一种陇东绒山羊羊绒性状相关的遗传标记及其应用的制作方法
本发明涉及陇东绒山羊羊绒性状相关的分子标记育种技术,具体涉及一种陇东绒山羊羊绒性状相关的遗传标记及其应用。
背景技术:
山羊绒指山羊被毛中直径在25um及以下的毛纤维,是由山羊皮肤中的次级毛囊形成的无髓毛纤维,在国际市场上称为“开士米”(cashmere)。由于其具有细而柔软、光泽良好、保暖性能强等诸多优点,山羊绒的价格昂贵,被称为“软黄金”。甘肃省是中国山羊绒的主产区之一,分布有河西绒山羊和陇东绒山羊2个甘肃省特有的绒山羊品种。在所有的羊绒性状中(例如产绒量、绒层高度、细度、净绒率等),产绒量和羊绒细度是最重要的2个性状,直接决定了羊绒的销售价格。羊绒性状主要受到遗传因素和非遗传因素(例如年龄、性别、营养等)的影响。研究证明,在遗传因素中,基因对羊绒性状起到了重要的调控作用。由于产绒量和羊绒细度是不能早期度量的性状,且羊绒细度的度量难度较大、成本较高。因此,利用常规育种方法(表型选择+后裔测定)对羊绒性状的改良难以在短期内取得良好的效果。分子标记辅助选择是改良此类性状的有效方法,可以明显缩短世代间隔,达到“早选、准选”目的,但前提是寻找到调控羊绒性状的关键基因。
角蛋白(keratins)和角蛋白关联蛋白(keratin-associatedproteins,kaps;其编码基因标识为krtaps)是羊绒纤维的主要蛋白成分,约占羊绒重量的90%。这2类蛋白决定了羊绒纤维的理化特性。kaps蛋白的典型特征是结构中含有高含量的半胱氨酸或甘氨酸/酪氨酸残基。根据氨基酸含量,kaps蛋白可被分为三类,分别是:①高硫kaps蛋白(high-sulfurkaps,hs-kaps,半胱氨酸的含量≤30%);②超高硫kaps蛋白(ultra-high-sulfurkaps,uhs-kaps,半胱氨酸的含量大于30%);③高甘氨酸/酪氨酸kaps蛋白(highglycine/tyrosinekaps,hgt-kaps,甘氨酸和酪氨酸的含量占35~60%)。根据氨基酸序列的同源性,kaps蛋白能被进一步分类为蛋白家族。
目前为止,krtaps基因的研究主要集中在人类和绵羊上。例如,在人类和绵羊中,分别有80个krtaps基因(属于25个基因家族)和29个krtaps基因(属于13个基因家族)被鉴别出来。但在山羊中只鉴别出10个krtaps基因,包括krtap1-1、krtap1-4、krtap6-2、krtap7-1、krtap8-1、krtap8-2、krtap9-2、krtap11-1、krtap13-1和krtap13-3,这表明大量的山羊krtaps基因还有待进一步鉴定。
虽然只在山羊中鉴别出少量的krtaps基因,但这些基因明显影响了山羊的羊绒性状。例如,krtap13-1和krtap13-3的核苷酸序列变异与绒山羊的产绒量、羊绒直径和绒层高度显著相关。在近缘物种—绵羊上,krtap1-2、krtap6-1、krtap6-3、krtap8-2和krtap22-1的核苷酸变异影响了包括产毛量在内的诸多重要的羊毛性状。这表明鉴别新的山羊krtaps基因以及研究基因多态性对羊绒性状的影响具有重要的意义,能为绒山羊的遗传改良提供理论依据和指导。
krtaps基因结构简单,长度较小。它只包含一个开放阅读框,没有内含子,长度约在600~1500bp之间。kap20属于高甘氨酸/酪氨酸kaps蛋白(hgt-kap)家族,它包括kap20-1和kap20-2两个成员。在人类中这两个蛋白及其编码基因已被鉴定,其序列特征、表达特点和遗传特征已被描述,但它们在包括山羊在内的家畜中尚未被鉴别出来。
技术实现要素:
本发明的目的就是针对家畜中krtap20-2基因的研究空白,提供了一种陇东绒山羊羊绒性状相关的遗传标记及其应用。
为了实现上述目的,本发明提供的技术方案为:一种陇东绒山羊羊绒性状相关的遗传标记,所述遗传标记为山羊krtap20-2基因的部分dna序列;所述遗传标记的核苷酸序列如序列表中seqidno:1-seqidno:3所示。
本发明的第二个目的是提供了一种用于扩增上述的陇东绒山羊羊绒性状相关的遗传标记的引物对,所述引物对的正向引物如序列表中seqidno:4所示,反向引物如序列表中seqidno:5所示。
