通过靶向糖蛋白A重复优势蛋白(GARP)治疗和诊断癌症以及单独或联合提供有效免疫疗法的方法与流程
本发明是在政府支持下由国家卫生研究院(nationalinstitutesofhealth)授予的基金号r01ai070603、p01ca186866、r01ca188419、和p30ca138313资助完成。政府对本发明拥有一定的权利。
序列表引用
创建于2017年3月23日的以“mescp0096wo_st25.txt”命名的7kb(于microsoft
背景技术:
1.发明领域
本发明一般涉及癌症生物学、免疫学和医学领域。更具体地,它涉及靶向garp(糖蛋白-a重复优势蛋白)的单克隆抗体用于治疗和检测癌症,以及使用免疫疗法治疗癌症的方法。具体地,提供了通过将t细胞疗法与抗血小板剂组合来治疗癌症的方法。
2.背景技术
tgf-β是在大多数组织中广泛表达的多效细胞因子。其信号传导中的异常与多种疾病和癌症特别相关(derynck等,2001;massague,2008)。除生长停滞外,tgf-β还诱导多种恶性细胞表型,包括侵袭,细胞粘附丧失,上皮-间质转化和转移(bhowmick等,2001;derynck等,2001;oft等,1998)。重要的是,tgf-β在塑造肿瘤微环境中的作用是其在癌发生中的功能的关键方面。例如,tgf-β1是一种有效的血管生成诱导剂(roberts等,1986),可直接通过诱导vegf表达(pertovaara等,1994)或通过募集其他细胞如单核细胞进而分泌促血管生成分子(sunderkotter等,1991)。tgf-β还可以通过篡改t细胞,nk细胞,b细胞或其他细胞的有效抗肿瘤功能(kehrl等,1986;kopp等,2009),通过其直接作用以及诱导foxp3+调节性t细胞的能力(li和flavell,2008),来促进癌细胞逃避免疫监视,从而控制肿瘤微环境。
生物化学上,tgf-β以至少4种不同的形式存在:1)易溶的活性tgf-β;2)与潜伏相关肽或lap相关的可溶性tgf-β(形成tgf-β-lap复合物,称为潜伏tgf-β或ltgf-β);3)与大tgf-β结合蛋白(ltbp)共价结合的ltgf-β,从而形成tgf-β-lap-ltbp复合物;和4)tgf-β的膜潜伏形式(mtgf-β)(li和flavell,2008;tran,2012)。仅已知不含lap的tgf-β具有生物学活性。因此,大量的tgf-β以潜伏形式被隔离在细胞外基质中,然后被蛋白酶如mmp2,mmp9和纤溶酶活化(lyons等,1990;sato和rifkin,1989;yu和stamenkovic,2000),其又由肿瘤细胞和肿瘤微环境中的其他细胞分泌。mltgf-β由两种造血细胞类型表达;血小板和调节性t细胞与跨膜蛋白糖蛋白a重复优势蛋白(garp)相关,也称为含富含亮氨酸重复32(lrrc32)(tran等,2009;wang等,2012)。除了作为mltgf-β停靠受体的作用外,garp对于调节tgfβ活化和生物利用度至关重要:garp增强protgf-β成熟并与整合素在mltgf-β活化中协同作用(wang等,2012)。本文描述了garp在癌症中的潜在作用。
通过过继转移大量肿瘤反应性淋巴细胞的被动免疫,称为过继性细胞疗法(act),目前是对转移性黑素瘤患者最有效的治疗方法,并且被广泛研究用于治疗其他人类癌症。act涉及给予大量对肿瘤抗原具有高亲合力的高选择性细胞。这些t细胞可以离体编程和活化以显示抗肿瘤效应功能。此外,t细胞输注之前可以用淋巴清除化学疗法或全身照射对患者进行“调节”,这使得能够减少免疫抑制细胞类型/因子,然后输注肿瘤特异性t细胞。尽管act似乎在许多方面都很有前途,但仍需要做大量的工作才能使治疗更加成功。
