一种促进细胞自噬和/或内吞的多肽或所述多肽的衍生物及其用途的制作方法
本发明属于蛋白质和肽化学领域,具体涉及一种促进细胞自噬和/或内吞的多肽或所述多肽的衍生物及其用途。
背景技术:
自噬是细胞溶质成分和细胞器被双膜结构吞噬和降解的过程,可被视为一种专门的多步膜运输过程。它开始于双膜自噬吞噬细胞质的部分,其后是这些囊泡与溶酶体的融合和自噬内含的降解。除了其重要的稳态作用外,这种降解途径还涉及各种人类疾病,包括代谢疾病、神经变性疾病、癌症和传染病。在细胞的营养或生长因子剥夺的条件下以及各种胁迫条件下诱导自噬。已表明在维持细胞内稳态和调节细胞存活方面发挥重要作用。自噬的抑制与许多类型的疾病病理学有关,包括但不限于癌症,帕金森病,阿尔茨海默病和亨廷顿病,胰腺炎,肥胖症和糖尿病,感染性疾病如细菌感染,艾滋病毒,肝炎和土热病(参见rubinszteindcetal.,natrevdrugdiscov.11(9),709-30(2012))。已知有几种分子参与自噬的起始和过程,包括但不限于1,2-棕榈酰磷酯酰肌醇-3-磷酸(pi3p)、含双重fyve结构域的蛋白(dfcp1)、amp依赖性激酶(ampk)以及iii类pi3激酶,例如vps34-vps15-beclin1-atg14l复合物。另外,一些调节分泌和内吞膜流量的rabgtp酶已被证明在自噬中发挥关键或辅助作用,包括但不限于rab1、rab5、rab7、rab9、rab24以及rab33。
内吞是指哺乳动物细胞吸收细胞外物质的过程。胞吞服用于许多重要的细胞功能,包括摄取细胞外营养物质,调节细胞表面受体表达,维持细胞极性和抗原呈递。内吞途径也被病毒,毒素和共生微生物所利用,以进入细胞。有几种类型的内吞途径,即受体介导的通过网格蛋白包被的细胞内吞,穴状病毒或膜芽,巨噬细胞增多症和吞噬作用。已经阐明了内吞作用在生理和病理过程中的作用,包括维持细胞极化,抗原呈递,葡萄糖转运,动脉粥样硬化,阿尔茨海默氏病和毒素和病毒的胞吞作用(mukherjee,s.etal.,physiolrev77(3),759-803,1997).
一般来说,p110β是pi3k的催化亚基,可以磷酸化磷脂酰肌醇和磷脂酰肌醇二磷酸产生3,4,5-三磷酸磷脂酰肌醇(pip3)。pip3通过向包括akt1和pdpk1的膜募集含有ph域(pleckstrin同源结构域)的蛋白质起着关键作用,激活参与细胞生长、存活、增殖、运动和形态的信号级联。此外,p110β在由g蛋白偶联受体(gpcr)配体如cxcl12,1-磷酸鞘氨醇和溶血磷脂酸刺激后参与akt1的激活。p110β在不同的信号通路中也需要稳定的血小板粘附和聚集,并且在由gpcrs、αiib/β-3整联蛋白(itga2b/itgb3)和itam(基于免疫受体酪氨酸的激活基序)引发的血小板激活信号中起作用。此外,p110β调节细胞传递收缩力所必需的itga2b/itgb3激活受体的粘附强度。需要由f2(凝血酶)和血栓素a2(txa2)诱导的血小板聚集并且在细胞存活中起作用。已经表明p110β在细胞迁移和自噬体形成和胞吞作用中起作用。p110β还调节磷脂酰肌醇3-磷酸的细胞内水平,并激活pik3c3激酶活性,并可作为支架,独立于其脂质激酶活性以积极调节自噬。此外,p110β可能在胰岛素信号传导中具有作为不需要脂质激酶活性的支架蛋白的作用。p110β可能在调节细胞增殖和网格蛋白介导的内吞作用中具有与激酶无关的功能。p110β作为缺乏pten的细胞系中致癌信号的介质。最后,p110β功能是erbb2和ras驱动肿瘤生长所必需的。
rab5是从rab5a基因表达产生的小gtp酶蛋白。它有两种形式:1)gtp结合,也称为其活性状态(本文中为“rab5-gtp,活性rab5或rab5”),以及2)gdp结合或其无活性形式。通常已知rabgtp酶调节分泌和内吞膜流量,并已被证明在自噬中发挥关键或辅助作用。在细胞自噬作用中起关键作用的小gtp酶rab5通过与vps34-beclin1复合体的相互作用参与自体吞噬体形成(ravikumar,b.etal.,jcellsci121,1649-1660,2008)。rab5的gtp结合形式是调节膜运输的活性形式(barbieri,maetal.,jbiolchem269,18720-18722,1994;stenmark,h.etal.,emboj13,1287-1296,1994)。此外,p110β缺乏和rab5失活在内吞和自噬途径中引起一些相似的改变,表明p110β和rab5可能在相同的信号通路中发挥其功能(dou,z.etal.,2010)。但两者关系未明。