本发明的第三个目的是提供了一种陇东绒山羊羊绒性状相关的遗传标记的筛选方法,包括以下步骤:从陇东绒山羊血液中提取基因组dna,用上述的引物对在绒山羊的基因组dna中进行pcr扩增,获得如序列表中seqidno:1-seqidno:3所示的核苷酸序列。
本发明的第四个目的是提供了一种陇东绒山羊羊绒性状相关的遗传标记在提高陇东绒山羊羊绒性状选育精准度中的应用。
本发明的第五个目的是提供了一种陇东绒山羊羊绒性状相关遗传标记的引物对在提高陇东绒山羊羊绒性状选育精准度中的应用。
本发明的有益效果为:本发明提供的一种陇东绒山羊羊绒性状相关的遗传标记及其应用。以陇东绒山羊为研究对象,首先鉴别出山羊的krtap20-2基因,进而利用pcr-sscp方法研究该基因的核苷酸序列变异,最后探讨基因多态性对陇东绒山羊羊绒性状的影响,以期为陇东绒山羊的遗传改良提供理论基础和指导。
krtap20-2的基因型和等位基因对陇东绒山羊的产绒量和绒层高度有极显著或显著影响(p<0.01)。含有等位基因a的个体的产绒量较缺失等位基因a的个体高38g(p<0.01),含有等位基因b的个体的产绒量和绒层高度分别较缺失等位基因b的个体低30g(p<0.01)和0.1cm(p<0.05)。因此,对陇东绒山羊的产绒量进行选种时,应选留含有等位基因a的个体,淘汰含有等位基因b的个体,以提高陇东绒山羊的产绒量。
附图说明
图1显示为山羊krtap20-2基因的pcr-sscp检测结果。
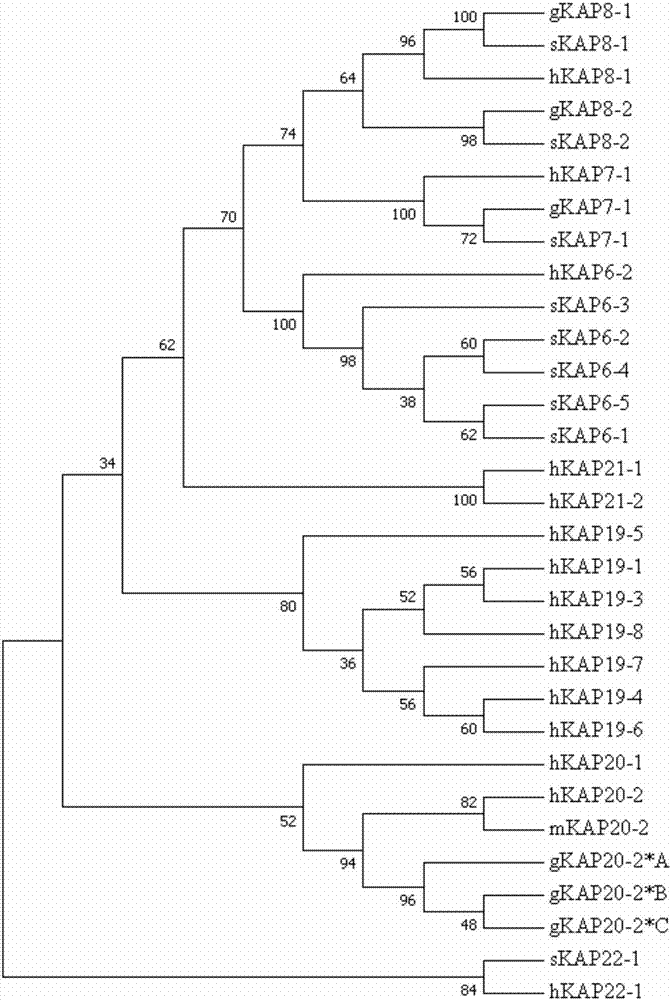
图2显示为基于山羊、绵羊和人类的hgt-kaps以及小鼠的kap20-2的氨基酸序列构建的最大简约树。
其中,山羊、绵羊、人类和小鼠的kaps序列分别用g、s、h和m标示。
具体实施方式
实施例1:
1材料与方法:
1.1实验材料:
在甘肃省环县钰生绒山羊繁育公司,以11只陇东绒山羊公羊作为父本,对陇东绒山羊母羊以人工授精方式配种。在所产后代中,随机选择373只陇东绒山羊作为试验对象,当后代羊只达到周岁时,现场测定产绒量,在背中线处测定绒层高度。同时在背中线处采集绒样用于实验室测定羊绒细度。另外,每只羊颈静脉采血8ml,加入酸性柠檬酸葡萄糖(citricaciddextrose,acd)抗凝,-20℃冻存,用于基因组dna的提取。
1.2基因组dna提取:
采用常规的苯酚-氯仿法提取血液基因组dna。
1.3引物设计和pcr扩增:
以人类krtap20-2基因的编码区序列作为模板(genbank登录号:nm_181616),应用genbank的blast程序在山羊基因组gcf_001704415.1(www.ncbi.nlm.nih.gov/assembly/gcf_001704415.1)中进行同源性搜索。搜索结果中同源性最高的序列被假定为山羊krtap20-2基因的序列。以此序列为模板,设计引物,扩增山羊krtap20-2基因的整个编码区序列。上游引物是5'-cagactatagagacagattcc-3',下游引物是5'-ccaattagttgagtttctctg-3'。引物由大连宝生物有限责任公司合成。
pcr扩增采用20μl体系,包括dna模板1μl(约50ng/μl)、10×pcrbuffer2.0μl、上下游引物各0.5μm、150μmdntps0.3μl、0.5utaqdnapolymerase0.2μl和ddh2o15.5μl。
pcr扩增条件:94℃预变性2分钟,94℃变性30秒,59℃退火30s,72℃延伸30秒,共35个循环,最后延伸5分钟。
pcr扩增结果用1.5%的琼脂糖凝胶电泳检测。
1.4山羊krtap20-2基因的sscp多态性检测:
取0.7μlpcr产物加入7μl上样缓冲液(98%甲酰胺、10mmedta、0.025%甲酰胺和0.025%二甲苯青)。95℃变性5分钟后立即置于冰水混合物中,上样于16×18cm、12%(acr:bis=37.5:1)的聚丙烯酰胺凝胶中,在0.5×tbe、16.5℃(恒温)、210v电压条件下电泳17小时。电泳结束后采用byun等的方法对聚丙烯酰胺凝胶进行银染显色。