act令人鼓舞的临床成就面临着主要障碍,这些障碍限制了该方法的临床益处和更广泛的应用。虽然一些固有的困难可归因于用于分离,繁殖或产生效应淋巴细胞的特定方法,但其他困难,例如完全分化的t细胞的增殖和存活潜力的耗尽,似乎是更普遍的效应表型相关现象。其他困难起因于在肿瘤部位施加的外在抑制机制,其通过与肿瘤细胞,基质细胞和调节性t细胞(treg)的直接细胞-细胞接触或通过抑制性细胞因子如tgf-β介导。结果,给予的t细胞表现出降低的肿瘤内持久性和功能受损,并且通常不能执行可检测的杀肿瘤效果。因此,需要避免或破坏这些抑制机制并增强act治愈结果的方法。
技术实现要素:
本公开的实施方式提供了治疗癌症的方法。在第一个实施方式中,提供了分离的单克隆抗体,其中所述抗体特异性结合garp,并且其中所述抗体与4d3或5c5单克隆抗体竞争结合garp表位。在一些方面,抗体包含(a)与4d3(seqidno:1)或5c5(seqidno:9)的vhcdr1至少80%相同的第一vhcdr;(b)与4d3(seqidno:2)或5c5(seqidno:10)的vhcdr2至少80%相同的第二vhcdr;(c)与4d3(seqidno:3)或5c5(seqidno:11)的vhcdr3至少80%相同的第三vhcdr;(d)与4d3(seqidno:5)或5c5(seqidno:13)的vlcdr1至少80%相同的第一vlcdr;(e)与4d3(seqidno:6)或5c5(seqidno:14)的vlcdr2至少80%相同的第二vlcdr;和(f)与4d3(seqidno:7)或5c5(seqidno:15)的vlcdr3至少80%相同的第三vlcdr。
在某些方面,抗体包含与seqidno:1至少80%相同的第一vhcdr,与seqidno:2至少80%相同的第二vhcdr,与seqidno:3至少80%相同的第三vhcdr,与seqidno:5至少80%相同的第一vlcdr,与seqidno:6至少80%相同的第二vlcdr,和与seqidno:7至少80%相同的第三vlcdr。在一个具体方面,抗体包含与seqidno:1相同的第一vhcdr,与seqidno:2相同的第二vhcdr,与seqidno:3相同的第三vhcdr,与seqidno:5相同的第一vlcdr,与seqidno:6相同的第二vlcdr,和与seqidno:7相同的第三vlcdr。
在其他方面,抗体包含与seqidno:9至少80%相同的第一vhcdr,与seqidno:10至少80%相同的第二vhcdr,与seqidno:11至少80%相同的第三vhcdr,与seqidno:13至少80%相同的第一vlcdr,与seqidno:14至少80%相同的第二vlcdr,和与seqidno:15至少80%相同的第三vlcdr。在一个具体方面,抗体包含与seqidno:9相同的第一vhcdr,与seqidno:10相同的第二vhcdr,与seqidno:11相同的第三vhcdr,与seqidno:13相同的第一vlcdr,与seqidno:14相同的第二vlcdr,和与seqidno:15相同的第三vlcdr。
在一些方面,抗体包含(i)与4d3的vh结构域(seqidno:4)至少约80%相同的vh结构域和与4d3的vl结构域(seqidno:8)至少约80%相同的vl结构域;或(ii)与5c5的vh结构域(seqidno:12)至少约80%相同的vh结构域和与5c5的vl结构域(seqidno:16)至少约80%相同的vl结构域。在一个具体方面,抗体包含与4d3的vh结构域(seqidno:4)相同的vh结构域和与4d3的vl结构域(seqidno:8)相同的vl结构域。在一个具体方面,抗体包含与5c5的vh结构域(seqidno:12)相同的vh结构域和与5c5的vl结构域(seqidno:16)相同的vl结构域。在一个具体方面,抗体是4d3或5c5抗体。在其他方面,抗体是重组的。
