体作为染色体外复制分子(附加体),或者更优选包含整合到所述宿主细胞核基因组或质体基因组中的嵌合基因。
术语“选择标记”是本领域技术人员公知的,在本文中用于描述在表达时可用于选择含有所述选择标记的一个或多个细胞的任何遗传实体。选择标记基因产物可提供例如抗生素抗性,或者更优选提供除草剂抗性或另外的选择性状例如表型性状(例如色素的改变)或营养需求。术语“报告子”主要是指可见的标记物例如绿色荧光蛋白(gfp)、egfp、萤光素酶、gus等。
术语基因或蛋白的“直系同源物”在本文中是指在另外的物种中发现的同源基因或同源蛋白,其具有与所述基因或蛋白相同的功能,但(通常)从含有所述基因的物种分化(即通过物种形成由共同的祖先进化而来的基因)的时间点开始出现序列分化。因此,可通过序列比较(例如,基于整个序列和/或具体结构域的百分比序列同一性)和/或功能分析在其他植物物种中鉴定番茄slarf9基因的直系同源物。
术语“同源的”和“异源的”是指核酸或氨基酸序列与其宿主细胞或宿主生物之间的关系(尤其是在转基因生物的情况下)。因此,同源序列是在宿主物种(例如用番茄基因转化的番茄植株)中天然存在的,而异源序列在天然条件下不存在于所述宿主细胞(例如用来自马铃薯植株的序列转化的番茄植株)中。依据上下文,术语“同源物”或“同源的”还可指来自共同的祖先序列的序列(例如它们可以是直系同源物)。
“严格杂交条件”可用于识别与给定的核苷酸序列基本上相同的核苷酸序列。严格条件是序列依赖的,并且在不同环境中是不同的。一般来说,将严格条件选择为比具体序列在确定的离子强度和ph下的热熔解点(tm)低约5℃。tm是(在确定的离子强度和ph下)50%的靶序列与完全配对的探针杂交的温度。通常将严格条件选择为:盐浓度为约0.02m(ph7),温度为至少60℃。降低盐浓度和/或增加温度将增加严格性。例如,rna-dna杂交(使用例如100nt的探针的rna印迹)的严格条件包括在63℃用0.2×ssc洗涤20min至少一次的条件或等价条件。例如,dna-dna杂交(使用例如100nt的探针的dna印迹)的严格条件包括在至少50℃(通常约55℃)的温度下用0.2×ssc洗涤20min至少一次(通常为2次)的条件或等价条件。也可参见sambrooketal.(1989)以及sambrookandrussell(2001)。
“序列同一性”和“序列相似性”可通过使用全局或局部比对算法比对两个肽序列或两个核苷酸序列来确定。然后,当序列至少共有某个最小百分比序列同一性(如下文中进一步定义)时(通过例如程序gap或bestfit或emboss程序“needle”(使用缺省参数,见下文)进行最佳比对时),所述序列就称为“基本相同”或“基本相似”。这些程序使用needleman和wunsch全局比对算法来在其全长上比对两个序列,使匹配数目最大化并使空位数目最小化。一般来说,使用缺省参数,其中空位产生(gapcreation)罚分=10,空位扩展(gapextension)罚分=0.5(对于核苷酸和蛋白质比对二者)。对于核苷酸,使用的缺省得分矩阵为nwsgapdna,对于蛋白质,使用的缺省得分矩阵为blosum62(henikoff&henikoff,1992,pnas89,915-919)。例如,百分比序列同一性的序列比对和得分可使用计算机程序例如gcgwisconsinpackage(10.3版,可得自:accelrysinc.,9685scrantonroad,sandiego,ca92121-3752usa)或者emboss(http://www.ebi.ac.uk/tools/webservices/services/emboss)来确定。或者,可通过搜索数据库例如fasta、blast等确定百分比相似性或同一性,但是应将找到的目标取回并进行两两比对以比较序列同一性。
在本文档及其权利要求书中,动词“包含”及其变化用其非限制性含义,意指包括该词之后所列的项,但不排除未具体提及的项。此外,除非上下文中清楚地要求有且只有一个要素,否则通过不定冠词“一”或“一个”表示的要素不排除存在超过一个所述要素的可能性。因此,不定冠词“一”或“一个”通常表示“至少一个”。还可以理解,本文所指的“序列”通常是指具有具体亚单位(例如氨基酸)序列的实际物理分子。
本文中使用的术语“植物”包括完整的植物或其任何部分或衍生物,例如植物器官(例如收获的或者未收获的贮藏器官、鳞茎、根茎、果实、叶等)、植物细胞、植物原生质体、可再生为完整植株的植物细胞或组织培养物、植物愈伤组织、植物细胞团以及完整植物中的植物细胞、植物的部分例如胚、花粉、胚珠、子房、果实(例如收获的组织或器官例如收获的番茄或其部分)、花、叶、种子、根茎、鳞茎、克隆繁殖植物、根、根状茎、茎、根尖等。并且,还包括任何发育阶段,例如未成熟或成熟的茎等。
“植物品种”是处于已知最低等级的相同植物学分类内的一组植物,其可基于来自某基因型或基因型组合的特征的表达进行定义(无论是否满足《植物育种者权利》(plantbreeder'srights)中的识别条件),可通过表达这些特征中的至少一个与任何其他植物组相区别,可被看作一个实体,因为其可以不发生任何改变地繁殖。因此,如果一组植物的特征是含有一个基因座或基因(或由该单个基因座或基因产生的一系列表型特征),另外对于其他基因座或基因彼此可以有很大的差异,那么即使它们属于同类,也不能使用术语“植物品种”来表述该组植物。
“f1、f2等”是指两个亲本植株或亲本株系之间杂交后的连续相关世代。由两个植株或株系杂交产生的种子长成的植株称为f1代。f1代植株自交产生f2代,等等。“f1代杂种”植株(或f1代种子)是使两个近交的亲本株系杂交得到的世代。“m1群”是某植物株系或栽培种的多个被诱变处理的种子/植株。“m1、m2、m3、m4等”是第一代被诱变处理的种子/植株(m1)自交后得到的连续世代。
术语“等位基因”是位于具体基因座的基因的一种或多种其他形式中的任一种,位于具体基因座的所有等位基因与一种性状或特征相关。在生物体的二倍体细胞中,给定基因的等位基因位于染色体上的具体位置或基因座。一个等位基因位于一对同源染色体中的每条染色体上。二倍体植物物种在某个基因座上可包含大量不同的等位基因。这些等位基因可以是所述基因的相同等位基因(同源的)或者两个不同等位基因(异源的)。
术语“基因座”是指染色体上的存在例如基因或遗传标记物的一个或多个具体位置或位点。因此,slarf9基因座是基因组中存在slarf9基因的位置。不意在限制本发明的,所述slarf9基因座被认为位于番茄基因组中的8号染色体。
“野生型等位基因”(wt)在本文中是指编码功能性蛋白(野生型蛋白)的基因形式。番茄栽培种moneymaker的野生型slarf9等位基因如seqidno:1(mrna/cdna)和seqidno:3(基因组dna,具有核苷酸2005到5879的slarf9编码区)所示。番茄栽培种heinz1706的野生型slarf9等位基因如seqidno:4(基因组dna,具有核苷酸1196到5869的slarf9编码区)所示。本文中“突变体等位基因”是指与所述野生型等位基因相比在编码序列(mrna、cdna或基因组序列)中含有一个或多个突变的等位基因。所述突变(例如,一个或多个核苷酸的插入、倒位、缺失和/或置换)可导致所编码蛋白具有降低的体外和/或体内功能性(功能降低)或者无体外和/或体内功能性(功能丧失),例如由于所述蛋白例如被截短或者具有其中一个或多个氨基酸被缺失、插入或置换的氨基酸序列。这些改变可导致蛋白质具有不同的3d构象、被靶向不同的亚细胞区室、具有改良的催化结构域、具有改良的对核酸或蛋白质的结合活性等。
“更大的果实大小”或“显著增大的果实大小”、“显著更大的果实大小”或“产生显著更大果实的植物”在本文中是指与适合的对照植物(例如野生型植物)相比果实大小(平均值)显著更大,即平均中纬直径和/或平均体积和/或平均鲜果实重显著高于对照。优选在果实生长期结束时(即当所述果实达到其最终大小时)或之后(在足大小果实期,例如破色期)确定所述果实大小。
“更多果皮细胞”或“具有更多细胞的果实”在本文中是指果皮(包括表皮、外果皮、中果皮和内果皮的细胞层)中果皮细胞的数目和/或细胞层的数目(平均值)显著多于对照。优选在细胞分裂期结束时(例如,番茄为约10dap或其后)和/或果实生长期结束时或其后(在足大小果实期,例如番茄的破色期)确定所述细胞数目。
“较小果皮细胞”是指果皮细胞(特别是中果皮细胞)的平均细胞大小显著小于对照(例如野生型果实)。优选在细胞分裂期结束时(例如,番茄为约10dap或其后)和/或果实生长期结束时或其后(在足大小果实期,例如破色期)确定所述细胞大小数值。
“野生型植物”和“野生型果实”在本文中是指含有编码功能性蛋白的野生型(wt)slarf9等位基因的植物(例如,与含有突变型slarf9等位基因的“突变型植物”相比)。这种植物是例如表型测定中适合的对照。优选地,野生型植物和/或突变型植物是“栽培的植物”,即人工栽培并具有很好的农艺特征的种的品种、育种系或栽培种;优选所述植物不是“野生植物”(即通常具有比栽培植物差得多的产量和农艺特征并且例如在野生群中自然生长的植物)。“野生植物”包括例如种的生态型、pi(植物引种)系、当地品种或者野生种质或野生近缘种,或者所谓的祖传品种或栽培种(即通常在人类历史的较早时期种植但是现代农艺中不再使用的品种或栽培种)。
slarf9基因或蛋白质的“变体”包括番茄种或番茄的野生近缘种中发现的天然的等位变体,以及在其他植物物种例如其他双子叶植物物种或单子叶植物物种中发现的直系同源物。
番茄的野生近缘种包括:s.arcanum、克梅留斯基番茄(s.chmielewskii)、小花番茄(s.neorickii=l.parviflorum)、契斯曼尼番茄(s.cheesmaniae)、s.galapagense、细叶番茄(s.pimpinellifolium)、智利番茄(s.chilense)、多腺番茄(s.corneliomulleri)、多毛番茄(s.habrochaites=l.hirsutum)、s.huaylasense、赤茄(s.sisymbriifolium)、秘鲁番茄(s.peruvianum)、多毛番茄(s.hirsutum)或者潘那利番茄(s.pennellii)。
本文中“平均值”是指算数平均数。
具体实施方式
发明人通过对已传粉的子房和ga3(赤霉素)处理的子房(vriezenetal.2008,newphytologist177:60-76)进行转录组分析(cdna-aflp),研究了在番茄果实形成过程中差异表达的基因。通过产生转基因植物,对在传粉的子房中高表达的一个基因进行了分析,以研究该基因的功能。该番茄基因被命名为slarf9(番茄arf9),因为其编码的658个氨基酸的蛋白质的氨基酸序列与拟南芥arf9蛋白的氨基酸序列最相似(用emboss程序“needle”,氨基酸序列同一性为52%,cdna序列同一性为61%,其中空位空缺(gapopening)=10、空位扩展=0.5)。
slarf9被发现在传粉的子房的胚珠、胎座和果皮中高表达(见图1)。更详细的分析表明,slarf9也在其他植物组织例如叶腋分生组织和根分生组织中转录。一般而言,这些是许多细胞分裂发生的组织。slarf9mrna水平增加的转基因植物形成了比野生型果实小的果实。而由于细胞分裂活性的增加,slarf9mrna水平减少的转基因株系的果实形成较大的果实。所述表达分析以及转基因株系的表型表明,slarf9在果实形成过程中充当细胞分裂的抑制因子。值得注意的是,通过在camv35s启动子控制下组成型表达rnai分子使slarf9mrna水平减少,没有引起任何其他负面表型。因此,尽管有slarf9基因不被认为是果实特异的基因的事实,但是rnaislarf9株系只显示出果实表型,这表明在其他植物组织中slarf9可能与arf蛋白家族的其他成员的作用有冗余。
slarf9涉及果实大小的发现可被用于通过减少果实生长过程中野生型slarf9蛋白(或其变体或直系同源物)的含量和/或野生型蛋白(或其变体或直系同源物)的功能,产生具有更大果实的转基因植物和/或非转基因植物,这将在下文进一步描述。具体地,据此增强果实生长过程中的细胞分裂,导致果皮中显著更多的细胞和/或显著更多的细胞层,和/或果实中显著更小的细胞。因此,植物产生具有更多固体组分的更大果实。
通过如本文中他处所述,在植物中过表达编码slarf9功能性蛋白(或其变体)的slarf9基因、变体或直系同源物,该发现也可用于相对于对照(例如野生型植物)显著减小果实大小。
下文及非限制性实施例中描述了本发明的不同实施方案。除非另有说明,否则本文中描述的适用于转基因方法的部分通常也可适用于非转基因方法,反之亦然。
在一个实施方案中,提供了至少在果实生长过程中内源slarf9表达下调或沉默并且产生显著更大的果实的转基因植物。在另一个实施方案中,本文提供了非转基因植物,其含有一个或多个突变型slarf9等位基因(以纯合或杂合的形式),并且其中所述突变型等位基因编码与野生型蛋白相比体外和/或体内功能性降低或者甚至没有功能性(例如通过翻译终止密码子或者移码突变)的slarf9蛋白,并且所述突变藉此产生与缺少所述突变型等位基因的植物(野生型植物)相比具有显著更大果实的植物(突变系或其子代)。在本发明的一个实施方案中,所述非转基因的、产生大果实的植物使用tilling或eco-tilling产生或鉴定,但也可以使用其他已知的诱变方法结合育种方法产生或鉴定。因此,在一个实施方案中,使用诱变技术人工诱导和/或鉴定所述突变型slarf9等位基因(“诱导的突变体”),而在本发明的另一个实施方案中,所述突变型slarf9等位基因是“天然突变体”,也就是说其是从天然的植物群(例如番茄的野生近缘种)中鉴定并被导入优质种质的。“诱导的突变体”优选在栽培种质中产生,并因此直接存在于具有农艺价值的株系中。另一方面,“天然突变体”或“自发突变体”或“天然变体”或“天然等位变体/变种”是基于种内发现的天然变种(多态性/突变),因此其可能存在于农艺质量较差、现代农业不栽培的植物材料例如野生植物中。然后,需要将天然突变体的等位基因转化到具有优良农艺特征的栽培植物中,这是本发明的一个实施方案。
用于本发明用途的核酸序列和蛋白质
在本发明的一个实施方案中,提供了slarf9的核酸序列和氨基酸序列,以及用于分离或鉴定其“变体”的方法,所述“变体”例如种(番茄)内或茄属(例如野生番茄近缘种例如潘那利番茄、多毛番茄等)内的等位基因变体,或者其他植物物种例如其他蔬菜种(例如茄科内的种例如辣椒或茄子;或葫芦科内的种例如甜瓜、西瓜或黄瓜)或田间作物物种(例如玉米、小麦、水稻)的slarf9直系同源物。
来自番茄栽培种moneymaker(生鲜市场番茄)的野生型slarf9转录因子蛋白如seqidno:2所示。它是包含数个结构域的658氨基酸蛋白,所述结构域即:a)位于n-末端区的dna-结合结构域(seqidno:2的氨基酸74-236),其可能结合到生长素调节基因的启动子区的顺式调节元件,b)中间区(“mr”,seqidno:2的氨基酸237到564),和c)两个二聚化结构域:结构域iii(seqidno:2的氨基酸565-602)和结构域iv(seqidno:2的氨基酸609到651)。
加工番茄栽培种heinz1706的所述蛋白如seqidno:4所示。其与seqidno:2的蛋白有99.8%的序列同一性,因为它仅包含一个不同氨基酸。最后一个氨基酸(氨基酸658)在栽培种moneymaker(seqidno:2)中是组氨酸(his),在栽培种heinz1706中是丝氨酸(ser)。因此,编码heinz1706蛋白的基因可认为是生鲜市场栽培种moneymaker中发现的该基因的等位基因变体。
“slarf9蛋白”(包括其“变体”,例如由所述基因的等位基因变体或所述基因的直系同源物编码的蛋白)可通过与seqidno:2在全长上的氨基酸序列同一性进行定义,即与seqidno:2具有至少约55%、60%、70%、80%、90%、95%、98%、99%或更高的序列同一性(通过使用emboss“needle”的两两比对确定,blossum62矩阵,空位空缺罚分=10、空位扩展罚分=0.5)并具有与slarf9基本相似的体内功能的蛋白。
本文还包括野生型slarf9蛋白(或如上所定义的其变体)的功能丧失的突变体或者野生型slarf9蛋白(或其变体)的功能降低的突变体(如别处所述),以及包含编码所述突变体的一个或多个核苷酸序列的植物或植物部分,所述突变体等位基因藉此使所述植物与含有编码野生型slarf9蛋白的核酸序列的植物相比产生显著更大的果实。显著更大的果实优选是重量(平均值)为野生型植物的果实的平均鲜果重的至少约105%,优选至少约110%、120%、130%、140%、150%或更大的果实。还任选地,平均果实直径是野生型植物果实直径的至少105%、110%、115%或更大。
还任选地,果皮组织中的平均细胞数目和/或果皮组织的细胞层数目显著多于对照(例如野生型植物的果实)。并且,果皮细胞(特别是中果皮细胞)的平均细胞大小优选显著小于对照。优选地,平均果皮细胞数目(按单位面积度量,见实施例)是对照的至少约105%、110%、120%、130%、140%、150%、160%或更多。优选地,所述细胞层数目是对照的至少105%、110%、120%或更多。还优选地,果皮细胞(特别是中果皮细胞)的平均大小是对照的平均细胞大小的至少约95%、92%、90%、85%、75%、72%或更小。
“与slarf9的功能基本相似的功能”在本文中是指所述蛋白具有证实的决定果实大小的功能。在至少发育中的果实组织中过表达slarf9或其变体的植物与对照(例如野生型植物或者空载体转化的植物)相比(平均)产生显著更小的果实。反之亦然,在至少发育中的果实组织中全功能(野生型)slarf9蛋白或其变体水平下降的植物与对照(例如野生型植物或者空载体转化的植物)相比(平均)产生显著更大的果实。如实施例中所示,slarf9过表达株系的平均果实重量是野生型重量的75%,平均果实直径是野生型的90%。与之相反,slarf9沉默株系的平均果实重量是野生型重量的125%,直径是野生型果实直径的大于105%。
因此,(推定的)slarf9蛋白的功能可以使用多种已知的方法测试,例如通过比较组成型表达要测试蛋白的转化体的表型与相同宿主种(和品种)的slarf9过表达转化体(优选包含稳定整合到所述宿主基因组中的嵌合slarf9编码基因)的表型,使得可直接比较对所述转化体表型的功能效应。
相似地,可使用slarf9基因(或其变体)沉默或者下调的转化体(例如,与野生型或对照转化体相比,至少在发育中的果实组织中slarf9mrna显著减少)来确定所述功能。slarf9转录本mrna“显著减少”是指存在的靶mrna水平小于或等于所述野生型或对照转化体(例如空载体转化体)中存在的转录本水平的90%、80%、70%、60%、50%、40%、30%、20%或更小(10%、5%或0%)。可以理解,在任何转化体实验中,转化体出现某种程度的表型变异通常是由于基因组中的位置效应和/或由于拷贝数。本领域技术人员能够知晓如何对转化体进行相互比较,例如通过选择单拷贝数事件并分析其表型。确定或验证体内基因/蛋白质功能的其他方法包括产生敲除突变体或功能下降突变体(例如通过tilling)或者瞬时表达研究。启动子-报告基因表达研究也可提供关于所述蛋白的时空表达模式和功能的信息。
上述方法可用于测试任何推定的slarf9基因例如来自番茄野生近缘种、栽培番茄植株、番茄育种系、pi(植物引种)系或者不同物种(例如,西瓜或其他果实或蔬菜物种或者田间作物物种)的等位基因是否的确为slarf9变体或直系同源物,然后可将其用于产生与适合的对照例如野生型植物相比产生(显著)更大果实的转基因植物和/或非转基因植物。可以理解,对于转基因植物,优选转化和再生具有良好农艺特征的植物,即栽培植物(例如高产的栽培种或育种系),最适合的对照是相同株系的空载体转化体或非转化株系本身的多株植物。
本文提供的序列可用于鉴定、产生和/或分离来自其他番茄植株、番茄的野生近缘种或其他物种的直系同源物的其他slarf9或slarf9等位基因。因此,所述序列可用于产生和/或鉴定在其基因组中含有一个或多个突变型slarf9等位基因(所述突变藉此导致显著增大的果实大小)的植物。因此,在一个实施方案中,提供了slarf9基因用于鉴定和/或产生含有一个或多个突变型slarf9等位基因、能够提供更大果实大小的植物的用途。
优选地,本发明的植物含有一个或多个突变型slarf9等位基因(或其变体),并且其较野生型植物产生更大的果实,但不产生更少的果实。因此,每株植物的果实数目优选不减少。在本发明的一个实施方案中,本发明的植物在果实生产季节结束时产生的果实与主要果实生产季节的野生型果实大小相似或者比其更大。在番茄中,由于环境条件导致在生产季节结束时野生型植物的果实大小较小。本发明的植物可补偿该季节结束效应。
可以通过计算机(insilico)来鉴定其他推定的slarf9基因/蛋白,例如通过在已有的核酸或蛋白质数据库(例如genbank、swissprot、trembl)中使用标准的序列分析软件例如序列相似度搜索工具(blastn、blastp、blastx、tblast、fasta等)鉴定核酸序列或蛋白质序列。选择、克隆或者从头合成含有slarf9蛋白(如上文定义的)的推定氨基酸序列或者编码slarf9蛋白的推定核酸序列,并通过例如在植物宿主中过表达和/或沉默测试其体内功能性。需要注意,标识slarf9在本文中也用于来自番茄之外的物种的蛋白,也就是说前缀sl在本文中并不将所述蛋白限定为来自具体的物种。
在一个实施方案中,提供了功能下降的或功能丧失的突变型slarf9蛋白(包括所定义的变体或直系同源物),以及其基因组中含有一个或多个slarf9等位基因——其编码功能下降或功能丧失的突变体,由此当存在于植物基因组中时所述功能下降或功能丧失显著增大果实大小——的植物和植物部分。
任何类型的突变都可能导致所编码的slarf9蛋白功能下降或功能丧失,所述突变例如在cdna(seqidno:1或其变体)或在相应的基因组slarf9序列(seqidno:3的核苷酸2005-5879、seqidno:4的核苷酸1196-5869或它们的变体)中,特别是在slarf9蛋白(或其变体,包括直系同源物)的14个外显子序列(见seqidno:3和seqidno:4)的任一个和/或内含子/外显子边界中,一个或多个核苷酸的插入、缺失或置换。在一个优选的实施方案中,提供了能够增大果实大小的slarf9核酸序列,其中所述核酸序列因编码以下的区域中的一个或多个突变而功能下降或功能丧失的slarf9蛋白:dna结合结构域(由外显子3-8编码的seqidno:2的氨基酸74-236)、mr(由外显子8-12编码的seqidno:2的氨基酸237-564,在mr中由外显子8-10编码的氨基酸256-332是特别优选的)、二聚化结构域iii(由外显子12和13编码的seqidno:2的氨基酸565-602)和/或二聚化结构域iv(由外显子13和14编码的seqidno:2的氨基酸609-651)。
可以通过sift分析(paulinec.ngandhenikoff2003,nucleicacidresearchvol.31,pp3812-3814)来预测或者通过评价对果实大小(表型)的效应来确定突变对蛋白质功能的效应(功能下降或功能丧失)。
如上文所述,可通过确定该突变型等位基因在显著增大果实大小方面的效应来测试所述蛋白在体内的功能下降或功能丧失。如下文所述,可例如使用tilling产生或使用ecotilling鉴定含有编码所述功能下降或功能丧失的突变型蛋白的核酸序列并且产生显著更大果实的植物。
在本发明的一个实施方案中,编码所述突变型蛋白的(cdna或基因组)核酸序列含有一个或多个无义和/或错义突变例如转换(用另一嘌呤置换嘌呤
在一个实施方案中,提供了含有外显子2编码序列、外显子3编码序列、外显子4编码序列、外显子5编码序列、外显子6编码序列和/或外显子7编码序列中的一个或多个无义和/或错义突变的slarf9核苷酸序列,以及含有所述突变等位基因并产生比仅含(编码功能性slarf9蛋白的)野生型等位基因的植物显著更大果实的植物。