1.5等位基因序列测定和序列分析:
凝胶银染显色后判定个体的基因型,测序方法因纯合子和杂合子而异。如果个体的sscp条带是纯合子,用pcr扩增产物直接测序。如果sscp条带是杂合子,按照gong等描述的方法切胶测序。序列测定由北京华大基因测序有限公司完成,每种基因型均选择3个不同的个体进行测序,以确保测序结果的准确性。
用dnaman(v5.2.10)软件进行核苷酸序列的比对和翻译,用genbank的在线blast软件(http://www.ncbi.nlm.nih.goc/)搜索同源性。基于氨基酸序列,用mega(v7.0)构建最大简约树(maximumparsimony),进化树的置信度以1000次自展分析进行重复检验。用于构建进化树的序列包括本研究获得的3条山羊的kap20-2序列,人类kap20-2序列(nm_181616)、小鼠kap20-2序列(nm_001163615)以及在山羊、绵羊和人类中鉴别出的其它hgt-kaps序列,包括绵羊kap6-1(nm_001193399)、绵羊kap6-2(kt725832)、人类kap6-2(nm_181604)、绵羊kap6-3(kt725837)、绵羊kap6-4(kt725840)、绵羊kap6-5(kt725845)、山羊kap7-1(ay510121)、绵羊kap7-1(x05638)、人类kap7-1(nm_181606)、山羊kap8-1(ay510122)、绵羊kap8-1(x05639)、人类kap8-1(nm_175857)、山羊kap8-2(ay510123)、绵羊kap8-2(kf220646)、人类kap19-1(nm_181607)、人类kap19-3(nm_181609)、人类kap19-4(nm_181610)、人类kap19-5(nm_181611)、人类kap19-6(nm_00130312)、人类kap19-7(nm_181614)、人类kap19-8(ab096964)、人类kap20-1(nm_181615)、人类kap21-1(nm_181619)、人类kap21-2(nm_181617)、绵羊kap22-1(kx377616)和人类kap22-1(nm_181620)。
1.6相关性分析:
采用spss(20.0)软件的一般线性混合效应模型(generallinermixed-effectmodels,glmms)分析频率大于5%的等位基因的状态(存在或缺失)和基因型对陇东绒山羊羊绒性状(产绒量、细度和绒层高度)的影响。相关性分析表明,性别和父本(配种公羊)对羊绒性状有极显著的影响(p<0.01),出生等级(单羔和双羔)对羊绒性状没有显著影响(p>0.05)。因此在模型中,等位基因状态(或基因型)和性别作为固定效应,父本作为随机效应。由于所有陇东绒山羊的羊绒性状均在周岁时被测定,且都在环县钰生绒山羊繁育公司这一相同的环境条件下饲养,因此在模型中没有考虑年龄和环境因素。
等位基因状态对羊绒性状影响的研究模型:y=μ+allele+gender+sire+e
基因型对羊绒性状影响的研究模型:y=μ+genotype+gender+sire+e
其中,y为羊绒性状表型值,μ为群体均值,allele为等位基因状态(用0和1分别表示缺失和存在),genotype为基因型,gender为性别,sire为父本(配种公羊),e为随机误差。
2结果:
2.1山羊krtap20-2基因的鉴定:
以人类krtap20-2基因的编码区序列(genbank登录号:nm_181616)为模板,应用genbank的blast程序在山羊基因组gcf_001704415.1中进行同源性搜索。结果显示,在山羊1号染色体上(nt3486283_3486471)发现了一个包含189bp开放阅读框的片段,这个片段与人类的krtap20-2基因有75%的序列同源性。