在另外的方面,抗体是igg,igm,iga或其抗原结合片段。在某些方面,抗体是fab′,f(ab′)2,f(ab′)3,单价scfv,二价scfv或单结构域抗体。在特定方面,抗体可以是人、人源化抗体或去免疫抗体。在一些方面,抗体与成像剂,化学治疗剂,毒素或放射性核素偶联。在某些方面,抗体具有至少第二结合特异性,例如结合garp和第二靶标的双特异性抗体。
本发明的另一个实施方式提供了包含本文所述实施方式和方面的抗体的组合物,其在药学上可接受的运载体中。
在又一个实施方式中,本发明提供分离的多核苷酸分子,其包含编码本文所述实施方式和方面的抗体的核酸序列。
又一个实施方式提供了重组多肽,其包含抗体vh结构域,其包含4d3的vh结构域的cdr1-3(seqidno:1,2和3);5c5的vh结构域的cdr1-3(seqidno:9,10和11)。
在又一个实施方式中,提供了重组多肽,其包含抗体vl结构域,其包含4d3(seqidno:5,6和7),或5c5(seqidno:13,14和15)的vl结构域的cdr1-3。
又一个实施方式提供分离的多核苷酸分子,其包含编码本文所述实施方式和方面的多肽的核酸序列。
在又一个实施方式中,本发明提供了宿主细胞,其包含编码实施方式的抗体或本文所述的实施方式和方面的重组多肽的一种或多种多核苷酸分子。在一些方面,宿主细胞是哺乳动物细胞,酵母细胞,细菌细胞,纤毛虫细胞或昆虫细胞。
本发明的又一个实施方式提供了制备抗体的方法,包括(a)在细胞中表达编码实施方式的抗体的vl和vh链的一个或多个多核苷酸分子;和(b)从细胞中纯化抗体。
在进一步的实施方式中,本发明提供治疗患有癌症的对象的方法,包括给予有效量的本文所述的任何一个实施方式和方面的抗体。在某些方面,癌症是乳腺癌,肺癌,头颈癌,前列腺癌,食道癌,气管癌,皮肤癌脑癌,肝癌,膀胱癌,胃癌,胰腺癌,卵巢癌,子宫癌,宫颈癌,睾丸癌,结肠癌,直肠癌或皮肤癌。在特定方面,抗体是药学上可接受的组合物。在一些具体方面,抗体全身给予。在其他方面,抗体通过静脉内,皮内,肿瘤内,肌肉内,腹膜内,皮下或局部给药。在其他方面,该方法另外包括向对象给予至少第二抗癌疗法。在这些方面中的一些方面,所述第二抗癌疗法是外科疗法、化疗、放疗、冷冻疗法、激素疗法、免疫疗法、或细胞因子疗法。
本发明的又一个实施方式提供了用于检测对象中的癌症的方法,包括测试相对于来自对象的样品中的对照升高的garp的存在,其中所述测试包括使所述样品与本文所述的任何实施方式和方面的抗体接触。在一些方面,该方法进一步定义为体外方法。
在又一个实施方式中,提供了分离的抗体,其中所述抗体包含与4d3(seqidno:1)或5c5(seqidno:9)的vhcdr1至少80%相同的第一vhcdr,与4d3(seqidno:2)或5c5(seqidno:10)的vhcdr2至少80%相同的第二vhcdr,与4d3(seqidno:3)或5c5(seqidno:11)的vhcdr3至少80%相同的第三vhcdr,与4d3(seqidno:5)或5c5(seqidno:13)的vlcdr1至少80%相同的第一vlcdr,与4d3(seqidno:6)或5c5(seqidno:14)的vlcdr2至少80%相同的第二vlcdr,和与4d3(seqidno:7)或5c5(seqidno:15)的vlcdr3至少80%相同的第三vlcdr。