在本发明的一个具体的实施方案中,提供了含有功能丧失或功能下降的突变型slarf9等位基因的植物和植物部分(果实、种子等)。
在一个实施方案中,所述功能丧失或功能下降的slarf9蛋白是截短的蛋白,即上文进一步定义的slarf9蛋白(包括其变体)的任一种的蛋白片段。一般来说,ems(甲基磺酸乙酯)可诱导鸟嘌呤/胞嘧啶替换腺嘌呤/胸腺嘧啶。对于谷氨酰胺(gln或q,由核苷酸caa或cag编码)或精氨酸(arg或r,由核苷酸cga编码)的密码子,将胞嘧啶替换为胸腺嘧啶可导致在读码框中引入终止密码子(例如caa/cag/cga变为taa/tag/tga),从而产生截短的蛋白。
还提供了编码slarf9蛋白的核酸序列(基因组dna、cdna、rna),所述slarf9蛋白例如如上文定义的seqidno:2所示的slarf9或其变体(包括任何嵌合或杂种蛋白、突变蛋白或截短蛋白),或者来自其他种的任何slarf9蛋白。由于遗传密码的简并性,多种核酸序列可编码相同的氨基酸序列。将编码slarf9蛋白(如上文定义,包括其变体)的任何核酸序列在本文中称为slarf9。所提供的核酸序列包括天然出现的、人工的或者合成的核酸序列。在seqidno:1(cdna)和3(来自番茄栽培种moneymaker的基因组序列,核苷酸2005-5879是带有内含子的蛋白编码区)以及seqidno:4(来自番茄栽培种heinz1706的基因组序列,核苷酸1196-5869是带有内含子的蛋白编码区)中提供了编码slarf9的核酸序列。
可以理解,当序列以dna序列显示时,也涉及rna,rna分子的实际碱基序列除了胸腺嘧啶(t)被替换为尿嘧啶(u)之外是相同的。
还提供了编码突变的slarf9蛋白即如上文定义的功能下降或功能丧失的slarf9蛋白的核酸序列(基因组dna、cdna、rna),以及含有这种突变型序列的植物或植物部分。例如,slarf9核酸序列包含在野生型slarf9编码序列中的一个或多个无义和/或错义突变,使得所编码的蛋白在体内无功能或功能降低。还提供了含有其他突变的序列,例如剪接位点突变体,即导致mrna前体的异常剪接的基因组slarf9序列的突变,和/或移码突变,和/或一个或多个核酸的插入(例如转座子插入)和/或缺失。
本文还提供了两种野生型基因组slarf9序列,一个来自生鲜市场番茄栽培种moneymaker(seqidno:3),另一个来自加工栽培种heinz1706(seqidno:4)。如所提及的,除最后一个密码子不同(在seqidno:3中编码组氨酸,在seqidno:4中编码丝氨酸)外,其编码区是相同的。编码dna序列(不包括内含子)具有99.9%的序列同一性。由于在内含子序列中有一些核苷酸差异,包括内含子的基因组编码dna(seqidno:3的核苷酸2005-5879,seqidno:4的核苷酸1996-5869)具有99.8%的序列同一性。所述基因的启动子(seqidno:3的核苷酸1-2004,seqidno:4的核苷酸1-1995)也具有98.7%的高序列同一性。
还包括slarf9核酸序列的变体及片段,例如在所定义的严格杂交条件下可与slarf9核酸序列例如seqidno:1或3(核苷酸2005-5879)杂交的核酸序列。slarf9核酸序列的变体还包括与seqidno:1、seqidno:3(核苷酸2005-5879)或seqidno:4(核苷酸1996-5869)的具有以下序列同一性的核酸序列:至少60%或更高,优选至少65%、70%、80%、85%、90%、95%、98%、99%或更高(通过emboss“needle”使用缺省参数(即,空位产生罚分=10,空位扩展罚分=0.5,评分矩阵nwsgapdna)确定)。所述变体还包括如所述的突变型slarf9变体。很显然,许多方法可用于鉴定、合成或分离slarf9核酸序列的变体或片段,例如核酸杂交、pcr技术、计算机分析和核酸合成等。seqidno:1、seqidno:3(核苷酸2005-5879)或seqidno:4(核苷酸1996-5869)的变体既可编码野生型的功能性slarf9蛋白(例如来自其他番茄品种或育种系或野生新种(accession)的等位基因,或者来自其他非番茄物种的直系同源物),也可编码所述蛋白中任一种的功能下降或功能丧失的突变型等位基因(例如通过例如tilling或ecotilling或者其他方法产生或鉴定的)。
片段包括上文slarf9核酸序列(或变体)的任一种的部分,其可以例如用作引物或探针或者基因沉默构建体,或者用于检测突变型slarf9等位基因(例如用于tilling的引物,例如引物seqidno:15-22)。部分可以是编码链(正义链)或互补链(反义链)的长度至少约10、15、19、20、21、22、23、24、25、50、60、100、200、300、420、450、500、600、700、800、900或更多个核苷酸的连续片段(stretch)。还包括所述片段的正义-反义构建体,其在植物细胞中转录时能够形成双链rna(任选在正义和反义片段之间有间隔序列)(见“基因沉默”)。因此,还包括slarf9核酸序列的片段,其中长度至少约10、15、20、30、40、50、60、100、150、200、300、400、420、450、500、600、700、800、900个核苷酸的片段与大约相同长度的slarf9核酸序列的另一片段有至少50%、60%、70%、75%,更优选地至少80%、90%、95%、98%、99%或更高(100%)的核酸序列同一性(通过emboss“needle”使用缺省参数(即,空位产生罚分=10,空位扩展罚分=0.5,评分矩阵nwsgapdna)两两比对确定)。
引物对可用于从植物组织dna样本pcr扩增slarf9或slarf9转录本(mrna或相应cdna或基因组dna)。引物对可用于检测和/或定量植物组织例如番茄果实组织中的slarf9或slarf9表达(mrna水平),或者例如用于确定内源slarf9mrna水平显著是否降低或基因组中是否存在突变型slarf9等位基因。同样,可基于seqidno:1或seqidno:3设计特异性引物或简并引物,并将所述引物用于从/在其他番茄株系(或在其中)、番茄的野生近缘种或其他植物物种扩增/检测slarf9的等位基因变体(例如,突变型slarf9等位基因)。
一旦产生并/或鉴定含有具体突变型slarf9等位基因的植物组织(例如通过tilling或ecotilling),即可设计所述突变型等位基因特异的引物或探针,并开发检测植物或植物部分中存在和/或不存在所述突变型等位基因的测定法(使用等位基因特异的检测测定法)。可开发用于检测并/或转移所述突变型等位基因的分子标记物测定法(例如,通过mas——标记物辅助选择)。例如,可开发检测植物的dna中突变型slarf9核酸序列存在情况并/或可用于将所述等位基因转移进入其他植物的snp检测测定法或caps标记物。
在一个实施方案中,提供了突变型slarf9核酸序列,其中所述slarf9核酸序列含有可导致slarf9蛋白功能丧失突变体或者slarf9蛋白的功能下降突变体的一个或多个突变。
还可鉴定或产生(例如通过同源重组,或者通过插入、缺失或置换一个或多个核苷酸,等等)植物,所述植物在slarf9调节区(例如启动子)中有一个或多个突变,藉此所述植物中的slarf9基因表达即(seqidno:1或变体的)mrna水平与野生型相比显著下降,并且藉此所述植物与含有野生型调节区(例如启动子)的野生型植物相比产生显著更大的果实。
slarf9的启动子区在seqidno:3的核苷酸1-2004和seqidno:4的核苷酸1-1995显示。这两个启动子序列有98.7%的序列同一性。野生型番茄slarf9启动子在果实发育阶段的早期(在传粉的子房的果皮、胚珠、胎座中)、叶腋分生组织、根尖和侧根原基中具有活性。seqidno:3和4两者在启动子区都含有两个生长素应答元件(auxre),分别位于seqidno:3的位置612-617(tgtcnc)和1224-1229(tgtctn)以及seqidno:4的位置612-617(tgtcnc)和1229-1234(tgtctn)。此外,seqidno:3还含有4个ntbbf1arrolb-元件(acttta,在位置888-893、1541-1546、1824-1829和1831-1836),而seqidno:4只含有三个这种元件(在位置888-839、1815-1820和1822-1827)。这些顺式活性调节元件可通过其他arf和/或dof-类蛋白提供转录调节。这些元件中的突变可减少slarf9转录本的产生,导致具有更大果实的植物。因此,在一个实施方案中,提供了在内源(野生型)slarf9启动子区特别是在一个或多个auxre元件和/或ntbbf1arrol元件中含有一个或多个突变的植物、植物部分或组织,这些植物藉此产生显著更大的果实。例如,可通过tilling产生这些植物。具体地,本文包括auxre元件中的g突变和/或ntbbf1arrolb元件中的c突变(转换或颠换)。例如,通过tilling或其他已知的方法可鉴定以下植物:slarf9启动子中含有突变,藉此所述启动子的活性至少在果实发育早期显著降低,并且藉此所述植物产生显著更大的果实。
本发明的“slarf9启动子”是与seqidno:3的核苷酸1-2004或seqidno:4的核苷酸1-1995有至少80%,优选至少90%、95%、98%、99%或100%的核酸序列同一性(使用缺省参数的两两比对程序needle)的启动子。在体内的天然环境中,这种启动子可驱动本发明的slarf9核酸序列或变体(例如野生型slarf9或突变型slarf9基因)的表达。本文包括突变型slarf9启动子(和含有这些启动子的植物)。
本文还包括含有slarf9启动子的嵌合基因和载体以及含有slarf9启动子的转基因植物,所述启动子可操作地连接到蛋白编码核酸或基因沉默构建体(正义和/或反义序列)。所述启动子可用于在发育的果实中表达嵌合基因。slarf9启动子的至少约2000、1700、1600、1500、1200、100、800、600、500、400、300个核苷酸的活性片段(例如通过5'末端制造启动子缺失获得的)也适合于果实优选的表达。
可将本发明的上文所述的slarf9核酸序列或其片段,特别是编码所述slarf9蛋白的dna序列(或它们的变体)插入到表达载体(在共抑制方法中)或基因沉默载体以产生具有更大果实的植物。
在本发明的一个实施方案中,通过例如rnai方法(如他处所述)可下调宿主细胞、植物或具体组织中的slarf9基因表达。
在另一个实施方案中,提供含有一个或多个内源突变型slarf9等位基因的植物,所述突变使植物与缺少所述突变型等位基因的植物相比具有更大的果实大小。突变型等位基因优选通过以下方式产生:诱变所述植物或种子,并鉴定这样的植物或种子,即这些植物或种子在靶slarf9等位基因中含有一个或多个突变,由此所述突变导致mrna转录和/或翻译减少或终止(使得减少slarf9蛋白产生或无slarf9蛋白产生),或者导致转录突变型slarf9等位基因(其被翻译成功能下降或功能丧失的slarf9蛋白)。至少在果实组织中(优选至少在果实发育阶段的早期)野生型slarf9蛋白的功能下降或终止赋予植物或种子产生显著更大果实的能力。在本发明的一个实施方案中,所述植物含有两个内源突变型slarf9等位基因,即是slarf9纯合的。这种植物可通过含有单个突变型slarf9等位基因的植物自交产生。还提供了在其基因组中含有至少一个或两个突变型slarf9等位基因的果实和种子,由此所述果实比含有野生型slarf9等位基因(编码功能性slarf9蛋白)的植物的果实显著更大。
在本发明的另一个实施方案中,提供了用于检测slarf9或slarf9dna序列的pcr引物和/或探针和试剂盒。如本领域所公知的,可基于seqidno:1或seqidno:3或4合成用于从样本中扩增slarf9或slarf9dna的简并pcr引物对或特异性pcr引物对(见dieffenbachanddveksler(1995)pcrprimer:alaboratorymanual,coldspringharborlaboratorypress,andmcphersonatal.(2000)pcr-basics:frombackgroundtobench,firstedition,springerverlag,germany)。同样,seqidno:1或seqidno:3(或其变体)的dna片段可用作杂交探针。slarf9或slarf9检测试剂盒可包含slarf9和/或slarf9特异的引物以及/或者slarf9和/或slarf9特异的探针,以及使用所述引物或探针检测样本中slarf9和/或slarf9dna的相关方案。例如,这种检测试剂盒可用于确定植物是否被本发明的slarf9基因(或其部分)转化或者植物是否含有一个或多个突变型slarf9等位基因。由于遗传密码的简并性,一些氨基酸密码子可被另一些密码子代替而不改变所述蛋白的氨基酸序列。
在另一个实施方案中,提供了可特异性结合到本发明的slarf9蛋白或突变型slarf9蛋白的抗体。特别地,本文包括结合到slarf9或结合到其片段或变体(例如突变型蛋白质)的单克隆抗体或多克隆抗体。使用本领域公知的方法,可在动物中通过使用本发明的slarf9蛋白作为抗原来制备抗体,如例如harlowandlane"usingantibodies:alaboratorymanual"(newyork:coldspringharborpress1998)和liddellandcryer"apracticalguidetomonoclonalantibodies"(wileyandsons,1991)中所述。所述抗体可随后用于分离、鉴定、表征或纯化其结合的slarf9蛋白,例如以检测样本中的slarf9蛋白,使得可形成免疫复合物并通过例如elisa(酶联免疫测定)或免疫印迹分析来检测所述免疫复合物的存在情况。还提供了可用于检测样本中slarf9蛋白、蛋白片段或表位的免疫试剂盒。样本可以是细胞、细胞上清液、细胞悬液、组织等。所述试剂盒至少包含可结合到slarf9蛋白的抗体以及一种或多种免疫检测试剂。所述抗体还可被用于例如通过elisa或蛋白质印迹分离/鉴定其他slarf9蛋白。
显然,许多方法可用于鉴定、合成或分离slarf9或slarf9核酸序列的变体或片段,例如核酸杂交、pcr技术、计算机分析、核酸合成等。因此,slarf9-蛋白编码核酸序列可以是化学合成的序列或者从任何植物物种克隆的序列。
具有更大果实的转基因植物
提供了这样的转基因植物、种子和植物部分,即在它们中slarf9(优选至少在果实组织中)被沉默,并且由于所述slarf9基因沉默它们与野生型(非转基因)对照植物和其他对照植物(例如空载体转化体)相比产生显著更大的果实。
在本发明的一个实施方案中,同源或异源的核酸序列用于沉默将被转化的宿主物种的内源slarf9基因。例如,马铃薯slarf9基因(或其变体或片段)可用于在转基因番茄、茄子或西瓜植株中沉默slarf9基因表达。或者,可使用同源slarf9核酸序列。例如,将来自某种植物物种(例如来自番茄)的序列重新引入所述物种(番茄)中。
因此,在一个实施方案中,slarf9dna与用作转化宿主的物种的内源slarf9dna相一致,或者是其修饰产物/变体。因此,优选使用番茄slarf9cdna或基因组dna(或其变体或片段)来转化番茄。此外,(由于监管部门的批准和公众接受的原因)可将同源或异源核酸序列可操作地连接到转录调节序列,特别是启动子,其也来自植物物种或者甚至来自与要转化的植物相同的植物。例如,上文定义的slarf9启动子可用于番茄中。
为产生含有嵌合基因(其表达时导致内源slarf9基因或基因家族的表达沉默)的植物,可使用本领域公知的方法。
“基因沉默”是指宿主细胞或组织中一个或多个靶基因例如slarf9基因的表达的下调或完全抑制。应理解,在任何转化实验中均会出现某种程度的转化体表型差异,这通常是由于在基因组中的位置效应和/或由于拷贝数。一般来说,本文区分“弱”和“强”基因沉默植物(都是本发明的实施方案),其中“弱”基因沉默(rnai)事件是指与对照组织相比内源靶基因表达下降约15%、20%或30%的植物或植物部分,“强”基因沉默(rnai)事件是指与对照组织(例如野生型)相比内源靶基因表达下降至少约50%、60%、70%、80%、90%或更大的植物或植物部分。沉默可通过例如定量所述靶基因的转录水平(例如使用定量rt-pcr)和/或通过确定并任选定量靶蛋白的酶活性和/或通过评价并任选定量所产生的表型(更大果实大小)来定量。
并非限制本发明的范围,可以选择具有最佳沉默水平的植物,使得形成的植物在果实发育过程中所暴露的环境条件下与对照相比具有显著更大的果实,同时具有最小的不良副作用例如产量降低等。优选地,在所述slarf9沉默的植物中产量显著提高。
使用抑制性rna来减少或终止基因表达是本领域常用的,并且是多篇综述的主题(例如,baulcombe1996,plantcell8(2):179-188;depickerandvanmontagu,1997,curropincellbiol.9(3):373-82)。有多种可用的技术来实现植物中的基因沉默,例如产生全部或部分靶基因的反义rna的嵌合基因(见,例如ep0140308b1、ep0240208b1和ep0223399b1),或者产生靶基因的正义rna的嵌合基因(也称为“共抑制”),见ep0465572b1。
然而,迄今为止最成功的方法是产生靶基因的正义rna和反义rna两者(“反向重复”),在由上游启动子转录时,它们在细胞中形成双链rna(dsrna)或茎环结构(发夹rna,hprna)并沉默靶基因。ep1068311、ep983370a1、ep1042462α1、ερ1071762a1和ep1080208a1中描述了dsrna和hprna产生以及基因沉默的方法和载体。
因此,用于植物转化的嵌合基因可包括转录调节区,所述转录调节区在植物细胞中具有活性并且可操作地连接到slarf9靶基因或基因家族的正义和/或反义dna片段(或完整核酸序列)或者与slarf9靶基因或基因家族互补的正义和/或反义dna片段(或完整核酸序列)或与slarf9靶基因或基因家族基本相似的正义和/或反义dna片段(或完整核酸序列)。
一般来说,靶基因序列的短(正义和反义)片段,例如靶基因的编码序列和/或非编码序列的17、18、19、20、21、22、23、24、25或26个核苷酸是足够的。也可使用更长的序列,例如至少约50、100、200、250、300、400、420、450、500、1000或更多个核苷酸。甚至与完整转录本rna或mrna一致或互补的dna也可用于产生正义和/或反义构建体。优选地,所述正义和反义片段/序列被间隔序列例如内含子分隔,所述间隔序列在dsrna形成时形成环(或发夹)。
原则上,可将任何slarf9基因或基因家族作为靶标。例如,通过选择对这些等位基因特异的其一级转录本或mrna转录本的核酸区,可沉默一个或多个具体slarf9等位基因(见byzovaetal.plant2004218:379-387中以器官特异性方式进行等位基因特异性沉默)。类似地,通过选择转录本中一个或多个保守区用于沉默构建体,可将整个基因家族作为沉默的靶标。如上所述,以正义和/或反义方向使用的dna区不需要是编码区的一部分,但也可与所述一级转录本的部分(含有5'和3'非翻译序列和内含子;番茄品种moneymaker的slarf9的一级转录本示于seqidno:3中,核苷酸1551到6323,编码区为核苷酸2005到5879)或者mrna转录本的部分(已除去任何内含子并已添加polya尾)一致或互补。可以理解,在与rna序列相对应的dna序列中,u替换为t。还需要注意,在可转录在宿主细胞中转录时可靶向并能够沉默slarf9基因表达的dsrna或hprna的嵌合基因中,正义和反义区的长度不需要相等,其中一个区可长于另一个区。
因此,例如seqidno:1或如上文所述的其变体、它们中任一个的片段或seqidno:3或4的基因组序列或一级转录本序列可用于产生slarf9基因沉默基因和载体,以及其中一个或多个slarf9基因在全部或一些组织或器官中(优选至少在发育中的果实中)或者诱导时被沉默(见,例如,wielopolskaetal.plantbiotechnolj.20056:583-90)的转基因植物。实施例中给出了基因沉默载体的实例,其中使用编码seqidno:1的一部分的中间区(mr)的反向重复420bp片段来沉默番茄中的内源slarf9,使用在camv35s启动子控制下组成型表达。
产生发夹结构的一个便利方式是使用通用载体例如基于
通过选择保守的核酸序列,可沉默宿主植物中所有slarf9基因家族成员。沉默宿主植物中所有基因家族成员是一个实施方案。
在一个实施方案中,可操作连接到所述正义和/或反义核酸序列(以制备嵌合沉默基因/rnai基因)的启动子选自组成型启动子、诱导型启动子(例如化学诱导型启动子等)、激素诱导型启动子(例如生长素诱导型)、果实特异型启动子或发育调节的启动子(例如在果实发育中有活性)。同时,slarf9基因本身的启动子,即如上文定义的slarf9启动子,也可用于沉默方法。任选地,可将3'utr可操作地连接到所述嵌合基因的3'端,使得所述可操作连接的dna元件包括启动子-slarf9rnai基因-3'utr。
优选的组成型启动子包括:花椰菜花叶病毒(camv)分离株cm1841(gardneretal,1981,nucleicacidsresearch9,2871-2887)、cabbb-s(francketal,1980,cell21,285-294)和cabbb-ji(hullandhowell,1987,virology86,482-493)的强组成型35s启动子或增强的35s启动子(所述“35s启动子”);odell等(1985,nature313,810-812)或us5164316中描述的35s启动子,来自泛素家族的启动子(例如christensenetal,1992,plantmol.biol.18,675-689、ep0342926、以及cornejoetal1993,plantmol.biol.23,567-581中的玉米泛素启动子),gos2启动子(depateretal,1992plantj.2,834-844),emu启动子(lastetal,1990,theor.appl.genet.81,581-588),拟南芥肌动蛋白启动子例如an等(1996,plantj.10,107)描述的启动子,水稻肌动蛋白启动子例如zhang等(1991,theplantcell3,1155-1165)描述的启动子,以及us5,641,876中描述的启动子或wo070067中描述的水稻肌动蛋白2启动子;木薯叶脉花叶病毒启动子(wo97/48819,verdagueretal1998,plantmol.biol.37,1055-1067)、来自地三叶草矮化病毒的pplex系列启动子(wo96/06932,特别是s7启动子)、醇脱氢酶启动子例如padh1s(genbank登录号x04049、x00581)、以及分别驱动t-dna的1'和2'基因表达以及驱动t-dna表达(veltenetal,1984,emboj3,2723-2730)的tr1'启动子和tr2'启动子(分别为“tr1'启动子”和“tr2'启动子”)、us6051753和ep426641中描述的玄参花叶病毒启动子、组蛋白基因启动子例如来自拟南芥的ph4a748启动子(pmb8:179-191),等等。
或者,可使用非组成型但是对植物中一个或多个组织或器官特异的启动子(组织优选的/组织特异的,包括发育调节的启动子)。例如,可以使用在果实组织和/或果实发育中具有活性的启动子,例如slarf9启动子或其活性片段。或者,可使用子房和幼小果实特异的tptp-f1启动子(carmietal.200,见上文),等等。
在根据本发明的方法中,本领域技术人员能够很容易地测试多种启动子的特异性和适应性。此外,通过缺失、插入或置换部分的所述启动子序列可改变启动子的特异性。可将所述改性的启动子可操作地连接到报告基因,以测试它们在转基因植物中的时空活性。