技术实现要素:
本发明为克服现有技术当中p110β和rab5两者相互作用关系不明,且细胞自噬和/或内吞的机理不十分明确的缺陷,提供了一种促进细胞自噬和/或内吞多肽或所述多肽的衍生物及其用途。所述多肽及其衍生物均具有p110β的功能,本发明研究发现所述多肽及其衍生物与p110β竞争性地与rab5结合,最终促进细胞自噬和/或内吞,并能够作为小gtp酶rab5的激动剂。
本发明提供一种促进细胞自噬和/或内吞的多肽或所述多肽的衍生物,所述多肽为位于磷酸肌醇3-激酶的催化亚基p110β上的肽段,所述肽段以p110β肽链的第596位谷氨酰胺以及第597位异亮氨酸为关键残基,所述谷氨酰胺的n端连接有至少7个氨基酸残基,所述异亮氨酸的c端连接有至少5个氨基酸残基,所述多肽至少包括14个氨基酸残基;所述的衍生物为所述多肽与细胞穿膜肽连接所形成的嵌合肽。
较佳地,所述谷氨酰胺关键残基的n端连接有7~10个氨基酸残基,所述异亮氨酸的c端连接有5~8个氨基酸残基,和/或所述多肽的氨基酸残基不超过20个;更佳地,所述多肽的氨基酸序列具有序列表中seqidno.1所示的序列;进一步更佳地,所述多肽的氨基酸序列如序列表中seqidno.2所示。
如本领域常规,所述细胞穿膜肽可以连接在所述多肽的n端或者c段,较佳地,所述的细胞穿膜肽连接在所述多肽的n段;更佳地,所述穿膜肽为tat结构域,所述tat结构域的氨基酸序列如seqidno.3所示;进一步更佳地,所述多肽衍生物的氨基酸序列如序列表中seqidno.4或seqidno.5所示。
本发明还提供一种上述促进细胞自噬和/或内吞的多肽或所述多肽的衍生物在用于制备小gtp酶rab5的激动剂或治疗小gtp酶rab5介导的疾病的药物中的用途。所述疾病较佳地包括代谢疾病、神经变性疾病、癌症和传染病;所述代谢疾病优选为胰腺病、肥胖或者糖尿病;所述神经变性疾病优选为帕金森病、阿尔兹海默病或者亨廷顿病;和/或,所述传染病优选为艾滋病、肝炎或者土热病。较佳地,所述多肽或者多肽衍生物为所述药物的唯一活性成分。
本发明还提供一种非治疗用途地调节细胞自噬作用或者胞吞作用的方法,将上述促进细胞自噬和/或内吞的多肽或所述多肽的衍生物与细胞混合;较佳地,所述多肽或所述多肽衍生物与细胞混合后的终浓度为30μm。
本发明还提供一种小gtp酶rab5的激动剂,所述激动剂包括上述促进细胞自噬和/或内吞的多肽或所述多肽。
在符合本领域常识的基础上,上述各优选条件,可任意组合,即得本发明各较佳实例。
本发明所用试剂和原料均市售可得。
本发明的积极进步效果在于:
本发明所提供的多肽及其衍生物能够高效诱导或者激活细胞的自噬和/或内吞;所述的多肽及其衍生物能够作为小gtp酶rab5的激动剂。
附图说明
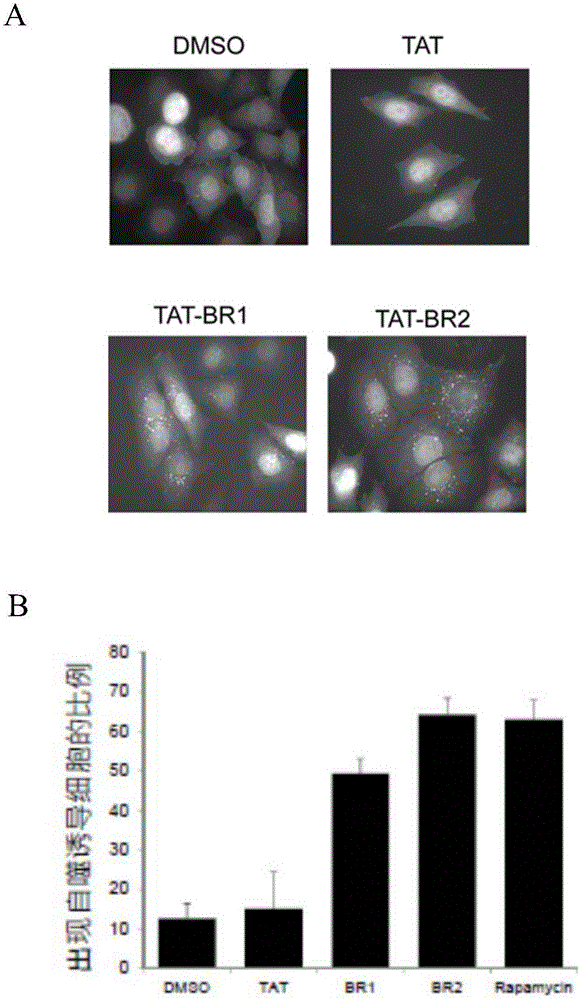
图1为p110β/rab5-结合肽tat-br1或tat-br2诱导细胞自噬,其中rapamycin为雷帕霉素的英文名称。
图2为tat-br1和tat-br2高效激活自噬功能。
图3为多肽br1为rab5的激动剂。
具体实施方式
下面通过实施例的方式进一步说明本发明,但并不因此将本发明限制在所述的实施例范围之中。下列实施例中未注明具体条件的实验方法,按照常规方法和条件,或按照商品说明书选择。
实施例1多肽的合成
多肽br1(seqidno.1)、tat-br1(seqidno.4)以及tat-br2(seqidno.5)由peptide2.0,inc.公司合成,所用合成方法为固态相合成法(solidphasepeptidesynthesis)。通常,固相肽合成包括含有包括但不限于-oh的反应性基团的合成聚合物的固体支持物。反应性基团与n-a保护的氨基酸的羧基容易反应,从而与聚合物共价结合。然后可以除去氨基保护基,并且可以将第二个n-a保护的氨基酸与所连接的氨基酸偶联。重复这些步骤,直到获得所需的序列。在合成结束时,应用不同的试剂来切割c-末端氨基酸和聚合物载体之间的键;肽然后进入溶液并且可以从溶液中获得。加入tat结构域以使肽在细胞培养和体内更易于渗透进入细胞(wagstaff,k.m.,jans,d.a.currentmedicinalchemistry13,1371-1387,2006)