在另一个实施方式中,提供了治疗对象的癌症的方法,包括与抗血小板剂联合给予免疫疗法。在一些方面,用于治疗对象的方法包括与抗血小板剂联合给予t细胞,nk细胞,nkt细胞疗法,免疫检查点抑制剂(例如,抗体),细胞因子,溶瘤或免疫刺激疗法(例如,疫苗)。在一些方面,所述对象是人对象。
在一些方面,t细胞疗法包括给予肿瘤浸润淋巴细胞(til),cd8+t细胞和/或cd4+t细胞。在某些方面,t细胞疗法包括给予肿瘤特异性t细胞。在一些方面,肿瘤特异性t细胞是自体的。在某些方面,肿瘤特异性t细胞经工程改造以表达对garp或肿瘤抗原具有抗原特异性的t细胞受体(tcr)或嵌合抗原受体(car)受体。例如,所述肿瘤抗原选自tegfr、her2、cd19、cd20、cd22、间皮素、cea、cd23、cd24、cd30、cd33、cd38、cd44、egfr、egp-2、egp-4、epha2、erbb2、fbp、mage-a1、muc1、ny-eso-1、和mart-1。在某些方面,car包含选自cd28,cd27,4-ibb,ox40icos及其组合的共刺激分子内结构域。
在一些方面,该方法还包括在给予t细胞疗法之前对对象进行淋巴细胞清除。例如,淋巴细胞消除包括给予环磷酰胺,氟达拉滨和/或照射(例如,低剂量的全身照射)。
在特定方面,抗血小板剂是抗garp抗体或其片段。例如,抗garp抗体是单克隆抗体。在一些方面,抗garp抗体是人、人源化抗体或去免疫抗体。在某些方面,抗garp抗体包含4d3或5c5抗体的cdr序列。
在其他方面,抗血小板剂不是抗garp抗体。在一些方面,抗血小板剂选自环加氧酶抑制剂,二磷酸腺苷(adp)抑制剂,磷酸二酯酶抑制剂,蛋白酶激活受体-1(par-1)拮抗剂如凝血酶抑制剂,糖蛋白iib/iiia抑制剂,腺苷再摄取抑制剂和血栓素抑制剂。例如,adp抑制剂包括氯吡格雷,普拉格雷或噻氯匹定。
在一些方面,该方法还包括给予至少一种其它治疗剂。在某些方面,该至少一种其它治疗剂为化疗、免疫疗法、手术、放疗或生物疗法。例如,至少一种其他治疗剂是tgfβ抑制剂。在一些方面,tgfβ抑制剂是加仑他滨(galunisertib)(ly2157299),曲贝森(trabedersen),弗瑞索单抗(fresolimumab),ly2382770,瘤苗(lucanix)或pf-03446962。在某些方面,免疫疗法是免疫检查点抑制剂。例如,至少一种免疫检查点抑制剂是抗ctla-4抗体。在一些方面,免疫检查点抑制剂是人程序性细胞死亡1(pd-1)结合拮抗剂,pdl1结合拮抗剂或pdl2结合拮抗剂。在某些方面,pd-1结合拮抗剂是单克隆抗体或其抗原结合片段。在特定方面,pd-1结合拮抗剂是尼莫单抗(nivolumab),彭美罗珠单抗(pembrolizumab),ct-011,bms936559,mpdl328oa或amp-224。仍然在其他方面,剂是靶向pd-l1和tgfβ的双功能融合蛋白,例如m7824(msb0011359c)。
在某些方面,t细胞疗法在抗血小板剂之前,与抗血小板剂同时或在抗血小板剂之后给予。在一些方面,t细胞疗法和抗血小板剂同时给予。在某些方面,t细胞疗法和抗血小板剂通过静脉内,腹膜内,气管内,肿瘤内,肌肉内,内窥镜,病灶内,经皮,皮下,区域性或通过直接注射或灌注给予。
在一些方面中,癌症是膀胱癌,乳腺癌,透明细胞肾癌,头/部鳞状细胞癌,肺部鳞状细胞癌,黑色素瘤,非小细胞肺癌(nsclc),卵巢癌,胰腺癌,前列腺癌,肾细胞癌,小-细胞肺癌(sclc),三阴性乳腺癌,急性淋巴细胞白血病(all),急性髓性白血病(aml),慢性淋巴细胞白血病(cll),慢性骨髓白血病(cml),弥漫性大b细胞淋巴瘤(dlbcl),滤泡性淋巴瘤,霍奇金淋巴瘤(hl),套细胞淋巴瘤(mcl),多发性骨髓瘤(mm),骨髓细胞白血病-1蛋白(mcl-1),骨髓增生异常综合征(mds),非霍奇金淋巴瘤(nhl),或小淋巴细胞淋巴瘤(sll)。在特定方面,所述癌症是乳腺癌。在其他方面,所述癌症是黑色素瘤。