另一个可选实施方案是使用其表达可诱导的启动子。诱导型启动子的实例是化学诱导启动子,例如aoyamaandchua(1997,plantjournal11:605-612)以及us6063985中描述的地塞米松诱导的启动子,或者四环素诱导的启动子(topfree或top10启动子,见gatz,1997,annurevplantphysiolplantmolbiol.48:89-108,以及loveetal.2000,plantj.21:579-88)。
任选地,启动子-slarf9rnai基因还可包含3'端转录调节信号(“3'端”或“3'utr”)(即转录本形成和多腺苷酸化信号)。多腺苷酸化和转录本形成信号包括在转化的植物细胞中充当3'-非翻译dna序列的胭脂碱合成酶基因的信号(“3'nos”)(depickeretal.,1982j.molec.appl.genetics1,561-573)、章鱼碱合成酶基因的信号(“3'ocs”)(gielenetal.,1984,emboj3,835-845)和t-dna基因7的信号(“3'基因7”)(veltenandschell,1985,nucleicacidsresearch13,6981-6998),等等。同时,可使用slarf9基因的3'端,即含有seqidno:3的核苷酸5880到6323或由seqidno:3的核苷酸5880到6323组成的序列。
可将所述嵌合slarf9沉默基因(即可操作连接到在植物细胞中转录时能够沉默内源slarf9基因表达的核酸序列的启动子)以常规方式稳定地插入到单个植物细胞的核基因组中,并且可以常规方式使用所转化的植物细胞来产生具有改变的表型的转化植物,所述改变的表型由于在某段时间在某些细胞中的slarf91沉默而导致。在这点上,可将含有可操作连接到正义和/或反义slarf9序列的启动子(以及任选的3'utr)的t-dna载体引入到农杆菌中,并用其转化植物细胞,然后使用例如ep0116718、ep0270822、pct公开wo84/02913以及公开的欧洲专利申请ep0242246中和gould等(1991,plantphysiol.95,426-434)中描述的方法,从所述转化植物细胞产生转化植物。用于农杆菌介导的植物转化的t-dna载体是本领域公知的。所述t-dna载体可以是ep0120561和ep0120515中描述的双元载体,或者是如ep0116718中所述可通过同源重组整合到农杆菌ti-质粒中的共整合载体。
每个优选的t-dna载体均含有与位于t-dna边界序列之间或者至少位于右边界序列的左边的slarf9沉默基因可操作连接的启动子。gielen等(1984,emboj3,835-845)描述了边界序列。当然,也可以使用其他类型的载体来转化植物细胞,使用的方法例如直接基因转化(例如ep0223247中所述)、花粉介导的转化(例如ep0270356和wo85/01856中所述)、原生质体转化(例如us4,684,611中所述)、植物rna病毒-介导的转化(例如ep0067553和us4,407,956中所述)、脂质体介导的转化(例如us4,536,475中所述)以及其他方法,例如用于转化某些玉米系的记载方法(例如e.g.,us6,140,553;frommetal,1990,bio/technology8,833-839;gordon-kammetal,1990,theplantcell2,603-618)和转化某些稻的记载方法(shimamotoetal,1989,nature338,274-276;dattaetal.1990,bio/technology8,736-740)以及通常用于转化单子叶植物的方法(pct公开wo92/09696)。棉花转化还可见wo00/71733、稻转化还可见w092/09696、w094/00977和w095/06722中描述的方法。高粱转化见例如jeoungjmetal.2002,hereditas137:20-8或者zhaozyetal.2000,plantmolbiol.44:789-98。番茄或烟草转化还可见ang.etal.,1986,plantphysiol.81:301-305;horschr.b.etal,1988,in:plantmolecularbiologymanuala5,dordrecht,netherlands,kluweracademicpublishers,pp1-9;koornneefm.etal,1986,in:nevinsd.j.andr.a.jones,eds.tomatobiotechnology,newyork,ny,usa,alanr.liss,inc.pp169-178。土豆转化见例如shermanandbevan(1988,plantcellrep.7:13-16)。还可依据dejongetal.(2008)plantjournal57:160-170和sunetal.(2006)plantcellphysiol.47:426-431进行番茄转化和再生。
同样,从转化细胞选择和再生转化植物是本领域公知的。显然,对于不同物种、甚至对于同一物种的不同品种或栽培种,可具体地调整实验方案以高频率再生转化体。
除转化核基因组之外,本发明还包括转化质体基因组,优选叶绿体基因组。质体基因组转化的一个优点在于其可降低转基因扩散的风险。可使用本领域公知的方法进行质体基因组转化,见例如sidorovvaetal.1999,plantj.19:209-216或者lutzkaetal.2004,plantj.37(6):906-13。
任何植物都可作为适合的宿主,例如单子叶植物或者双子叶植物,但是最优选可从产生更大的果实受益的植物,例如但不限于:番茄、辣椒、黄瓜、茄子、甜瓜、西瓜、南瓜、西葫芦、葡萄以及许多其他植物,例如玉米、小麦、稻、高粱、向日葵、果树、草莓、柑橘类(citrusfruit)、豆、豌豆、大豆等。基本上,本文中的宿主包括由子房产生可食用果实(植物学意义上)的任何开花植物物种。特别优选的是肉质果实物种(产生具有肉质果皮的果实)。
优选的宿主是茄科(solanaceae)宿主,例如茄属(solanum)的种,例如番茄(s.lycopersicum)、树番茄(s.betaceum,syn.cyphomandrabetaceae)以及其他茄属的种如茄子(solanummelongena)、凤果(s.muricatum)、科科纳果(s.sessiliflorum)和奎东茄(s.quitoense)。茄科还包括辣椒(capsicumannuum,capsicumfrutescens)。
在一个优选的实施方案中,所述宿主属于茄科或葫芦科(cucurbitaceae)。在一个更优选的实施方案中,所述宿主属于茄属。在一个甚至更优选的实施方案中,所述宿主属于番茄种。优选地,所述宿主是番茄种的栽培番茄,即高产量的种系或品种,例如果实平均鲜重为至少50g或更高,例如至少约80g、90g、100g、200g、300g或者甚至达到600g(牛肉番茄型)。还包括小类型,例如樱桃番茄或开胃番茄(cocktailtomato),以及全果肉番茄,例如纽海姆公司的番茄品种intense,例如种子空腔中无凝胶。所述宿主番茄植株可具有确定的或不确定的不同果实大小和形状,例如罗马型(romatype)、簇型(clustertype)、圆形。其可以是加工型番茄或者生鲜市场型番茄。同时,本文包括自由传粉和杂交的番茄。在一个实施方案中,所述番茄植株是由f1代杂种种子长成的f1代杂种植株。为产生本发明的转基因植物的f1代杂种种子,可产生两个近交亲本系,其基因组中各含有一个拷贝的转基因。当这些植物交叉受精后,收集f1代种子,其由于所述转基因而产生高产量且大果实的转基因f1代杂种植物。
本文中“宿主”植物的实施方案也适用于本文中他处描述的非转基因的突变型植物,其基因组中存在内源的突变型slarf9等位基因而非转基因。
其他适合的宿主是产生肉质果实的其他植物物种和多个物种(葡萄、桃、李、草莓、芒果、木瓜等)。葫芦科植物例如甜瓜(citrulluslanatus,cucumismelo)和黄瓜(cucumissativus)以及南瓜和葫芦(cucurbita)也是适合的宿主。同样,蔷薇科(rosaceae)例如苹果、梨、李等也是适合的宿主。
本发明还提供了具有更大(slarf9沉默或突变的slarf9)或更小(slarf9过表达)果实(植物学意义上的)的田间植物。例如,玉米/谷物(玉蜀黍种,例如玉米、大刍草(chapule)、繁茂大刍草(危地马拉玉米草)、委委特南戈类玉米亚种(圣安东尼奥huista玉米草)、墨西哥类玉米亚种(墨西哥玉米)、小颖类玉米亚种(巴尔萨斯玉米草)、z.perennis(多年生玉米草)和z.ramosa)、小麦(小麦种)、大麦(例如hordeumvulgare)、燕麦(例如avenasativa)、高粱(二色高粱)、黑麦(secalecereale)、大豆(大豆属例如g.max)、棉花(棉花种,例如陆地棉、海岛棉)、芸苔属(例如油菜、芥菜、花椰菜、芜菁等)、稻(稻种,例如籼稻栽培种或粳稻栽培种)、珍珠粟(狼尾草属属例如p.glaucum)。
基本上,任何作物植物物种都是适合的。本文中作物植物或栽培植物是指人工栽培和培育的植物物种,不包括杂草例如拟南芥或野生近缘种例如番茄近缘种和其他近缘种(尽管可从这种植物中得到突变型slarf9等位基因并通过育种方法将其转移到栽培植物中,详见下文)。作物植物可以为食物或饲料(例如蔬菜作物或田间作物)目的或者为观赏目而栽培。本文中定义的作物植物还包括可收获非食物产品(例如用作燃料的油、塑料聚合物、药制品、软木、纤维等)的植物。
因此,在本发明的一个实施方案中,转基因植物含有与在宿主细胞中转录时能够沉默内源slarf9基因表达的核酸分子可操作连接的转录调节元件(特别是上文描述的启动子)。
构建用于将slarf9沉默基因导入(优选稳定地导入)宿主细胞基因组的嵌合基因和载体是本领域公知的。为产生嵌合基因,用常规分子生物学技术将正义和/或反义slarf9序列可操作地连接到适于在宿主细胞中表达的启动子序列。所述启动子序列可以是已经存在于载体中的,因此可简单地将所述核酸序列插入载体中所述启动子序列的下游。然后,可使用所述载体来转化宿主细胞,并将所述嵌合基因插入核基因组或质体(线粒体或叶绿体)基因组,并使用适合的启动子而表达(例如mcbrideetal,1995bio/technology13,362;us5,693,507)。
可将形成的转化植物用于常规的植物育种方案中,以产生更多的具有相同特征的转化植物,或者将所述基因部分导入相同或相近植物物种的其他品种。可选择“良种事件(eliteevent)”,其为所述转基因插入基因组中具体位点并且所述插入导致所需要表型(例如最佳的沉默和大果实大小)良好表达的转化事件。
其中slarf9被沉默的转基因植物或其部分具有显著更大的果实,优选在果皮组织中具有显著更多的细胞和/或显著更多的细胞层和/或显著更小的细胞。本文中使用的显著更大的果实(如上文所述)是指与对照相比平均果实重量和/或(任选)果实直径和/或果实体积增大。例如,可在果实生长期结束当果实达到最终的大小时比较果实重量。果实直径对于圆形果实很容易测量,但是难以与其他形状比较。通过例如测量容器中被果实替代的水的液体(例如水)的体积或其他方法,可很容易地确定果实体积。
可以理解,当分析突变体植物的表型时,对照植物优选为所述突变体的近等基因系,其含有野生型等位基因。
最后,可使用田间试验显示与野生型植物相比产生显著更大果实的转化体(或下文描述的突变型植物)。
如已提到的,可通过例如分析拷贝数(dna印迹分析)、mrna转录水平(例如使用slarf9引物对的rt-pcr)或者通过分析多个组织中slarf9蛋白存在情况和/或水平(例如sds-page、elisa测定等)选择具有最佳沉默水平的转化体。然后,将最佳的转基因事件用于后续的杂交/回交/自交直到获得具有稳定转基因的高性能良种事件。
表达一个或多个本发明slarf9基因(或基因沉默)的转化体也可含有其他的转基因,例如提供干旱抗性或者生物或非生物胁迫抗性、除草剂抗性等的基因。为获得这种具有“叠加的”转基因的植物,可将其他转基因整合进所述slarf9转化体,或者可后续用一个或者多个其他基因转化所述转化体,或者也可使用几个嵌合基因转化植物株系或品种。例如,几个嵌合基因可存在于单个载体中,或者存在于共转化的不同载体中。也可将含有几个嵌合基因的微型染色体导入植物中(见,例如,yuetal.2007,pnas104:8924-9,和houbenandschubert2007,plantcell19:2323-2327)。
任意上文所述转化植物的完整植株、种子、细胞、组织和子代(例如f1、f2种子/植株等)包括在本文中,并且可以通过dna中存在情况进行鉴定,例如通过pcr分析。还可开发“事件特异的”pcr诊断方法,其中pcr引物是基于插入的嵌合基因侧翼的植物dna,见us6563026。类似地,可开发鉴定所述转基因植物或者其衍生的任何植株、种子、组织或细胞的事件特异的aflp指纹或rflp指纹。
可以理解,本发明的转基因植物优选不显示不希望的表型,例如果实质量下降、每个植株果实减少、对疾病的敏感性增加或者不希望的株型改变(矮化、畸形)等,并且如果这种表型出现在一级转化体中,可通过常规的育种和选择方法(杂交/回交/自交等)将其除去。本文所述的任何转基因植物可以是纯合的或者半纯合的所述转基因。
产生更大果实的非转基因植物及制备所述植物的方法
本发明的另一个实施方案使用非转基因方法例如靶突变体产生和鉴定系统如tilling(targetinginducedlocallesionsingenomics;mccallumetal,2000,natbiotech18:455,andmccallumetal.2000,plantphysiol.123,439-442,henikoffetal.2004,plantphysiol.135:630-636,其通过引用纳入本文)并进行选择,以产生这样的植物株系,即这些植物株系在内源slarf9等位基因中含有至少一个突变,以此含有纯合的或杂合的所述突变型slarf9等位基因的植物与不含所述突变型等位基因的植物(其在slarf9基因座具有野生型等位基因)相比产生显著更大的果实。因此,在本发明的一个实施方案中,本文提供了基因组中含有一个或多个突变型slarf9等位基因并且与不含所述突变型等位基因而含有野生型等位基因的植物相比产生显著更大果实的植物,以及含有所述突变型等位基因的这类植物的植物部分(例如收获的果实、收获的叶等)、种子、克隆繁殖产物、后代。
术语“显著更大的果实”在本文中是指与适合的对照植物(例如野生型植物)相比果实大小(平均)显著更大,即平均中纬直径和/或平均体积和/或平均鲜果实重显著大于对照。优选地,在果实生长期结束或其后(即当所述果实达到其最终大小时)确定所述果实大小。例如,在番茄中,通过在果实生长期结束(例如破色期)测定平均果实鲜重和/或通过测量平均中纬直径并将所得值与对照(来自基因组中含有野生型slarf9等位基因的植物的果实)比较,可确定每个植株中至少约5、8、10、15、20或更多个果实的果实大小。产生显著更大果实的植物可产生例如具有对照果实的至少约105%、110%、115%、120%、130%、140%、150%或更大的平均重量的果实。任选地或可选择地,平均中纬直径可以是所述对照果实的平均中纬直径的至少约105%、110%、115%、120%或更大。
如他处所述,任选地,与对照(例如野生型果实)相比,果皮组织中的细胞数目和/或细胞层数目也显著更多,并且/或者细胞大小显著更小。
不意在限制本发明,由于果实发育过程中(即,特别是果实发育的细胞分裂期过程中,例如番茄受精后的头10-14天)细胞分裂增加,应认为含有编码无功能或功能下降的slarf9蛋白的突变型slarf9等位基因的植物的果实显著更大并具有更多的细胞。
优选地,上文所述产生更大果实的植物是纯合的突变型slarf9等位基因,但杂合体植物也可产生显著更大的果实。为产生含有纯合形式的突变型等位基因的植物,可使用自交,任选结合基因分型(例如通过使用等位基因特异的引物pcr和/或通过测序检测所述突变型等位基因的存在情况)。如果使用tilling群,则优选使所述突变型植物(m1)自交一次或多次以产生例如m2群,或者优选m3或m4群用于表型分析。在m2群中,所述突变型等位基因以1(纯合的突变型等位基因):2(杂合的突变型等位基因):1(纯合的野生型等位基因)的比例存在。果实大小的分离与所述突变型等位基因的分离相关。
含有所述突变型slarf9等位基因或其变体并产生显著更大果实的植物可以是任何物种,因为本文提供的番茄序列可用于产生和鉴定在所述基因的同源基因和直系同源物中含有突变的植物,如下文进一步所述。可鉴定所述植物中的内源slarf9变体核酸序列,然后可将其用作产生和/或鉴定含有slarf9变体的突变型等位基因的植物中的靶基因。因此,所述突变型植物(即,含有突变型slarf9等位基因的植物)可以是双子叶植物物种或单子叶植物物种。
优选地,所述植物是栽培植物,但鉴定野生型植物或非栽培植物中突变型等位基因并通过育种技术将其转移到栽培植物中也是本发明的一个实施方案。
在一个实施方案中,含有至少一个突变型slarf9等位基因(纯合体或杂合体形式)并具有显著更大果实的所述植物是茄科(即包括茄属、辣椒属、烟草属等)或葫芦科(其包括黄瓜属种例如甜瓜和黄瓜)。在另一个实施方案中,所述植物是番茄属,例如包括栽培的番茄、土豆、茄子等。
在一个具体的实施方案中,所述植物是番茄种。可产生和/或鉴定其基因组中具有至少一个突变型slarf9等位基因并因此产生更大果实的任何番茄。因此,所述番茄植株可以是任何栽培番茄、任何商业品种、任何育种系等,其可以是确定的或不确定的、自由传粉或杂交的,并可产生任何颜色、形状和大小的果实。可通过育种(与含有突变型等位基因的植物杂交,然后选择含有所述突变型等位基因的子代)容易地将在具体番茄植株或番茄的性相容近缘种中产生和/或鉴定的突变型等位基因转移到任何其他的番茄植株中。
所述植物可以是茄科或茄属的任何种,所述种即可自身诱变产生突变型等位基因(例如,通过tilling或其他方法),或者在所述种中例如通过ecotilling鉴定slarf9基因(或变体)中的一个或多个天然或自发突变。
使用锌指核酸酶在内源slarf9基因中产生突变也是本发明的一个实施方案,如townsendetal.2009,nature459:442-445和shuklaetal.2009,nature459:437-441中所述。
在一个实施方案中,在栽培植物中产生或鉴定所述突变型等位基因,但是也可在野生型植物或非栽培植物中产生和/或鉴定所述突变型等位基因然后使用例如杂交和选择(任选使用种间杂交与例如胚拯救来转移所述突变型等位基因)将其转移到栽培植物中。因此,可在其他番茄种中产生(使用诱变技术诱变所述靶slarf9基因或其变体的人工诱导突变)和/或鉴定(自发的或天然的等位基因变异)突变型slarf9等位基因,然后通过常规育种技术将其转移到栽培的茄属植物例如番茄中,所述番茄种包括例如番茄的野生近缘种,例如契斯曼尼番茄(s.cheesmanii)、智利番茄、多毛番茄(s.habrochaites,l.hirsutum)、克梅留斯基番茄、s.lycopersicumxs.peruvianum、s.glandulosum、多毛番茄(s.hirsutum)、小花番茄(s.minutum)、小花番茄(s.parviflorum)、潘那利番茄、秘鲁番茄、s.peruvianumvar.humifusum和细叶番茄。术语“常规育种技术”在本文中包括育种者公知的杂交、自交、选择、双单倍体产生、胚拯救、原生质体融合、通过中间物种转移等,即遗传改性之外的可转移等位基因的方法。
优选地,slarf9等位基因的突变使所述植物与不含所述突变型等位基因(即含有野生型slarf9等位基因)的植物相比具有显著更大的果实,如上文所述。
不意在限制本发明,slarf9(seqidno:1或其变体,或者相应基因组序列例如seqidno:3的核苷酸2005-5879或seqidno:4的核苷酸1196-5869的基因组序列或其变体)的突变可导致slarf9蛋白功能下降或功能丧失,例如通过单个碱基的转换、错义或无义突变,或者插入或缺失一个或多个氨基酸,或者编码序列的移码,slarf9蛋白功能下降或功能丧失会进而导致表型改变。使用slarf9和/或slarf9特异的引物,通过基因测序可以分析突变的存在情况和类型。优选地,通过杂合的突变型等位基因或者优选纯合的突变型等位基因的植物的表型(即显著更大的果实),在体内间接确定slarf9蛋白的功能“显著下降”。所述果实大小增大的表型与所述突变型等位基因共分离。
在本发明的一个实施方案中,提供了植物(优选番茄植株),其在seqidno:1、seqidno:3(核苷酸2005-5879)或seqidno:4(核苷酸1196-5869)中或者在与这些序列的任一个具有至少约60%、62%、65%、70%、80%、90%、95%、97%、97.5%、98%、98.5%、99%或更高的序列同一性的核酸序列(如所定义的)中含有一个或多个突变,由此所述突变导致所编码的slarf9蛋白(或变体)在体内具有降低的活性(与野生型功能性蛋白相比)或者无活性,并且其中所述植物与含有编码野生型功能性slarf9蛋白(或变体)的核酸序列的植物(优选为番茄)相比产生显著更大的果实。
在一个实施方案中,提供了植物(优选番茄植株),其在编码如下的蛋白的核苷酸序列中含有一个或多个突变:seqidno:2的蛋白或与seqidno:2有至少53%、54%、55%、58%、60%、70%、80%、90%、95%、98%、99%或更高序列同一性的蛋白(如所定义的),并且其中所述(番茄)植物与不含所述一个或多个突变的(番茄)植物相比具有显著更大的果实。
在一个实施方案中,提供了含有突变型slarf9等位基因的产生更大果实的植物(优选番茄植株),其特征在于所述突变是所编码的slarf9蛋白功能丧失或功能下降的突变,所述蛋白是与seqidno:2有至少53%、54%、55%、58%、60%、70%、80%、90%、95%、98%、99%或更高氨基酸序列同一性的蛋白。
所述植物(例如番茄植株)优选为突变型slarf9等位基因的纯合体。
在一方面,提供了在其基因组中,特别是在seqidno:3(或其变体)的任一外显子中(尤其是在外显子2和/或外显子7中),含有突变型slarf9等位基因(特别是具有一个或多个单点突变的等位基因)的番茄植株。一方面,所述突变位于以下氨基酸之一的编码序列中:seqidno:2的氨基酸52、191和/或193。本发明的一个实施方案是含有突变型slarf9等位基因并产生显著更大果实的番茄植株,其中所述突变型等位基因含有一个或多个下述突变(表示在前的野生型slarf9蛋白的氨基酸被转换为突变体的不同氨基酸,位置如下标所示):gly52→ser52、arg191→trp191或his193→tyr193。在一方面,所述突变位于编码slarf9蛋白保守结构域,特别是编码b3-衍生的dna结合结构域,即seqidno.2或其变体的氨基酸74到236,的序列中。在一方面,本文提供了含有突变型slarf9等位基因的番茄植株,如可从以ncimb41827、41828、41829、41839和/或41831保藏的种子获得(或者从来自所述种子的植物或者从这些植物的子代),其中所述番茄植株的果实显著大于在slarf9基因座具有野生型slarf9等位基因的番茄植株的果实。
在另一个实施方案中,所述含有内源突变型slarf9等位基因的植物是西瓜植株,结果是其与含有野生型slarf9等位基因的西瓜植株相比产生显著更大的西瓜果实。在另一个实施方案中,所述植物是黄瓜植株、甜瓜植株或辣椒植株。
通过分子方法例如slarf9基因组dna或mrna(cdna)中存在的突变、slarf9蛋白水平和/或蛋白活性等以及通过与野生型相比改变的表型特征,可将突变型植物与非突变体区分。通过常规的杂交和选择,可将所述突变型等位基因转移到与所述突变型植物性相容的其他植物。因此,可将所述突变型等位基因用于产生任何类型的大果实番茄品种例如自由传粉品种、杂交品种、f1代杂种、罗马型、樱桃型、确定或不确定的类型等。在一个实施方案中,含有突变型slarf9等位基因并具有显著更大果实的所述植物(优选番茄)是f1代杂种植物或可长成f1代杂种植物的f1代种子。用于产生f代杂种的近交亲本在它们的基因组中优选都含有相同的纯合形式的突变型slarf9等位基因。