以以上片段作为模板,设计引物,经pcr扩增和测序,得到包含开放阅读框序列在内的273bp的扩增片段,与目的大小一致。经sscp分析,在373只陇东绒山羊个体中,鉴定出a、b和c3个等位基因(图1)。blast比对显示,3个等位基因之间的核苷酸序列有所差别,但它们与山羊基因组gcf_001704415.1的序列同源性都大于或等于98%。
以氨基酸序列为基础构建的最大简约进化树显示,在所有已知的山羊、绵羊和人类的hgt-kaps序列中,这3个等位基因的序列与人类和小鼠的kap20-2序列有最高的序列同源性(首先与这些物种的kap20-2序列聚为一支)(图2)。根据与山羊基因组的高度同源性以及系统进化树结果,这3个等位基因就是山羊krtap20-2基因的等位基因。这3个等位基因序列都被录入到genbank中,登录号为mf973462-mf973464。
2.2陇东绒山羊krtap20-2基因的snps:
测序结果表明,3个等位基因在189bp的编码区内存在4个snps,分别是c.27c/t、c.37c/t、c.125t/c和c.126g/a。其中3个snps是非同义突变,snpc.37c/t导致了p.his13tyr氨基酸的改变,c.125t/c和c.126g/a位于同一个密码子内,导致了p.met42thr氨基酸的改变,表1显示为山羊krtap20-2基因的snps。
表1
2.3krtap20-2在陇东绒山羊中的等位基因和基因型频率:
在373只陇东绒山羊中,共鉴别出a、b和c3种等位基因和5种基因型(aa、ab、bb、ac和bc)。a、b和c3种等位基因的频率分别为68.37%、28.95%和2.68%。5种基因型中,aa、ab和bb为优势基因型,它们的基因型总频率为94%,剩余2种基因型的频率不足5%。
2.4陇东绒山羊中3种羊绒性状间的表型相关性:
表型相关性结果显示,产绒量与绒层高度有中等的(0.3<|r|≤0.7)正相关(r=0.490),羊绒细度与产绒量和绒层高度有微弱的(|r|≤0.3)正相关,表2显示为陇东绒山羊3种羊绒性状间的表型相关性。
表2
对于每一个性状,第一行是相关系数(r),第二行是p值。
2.4krtap20-2基因型与陇东绒山羊羊绒性状的相关性分析:
相关性分析结果表明,性别和父本对陇东绒山羊的产绒量、细度和绒层高度有极显著影响(p<0.01),出生等级对羊毛性状没有显著影响(p>0.05)(表3)。因此,应用一般线性混合效应模型研究krtap20-2基因多态性对陇东绒山羊羊绒性状的影响时,性别作为固定效应,父本作为随机效应,在模型中没有考虑出生等级。
在陇东绒山羊上发现的5种基因型中,aa、ab和bb为优势基因型,它们的基因型频率均大于5%,其余2种基因型的频率均小于5%。因此,比较了3种优势基因型之间的羊绒性状的差异。基因型对陇东绒山羊的羊绒细度没有显著影响(p>0.05),但对产绒量和绒层高度有显著影响(p<0.05)。aa型个体的产绒量分别比ab和bb基因型个体高20g和47g(p<0.01),aa型个体的绒层高度比ab和bb基因型个体高0.1cm(p<0.05),表3显示为krtap20-2基因型与陇东绒山羊羊绒性状的相关性。
表3
2.5krtap20-2等位基因对陇东绒山羊羊绒性状的影响:
在陇东绒山羊上发现的4个等位基因中,等位基因a和b为优势等位基因,等位基因c的频率小于5%,因此分析了优势等位基因的存在或缺失对羊绒性状的影响(表4)。结果表明,2个等位基因对羊绒细度均没有显著影响(p>0.05),等位基因a对产绒量有极显著影响(p<0.01),等位基因b对产绒量和绒层高度有极显著或显著影响。含有等位基因a的个体的产绒量较缺失等位基因a的个体高38g(p<0.01),含有等位基因b的个体的产绒量和绒层高度分别较缺失等位基因b的个体低30g(p<0.01)和0.1cm(p<0.05)。表4显示为krtap20-2等位基因对陇东绒山羊羊绒性状的影响。
表4
4结论:
krtap20-2的基因型和等位基因对陇东绒山羊的产绒量和绒层高度有极显著或显著影响(p<0.01)。含有等位基因a的个体的产绒量较缺失等位基因a的个体高38g(p<0.01),含有等位基因b的个体的产绒量和绒层高度分别较缺失等位基因b的个体低30g(p<0.01)和0.1cm(p<0.05)。因此,对陇东绒山羊的产绒量进行选种时,应选留含有等位基因a的个体,淘汰含有等位基因b的个体,以提高陇东绒山羊的产绒量。