在某些方面,相对于给予治疗之前,对象中调节性t细胞(treg)的数量减少。在特定方面,treg是foxp3+treg。
本文所述“基本上不含”,就特定成分而言,指所述的特定成分均非有意配制进组合物并且/或者仅作为污染物或以痕量存在。因此,组合物的任何非有意污染所导致的特定成分的总量远小于0.05%。最优选的是其中所述特定成分的量无法使用标准分析方法检测到的组合物。
说明书和权利要求中所用“一个”或“一种”可指一个/种或多个/种。如本文说明书和权利要求中所用,当与词语“包括”联用时,“一种”或“一个”可指一种/个或多于一种/个。如本文所用,在说明书和权利要求书中,“另一个”或“另外的”可以表示至少第二个或更多个。
如本说明书和权利要求书中所用,术语“约”用于表示包括装置,用于测定数值的方法,或存在于研究对象中的差异造成的包括固有误差的固有变动的值。
通过以下详细说明,本发明的其他目的、特征和优点将显而易见。然而,应当理解的是以下详述和具体实施例以及本发明某些实施方式仅仅是用于举例说明,本领域普通技术人员通过阅读所述详述,可以显而易见地得知本发明精神和范围之内的各种变化和改良。
附图简述
以下附图形成本说明书的部分并且包括以进一步证明本文提供的某些方面。可参照这些附图中的一幅或者多幅并结合本文所示具体实施方式的详细描述来更好地理解本发明。
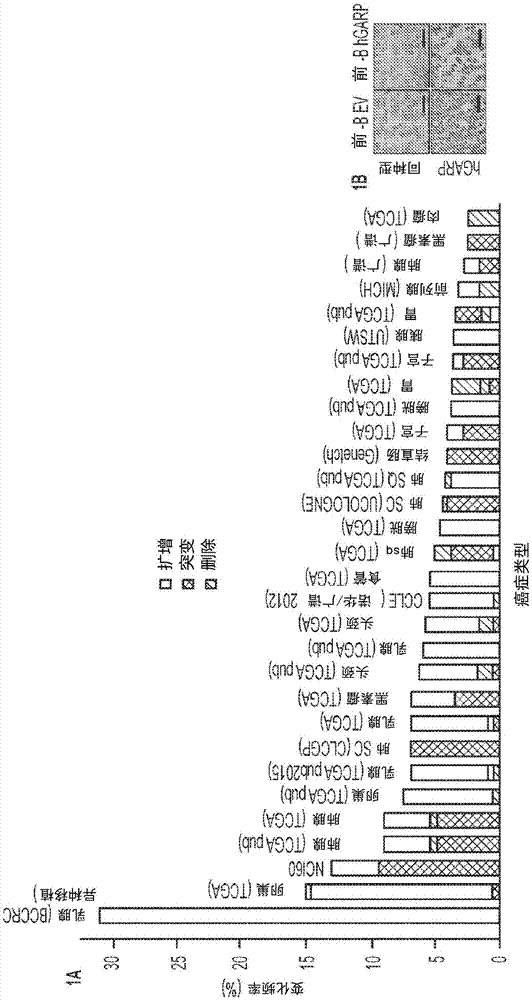
图1a-1f。癌症中的garp上调与不良预后相关。(1a)garp的跨癌症改变研究综述。数据来自www.cbioportal.org以响应于2015年11月16日对garp基因lrrc32的查询。(1b)hgarp抗体在表达hgarp的前bev和前b白血病细胞中的特异性分析。(1c)患者匹配的未受累和原发性乳腺癌。显示的是代表性图像和ihcgarp分数。(1d)正常组织和癌症的garpihc(暗色区域)的代表性图像。比例尺:20μm。(1e)garp阳性细胞的表达强度。(1f)garp表达与结肠癌和肺癌(左图和中图)的总体存活率以及前列腺癌的gleason评分(右图)之间的相关性。指示样品数量(n)。kaplanmeier曲线显示在f组中,针对肺癌和结肠癌,p值通过对数秩检验计算。两个样品t检验用于比较图1c,1e中的组差异和1f中的前列腺癌。hr代表风险比。*p<0.05。**p<0.01。***p<0.001。****p<0.0001。
图2a-2f。从癌细胞中脱落膜结合的garp及其作为潜在癌症生物标志物的意义。(2a)er后隔室中的garp裂解仅在grp94存在下发生。n-末端flag标记的garp在wt或grp94前bko细胞中稳定表达。用endoh或pngasef处理全细胞裂解物,然后用flag抗体进行免疫印迹。(2b)基于免疫反应性和质谱分析,下部片段蛋白质是garp。显示了通过质谱法鉴定的来自garp的肽序列(seqidno:17)。(2c)前列腺癌患者和对照正常对象的血清中可溶性garp。(2d)garp阳性与psa1水平(左图),garp阳性和前列腺癌转移状态(右图)之间的相关性分析。(2e)通过夹心elisa定量前列腺癌患者和正常对象血清中的garp-tgf-β1复合物。(2f)来自纯化的重组可溶性garp-fc的活性tgfβelisa水平。通过卡方检验计算图2d中的分布差异。使用两个样品t检验来比较图2e中的组差异。*p<0.05。***p<0.001。