在另一个实施方案中,将所述含有突变型slarf9等位基因的植物(例如番茄)与相同种或近缘种的另一植物杂交,以产生含有突变型slarf9等位基因的杂种植物(杂种种子)。所述杂种植物也是本发明的一个实施方案。还提供了将突变型slarf9等位基因转移到另一植物的方法,包括提供在其基因组中包含一个突变型slarf9等位基因的植物,由此所述突变型等位基因产生更大的果实(如上文所述),将所述植物与另一植物杂交并获得所述杂交的种子。任选地,可将这些种子获得的植物进一步自交和/或杂交,并选择含有所述突变型等位基因且由于所述突变型等位基因的存在而与含有野生型slarf9等位基因的植物相比产生显著更大果实的子代。
在一个实施方案中,用于产生f1代杂种的亲本均含有纯合形式的不同突变型slarf9等位基因,使得所述杂种含有两种不同的突变型slarf9等位基因。例如,亲本1可含有功能丧失的突变体,而亲本2含有功能下降的突变体。那么,所述f1代杂种含有来自每个亲本的一个等位基因。因此,本文还包括在slarf9基因座含有两个不同的突变型slarf9等位基因并且产生显著更大果实的番茄植株。
在一方面,从以ncimb41827、41828、41829、41839和/或41831保藏的种子(或者从来自所述种子的植物或者从这些植物的子代)获得的突变型等位基因在植物中可以是纯合的,或者可以与野生型等位基因或与另一突变型slarf9等位基因,例如从以ncimb41827、41828、41829、41839和/或41831保藏的种子(或者从来自所述种子的植物或者从这些植物的子代)获得的等位基因的任一个,相结合。因此,含有编码突变gly52→ser52的等位基因的番茄植株可与等位基因arg191→trp191或his193→tyr193结合。类似地,编码arg191→trp191和his193→tyr193的突变型等位基因可在同一植株中结合。同时,剪接位点突变型等位基因例如从以登录号ncimb41827保藏的种子获得的等位基因也可为纯合形式或杂合形式,并且任选与野生型slarf9等位基因或与另一突变型slarf9等位基因,例如从以ncimb41828、41829、41839和/或41831保藏的种子(或者从来自所述种子的植物或者从这些植物的子代)获得的编码突变gly52→ser52、arg191→trp191或his193→tyr193的等位基因,相结合。
通过使用诱变方法或通过从天然群中筛选slarf9等位基因的天然变体,可产生和/或鉴定含有编码功能丧失或功能下降的蛋白(例如因无义突变产生的截短蛋白;例如由于错义突变、移码突变和/或剪接位点突变而具有改变的氨基酸序列从而产生例如改变的催化位点、改变的折叠等的蛋白)的突变型slarf9等位基因的植物。在本发明的一个实施方案中,tilling用于产生这类植物和/或鉴定这类诱变诱导的突变,并且/或者ecotilling用于鉴定slarf9基因中含有天然(自发)突变的植物例如野生植物或非栽培植物,然后通过常规育种方法将所述slarf9基因转移到栽培植物中。但是,也可使用任何其他诱变方法,可以理解,本文包括在栽培植物或作物植物中产生或者通过常规育种转移到栽培植物或作物植物中的slarf9基因的人工诱导突变体(uv或x-射线诱变、化学诱变剂等)以及自发突变体两者。还可使用定向诱变(使用例如锌指内切核酸酶)来突变所述内源slarf9基因并产生编码功能下降或功能丧失的slarf9蛋白的slarf9等位基因,或者产生导致至少在果实发育过程中制备的slarf9蛋白减少或无slarf9蛋白的内源slarf9启动子突变。
在本发明的一个具体实施方案中,所述突变体植物(即含有突变型slarf9等位基因的植物)是与番茄不同种的植物,例如其基因组中含有突变型slarf9等位基因的单子叶栽培植物,优选稻、玉米、小麦或大麦,或者蔬菜物种或水果物种,例如西瓜、甜瓜、黄瓜、辣椒、南瓜、西葫芦、草莓、苹果、桃、樱桃、李、葡萄、柠檬、柑橘、梨、覆盆子、醋栗、蓝莓等。当使用例如tilling的方法时,可基于seqidno:1或其片段扩增靶基因片段(例如使用特异引物或简并引物,例如基于slarf9的一个或多个保守结构域设计的),或者可以首先分离slarf9直系同源物并基于所述直系同源序列设计引物。也可基于内含子序列或者内含子-外显子边界序列设计用于扩增靶基因片段的引物。例如,当在大外显子中筛选突变时,可使用两个pcr反应和两对引物扩增外显子,其中一条或多条引物可存在于所述外显子侧翼的内含子序列中。因此,引物对也可基于slarf9的基因组序列,例如seqidno:3(特别是核苷酸从约1977到5940或者2005到5879)和seqidno:4(特别是核苷酸从约1950到5909或者从1996到5869)所示的。
tilling(定向诱导基因组局部突变)是一种通用反向遗传学技术,其使用常规的化学诱变方法产生诱变个体文库,随后对所述文库进行高通量筛选来发现突变。tilling将化学诱变与混合的pcr产物的突变筛选相结合,形成靶基因的错义和无义突变型等位基因的分离。因此,tilling使用常规化学诱变(例如ems或mnu诱变)或其他诱变方法(例如辐射例如uv),然后高通量筛选具体靶基因例如本发明的slarf9中的突变。将s1核酸酶例如cel1或endo1用于切割突变型和野生型靶dna的杂合双链,并使用例如电泳如li-cor凝胶分析仪系统检测切割产物,见例如henikoffetal.plantphysiology2004,135:630-636。tilling已经被应用于许多植物物种,例如番茄(见http://tilling.ucdavis.edu/index.php/tomato_tilling)、稻(tilletal.2007,bmcplantbiol7:19)、拟南芥(tilletal.2006,methodsmolbiol323:127-35)、芸苔、玉米(tilletal.2004,bmcplantbiol4:12)等。在天然群中检测突变的ecotilling也被广泛使用,见tilletal.2006(natprotoc1:2465-77)和comaietal.2004(plantj37:778-86)。本发明的一个实施方案中,将经典的tilling进行改良,使用此前只用于人的两种不同的高通量检测系统代替基于酶的突变体检测(使用单链特异的核酸酶进行酶消化,并进行高分辨率的聚丙烯酰胺凝胶电泳)。这些检测方案是csce(构象敏感毛细管电泳,见rozyckaetal.2000,genomics70,34-40)或hrm(高分辨率熔解曲线,见clinchem49,853-860)的修改版本。见gadyetal.2009,plantmethods5:13。
因此,本文还包括在一个或多个组织中含有突变型slarf9等位基因并具有由本发明的功能下降或功能丧失的slarf9蛋白导致的一种或多种表型(例如如上述的更大果实)的非转基因植物、种子、果实和组织,以及产生和/或鉴定所述植物的方法。
还提供了产生和/或鉴定适于产生具有更大果实的植物的突变型slarf9等位基因的方法,和/或产生具有更大果实的植物的方法,包括步骤:
(a)诱变植物种子(例如通过ems诱变)以产生m1群,或者提供被诱变的植物种子或提供含有天然变异的植物,
(b)任选使(a)的植物自交一次或多次,以产生m2、m3或m4家庭成员,
(c)制备(a)或(b)的植物的dna,合并个体的dna,或者合并个体的组织样本并从所述合并的样本制备dna,
(d)从所述dna合并物或从来自所述合并的组织样本的dna,pcr扩增slarf9靶基因(基因组或cdna)或其变体的全部或部分,
(e)检测突变的slarf9等位基因在pcr扩增产物中的存在情况,由此在所述dna合并物中或在来自所述合并的组织样本的dna中的存在情况,
(f)选择含有突变型slarf9等位基因的相应个体植物,
(g)任选对所述植物的突变型slarf9等位基因进行测序;
(h)对(f)的植物或其后代就更大的果实大小产生进行表型分析,并且
(i)选择较对照产生更大果实的植物,并且任选地
(j)对(i)的植物育种以产生可产生更大果实并具有良好的农艺特征的栽培植物。
任选地在(e)和(f)步骤之间可进行sift分析,以预测哪些突变可在植物上产生更大的果实大小。
在步骤(d)中,使用常规方法例如coddle(http://www.proweb.org/doddle)设计扩增全部或部分所述靶基因(slarf9(seqidno:1、seqidno:3或4)或其变体)的引物。可设计引物以扩增所述靶基因(即seqidno:1、3或4,或者seqidno:1、3或4的变体)的例如至少约50bp、100bp、200bp、250bp、300bp、400bp、500bp、600bp、800bp或至少约1000bp或更多。优选地,通过引物扩增编码slarf9蛋白的保守结构域的全部或部分的片段,例如所述片段编码所述mr、二聚化结构域iii或二聚化结构域iv的dna-结合结构域的全部或部分的片段。
对于除番茄外的植物物种,需要首先鉴定内源slarf9基因的序列,以能够设计好的引物序列。可通过计算机或通过例如设计简并pcr引物并从所述植物物种的基因组中扩增所述slarf9基因变体(番茄slarf9基因的直系同源物)的全部或部分来鉴定所述序列。然后,可将所述内源slarf9基因的序列优选用于设计tilling的适合引物。
步骤(e)中可使用s1核酸酶,例如cel1,来检测所述pcr扩增产物之间(即形成杂合双链的所述野生型slarf9pcr产物和突变型slarf9pcr之间)的错配。或者,步骤(e)中可使用csce或hrm进行检测。在csce中,形成纯合双链(wt/wt或者突变体/突变体片段)以及杂合双链(突变体/wt片段)。由于形成了错配,杂合双链以与纯合双链不同的速率迁移通过毛细管,因此可鉴定靶片段中含有突变的合并物。hrm也是一种非酶的技术。在对所述靶基因片段的pcr扩增过程中,lcgreenplus+tm分子被整合到双链dna分子的每对退火的碱基对之间,其在被捕获到所述分子时会发出荧光。当板被逐渐加热时,使用光扫描仪记录荧光强度。在某温度下,所述pcr产物开始熔解并释放lcgreenplus+tm,从而所述荧光减弱。由于杂合双链的熔解温度低于纯合双链的熔解温度,所以可鉴定含有突变(杂合双链)的dna合并物。
步骤(j)可包含常规育种方法,以及表型和/或标记物辅助选择方法。该方式可产生含有一个或多个突变型slarf9等位基因并较含有一个或多个野生型slarf9等位基因的植物产生显著更大果实的许多不同品种。
已公开了许多进行tilling的方法,见例如http://blocks.fhcrc.org/%7esteveh/tilling_publications.html和tilletal.(2006)natureprotocols1:2465-2477;tilletal.(2006)methodsmolbiol.323:127-135,以及tilletal.(2003)methodsmolbiol.236:205-220,它们都以引用的方式纳入本文。
一旦鉴定了含有赋予需要的表型的突变型等位基因的植物,通过常规育种技术例如通过将所述植物与另一植物杂交并收集所述杂交的子代,可将所述等位基因转移到其他植物。因此,在步骤(j)中,所述等位基因可用于产生可产生大果实并提供好的农艺特征的植物。
如所述,可以理解,其他诱变和/或选择方法可等价地用于产生本发明的突变型植物。可以例如对种子进行辐射或化学处理以产生突变型群。也可对slarf9进行直接基因测序以从诱变的植物群筛选突变型等位基因。例如,keypoint筛选是可用于鉴定含有突变型slarf9等位基因的植物的基于测序的方法(rigolaetal.plosone,march2009,vol4(3):e4761)。
因此,提供了在一个或多个组织中(优选至少在果实组织中)产生较低水平的(功能性)野生型slarf9蛋白的非转基因突变型植物,或者在具体组织中完全缺乏功能性slarf9蛋白或者在某些组织中产生无功能slarf9蛋白的非转基因突变型植物,例如由于一个或多个内源slarf9等位基因的突变。可通过诱变方法例如tilling或其变型产生所述突变体,或者可通过ecotilling或其他任何方法鉴定所述突变体。可将编码无功能或功能下降的slarf9蛋白的slarf9等位基因分离并测序,或者可通过常规育种方法将其转移到其他植物。
提供了所述植物或其后代的任意部分,包括基因组中含有本发明的突变型slarf9等位基因的收获的果实、收获的组织或器官、种子、花粉、花、子房等。还提供了其基因组中含有突变型slarf9等位基因的植物细胞培养物或植物组织培养物。优选地,所述植物细胞培养物或植物组织培养物可再生为其基因组中含有突变型slarf9等位基因的完整植物。本文还包括通过含有slarf9突变型等位基因单倍体细胞染色体加倍产生的双单倍体植物(以及可长成所述双单倍体植物的种子),和其基因组中含有突变型slarf9等位基因的杂种植物(以及可长成所述杂种植物的种子),从而根据本发明,所述双单倍体植物和杂种植物产生显著更大的果实。
还提供了用于检测植物是否含有本发明的突变型slarf9等位基因的试剂盒。这种试剂盒可包含用于在组织样本或从所述组织获得的dna或rna中检测该等位基因的pcr引物或探针。
优选地,所述突变体植物还具有其他良好的农艺特征,即与野生型植物相比它们的果实数目不减少和/或果实质量不降低。优选地,由于所述植物的果实较大,所以其产量较高。同时,果皮组织中更多的细胞和/或更小的细胞大小导致其固体含量更高(每克鲜重具有更多的细胞壁)。因此,所述果实的可溶的和不溶的固体含量更高。在一个优选的实施方案中,所述植物是番茄植株,所述果实是番茄果实例如具有任何形状、大小或颜色的加工番茄、生鲜市场番茄。因此,还提供了含有一个或两个突变型slarf9等位基因的植物或植物部分的收获产品。所述产品包括下游的加工产品,例如番茄糊、番茄酱、番茄汁、切开的番茄果实、灌装果实、干果、去皮的果实等。这也适用于其他植物物种。通过其基因组dna中含有所述突变型等位基因,可以对所述产品进行鉴定。
本发明的其他用途
根据本发明,提供了编码slarf9蛋白的核酸序列用于改良植物果实大小的用途。在一个优选的实施方案中,该用途涉及改良(增加或减少)植物或具体植物部分中(例如至少在果实中)功能性slarf9蛋白的水平。
在一个实施方案中,本文提供了编码slarf9蛋白的核酸序列用于产生可产生大果实的转基因植物或非转基因植物的用途,其中所述slarf9蛋白与seqidno:2有至少60%、70%、75%、80%、85%、90%、95%、98%或99%的氨基酸序列同一性。本文中,该用途包括涉及在基因组中含有本发明的slarf9等位基因的植物、种子或者植物细胞或组织的用于产生或使用更大果实的任何用途。
类似地,提供了编码slarf9蛋白的核酸序列用于增大植物果实大小的用途,其中所述slarf9蛋白与seqidno:2有至少60%、70%、75%、80%、85%、90%、95%、98%或99%氨基酸序列同一性。
在一方面,本文提供了含有突变型slarf9等位基因的植物或种子用于产生增大的果实的用途,其中所述slarf9等位基因是编码与seqidno:2有至少60%、70%、75%、80%、85%、90%、95%、98%或99%氨基酸序列同一性的蛋白的等位基因。所述一个或多个突变的结果是,其基因组中含有所述突变型等位基因的植物与其基因组中含有野生型slarf9等位基因的植物相比产生显著更大的果实。
在另一方面,本文提供了含有突变型slarf9等位基因的体外植物细胞或组织培养物用于产生增大的果实的植物的用途,其中slarf9等位基因是编码与seqidno:2有至少60%、70%、75%、80%、85%、90%、95%、98%或99%氨基酸序列同一性的蛋白的等位基因。使用已知的方法,可将所述的植物细胞或组织培养物再生为完整植株。
同样,还提供了其基因组中含有突变型slarf9等位基因的增大的果实用于收获、储存、加工或出售的用途,其中slarf9等位基因是编码与seqidno:2有至少60%、70%、75%、80%、85%、90%、95%、98%或99%氨基酸序列同一性的蛋白的等位基因。
本文还提供了编码slarf9蛋白的核酸序列用于产生可产生更小果实的转基因植物或非转基因植物的用途。如本文中他处所述,slarf9蛋白的表达增加,所述slarf9蛋白是功能性蛋白并与seqidno:2有至少60%、70%、75%、80%、85%、90%、95%、98%或99%的氨基酸序列同一性。
所述植物优选是番茄属、辣椒属或黄瓜属。在一方面,所述植物优选为番茄、辣椒、黄瓜或甜瓜。
在一方面,所述突变型slarf9等位基因是从由以登录号ncimb41827、41828、41829、41830或41831保藏的种子长成的植物获得的等位基因。
因此,在一方面,本文提供了编码突变型slarf9等位基因的核酸序列用于产生可产生大果实的非转基因植物的用途,在一方面,所述突变型slarf9等位基因是来自上述种子保藏物的等位基因。
序列
seqidno1:来自番茄栽培种moneymaker的野生型slarf9等位基因的cdna序列
seqidno2:由seqidno:1编码的野生型slarf9蛋白的蛋白序列。氨基酸74–236包含b3-衍生的dna结合结构域。氨基酸237–564包含中间区(mr)。氨基酸256–332包含推定的生长素应答区。氨基酸565–602包含二聚化结构域iii。氨基酸609-651包含二聚化结构域iv。
seqidno3:来自番茄栽培种moneymaker的野生型slarf9的启动子区(核苷酸1-2004)和基因组dna。转录(mrna)起始点位于核苷酸1551,转录终止点位于核苷酸6323,翻译起始密码子是位于核苷酸2005-2007的atg,翻译终止密码子是位于核苷酸5877-5879的taa。因此,5'utr从碱基1551到2004,3'utr从碱基5880到6323。
seqidno4:来自番茄栽培种heinz1706的野生型slarf9的启动子区(核苷酸1-1995)和基因组dna。转录(mrna)起始点位于核苷酸1543,转录终止点位于核苷酸6313,翻译起始密码子是位于核苷酸1996-1998的atg,翻译终止密码子是位于核苷酸5867-5869的taa。因此,5'utr从碱基1543到1995,3'utr从碱基5870到6313。
seqidno5:肌动蛋白引物,正向
seqidno6:肌动蛋白引物,反向
seqidno7:slarf9引物,正向,用于mrna检测
seqidno8:slarf9引物,反向,用于mrna检测
seqidno9:slarf9引物,正向,用于编码序列扩增
seqidno10:slarf9引物,反向,用于编码序列扩增
seqidno11:slarf9引物,正向,用于rnai片段扩增
seqidno12:slarf9引物,反向,用于rnai片段扩增
seqidno13:启动子slarf9引物,正向,用于启动子扩增
seqidno14:启动子slarf9引物,反向,用于启动子扩增
seqidno15:用于从植物群中筛选外显子2中突变的正向引物
seqidno16:用于从植物群中筛选外显子2中突变的反向引物
seqidno17:用于从植物群中筛选外显子2中突变的正向引物
seqidno18:用于从植物群中筛选外显子2中突变的反向引物
seqidno19:用于从植物群中筛选外显子6中突变的正向引物
seqidno20:用于从植物群中筛选外显子6中突变的反向引物
seqidno21:用于从植物群中筛选外显子7中突变的正向引物
seqidno22:用于从植物群中筛选外显子7中突变的反向引物
附图说明
图1-番茄果实形成过程中的相对slarf9mrna水平
(a)在处理后1、2和3天(d),在胎座和胚珠组织、以及子房壁中实时定量pcr的slarf9的表达模式。总rna分离自去雄的花(对照)、赤霉酸处理的去雄的花(ga3)和人工传粉后去雄的花(传粉的)。
(b)在花发育的七个不同阶段收集的番茄子房中的相对slarf9mrna水平。1-4阶段示出了花芽大小。第4阶段代表去雄阶段的花。第5阶段,花完全开放(开花期)。第6阶段,开花3d(daa)收集的花。第7阶段,人工传粉后3d收集的花。示出了标准误差(n=2)。
(c)从开花期未传粉的番茄子房和人工传粉后3d的子房——分离成胚珠、胎座和子房壁组织样本——中的相对slarf9mrna水平。示出了标准误差(n=2)。
(d)生长素处理(iaa)6或24h后收集的去雄的花的番茄子房中的相对slarf9mrna水平。未处理的子房作为对照。示出了标准误差(n=2)。
(e)幼花芽、未传粉的子房、从去雄阶段的花收集的多个其他花器官、传粉的子房(3dap)以及10d幼苗的胚轴和根中的相对slarf9mrna水平。示出了标准误差(n=2)。以上(a)到(e)的每个图中,最大值设定为1。
图2:发育中的野生型果实和转基因果实中的slarf9mrna水平
(a)从野生型和slarf9-oe株系收集的子房和果实中的相对slarf9mrna水平。示出了标准误差(n=2)。3-4mm果实(6dap)中的野生型水平被设定为1。
(b)从野生型和rnaislarf9株系收集的子房和果实中的相对slarf9mrna水平。示出了标准误差(n=2)。3-4mm果实(6dap)中的野生型水平被设定为1。
图3:
来自slarf9-rnai植株(左)和野生型对照(右)的番茄果实的照片。
图4:
来自slarf9-rnai植株(左)和野生型对照(右)的番茄果实(10dap)的果皮细胞的显微照片。
下述非限制性实施例描述了slarf9和slarf9基因用于改良植物表型的用途。除非实施例中另有说明,否则所有的重组dna技术都依据sambrooketal.(1989)molecularcloning:alaboratorymanual,第2版,coldspringharborlaboratorypress和sambrookandrussell(2001)molecularcloning:alaboratorymanual,第3版,coldspringharborlaboratorypress,ny以及ausubeletal.(1994)currentprotocolsinmolecularbiology,currentprotocols,usa的卷1和2中所述的常规方法进行。用于植物分子工作的标准材料和方法见biosscientificpublicationsltd(uk)和blackwellscientificpublications,uk联合出版的,r.d.d.croy著的plantmolecularbiologylabfax(1993)中所述。
种子保藏
实施例3中的5种番茄tilling突变体的代表性种子样本由nunhemsb.v.依据布达佩斯条约的expertsolution(epc2000,rule32(1))于2011年4月14日保藏于英国食品工业与海洋细菌菌种保藏有限公司(ncimb有限公司)(英国苏格兰bucksburn阿伯丁郡craibstone区ferguson大厦,ab219ya(fergusonbuilding,craibstoneestate,bucksburnaberdeen,scotlandab219ya,uk))。种子的保藏号如下:ncimb41827(突变体1719)、ncimb41828(突变体2484)、ncimb41829(突变体3175)、ncimb41830(突变体6725)和ncimb41831(突变体6932)。
申请人要求,所述生物材料及源自其的任何材料的样本依据rule32(1)epc或具有类似条款和规章的国家或条约的相关法规只向指定的专业人员提供,直至授权公告或者提出申请之日起20年(如果所述申请被驳回、撤回或视为撤回)。