最后应说明的是:以上所述仅为本发明的优选实施例而已,并不用于限制本发明,尽管参照前述实施例对本发明进行了详细的说明,对于本领域的技术人员来说,其依然可以对前述各实施例所记载的技术方案进行修改,或者对其中部分技术特征进行等同替换。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
序列表
<110>甘肃农业大学
<120>一种陇东绒山羊羊绒性状相关的遗传标记及其应用
<160>5
<170>siposequencelisting1.0
<210>1
<211>273
<212>dna
<213>陇东绒山羊krtap20-2等位基因a的核苷酸序列(2ambystomalateralexambystomajeffersonianum)
<400>1
cagactatagagacagattccctgctcttcaaccaaatcctccatttctgaaaccatgtg60
ctactatggaaactactatggcggcctgggtcatggctacagtggcctgggttgtggtta120
tggttgtggctacagtagtctaggctgtggctatggtggcctaggctgtggctatggcat180
gtatgggtatggctgctgccgtccatcctgttgcagaagatactggtcatatgggttcca240
ctgagaatctttcagagaaactcaactaattgg273
<210>2
<211>273
<212>dna
<213>陇东绒山羊krtap20-2等位基因b的核苷酸序列(2ambystomalateralexambystomajeffersonianum)
<400>2
cagactatagagacagattccctgctcttcaaccaaatcctccatttctgaaaccatgtg60
ctactatggaaactactatggtggcctgggttatggctacagtggcctgggttgtggtta120
tggttgtggctacagtagtctaggctgtggctatggtggcctaggctgtggctatggcac180
atatgggtatggctgctgccgtccatcctgttgcagaagatactggtcatatgggttcca240
ctgagaatctttcagagaaactcaactaattgg273
<210>3
<211>273
<212>dna
<213>陇东绒山羊krtap20-2等位基因b的核苷酸序列(2ambystomalateralexambystomajeffersonianum)
<400>3
cagactatagagacagattccctgctcttcaaccaaatcctccatttctgaaaccatgtg60
ctactatggaaactactatggcggcctgggttatggctacagtggcctgggttgtggtta120
tggttgtggctacagtagtctaggctgtggctatggtggcctaggctgtggctatggcac180
atatgggtatggctgctgccgtccatcctgttgcagaagatactggtcatatgggttcca240
ctgagaatctttcagagaaactcaactaattgg273
<210>4
<211>21
<212>dna
<213>krtap20-2基因扩增的上游引物(2ambystomalateralexambystomajeffersonianum)
<400>4
cagactatagagacagattcc21
<210>5
<211>21
<212>dna
<213>krtap20-2基因扩增的下游引物(2ambystomalateralexambystomajeffersonianum)
<400>5
ccaattagttgagtttctctg21
- 还没有人留言评论。精彩留言会获得点赞!