图3a-3j。在正常乳腺上皮细胞上强化的garp表达增强tgf-β信号传导并驱动上皮-间充质细胞转化(emt)和侵袭。(3a)转染nmumg细胞以稳定表达膜结合的garp,然后对e-钙粘蛋白,波形蛋白和磷-smad-2/3进行western印迹。(3b)用重组人tgf-β1,可溶性garp和同种型抗体对照处理nmumg细胞或在无血清培养基中处理24小时,然后进行形态学分析。(3c)在无血清培养基中用可溶性garp-fc(sgarp)以指定时间处理nmumg细胞。通过western印迹分析来检测波形蛋白上调。(3d)用增加剂量的可溶性garp处理nmumg细胞,然后进行针对波形蛋白的免疫印迹。(3e)garp,tgfβ和β-肌动蛋白对照的免疫印迹。(3f)在nmumgev,garp和garp-fc细胞的条件培养基中可溶性garp的elisa定量。(3g)体外划痕试验,表明24小时间隙闭合的差异。(3h)三次独立划痕试验的总结。(3i)在第3周和第6周注射garp,garp-fc和对照nmumg细胞后,对小鼠中荧光素增强的生物发光的体内成像。(3j)通过h&e对nmumg-garp肿瘤进行组织学分析,并且通过ihc对e-钙粘蛋白和波形蛋白表达进行组织学分析。比例尺:20μm。使用两个样品t检验来比较图3h中的组差异。*p<0.05。**p<0.01。进行了两次独立实验,结果相似。
图4a-4g。garp沉默阻断乳腺癌的生长和转移。(4a)nmumg*细胞中garpmrna的shrna敲减。用乱序shrna(scr)处理的细胞用作对照。(4b)对来自garpkd和scrnmumg*细胞的细胞表面garp表达的流式细胞术分析。(4c)garpkd和scrnmumg细胞中总garp和tgf-β水平的免疫印迹。(4d)mtt测定以比较nmumg*-scr与nmumg*-garp-kd细胞的生长动力学。(4e-4g)将nmumg*scr和nmumg*-garpkd细胞注射到nod-rag1-/-小鼠中,然后监测肿瘤生长动力学(4g)和肿瘤转移(4f和4g)。通过双因素anova计算图4d和4e中的肿瘤生长差异。使用两个样品t检验来比较图4f和4g中的组差异。**p<0.01。
图5a-5j。鼠乳腺癌细胞中的garp上调促进tgf-β活化,肿瘤生长,转移和免疫耐受。(5a)稳定工程化以表达garp,garp-fc或对照ev的4t1细胞中garp,tgf-β和β-肌动蛋白对照的免疫印迹。(5b)在来自4t1ev,garp和garp-fc细胞的72小时条件培养基中通过elisa定量活性tgf-β1。(5c)在来自4t1-ev,4t1-garp和4t1-garp-fc细胞的50%3天条件培养基存在下,用抗-cd3和抗cd-28mab刺激原初cd4+t细胞。在第3天通过流式细胞术分析foxp3表达。(5d)将指定肿瘤注射到雌性balb/c小鼠的第4乳腺脂肪垫中。每3天测量肿瘤体积。(5e)在(5d)的终点以克为单位的肿瘤重量。(5f)分离肺并石蜡包埋。计算肺中肿瘤结节的数量。(5g)分离3周肿瘤并包埋在oct中。对新鲜冷冻切片进行p-smad-2/3mab染色。比例尺:100μm。(5h)p-smad-2/3染色强度的汇总统计,由研究病理学家独立定义。(5i-5j)分离肿瘤浸润淋巴细胞,通过流式细胞术计数cd4+cd25+foxp3+treg的数目。(5i)代表性的流程图。(5j)肿瘤微环境中treg百分比的总结。通过双因素anova计算图5d中的肿瘤生长差异。使用两个样品t检验来比其他组中的组差异。*p<0.05。**p<0.01。***p<0.001。