seqidno:1–番茄栽培种moneymaker的arf9编码序列
seqidno:2-番茄栽培种moneymaker的arf9蛋白
结构域(74)..(236)b3衍生的dna结合结构域
结构域(237)..(564)中间区(mr)
结构域(256)..(332)生长素应答区,中间区(mr)的一部分
结构域(565)..(602)二聚化结构域iii
结构域(609)..(651)二聚化结构域iv
实施例
实施例1-slarf9的分离和表征
1.1材料和方法
1.1.1植物材料和生长条件
番茄植株(番茄栽培种moneymaker)的种植如dejongetal.(2009,theplantjournal57,160-170)所述。同时,体外培养依照dejongetal.(2009,见上文)的方案进行。对于子房中slarf9的表达分析,在开花之前3天(d)将花去雄。人工传粉或激素处理在开花期进行。在以2μl的2%乙醇中1mm4-cl-iaa(sigma-aldrich,http://www.sigmaaldrich.com)处理的花的子房中,分析了生长素影响下的slarf9表达。在第一次施用后6h重复所述处理。对照花收集自开花期。
为分析转基因株系中slarf9的表达,从slarf9-oe株系(过表达株系)的第2代(t2)和rnaislarf9株系的第1代(t1)形成的子房和果实收集果皮组织。所有收集的组织于n2中冷冻并于-80℃储存至rna提取。
1.1.2实时定量pcr
使用
对于实时定量pcr,将5μl25-倍稀释的cdna用于25μlpcr反应中,所述pcr反应含有:400nm的每条引物以及12.5μliqtmsybrgreensupermix(bio-rad)。所述pcr反应在96-孔icycler(bio-rad)中进行,温度程序起始为95℃3min,然后40个循环(95℃15sec,60℃45sec。最后,确定所得产物的熔解温度以验证所扩增的片段的特异性。
使用计算机程序(beacondesigner5.01,premierbiosoftinternational,http://www.premierbiosoft.com)设计用于实时定量pcr引物,如下:
slactin正向引物:
5′-ggactctggtgatggtgttag-3′(seqidno:5)
slactin反向引物:
5′-ccgttcagcagtagtggtg-3′(seqidno:6)
slarf9正向引物:
5′-cgtaggcgtcaacaaatacttagagg-3′(seqidno:7)
slarf9反向引物:
5′-tccactgtgaagaaagatcatcaattcc-3′(seqidno:8)
该slarf9引物对可扩增slarf9mrna转录本中146个核苷酸的片段(seqidno:1的核苷酸834-979)。
1.1.3基因组slarf9序列的分离
为表征slarf9的结构特征,对从番茄幼叶组织分离的番茄基因组dna扩增几种pcr产物。引物来自slarf9的编码序列(genbank登录号bt013639)。将所述pcr产物完全测序并比对,以提供slarf9的外显子-内含子结构的信息。所述基因组序列如seqidno:3所示。栽培种heinz1706的基因组序列(seqidno:4)来自sgn数据库,scaffoldsl1.03sc03144(http:solgenomics.net)。
使用基因特异引物5'-ttcttcagccaggaaatgactattgataactcg-3'(反向)和巢式引物5'-ggagaattcatattcggctgagac-3'(反向)在snai(fermentas)染色体步移文库上进行染色体步移(genomewalkeruniversalkit,bdbiosciences,http://www.bdbiosciences.com),得到含有slarf9启动子的分离的3kb片段(在seqidno:3中显示,atg密码子的上游)。使用erase-a-base系统(promega,http://www.promega.com)产生含有该片段的逐步单向缺失的亚克隆。随后,将所述亚克隆测序并用clustalw(http://www.ebi.ac.uk/clustalw)进行比对。
1.1.4植物转化
为产生slarf9过表达株系(oe),将slarf9的编码序列(正向引物5'-caccatggcaactataaatgggtggtg-3'(seqidno:9),反向引物5'-ttaactgtctgcgcgagacaggg-3'-seqidno:10)克隆到pentrtm/d-topo进入载体(entryvector)(invitrogen)中。将该克隆与pgd625双元载体(drs.defolter,wageningenuniversity,荷兰)重组,在所述双元载体中花椰菜花叶病毒(camv)35s启动子由m.busscher(plantresearchinternational,荷兰)替换为子房和幼果特异的tprp-f1启动子(carmietal.,2003,见上文)。
为产生rnaislarf9株系,将slarf9中间区的cdna片段(氨基酸367-506,正向引物5'-aaaaagcaggctgtcccaccaaccgcagagaagaac-3'-seqidno:11;反向引物5'-agaaaagctgggtgctgtagtcgtgcctcagtagtgc-3'-seqidno:12)克隆到pdonrtm221进入载体(invitrogen),然后使所述载体与双元载体pk7gwiwg2(i)(karimietal,2002,trendsinplantsciences7,193-195)重组,正义和反义方向均受camv35s启动子和终止子的转录调节。
为产生pslarf9::gus株系,将slarf9启动子片段(2200bp,正向引物5'-caccttttcaaagaggtgtgacattttcaataac-3'-seqidno:13;反向引物5'-caaccttcaattccaaaaactaaagaacaccc-3'-seqidno:14)克隆到pentrtm/d-topo进入载体。使该进入克隆与目标载体pkgwfs7(karimietal.,2002,见上文)重组。
通过农杆菌介导的转化产生转基因番茄植株,如dejongetal.(2009,见上文)中所述。虽然在含卡那霉素的培养基上培养,仍然通过用基因组dna上卡那霉素抗性基因特异的引物(正向引物5'-gactgggcacaacagacaatcg-3';反向引物5'-gctcagaagaactcgtcaagaagg-3')的pcr检测了可能的逃逸。
随后,测试所述株系的四倍性,因为只使用二倍体株系进行后续分析。
1.1.5gus活性的组织化学分析
将pslarf9::gus株系的第一代成株(t1)和15d的幼苗(t2)的组织浸入含gus染色缓冲液中,所述gus染色缓冲液在50mm磷酸缓冲液(ph7.0)中含有0.1%tritonx-100、0.5mmfe2+cn、0.5mmfe3+cn、10mmedta、1mgml-1x-gluc、0.1mgml-1。37℃孵育后,用70%乙醇清洗所述组织,并在解剖镜(leicamzfliii,leicamicrosystems,http://leica-microsystems.com)下观察。为通过光学显微镜详细分析侧根和胚珠,将所述gus染色的组织包埋入technovit7100(heraeuskulzer,http://www.heraeus-kulzer.com)中。将所述包埋的组织切成5μm的切片。用0.5%番红将所述侧根的切片复染,随后用70%乙醇部分地脱色。将所述切片在leitzorthoplan显微镜(leicamicrosystems)下观察。使用leica数码相机(型号dfc420c,leicamicrosystems)拍摄图像。
1.1.6细胞面积和细胞层数目的定量方法
将7-8mm直径的果实的果皮组织在2%戊二醛、0.1m磷酸缓冲液(ph7.2)中于4℃固定过夜。随后,将所述组织在乙醇系列中脱水并包埋于spurr中。用甲苯胺蓝溶液(在1%硼砂中0.1%)将1μm的切片染色。
将破色期的成熟果实的果皮组织在faa(5%乙酸、3.7-4.1%甲醛溶液和50%乙醇)中固定,在乙醇系列中脱水,随后将其包埋于technovit中。用甲苯胺蓝溶液将5μm的切片染色。将所述切片在leitzorthoplan显微镜(leicamicrosystems)下观察。使用leica数码相机(型号dfc420c,leicamicrosystems)拍摄显微照片。将所述显微照片用于后续分析。
为分析7-8mm的果实,对0.16mm2的方形切片划界,并从内果皮(包括表皮层)定位大约0.1mm。为分析成熟的果实,对9mm2的切片划界,并从内果皮定位大约0.1mm。然后,计数这些方形内部的细胞总数目。将其大小的2/3或更大位于所述切片中的细胞包括在内。为估计所述果皮中的细胞层数目,划一条横过所述果皮切片的线,并记录沿该线的细胞数目,包括表皮、外果皮、中果皮和内果皮的细胞层。总计,分析了每个果实的一个区域,每条线5个果实。
1.2.结果
1.2.1番茄中slarf9的表达
slarf9的相对转录水平在传粉后2天内增加,但是用植物激素赤霉酸(ga3)处理后没有增加。slarf9在胎座、胚珠组织和子房壁中表达。见图1a中实时定量pcr。
对在花发育的多个阶段收集的子房mrna的分析显示,在花发育早期slarf9的转录量也非常丰富,但是在其后的阶段中下降,在开花期达到最低水平(图1b)。slarf9的转录本水平保持较低,除非发生成功传粉和受精。这些过程主要在胎座组织和子房壁中增加了slarf9的转录本水平(图1c)。
虽然用ga处理未传粉的成熟子房没有效应,但是生长素(iaa)的施用诱导了slarf9的表达(图1d),表明slarf9本身可对生长素应答。迄今为止,只发现atarf4、atarf19和稻(oryzasativa)arf23的表达是生长素应答的(ulmasovetal,1999,proceedingsofthenationalacademyofsciences,usa96,5844-5849;okushimaetal,2005,theplantcell17,444-463;overvoordeetal,2005,theplantcell17,3282-3300;wangetal,2007,gene394,13-24)。
在其他植物组织中,slarf9转录本的水平很低(图1e),表明slarf9功能可能主要是果实特异的。
为更详细地研究slarf9的表达,构建了slarf9启动子-gus融合物,将2200bp的slarf9编码区的5'侧翼序列连接到β-葡糖醛酸糖苷酶(gus)的编码序列之前。然后,通过农杆菌介导的基因转移将pslarf9::gus构建体导入番茄中。在所研究的14个独立株系的7个中,在组织化学gus染色后,在分析的几个组织中观察到了gus表达。
在5-6mm直径(对应于约8dap(传粉后的天数))的番茄果实中,在果皮、发育成凝胶状物质的胎座的外细胞层以及胚珠中观察到gus染色(数据未示出)。
通过胚珠的横切片的显微分析显示,gus染色位于胚囊的珠孔端。染色的面积和位置表明,gus并非由胚体(其处于4-16细胞的发育期)表达(al-hammadietal,2003,plantphysiology133,113-125),而是由胚柄或在胚柄基周围快速发育的壁向内生长物表达(briggs,1995,annalsofbotany76,429-439)。gus还在叶表面和茎表面的腺毛以及位于茎中叶基部的叶腋分生组织中表达。此外,还在初生根根尖、侧根原基以及长出的侧根中观察到了gus染色。所述染色位于根尖的分生组织区、中柱鞘以及薄壁组织的少数细胞层。
总之,所述结果表明,slarf9启动子活性不局限于果实中。有趣的是,该基因在其中转录的大多数组织都是在其中发生许多细胞分裂的组织。
1.2.2番茄中slarf9的过表达和沉默
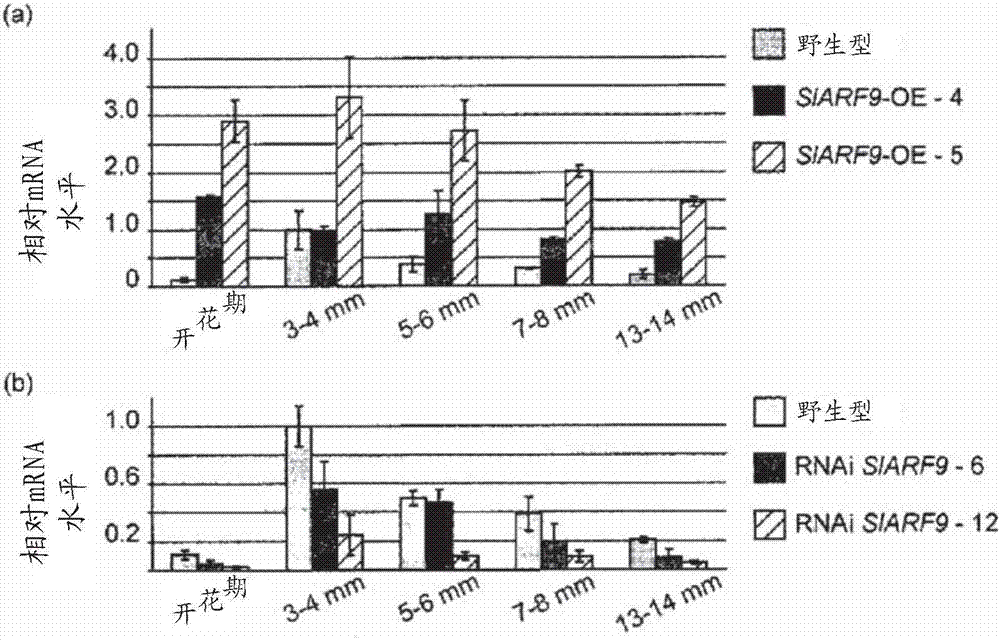
为研究slarf9在番茄果实形成和发育中的生理作用,产生了所述基因在其中过表达的转基因番茄株系。为产生这些slarf9过表达株系(slarf9-oe),将slarf9的编码序列连接到子房和幼果特异的tprp-f1启动子(carmietal,2003,见上文)。所产生的11个独立的转基因株系中,选择两个具有最高表达的slarf9-oe株系(分别为-4和-5)用于后续分析。
此外,使用基于slarf9中间区420bp的片段(氨基酸367-506),通过rna干扰(rnai)方法产生了在其中slarf9基因被沉默的转基因番茄株系。通过基因组dna的dna印迹分析测试了该片段的特异性,其产生了一个强的杂交信号(结果未示出)。
将所述片段克隆到rnai双元载体中,使其受camv35s启动子的转录调节,通过农杆菌介导的转化将其转移到番茄中。所产生的12个转基因株系中的2个中,slarf9的转录水平下降。将这两个rnaislarf9株系(-6和-12)用于后续分析。
果实发育早期的几个阶段中对slarf9的表达分析表明,在野生型中,传粉和受精后slarf9的相对mrna水平快速升高,在直径为3-4mm的果实(对应于6dap)中达到最高。在之后的阶段中,转录水平再次下降(图2)。
在slarf9-oe株系中,slarf9的转录水平在开花期已经很高,其不依赖于传粉,并且与野生型果实中的转录水平相比其较长时间地保持在高水平(图2a)。
在rnaislarf9株系中,slarf9的表达模式与野生型中类似,但是总的转录水平降低40-70%(图2b)。
虽然所述rnai构建体受组成型35s启动子调控,但是没有观察到生长表型,例如在根发育或茎分叉中。但是,slarf9-oe和rnaislarf9株系二者都显示出明显的果实发育的表型。研究了直径为7-8mm果实的组织学横切片。在细胞分离阶段期结束时,约10dap收集所述果实。定量了每单位面积的细胞数目和果皮中的细胞层数目。一般而言,果皮分化为三层:内果皮、中果皮和外果皮(gillaspyetal.,1993,见上文)。slarf9-oe果实的果皮中,与野生型果实的中果皮细胞数目相比,每mm2中果皮细胞的数目显著较少(p<0.05,student'sttest),而rnaislarf9株系的果皮中每mm2的细胞数目显著较多(p<0.05,student'sttest)(表1)。此外,所述转基因果实的果皮中的细胞层数目也可能受到了影响。在slarf9-oe果实中,该数目少于野生型果实中的数目,而在rnaislarf9果实中该数目增多。但是,由于果实的很大变异,对于所有转基因株系,这些差异在统计学上不显著(表1)。
表1
直径为7-8mm(10dap)的野生型果实和转基因果实的果皮中单位面积细胞数目或细胞层数目的定量。数据代表5个果实的平均值±标准误差。对于所有测量值,均测试了野生型和转基因株系之间差异的统计学显著性。示出了p-值(student'sttest)。
总之,这些结果表明,slarf9过表达时果皮中的细胞总数目减少,而在通过rnai方法使slarf9的转录物水平下降时果皮中的细胞总数目增多。
对破色期的成熟果实的果实重量和直径的分析表明,slarf9-oe株系的果实比野生型果实显著更小(重量p<0.05,直径p<0.05,student'sttest)。相反,rnaislarf9株系的果实比野生型果实显著更大(重量p<0.05,直径p<0.05,student'sttest)(表2)。
表2
对破色期收集的成熟野生型果实和转基因果实的果实重量和大小(直径)的分析。数据均代表5-20个果实的平均值±标准误差。对于所有测量值,野生型和转基因株系之间的差异均是统计学上显著的(p<0.05,student'sttest)。
此外,显微镜分析表明,rnaislarf9果实的果皮中的细胞层数目和单位面积的细胞数目多于野生型果实(表3)。rnaislarf9果实的更大果实大小(见图3)可能主要是由果皮中额外的垂周细胞分裂的结果。
所述显微照片还显示,rnaislarf9果实的果皮细胞不仅比野生型果实多,而且比野生型果实小(见图4)。下表3中,从细胞数目/mm2计算了细胞大小(细胞表面积)。
表3
对在破色期收集的成熟野生型果实和rnaislarf9果实的果皮中单位面积细胞数目或细胞层数目的定量。数据均代表5个果实的平均值±标准误差。示出了p-值(p;student'sttest)。
讨论
当子房在传粉和受精后转变为果实时,参与细胞周期和细胞膨大的几个基因被诱导(vriezenetal,2008,见上文;pascualetal.,2009,bmcplantbiology9,67;wangetal,2009,theplantcell21,1428-1452)。这些基因的诱导可能受到生长素和赤霉素的介导,这是由于以生长素处理未传粉的子房导致了具有更多果皮细胞数目的果实的形成,而ga-诱导的果实的果皮含有更少但更大的细胞(bünger-kiblerandbangerth,1982,见上文;serranietal.,2007,见上文)。与之相一致,发明人此前鉴定参与果实形成的基因的工作表明,传粉后生长素-和ga-相关基因二者的表达都上调(vriezenetal.,2008,见上文)。
本文中描述了被命名为slarf9的基因的功能分析。在拟南芥中,atarf9被表征为转录抑制因子(ulmasovetal.,1999见上文;tiwarietal.,2003,见上文),但是该转录因子的功能在很大程度上仍然是未知的,这是由于大多数t-dna插入突变株系未显示明显的表型(okushimaetal.,2005,见上文)。但是,缺少转录本3'端的atarf9突变株系在重力刺激后过度反应,表明atarf9可能参与重力信号转导。此外,还发现atarf9在胚的胚柄以及在arf9和arf13都被沉默的双敲除株系中表达,表明atarf9是控制胚柄发育所必需的(liuetal.,2008,见上文)。
尚未发现atarf9功能与拟南芥果实发育有关。已知参与该过程的仅有的arf是fruitwithoutfertilization(fwf)/arf8,因为fwf/arf8突变型株系形成了单性长角果(goetzetal,2006,见上文)。在番茄中,具有降低的slarf7转录水平的转基因株系也形成了单性果实,这表明slarf7充当了果实形成的负调节因子(dejongetal.,2009,见上文)。迄今为止,已经表征的番茄arf家族的唯一其他成员是developmentallyregulatedgene12(dr12),其为atarf4的同源物。整个果实发育过程中dr12的mrna水平都增加,在早红熟期的果实中达到最高水平。通过反义方法下调该基因影响了红熟期果实的硬度(jonesetal.,2002,见上文)。
相反,slarf9被发现主要在果实发育的早期表达。这些时期对应于番茄果实的生长主要依靠细胞分裂的阶段(mapellietal.,1978,见上文;bünger-kiblerandbangerth,1982,见上文;gillaspyetal,1993,见上文)。在生长素处理的未传粉的子房中slarf9的表达也被诱导,而在赤霉素施用后形成的单性果实中slarf9的转录水平未增加。
此外,gus分析的结果显示,slarf9也在发生许多细胞分裂的其他植物组织中表达。总之,这些结果表明slarf9可调节细胞分裂活性。
尽管有不能将slarf9看作果实特异的基因的事实,但是rnaislarf9株系只显示出果实表型,表明在其他植物组织中slarf9可能与arf蛋白家族的其他成员冗余地发挥功能。
slarf9转录本水平下降导致了显著更大果实的形成,可能主要是由于果皮中额外的垂周细胞分裂,而slarf9转录本水平增加导致了与野生型相比更小果实的形成。这些相反关系表明slarf9可调节细胞分裂,即slarf9蛋白是细胞分裂的负调节因子(抑制因子)。
slarf9下调可导致与野生型植物相比显著更大果实的结果使得该基因可用于改良植物果实大小,通过转基因或者通过提供含有突变型slarf9等位基因或slarf9启动子突变并籍此slarf9转录水平及蛋白水平下降或消失的(非转基因)植物。
实施例2–拟南芥中的slarf9启动子分析
2.1材料和方法
2.1.1植物材料和生长条件
col-0背景的拟南芥转基因植物在标准温室条件下,以温度22℃和16h光照/8h黑暗周期生长。将浸花(floraldip)转化产生的种子用100%乙醇处理1min并用2%盐酸溶液处理10min进行灭菌。用无菌蒸馏水冲洗3次后,将种子播种到1/2murashige和skoog(ms)培养基上(含有gamborgb5维生素、0.05%(w/v)mes、0.7%(w/v)琼脂(phytoagar)和30mgl-1卡那霉素,ph5.7)。在温室(16h光照/8h黑暗,22℃)下培育10d后,将抗性植株转移到土壤中。
如以前dejongetal.(2009,见上文)所述培养番茄植株(番茄栽培种moneymaker)。为分析生长素应答基因的表达,将去雄的花的子房用2μl的2%乙醇中1mm4-cl-iaa(sigma-aldrich,http://www.sigmaaldrich.com)处理。在第1次施用后6h,重复所述处理。在开花期收集对照花。将所收集的组织在n2中冷冻并在-80℃下储存至rna提取。
2.1.2植物转化
为产生转基因启动子::uida株系,将slarf9启动子片段(2200bp,正向引物5'-caccttttcaaagaggtgtgacattttcaataac-3'-seqidno:13;反向引物5'-caaccttcaattccaaaaactaaagaacaccc-3'-seqidno:14)和atarf9启动子片段(2466bp,正向引物5'-aaaaagcaggcttggtggtgggttttaaggcatc-3';反向引物5'-gaaaagctgggtcacacagtctctctatctctctcc-3')克隆到pentrtm/d-topo或pdonrtm221进入载体中(invitrogen,http://www.invitrogen.com)。之后,使所述进入克隆与目标载体pkgwfs7(karimietal,2002,trendsinplantsciences7,193-195)重组。用冻融转化将所述构建体转化到农杆菌菌株eha105中(chenetal.,1994,biotechniques16,664-670)。通过cloughandbent(1998,theplantjournal16,735-743)所述的浸花法进行拟南芥植株的转化。
2.1.3激素应答测定
为测试slarf9和atarf9启动子的生长素应答,将拟南芥pslarf9::gus株系的10d龄幼苗和成熟叶在0.05%乙醇或0.05%乙醇中50μμ吲哚乙酸(iaa)中孵育。在3h和9h后,将所述组织于n2中冷冻并在-80℃下储存至rna提取。
2.1.4实时定量pcr
依据实施例1.1.2中的方案,提取总rna,并反转录为cdna。同时,对于实时定量pcr,使用与实施例1.1.2相同的方法。,用于实时定量pcr的引物序列对于alarf9是seqidno:7和8,对于atarf9是如下:
正向引物:5'-agaagccatgagcaataagttctctgtagg-3'
反向引物:r5'-gggagcagtctttcacaccaataacc-3'
用于gus(uida)的是:
正向引物:5'-ctcctaccgtacctcgcattac-3'
反向引物:5'-ccgttgactgcctcttcgc-3'
2.1.5gus活性的组织化学分析
将拟南芥pslarf9::gus和patarf9::gus株系的成株(t1)和10d龄幼苗(t2)的组织浸入gus染色缓冲液(在50mm磷酸缓冲液(ph7.0)中含有0.1%tritonx-100、0.5mmfe2+cn、0.5mmfe3+cn、10mmedta、1mgml-1x-gluc、0.1mgml-1)中。在37℃下孵育后,用70%乙醇清洗所述组织。在解剖镜下观察(leicamzfliii,leicamicrosystems,http://leica-microsystems.com)染色的组织。使用leica数码相机(型号dfc420c,leicamicrosystems)拍摄图像。