实施例2多肽衍生物tat-br1和tat-br2诱导hela细胞自噬
hela细胞(人宫颈癌细胞系,购自atcc)稳定表达gfp-lc3,取4份等体积细胞(细胞浓度均为1×106个/ml)加入分别加入二甲基亚砜(dmso)、tat(30μm)、雷帕霉素(1μm)、tat-br1(30μm)以及tat-br2(30μm)。24小时后,在荧光显微镜下观察拍照。诱导自噬是由点状gfp-lc3的形成表示的(图1a)。
显示点状gfp-lc3的细胞被计算和量化为总细胞的百分比,结果表明tat-br1和tat-br2肽可以诱导自噬gfp-lc3(图1b)。其中雷帕霉素(rapamycin)被用作诱导自噬的阳性对照。
实施例3多肽衍生物tat-br1和tat-br2高效激活细胞自噬
hela人宫颈癌细胞(实验室培养过程中被支原体污染)分别用二甲基亚砜、tat结构域(30μm)、tat-br1肽(30μm)、tat-br2肽(30μm)进行24小时的培养(所用培养基为含有10%胎牛血清、2mml-谷氨酰胺,100单位/ml青霉素(penicillin)、100μg/ml链霉素的dmem,温度为37℃)后,用1毫克/毫升的dna染料4',6-二脒基-2-苯基吲哚(dapi)对细胞进行染色。结果表明,二甲基亚砜和tat结构域治疗细胞显示出dna的细胞质染色,表明支原体污染。tat-br1和tat-br2细胞支原体被显著清除。
实施例4多肽br1为rab5的激动剂
具有特异性的br1以及随机排列的非特异肽段(错配肽,seqidno.6)分别添加到p110β+/+或-/-的mefs细胞裂解液中。gst-r5bd沉淀显示(图3),多肽br1可以恢复p110β缺陷细胞裂解液中rab5的活性(rab5-gtp),说明多肽br1为rab5的激动剂。
sequencelisting
<110>武汉三成生物医药有限公司
<120>一种促进细胞自噬和/或内吞的多肽或所述多肽的衍生物及用途
<130>p1711100c
<160>6
<170>patentinversion3.5
<210>1
<211>14
<212>prt
<213>artificialsequence
<220>
<223>br1
<400>1
alaglnleuglnalaleuleuglniletrpprolysleupro
1510
<210>2
<211>20
<212>prt
<213>artificialsequence
<220>
<223>br2
<400>2
gluaspvalalaglnleuglnalaleuleuglniletrpprolysleu
151015
proproargglu
20
<210>3
<211>10
<212>prt
<213>artificialsequence
<220>
<223>tat结构域
<400>3
glyarglyslysargargglnargargarg
1510
<210>4
<211>24
<212>prt
<213>artificialsequence
<220>
<223>tat-br1
<400>4
glyarglyslysargargglnargargargalaglnleuglnalaleu
151015
leuglniletrpprolysleupro
20
<210>5
<211>30
<212>prt
<213>artificialsequence
<220>
<223>tat-br2
<400>5
glyarglyslysargargglnargargarggluaspvalalaglnleu
151015
glnalaleuleuglniletrpprolysleuproproargglu
202530
<210>6
<211>13
<212>prt
<213>artificialsequence
<220>
<223>错配肽
<400>6
leuglnleuvalalaaspalaglnleuileglnleuglu
1510
- 还没有人留言评论。精彩留言会获得点赞!