2.1.6计算机启动子分析
使用plantcare(lescotetal,2002nucleicacidsresearch30,325-327http://bioinformatics.psb.ugent.be/webtools/plantcare/html)和place(higoetal,1999,nucleicacidsresearch27,297-300,http://www.dna.affrc.go.jp/place/)分析slarf9和atarf9(at4g2323980)(包括这些基因的5'非翻译区)的启动子序列。
2.2结果
2.2.1slarf9和atarf9的计算机启动子分析
番茄arf9(slarf9)转录本水平在传粉后48h内增加。但是,在以激素生长素处理的未传粉子房中,slarf9表达也被诱导(图1c)。迄今为止,只发现atarf4、atarf19和水稻arf23的基因表达被生长素诱导(ulmasovetal.,1999,见上文;okushimaetal,2005,见上文;overvoordeetal.,2005,见上文;wangetal.,2007,见上文)。在本研究中,发明人使用plantcare(lescotetal,2002,见上文)和place(higoetal,1999,见上文)软件分析了slarf9基因上游1500bp的5'区中生长素相关的顺式作用调节元件的存在情况,结果鉴定了两个简并性的(degenerated)生长素应答元件(auxre)。这些元件通常出现在生长素应答基因的启动子序列中,被arf转录因子结合(ulmasovetal,1999,见上文)。此外,所述启动子序列含有数个ntbbf1arrolb-元件。这些元件首先在rolb(存在于毛根农杆菌(agrobacteriumrhizogenes)的t-dna序列中的癌基因之一)启动子序列中被鉴定,参与植物中生长素诱导的rolb表达(baumannetal.,1999,theplantcell11,323-333)。auxre元件和ntbbf1arrolb元件也大量存在于sliaa2和sliaa14启动子序列中。这些基因是番茄aux/iaa基因家族的两个成员,该家族是调节生长素应答基因表达的转录抑制因子家族。但是,许多aux/iaa本身被生长素诱导(reed,2001,trendsinplantscience6,420-425)。与slarf9相似,sliaa2和sliaa14的表达在传粉后(vriezenetal,2008,见上文)以及在以生长素处理的未传粉子房中上调(图1)。对atarf9启动子序列的分析鉴定了几个生长素调节的顺式作用元件(结果未示出)。存在auxre元件、简并性auxre元件以及ntbbf1arrolb元件。此外,还大量存在元件asf1motifcamv。该元件存在于许多生长素应答基因中,并且最初在camv35s启动子中被检测到(liuandlam,1994,thejournalofbiologicalchemistry269,668-675)。类似的生长素相关元件也存在于生长素诱导的atiaa1和atiaa5启动子序列中(abeletal,1995,journalofmolecularbiology251,533-549)。
计算机启动子分析显示,arf9基因的5'端上游区中有与生长素诱导的aux/iaa基因的启动子区中出现的相同的生长素相关顺式作用元件,表明slarf9和atarf9二者的表达都受生长素调节。此外,所鉴定顺式元件的大多数在番茄和拟南芥启动子序列中都是类似的。
2.2.2生长素诱导的slarf9和atarf9表达
由于在slarf9和atarf9的启动子序列中检测到了相似的生长素相关的顺式作用元件,可以预测在拟南芥中保留了slarf9启动子的生长素可诱导性。因此,将slarf9编码区2200bp5'端侧翼序列连接到uida基因的β-葡糖醛酸糖苷酶(gus)编码序列之前。其后,通过农杆菌介导的基因转移,将该pslarf9::uida构建体导入拟南芥中。将所产生的转基因株系的成熟莲座叶假处理或在50μμ吲哚乙酸(iaa)中处理。在孵育3h和9h后,通过实时定量pcr分析所述叶样本中的uida-表达。此外,将这些组织样本用于研究atarf9表达的生长素诱导性。
虽然可以检测到uida的表达,但是其水平太低而很难可靠地定量。相反,atarf9的转录本水平可以定量。iaa处理3h和9h后,atarf9的转录本水平增加。但是,表达在假处理的样本中也上调(数据未示出)。因此,分析了atiaa1和atiaa5的表达。在iaa处理的样本中所述基因的转录本水平被强诱导,而在假处理样本中该转录本水平保持低水平(数据未示出)。这些结果表明,该实验设计是正确的。在pslarf9::uida株系的10d龄幼苗重复相同的实验,但是获得相似的结果。该结果表明,尽管其启动子序列中存在推定的生长素相关元件,但是atarf9的表达不被生长素诱导。
2.2.3拟南芥中slarf9和atarf9的表达模式
由于pslarf9::uida株系的表达太低而无法定量,在拟南芥中出现了slarf9启动子序列中的调节元件是否仍然具有功能的问题。因此,组织化学gus染色后对pslarf9::uida株系进行了分析。在10d龄幼苗中,托叶、正在发育的幼叶、正在发育的叶的表皮毛、早的侧根原基以及侧根的根尖被染成蓝色。此外,在花形态建成过程中,在几种组织中检测了gus活性。幼花芽中显示无gus活性,但是较大的花芽中观察到gus在萼片的顶端和柱头表达。传粉后,在发育中的种子中也检测到gus活性。但是,在传粉后约6天(6dap)收集的角果中观察到无gus活性。
为更详细研究atarf9的表达,使用连接到uida编码序列之前的atarf9编码区的2466bp5'端侧翼序列生成了转基因株系,并在组织化学gus染色后对其进行了分析。在10d龄的幼苗中,发育中的叶片的托叶和表皮毛被染色。此外,在根的中柱中也能检测到gus染色。花形态建成过程中,在幼花芽中观察到无gus表达。在较大的花芽中,只有雄蕊被染色。更加放大的视野显示,该染色位于正在发育的花粉粒、内壁细胞和花药的薄壁组织细胞中。在开花期之前不久收集的成熟花芽中,雄蕊中的gus表达减少,但是整个雌蕊中的gus表达增加。在传粉后,雌蕊的gus表达变得更加明显,保持在正在发育的角果中。此外,角果的离区被染色。
这些结果表明,在拟南芥中,slarf9的启动子序列中的调节元件仍具有功能。但是,slarf9启动子和atarf9启动子在不同组织中有活性。
实施例3–slarf9tilling突变体
3.1番茄tilling群
将商业加工番茄育种中使用的高度纯合的近交株系用于通过下述方案的诱变处理。约20,000粒种子(分为各2500粒的8批)在湿润的
3.2来自tilling群的用于pcr扩增的靶slarf9基因
对上述番茄tilling群的dna筛选slarf9靶基因的单核苷酸多态性。为此目的,设计下述pcr引物以扩增n端b3超家族dna结合结构域(dbd)的保守部分。该区域的突变可导致保守残基置换,其可导致突变的slarf9蛋白对其靶基因的启动子序列的亲和力下降。此外,在该区域引入可能的终止密码子将导致非常短的截短slarf9蛋白,其非常可能是无活性/无功能的。
如果需要,可设计其他引物,以筛选dbd的低保守性部分。
将所述引物对用于从tilling群的m2或m3dna扩增靶序列,并且按如下所述使用csce或hrm检测突变型和野生型靶序列之间的杂合双链。dna样本的id编号与具有杂合形式或纯合形式的野生型等位基因或突变型等位基因的植物的种子批相关联。
使种子萌发,并通过pcr确认个体植物中具体突变的存在情况,所述pcr使用突变位点侧翼的引物并以这些植物的基因组dna作为模板。对所述片段进行dna测序可鉴定对于所需突变纯合和杂合的突变体。在自交并和随后选择后选择或获得纯合突变体,并确定所述突变对所述植物的相应蛋白和表型的效应。
用引物对seqidno:19和22(突变体1719、突变体2484、突变体6725和突变体6932)或引物对seqidno:15和16(突变体3175)鉴定了下述突变体,并将种子以如下登录号保藏在ncimb。
因此,一个slarf9突变体可影响mrna前体剪接(突变体1719)、一个突变体位于外显子2(突变体3175)并且三个突变体位于外显子7(突变体2484、6725和6932),外显子7是slarf9的b3衍生的dna结合结构域的部分。
对靶序列中含有突变的植物,例如上述的突变型植物或由其产生的植物(例如通过自交或杂交)以及含有突变型slarf9等位基因的植物,基于表型筛选显著更大果实的形成。
也可以通过具有不同突变的植物杂交使两个突变型等位基因在一个植物中组合,以此确定对果实大小的效应。
3.3构象敏感的毛细管电泳(csce)
用0.15ng、4倍合并的基因组dna在10μl体积中进行多重pcr。将标记的引物加入pcr主混合物(mastermix)中使其浓度为未标记引物的1/5(1μμ)。在pcr后,将样本稀释10倍。将2μl所述稀释产物加入到38μlmq水中,然后进行csce。
将所述样本加样到50cm毛细管中(注射时间和电压:16秒,10千伏;运行电压:15千伏),所述毛细管来自装满具有下述组分的半变性聚合物的abi3130x1装置:5g构象分析聚合物(cap)(appliedbiosystems,434037,9%)、2.16g尿素、0.45g20×tte(nationaldiagnostics,ec-871),用mq水补充到9g。用1×稀释的tte和10%甘油制备运行缓冲液。装置温度设定为18℃。
使用bionumerics的heteroduplexanalysis(hda)软件分析原始数据。该程序能够区分杂合双链分子(突变体)和纯合双链分子(野生型)的峰值模式,因此提供了选择含有靶基因中有突变的个体株系的dna合并物的可能性。
3.4高分辨率熔解曲线分析(hrm)
在framstar96孔板(4titude,uk)的8×平孔中进行lcgreenpcr。根据说明书的建议,将2μ1(15ng)合并的dna与2μ1f-524phiretm5×反应缓冲液(finnzymes,finland)、0.1μlphiretm热启动dna聚合酶(finnzymes,finland)、1μllcgreentmplus+(biochem,usa)、0.25μl5mm引物混合,并用mq水补充到10μl。使用
实施例4-将突变型slarf9等位基因转移到番茄栽培种
将含有突变型slarf9等位基因(例如上述突变型等位基因的任一个)的tilling突变体与不同番茄株系杂交,以将所述突变型等位基因转移到这些株系,产生具有良好农艺特征和显著更大果实的番茄植株。
开发
序列表
<110>纽海姆有限公司
<120>果实大小增大的植物
<130>bcs10-2006pct
<141>2017-09-28
<150>ep10005603.5
<151>2010-05-28
<160>32
<170>siposequencelisting1.0
<210>1
<211>1977
<212>dna
<213>未知的(unknown)
<220>
<221>allele
<222>()..()
<223>番茄栽培种moneymaker的arf9编码序列
<400>1
atggcaactataaatgggtggtgttatgagtctcagccgaatatgaattctccaggtaaa60
aaagatgctctgtatcatgagctatggcagttgtgtgcaggtccagtagttgatgttccc120
agggaaggagaaagagtttattattttcctcaaggccacatggaacaattggtagcatca180
attaatcaagaaatggatcaaagagttccatcattcaatctcaaatcaaaggtcctttgt240
cgagttatcaatagtcatttcctggctgaagaagacaatgatgaggtctatgtccagatc300
actttgatgccagaggcaccacatgtacccgagccgactactccggatccattaattccg360
caggatgtaaagcctagattccattctttctgcaaggtcctgacagcctccgatacaagc420
actcatggtggattttctgttctaaggaaacatgctaatgaatgccttcctccattggac480
ttgaaccagcagactccgacccaggaattgattgcgaaagaccttcatgacgtggagtgg540
cgcttcaagcatatatttagaggccaacctcggagacatttacttaccacagggtggagt600
acctttgtttcttcaaagaaattagtggcaggggattcttttgtattcttgaggggtaat660
aatggacagctgcgagttggggtcaaaaggcttgttcgccagcagagctcaatgccgtca720
tcggtgatgtcaagccagagcatgcacctaggagtcttggctacagcatctcatgctgtt780
acaacccagacaatgtttgttgtttactacaaaccgagaaccactcagttcatcgtaggc840
gtcaacaaatacttagaggctcttaaacatgaatatgcagttggcatgcgattcaaaatg900
cagtttgaagccgaagggaatcctgatagaagatttatgggcactatagttggaattgat960
gatctttcttcacagtggaaaaattctgcgtggcgatccttgaaggtccgatgggacgag1020
cctgcagccattgcaaggcctgacagagtttctccttgggaaattaaaccttatgtgtgt1080
tcaattccaaatgtccttgtcccaccaaccgcagagaagaacaaaaggcatcggctacat1140
agtgaaatcaaaatatcagaacaaccttcatcctcaaatgcttcggcggtttggaatcct1200
tcccttcgatcgcctcagtttaacacctttggcatcaacagcagtactaattgcgcatta1260
gcgtctcttacagagagtggttggcagcttcctcatttaaatacttcaggtatgcttgtg1320
gatgagccagaagacggtaggagtgctccaacttggtgtggttttccatgcgttttggcc1380
ccgcagttcggtcaagggactaatcagccgattgttattcctactgatgggagaaaatgt1440
gataccaaaaaaacctgtagattatttggtattgacttgaaaagttcctcaattagcact1500
actgaggcacgactacagctacagccagccggcatttcttgtgtctttgcagagagagca1560
cctccaaacacggtgcctgctggtgattcagatcaaaagtctgagctttcagtagacttc1620
aaagatcaaatgcaaggccatttgcggttacccctaaaggaggttcaaagcaagcagagt1680
tgttccaccaggtctcgcacaaaggtgcaaatgcaaggcgtagctgtaggtcgtgcagtg1740
gatttaaccatattgaaaggatacgatgagcttacaaaggagcttgaggagatgtttgaa1800
atccaaggagagcttcagtcacgacagaaatgggggatcttgtttacagatgatgaaggg1860
gatacaatgcttatgggtgattatccgtggcaagacttttgcaatgtggtgaggaagatt1920
ttcatttgttcaagtcaggatatgaaaaaattgaccctgtctcgcgcagaccattaa1977
<210>2
<211>658
<212>prt
<213>未知的(unknown)
<220>
<221>peptide
<222>()..(564)
<223>番茄栽培种moneymaker的arf9蛋白
<220>
<221>domain
<222>(74)..(236)
<223>b3衍生的dna结合结构域
<220>
<221>domain
<222>(237)..(564)
<223>中间区(mr)
<220>
<221>domain
<222>(256)..(332)
<223>生长素应答区,中间区(mr)的部分
<220>
<221>domain
<222>(565)..(602)
<223>二聚化结构域iii
<220>
<221>domain
<222>(609)..(651)
<223>二聚化结构域iv
<400>2
metalathrileasnglytrpcystyrgluserglnproasnmetasn
151015
serproglylyslysaspalaleutyrhisgluleutrpglnleucys
202530
alaglyprovalvalaspvalproarggluglygluargvaltyrtyr
354045
pheproglnglyhismetgluglnleuvalalaserileasnglnglu
505560
metaspglnargvalproserpheasnleulysserlysvalleucys
65707580
argvalileasnserhispheleualaglugluaspasnaspgluval
859095
tyrvalglnilethrleumetproglualaprohisvalproglupro
100105110
thrthrproaspproleuileproglnaspvallysproargphehis
115120125
serphecyslysvalleuthralaseraspthrserthrhisglygly
130135140
pheservalleuarglyshisalaasnglucysleuproproleuasp
145150155160
leuasnglnglnthrprothrglngluleuilealalysaspleuhis
165170175
aspvalglutrpargphelyshisilepheargglyglnproargarg
180185190
hisleuleuthrthrglytrpserthrphevalserserlyslysleu
195200205
valalaglyaspserphevalpheleuargglyasnasnglyglnleu
210215220
argvalglyvallysargleuvalargglnglnsersermetproser
225230235240
servalmetserserglnsermethisleuglyvalleualathrala
245250255
serhisalavalthrthrglnthrmetphevalvaltyrtyrlyspro
260265270
argthrthrglnpheilevalglyvalasnlystyrleuglualaleu
275280285
lyshisglutyralavalglymetargphelysmetglnphegluala
290295300
gluglyasnproaspargargphemetglythrilevalglyileasp
305310315320
aspleuserserglntrplysasnseralatrpargserleulysval
325330335
argtrpaspgluproalaalailealaargproaspargvalserpro
340345350
trpgluilelysprotyrvalcysserileproasnvalleuvalpro
355360365
prothralaglulysasnlysarghisargleuhissergluilelys
370375380
ilesergluglnproserserserasnalaseralavaltrpasnpro
385390395400
serleuargserproglnpheasnthrpheglyileasnserserthr
405410415
asncysalaleualaserleuthrgluserglytrpglnleuprohis
420425430
leuasnthrserglymetleuvalaspgluprogluaspglyargser
435440445
alaprothrtrpcysglypheprocysvalleualaproglnphegly
450455460
glnglythrasnglnproilevalileprothraspglyarglyscys
465470475480
aspthrlyslysthrcysargleupheglyileaspleulysserser
485490495
serileserthrthrglualaargleuglnleuglnproalaglyile
500505510
sercysvalphealagluargalaproproasnthrvalproalagly
515520525
aspseraspglnlyssergluleuservalaspphelysaspglnmet
530535540
glnglyhisleuargleuproleulysgluvalglnserlysglnser
545550555560
cysserthrargserargthrlysvalglnmetglnglyvalalaval
565570575
glyargalavalaspleuthrileleulysglytyraspgluleuthr
580585590
lysgluleugluglumetphegluileglnglygluleuglnserarg
595600605
glnlystrpglyileleuphethraspaspgluglyaspthrmetleu
610615620
metglyasptyrprotrpglnaspphecysasnvalvalarglysile
625630635640
pheilecysserserglnaspmetlyslysleuthrleuserargala
645650655
asphis
<210>3
<211>7510
<212>dna
<213>未知的(unknown)
<220>
<221>allele
<222>()..()
<223>番茄栽培种moneymaker的arf9启动子和基因
<220>
<221>promoter
<222>(1)..(2004)
<223>arf9启动子
<220>
<221>misc_feature
<222>(612)..(617)
<223>auxre元件
<220>
<221>misc_feature
<222>(888)..(893)
<223>ntbbf1arrolb元件
<220>
<221>misc_feature
<222>(1224)..(1229)
<223>auxre元件
<220>
<221>misc_feature
<222>(1541)..(1546)
<223>ntbbf1arrolb元件
<220>
<221>misc_feature
<222>(1824)..(1829)
<223>ntbbf1arrolb元件
<220>
<221>misc_feature
<222>(1831)..(1836)
<223>ntbbf1arrolb元件
<220>
<221>exon
<222>(2005)..(2059)
<223>外显子1从翻译起始密码子atg开始
<220>
<221>exon
<222>(2156)..(2268)
<223>外显子2
<220>
<221>exon
<222>(2495)..(2590)
<223>外显子3
<220>
<221>exon
<222>(2949)..(3008)
<223>外显子4
<220>
<221>exon
<222>(3095)..(3247)
<223>外显子5
<220>
<221>exon
<222>(3347)..(3431)
<223>外显子6
<220>
<221>exon
<222>(3532)..(3622)
<223>外显子7
<220>
<221>exon
<222>(3765)..(3929)
<223>外显子8
<220>
<221>exon
<222>(4054)..(4167)
<223>外显子9
<220>
<221>exon
<222>(4433)..(4505)
<223>外显子10
<220>
<221>exon
<222>(4581)..(4734)
<223>外显子11
<220>
<221>exon
<222>(4843)..(5387)
<223>外显子12
<220>
<221>exon
<222>(5495)..(5682)
<223>外显子13
<220>
<221>exon
<222>(5795)..(5879)
<223>外显子14至翻译终止密码子
<400>3
acagagtcactcgtaatgtaaattttttcatatctaatatttaatttaatgttctcgatt60
tagagtaaagaattctcaacatgataattagtactttttaattttcgcacgcttttactt120
tttaatatgatagcggagtcacttatgacgtaaatttctccacatctaagactacaaatc180
taatgtcttcgattgagacttaagaattctcaccgttacatattaaattataatcttgac240
ttttatgcaatttttacggtagttattccaccattacacgacaggcggtgttaactttac300
ttagttaataaccagcccgctacttacccaaccaacgccccacacacttaaatccaaggt360
ccccaattaagactacagaaattttgccgttacataataattaattctcttgactttcac420
acttttttacttcttaataatataacagagtcacccgtaacgtaaatttttccatatcta480
agatttaaatctaatgtcctcgattaagagcaaaaaactcccaccattacatgataatta540
gtactttttgactttcgcacgctttcactttttaatacaataacgggatcactcctaagt600
cgtaacataaatgtcaccgcatctaacacttaaatctaatatcctctattaagactaaag660
aaatccaaccattacatgataattagtacttctcgagtttcgcacgcttttacttcttaa720
cataataactgagtcatttgtaacgtaaatttctccacatctaaaactcacatccaatat780
ctttgattaagacttaagaattctcaccgttacataataattaataatcttgacttttat840
gcaatttttacggtagttgttccaccattgcttgacaggtggtgttaactttacttagtt900
aataaccacccgttacttacccaaccaacccccccccccgcacacacacttaaatccaag960
gtccccgattaaagactaaagaaatctcgccgttacataataattaatactcttgacttt1020
cacacctttttacttcttaatacaataacactcgtaacgtaaatttttccatatctaaga1080
tttaaatctaatgtcctcgattaagagtaaaatctcttaccattacatgataattagtac1140
tttttgactttcgcacgcttttactttttaatacaataacggggtcacttccaattagtc1200
gtaacgtaaatatcaccgcatctaagacataaatccaatgtactcgattaagactaaaaa1260
aatccaaccattacatgataattaacacttctggactttcgcacgcttttacttcttaac1320
acaaataatggagtcacttgtaaaccataaatttctccacctttaacactcaaatctact1380
atctccgattaggactaaaaaacaaaaaactccacggttacatactaattaatatgtttg1440
acttttaaccacaattttttacagtagttattccgcccaatacacgaaagcggggtttac1500
tctacttttaaccaagcttataaaacccaaccaaccccccactttacttcacactacaca1560
gatgaaaaaaaggagagaaacacgttctctgcagggagcggtagaaagcaacaaatcaag1620
ccatatattaagctgtttttgggtaaactccgttacagagttttctctttctctctctca1680
ctttctctctttctctcacgtacaaaatagaaagaaaaaaaaaatgaaattattaagctg1740
ttgcttcttctgttgttgctcctgtttgttccagtagaactagtactactattgttagtt1800
aatttttttttatattcagcactactttaaactttatttttagttgttgggggtgtttct1860
ttaaggcttttttatgtgttgttgttatagcagggaagaaatgtggtgaattttagctag1920
ttttgtgatgtttttttgagctaattacttgattttgaattctgggtgttctttagtttt1980
tggaattgaaggttgattatactcatggcaactataaatgggtggtgttatgagtctcag2040
ccgaatatgaattctccaggtgaaagtttgattttttgaagtttaatttgagaattttgg2100
agctttttttgttttttttgtttaaaatgtggattttttattttttgagtttcaggtaaa2160
aaagatgctctgtatcatgagctatggcagttgtgtgcaggtccagtagttgatgttccc2220
agggaaggagaaagagtttattattttcctcaaggccacatggaacaagtaaaaaatttt2280
attttaaaaaaataattgaattctgtgtttttttttctttctgttttaggtgaatctggt2340
tgattcaaggctaagttcttgtttttgagttatggagttgtgaatttttattgaatttct2400
caatttgagtgtatatttttagttatgttttctggggattgtaaaatctgtatctttctc2460
tttttgtctcactttttttggttttgtggaaaagttggtagcatcaattaatcaagaaat2520
ggatcaaagagttccatcattcaatctcaaatcaaaggtcctttgtcgagttatcaatag2580
tcatttcctggtatgtttatatttcatggtgttatttgatcagtttattttacatacatt2640
gtattcatacttgttagtaattcatccaaaaaaagtgatgaatttattgttgggttcaaa2700
attcaaacatcggaaggtgagccttggcgtattcgtagctggcaaagttgtcggtttgtc2760
gccaagtgaacgaccatgaggtcaccgttcaatgcgtgtaagcaatctcttgcagaaatg2820
cagggtaaggttgtgtccaacagacccttatagtccagtctagcgggagctttagtgcac2880
ctgaatgcgtgtaaagttcatacaattttgtatttgaaatgtaaaatgtttttgatgatt2940
gcttgtaggctgaagaagacaatgatgaggtctatgtccagatcactttgatgccagagg3000
caccacatgtaagaagttaaaattgttgtagtattcacttgtttatatgcattttattga3060
tcaatttggttttaattattgatctgatttttaggtacccgagccgactactccggatcc3120
attaattccgcaggatgtaaagcctagattccattctttctgcaaggtcctgacagcctc3180
cgatacaagcactcatggtggattttctgttctaaggaaacatgctaatgaatgccttcc3240
tccattggtaatataagtgtgatcattttgtataggctagactacgtttccccccctgtt3300
ttgacaagttgttgaattagctgtcgtgtttgttatactttgttaggacttgaaccagca3360
gactccgacccaggaattgattgcgaaagaccttcatgacgtggagtggcgcttcaagca3420
tatatttagaggtaattgttttgtttgtagagtgtaacagaagatatcaatgaagcttta3480
tcatgtattgtaaaggtcaaaagtcatccgtctttgttttaattataacaggccaacctc3540
ggagacatttacttaccacagggtggagtacctttgtttcttcaaagaaattagtggcag3600
gggattcttttgtattcttgaggtatctctctctggtcctagcttctcgtatgagtatga3660
tatttgattgcattgtctatcactattgcatgtcatgtcatatatccagaacagttaatg3720
ttggtttgttaataatcatattctaatggatatgtacgctctaggggtaataatggacag3780
ctgcgagttggggtcaaaaggcttgttcgccagcagagctcaatgccgtcatcggtgatg3840
tcaagccagagcatgcacctaggagtcttggctacagcatctcatgctgttacaacccag3900
acaatgtttgttgtttactacaaaccgaggtttgtcacatacacctcctttcaattatcc3960
cgtcatccttacgcagtagagatcattttcatgccagagaacatactaaatctttcatat4020
gctcttccaattttctgctttgacaaccttcagaaccactcagttcatcgtaggcgtcaa4080
caaatacttagaggctcttaaacatgaatatgcagttggcatgcgattcaaaatgcagtt4140
tgaagccgaagggaatcctgatagaaggtagtgatatcaattactcgctagctccatttt4200
gatttatctgtcttactttcctttatagtttgtttaaaagaatgcatctttccctttttg4260
gctttaattcaacttaaatcccgtgccaagttaaaaccagcaaataaattgaaatggtgg4320
gagtattcagaagtcctccaattagatgtggaagtctttgatcaagatgtttatttgcca4380
actgcttagttctgacatgtatttgtttgggctctcccctattatttctcagatttatgg4440
gcactatagttggaattgatgatctttcttcacagtggaaaaattctgcgtggcgatcct4500
tgaaggttgatatctacatttatatttcagcaatattctttaagttcaattggaaatcac4560
caattctcccttcgttccaggtccgatgggacgagcctgcagccattgcaaggcctgaca4620
gagtttctccttgggaaattaaaccttatgtgtgttcaattccaaatgtccttgtcccac4680
caaccgcagagaagaacaaaaggcatcggctacatagtgaaatcaaaatatcaggtctga4740
atgaaaattctcaacagcatggtggtttttcatgtgttggttgctgatattttaatttcc4800
attttcattttcttaattatcttgcctttcttttcctgttagaacaaccttcatcctcaa4860
atgcttcggcggtttggaatccttcccttcgatcgcctcagtttaacacctttggcatca4920
acagcagtactaattgcgcattagcgtctcttacagagagtggttggcagcttcctcatt4980
taaatacttcaggtatgcttgtggatgagccagaagacggtaggagtgctccaacttggt5040
gtggttttccatgcgttttggccccgcagttcggtcaagggactaatcagccgattgtta5100
ttcctactgatgggagaaaatgtgataccaaaaaaacctgtagattatttggtattgact5160
tgaaaagttcctcaattagcactactgaggcacgactacagctacagccagccggcattt5220
cttgtgtctttgcagagagagcacctccaaacacggtgcctgctggtgattcagatcaaa5280
agtctgagctttcagtagacttcaaagatcaaatgcaaggccatttgcggttacccctaa5340
aggaggttcaaagcaagcagagttgttccaccaggtctcgcacaaaggtattaaatcaaa5400
agccaaaaaattgcattcttaccaactcattttcgccacaaaacatattattaaactctc5460
taacatcttttcacatgtggaatcccttgagaaggtgcaaatgcaaggcgtagctgtagg5520
tcgtgcagtggatttaaccatattgaaaggatacgatgagcttacaaaggagcttgagga5580
gatgtttgaaatccaaggagagcttcagtcacgacagaaatgggggatcttgtttacaga5640
tgatgaaggggatacaatgcttatgggtgattatccgtggcagtaagttctctctcactt5700
aaaaaaatttaagactgaatacatatacaaccactgcataagatcttttctacaacctgt5760
aatttaacacatcacttaaatcacttaattgcagagacttttgcaatgtggtgaggaaga5820
ttttcatttgttcaagtcaggatatgaaaaaattgaccctgtctcgcgcagaccattaag5880
cagctcctttcaaatgaaatgatggcggaagatggtgaaagttaaagactccgccttttg5940
aagtggtaccgttttgtgatctctttggcttcattctctatatatagtaggaaagaacga6000
caggttgttgacatagcaaggcgtagaagcttgtttttacgctctttttagcctagtgca6060
gctccgttttgagagcagttaagtaagtaactagttataataggataagactagtaaaaa6120
tatagtactagaagtagtaaaaagttaagggtgttctctaatggggagtctatatagtag6180
tagtttatgtagttttctattcgcgcgttaacctgtaaatactcatgtgccattgtagta6240
ttctggaaatactacaatgtatggtcttgttaacgtgctttatgccttatcttatatgaa6300
aactccatggtggagattgctctgtacttgctctttctgtatgacttttgtgatgcacaa6360
tagggatttcttgtctaaatttagactcttttggcacaatcatgcatcactttgtaactt6420
tcccctttactgaacaaaaaaatctttcaaaacgctccttcgcgtgtgtgtttgttttat6480
tctttgctaaagggtgatcctaacatttaaattagataccgttttctttactataagggt6540
gacttgaactcaaaacctgttttattagtatgttactctatttagaagtttaataattta6600
ttagatatgacatttctatttaccttgctatttcttttaacaatatttacctttagattt6660
atataagtttttgattcattaactgactttggttaagatgttgagtgcaaaataaataga6720
tggttcatcttttgagctagcgtagagtcattttcctttaaactttataaagcacttaca6780
atcacatatttgtgcattcttagtgcacgtttgaccatggataattttcactattttatg6840
gaactgtatattataatttttttggaattagaagaactcaaaaagactcttttcataatt6900
ttattccgaatcactcgtacaaaaatcaaaaaacaatttcaagttgtattcattttcaag6960
caaaagttcaatttgtcactatcacttgtaacacaaatattttccctttcgaatttcaca7020
attttaatattctaagttcctttttgagaacttttagtgaaaggaaaattgggctttgtg7080
aattgtttggtcttcaatatccctatcctagctaggaagagtgtccatatagtcatgggc7140
ctcatgagttgtcaaatgttacgtaaaggtatttatctttgcatgagtgtttggtcacgc7200
tcaaaacttctttattataagtctgagtctttgtgttatgtatccatgcttggcttgcat7260
gatgtttgtgtcattcctaatcttattgaatggactataaaggacttagtcaagtataac7320
tataaaatgacgatagcttatagacaacttcgtagttgttggagtatacaacacgctgac7380
atcgacttcttggagtttagtaatcataattttttttaaaaaaaacaaatctcctcaatc7440
aacttcttgattttagtagtaatccatcgattcaggtacattcatgacgtgatgtgttcg7500
attaatattt7510
<210>4
<211>7500
<212>dna
<213>未知的(unknown)
<220>
<221>allele
<222>()..()
<223>番茄栽培种heinz1706的arf9启动子和基因
<220>
<221>promoter
<222>(1)..(1995)
<220>
<221>misc_feature
<222>(612)..(617)
<223>auxre
<220>
<221>misc_feature
<222>(888)..(893)
<223>ntbbf1arrolb元件
<220>
<221>misc_feature
<222>(1229)..(1234)
<223>auxre
<220>
<221>misc_feature
<222>(1815)..(1820)
<223>ntbbf1arrolb元件
<220>
<221>misc_feature
<222>(1822)..(1827)
<223>ntbbf1arrolb元件
<220>
<221>exon
<222>(1996)..(2050)
<223>外显子1
<220>
<221>exon
<222>(2146)..(2258)
<223>外显子2
<220>
<221>exon
<222>(2485)..(2580)
<223>外显子3
<220>
<221>exon
<222>(2939)..(2998)
<223>外显子4
<220>
<221>exon
<222>(3085)..(3237)
<223>外显子5
<220>
<221>exon
<222>(3337)..(3421)
<223>外显子6
<220>
<221>exon
<222>(3522)..(3612)
<223>外显子7
<220>
<221>exon
<222>(3755)..(3919)
<223>外显子8
<220>
<221>exon
<222>(4044)..(4157)
<223>外显子9
<220>
<221>exon
<222>(4423)..(4495)
<223>外显子10
<220>
<221>exon
<222>(4571)..(4724)
<223>外显子11
<220>
<221>exon
<222>(4833)..(5377)
<223>外显子12
<220>
<221>exon
<222>(5485)..(5672)
<223>外显子13
<220>
<221>exon
<222>(5785)..(5869)
<223>外显子14至翻译终止密码子
<400>4
acagagtcactcgtaatgtaaattttttcatatctaatatttaatttaatgttctcgatt60
tagagtaaagaattctcaacatgataattagtactttttaattttcgcacgcttttactt120
tttaatatgatagcggagtcacttatgacgtaaatttctccacatctaagactacaaatc180
taatgtcttcgattaagacttaagaattctcaccgttacatattaaattataatcttgac240
ttttatgcaatttttacggtagttattccaccattacacgacaggcggtgttaactttac300
ttagttaataaccagcccgttacttacccaaccaacgccccacacacttaaatccaaggt360
ccccaattaagactacagaaattttgccgttacataataattaattctcttgactttcac420
acttttttacttcttaataatataacagagtcacccgtaacgtaaatttttccatatcta480
agatttaaatctaatgtcctcgattaagagcaaaaaactcccaccattacatgataatta540
gtactttttgactttcgcacgctttcactttttaatacaataacgggatcactcctaagt600
cgtaacataaatgtcaccgcatctaacacttaaatctaatatcctctattaagactaaag660
aaatccaaccattacatgataattagtacttctcgagtttcgcacgcttttacttcttaa720
cataataactgagtcatttgtaacgtaaatttctccacatctaaaactcacatccaatat780
ctttgattaagacttaagaattctcaccgttacataataattaataatcttgacttttat840
gcaatttttacggtagttgttccaccattgcttgacaggtggtgttaactttacttagtt900
aataaccacccgttacttacccaaccaaccccccccccccccccgcacacacacttaaat960
ccaaggtccccgattaaagactaaagaaatctcgccgttacataataattaatactcttg1020
actttcacacctttttacttcttaatacaataacactcgtaacgtaaatttttccatatc1080
taagatttaaatctaatgtcctcgattaagagtaaaatctcttaccattacatgataatt1140
agtactttttgactttcgcacgcttttactttttaatacaataacggggtcacttccaat1200
tagtcgtaacgtaaatatcaccgcatctaagacataaatccaatgtactcgattaagact1260
aaaaaaatccaaccattacatgataattaacacttctggactttcgcacgcttttacttc1320
ttaacacaataatggagtcacttgtaacataaatttctccacctttaacactcaaatcta1380
ctatctccgattaggactaaaacaaaactcacggttacatactaattaatatgtttgact1440
tttacacaatttttacagtagttattccgccaatacacgaaagcggtgttaactctactt1500
ctaaccaagcttataaaacccaaccaaccccccactctacttcacactacacagatgaaa1560
aaaggagagaaacacgttctctgcagggagcggtagaaagcaacaaatcaagccatatat1620
taagctgtttttgggtaaactccgttacagagttttctctttctctctctcactttctct1680
ctttctctcacatacaaaatagaaagaaaaaaaaaatgaaattattaagctgttgcttct1740
tctgttgttgctcctgtttgttccagtagaactagtactactattgttagttaatttttt1800
ttatattcagcactactttaaactttatttttagttgttgggggtgtttctttaaggctt1860
ttttatgtgttgttgttatagcagggaagaaatgtggtgaattttagctagttttgtgat1920
gtttttttgagctaattacttgattttgaattctgggtgttctttagtttttggaattga1980
aggttgattatactcatggcaactataaatgggtggtgttatgagtctcagccgaatatg2040
aattctccaggtgaaagtttgattttttgaagtttaatttgagaattttggagctttttt2100
tgtttttttgtttaaaatgtggattttttattttttgagtttcaggtaaaaaagatgctc2160
tgtatcatgagctatggcagttgtgtgcaggtccagtagttgatgttcccagggaaggag2220
aaagagtttattattttcctcaaggccacatggaacaagtaaaaaattttattttaaaaa2280
aataattgaattctgtgttgttgtttttttctgttttaggtgaatctggttgattcaagg2340
ctaagttcttgtttttgagttatggagttgtgaatttttattgaatttctcaatttgagt2400
gtatatttttagttatgttttctggggattgtaaaatctgtatctttctctttttgtctc2460
actttttttggttttgtggaaaagttggtagcatcaattaatcaagaaatggatcaaaga2520
gttccatcattcaatctcaaatcaaaggtcctttgtcgagttatcaatagtcatttcctg2580
gtatgtttatatttcattgtgttatttgatcagtttattttacatacattgtattcatac2640
ttgttagtaattcatccaaaaaaagtgatgaatttattgttgggttcaaaattcaaacat2700
cggaaggtgagccttggcgtattcgtagctggcaaagttgtcggtttgtcgccaagtgaa2760
cgaccatgaggtcaccgttcaatgcgtgtaagcaatctcttgcagaaatgcagggtaagg2820
ttgtgtccaacagacccttatagtccagtctagcgggagctttagtgcacctgaatgcgt2880
gtaaagttcatacaattttgtatttgaaatgtaaaatgtttttgatgattgcttgtaggc2940
tgaagaagacaatgatgaggtctatgtccagatcactttgatgccagaggcaccacatgt3000
aagaagttaaaattgttgtagtattcacttgtttatatgcattttattgatcaatttggt3060
tttaattattgatctgatttttaggtacccgagccgactactccggatccattaattccg3120
caggatgtaaagcctagattccattctttctgcaaggtcctgacagcctccgatacaagc3180
actcatggtggattttctgttctaaggaaacatgctaatgaatgccttcctccattggta3240
atataagtgtgatcattttgtataggctagactacgtttccccccctgttttgacaagtt3300
gttgaattagctgtcgtgtttgttatactttgttaggacttgaaccagcagactccgacc3360
caggaattgattgcgaaagaccttcatgacgtggagtggcgcttcaagcatatatttaga3420
ggtaattgttttgtttgtagagtgtaacagaagatatcaatgaagctttatcatgtattg3480
taaaggtcaaaagtcatccgtctttgttttaattataacaggccaacctcggagacattt3540
acttaccacagggtggagtacctttgtttcttcaaagaaattagtggcaggggattcttt3600
tgtattcttgaggtatctctctctggtcctagcttctcgtatgagtatgatatttgattg3660
cattgtctatcactattgcatgtcatgtcatatatccagaacagttaatgttggtttgtt3720
aataatcatattctaatggatatgtacgctctaggggtaataatggacagctgcgagttg3780
gggtcaaaaggcttgttcgccagcagagctcaatgccgtcatcggtgatgtcaagccaga3840
gcatgcacctaggagtcttggctacagcatctcatgctgttacaacccagacaatgtttg3900
ttgtttactacaaaccgaggtttgtcacatacacctcctttcaattatcccgtcatcctt3960
acgcagtagagatcattttcatgccagagaacatactaaatctttcatatgctcttccaa4020
ttttctgctttgacaaccttcagaaccactcagttcatcgtaggcgtcaacaaatactta4080
gaggctcttaaacatgaatatgcagttggcatgcgattcaaaatgcagtttgaagccgaa4140
gggaatcctgatagaaggtagtgatatcaattactcgctagctccattttgatttatctg4200
tcttactttcctttatagtttgtttaaaagaatgcatctttccctttttggctttaattc4260
aacttaaattccgtgccaagttaaaaccagcaaataaattgaaatggtgggagtattcag4320
aagtcctccaattagatgtggaagtctttgatcaagatgtttatttgccaactgcttagt4380
tctgacatgtatttgtttgggctctcccctattatttctcagatttatgggcactatagt4440
tggaattgatgatctttcttcacagtggaaaaattctgcgtggcgatccttgaaggttga4500
tatctacatttatatttcagcaatattctttaagttcaattggaaatcaccaattctccc4560
ttcgttccaggtccgatgggacgagcctgcagccattgcaaggcctgacagagtttctcc4620
ttgggaaattaaaccttatgtgtgttcaattccaaatgtccttgtcccaccaaccgcaga4680
gaagaacaaaaggcatcggctacatagtgaaatcaaaatatcaggtctgaatgaaaattc4740
tcaacagcatggtggtttttcatgtgttggttgctgatattttaatttccattttcattt4800
tcttaattatcttgcctttcttttcctgttagaacaaccttcatcctcaaatgcttcggc4860
ggtttggaatccttcccttcgatcgcctcagtttaacacctttggcatcaacagcagtac4920
taattgcgcattagcgtctcttacagagagtggttggcagcttcctcatttaaatacttc4980
aggtatgcttgtggatgagccagaagacggtaggagtgctccaacttggtgtggttttcc5040
atgcgttttggccccgcagttcggtcaagggactaatcagccgattgttattcctactga5100
tgggagaaaatgtgataccaaaaaaacctgtagattatttggtattgacttgaaaagttc5160
ctcaattagcactactgaggcacgactacagctacagccagccggcatttcttgtgtctt5220
tgcagagagagcacctccaaacacggtgcctgctggtgattcagatcaaaagtctgagct5280
ttcagtagacttcaaagatcaaatgcaaggccatttgcggttacccctaaaggaggttca5340
aagcaagcagagttgttccaccaggtctcgcacaaaggtattaaatcaaaagccaaaaaa5400
ttgcattcttaccaactcattttcgccacaaaacatattattaaactctctaacatcttt5460
tcacatgtggaatcccttgagaaggtgcaaatgcaaggcgtagctgtaggtcgtgcagtg5520
gatttaaccatattgaaaggatacgatgagcttacaaaggagcttgaggagatgtttgaa5580
atccaaggagagcttcagtcacgacagaaatgggggatcttgtttacagatgatgaaggg5640
gatacaatgcttatgggtgattatccgtggcagtaagttctctctcacttaaaaaaattt5700
aagactgaatacatatacaaccactgcataagatcttttctacaacctgtaatttaacac5760
atcacttaaatcacttaattgcagagacttttgcaatgtggtgaggaagattttcatttg5820
ttcaagtcaggatatgaaaaaattgaccctgtctcgcgcagacagttaagcagctccttt5880
caaatgaaatgatggcggaagatggtgaaagttaaagactccgccttttgaagtggtacc5940
gttttgtgatctctttggcttcattctctatatatagtaggaaagaacgacaggttgttg6000
acatagcaaggcgtagaagcttgtttttacgctctttttagcctagtgcagctccgtttt6060
gagagcagttaagtaagtaactagttataataggataagactagtaaaaatatagtacta6120
gaagtagtaaaaagttaagggtgttctctaatggggagtctatatagtagtagtttatgt6180
agttttctattcgcgcgttaacctgtaaatactcatgtgccattgtagtattctggaaat6240
actacaatgtatggtcttgttaacgtgctttatgccttatcttatatgaaaactccatgg6300
tggagattgctctgtacttgctctttctgtatgacttttgtgatgcacaatagggatttc6360
ttgtctaaatttagactcttttggcacaatcatgcatcactttgtaactttcccctttac6420
tgaacaaaaaaatctttcaaaacgctccttcgcgtgtgtgtttgttttattctttgctaa6480
agggtgatcctaacatttaaattagataccgttttctttactataagggtgacttgaact6540
caaaacctgttttattagtatgttactctatttagaagtttaataatttattagatatga6600
catttctatttaccttgctatttcttttaacaatatttacctttagatttatataagttt6660
ttgattcattaactgactttggttaagatgttgagtgcaaaataaatagatggttcatct6720
tttgagctagcgtagagtcattttcctttaaactttataaagcacttacaatcacatatt6780
tgtgcattcttagtgcacgtttgaccatggataattttcactattttatggaactgtata6840
ttataatttttttggaattagaagaactcaaaaagactcttttcataattttattccgaa6900
tcactcgtacaaaaatcaaaaaacaatttcaagttgtattcattttcaagcaaaagttca6960
atttgtcactatcacttgtaacacaaatattttccctttcgaatttcacaattttaatat7020
tctaagttcctttttgagaacttttagtgaaaggaaaattgggctttgtgaattgtttgg7080
tcttcaatatccctatcctagctaggaagagtgtccatatagtcatgggcctcatgagtt7140
gtcaaatgttacgtaaaggtatttatctttgcatgagtgtttggtcacgctcaaaacttc7200
tttattataagtctgagtctttgtgttatgtatccatgcttggcttgcatgatgtttgtg7260
tcattcctaatcttattgaatggactataaaggacttagtcaagtataactataaaatga7320
cgatagcttatagacaacttcgtagttgttggagtatacaacacgctgacatcgacttct7380
tggagtttagtaatcataattttttttaaaaaaaacaaatctcctcaatcaacttcttga7440
ttttagtagtaatccatcgattcaggtacattcatgacgtgatgtgttcgattaatattt7500
<210>5
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>5
ggactctggtgatggtgttag21
<210>6
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>6
ccgttcagcagtagtggtg19
<210>7
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>7
cgtaggcgtcaacaaatacttagagg26
<210>8
<211>28
<212>dna
<213>人工序列(artificialsequence)
<400>8
tccactgtgaagaaagatcatcaattcc28
<210>9
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>9
caccatggcaactataaatgggtggtg27
<210>10
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>10
ttaactgtctgcgcgagacaggg23
<210>11
<211>36
<212>dna
<213>人工序列(artificialsequence)
<400>11
aaaaagcaggctgtcccaccaaccgcagagaagaac36
<210>12
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>12
agaaaagctgggtgctgtagtcgtgcctcagtagtgc37
<210>13
<211>34
<212>dna
<213>人工序列(artificialsequence)
<400>13
caccttttcaaagaggtgtgacattttcaataac34
<210>14
<211>32
<212>dna
<213>人工序列(artificialsequence)
<400>14
caaccttcaattccaaaaactaaagaacaccc32
<210>15
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>15
aatttgagaattttggagcttttt24
<210>16
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>16
agaacttagccttgaatcaacca23
<210>17
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>17
tgagtttcaggtaaaaaagatgc23
<210>18
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>18
acagaaaaaaacaacaacacagaa24
<210>19
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>19
cccctgttttgacaagttgttg22
<210>20
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>20
cattgatatcttctgttacactctac26
<210>21
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>21
gtagagtgtaacagaagatatcaatg26
<210>22
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>22
ccagagagagatacctcaagaatac25
<210>23
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>23
ttcttcagccaggaaatgactattgataactcg33
<210>24
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>24
ggagaattcatattcggctgagac24
<210>25
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>25
gactgggcacaacagacaatcg22
<210>26
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>26
gctcagaagaactcgtcaagaagg24
<210>27
<211>34
<212>dna
<213>人工序列(artificialsequence)
<400>27
aaaaagcaggcttggtggtgggttttaaggcatc34
<210>28
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>28
agaaaagctgggtcacacagtctctctatctctctcc37
<210>29
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>29
agaagccatgagcaataagttctctgtagg30
<210>30
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>30
gggagcagtctttcacaccaataacc26
<210>31
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>31
ctcctaccgtacctcgcattac22
<210>32
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>32
ccgttgactgcctcttcgc19
- 还没有人留言评论。精彩留言会获得点赞!