图6a-6g。b16小鼠黑素瘤肿瘤中的garp上调降低了过继性t细胞免疫疗法的效果。(6a)实验方案。(6b)b16-garp-fc和b16-ev的平均肿瘤生长动力学(n=6)。(6c)如所示的两个实验组之间存活率的差异。(6d)由cd8+cd90.1+表面标志物指示的外周血中抗原特异性供体t细胞的代表性facs图。(6e)act后不同时间点荷瘤小鼠外周血中供体t细胞的频率。(6f)响应于同源gp100肽刺激的外周血抗原特异性供体t细胞的细胞内ifnγ染色的代表性facs图。(6g)定量接受b16-garp-fc或b16-ev的小鼠外周血中产生ifnγ的供体t细胞的频率。通过对数秩检验计算图6c中的p值。使用两个样品t检验来比较其他图中的组差异。*p<0.05。***p<0.001。
图7a-7e。garp特异性抗体阻断外源ltgf-β与garp的结合,并具有针对乳腺癌临床前模型的治疗价值。(7a)通过各种garp抗体对稳定表达人garp(pre-b-hgarp)的前b细胞进行表面染色。灰色直方图表示用同种型对照抗体染色。(7b)在各种garp抗体或同种型对照抗体的存在下,不与或与人ltgf-β(hultgf-β)一起孵育前b-hgarp细胞,然后对细胞表面hultgf-β染色。(7c)balb/c小鼠原位注射4t1-hugarp乳腺肿瘤,然后给予指示治疗方案。显示的数据是在接种后35天(左图,联合治疗组)或30天(右图,单独的抗体治疗)分离的原发肿瘤重量的比较。(7d)单独的抗garp抗体治疗(左图)或与cy联合对肺中转移性肿瘤结节数量的影响。(7e)用同种型对照抗体或抗garp抗体(4d3)处理的小鼠脾脏中的脾treg和mdsc的百分比。使用两个样品t检验来比较所有图中的组差异。*p<0.05。***p<0.001。
图8a-8f。血小板固有的garp在产生活性tgfβ中起关键作用。(8a)血小板的消耗导致活性和总tgfβ的完全丧失。(8b-8d)指示的小鼠模型中garp和lap的表达。与wt血小板相比,来自plt-tgfβ1ko小鼠的血小板表达相似水平的表面garp-tgfβ1复合物。(8e)测量小鼠中的活性tgfβ。在wt小鼠中,与血浆相比,活性tgfβ在血清中升高。(8f)测量小鼠中的总tgfβ。在plt-tgfβ1ko小鼠中血清中的总潜伏tgfβ水平降低,但在plt-gp96ko或plt-garpko小鼠中未降低。
图9a-9d。在wt,plt-tgfβ1ko和plt-garpko受体小鼠中过继性t细胞治疗黑素瘤的功效。(9a)与wt小鼠相比,肿瘤生长在plt-garpko小鼠中得到更有效地控制。plt-garpko小鼠外周血中pmel细胞增强的持久性(9b)和功能(9c)。(9d)plt-tgfβ1ko小鼠,其血小板表达garp并且仍然能够激活tgfβ,对肿瘤没有改善的控制。
图10a-10h。血小板衍生的garp-tgfβ复合物钝化抗肿瘤t细胞免疫。(10a-10c)wt和plt-garpko小鼠的肿瘤大小(10a)和总存活率。与wt小鼠相比,plt-garpko小鼠中mc38的生长显著减少。(10d)带有mc38的plt-garpko小鼠具有降低的活性tgfβ的血清水平。(10e-10f)mc38肿瘤切片中p-smad2/3(p-smad2/3)的免疫组织化学染色显示plt-garpko小鼠中mc38细胞中tgfβ信号传导的显著减弱。(10g)plt-garpko小鼠中全身性骨髓来源的抑制细胞(10h)和肿瘤浸润性调节性t细胞的减少。
图11a-11d。抗血小板药剂加强癌症的过继性t细胞疗法。(11a)cy和ap对肿瘤生长的影响(左)。抗血小板剂和过继性t细胞转移对b16-f1非常有效,大多数小鼠的无复发存活超过3个月(右)。(11b)在同时接受抗血小板治疗和act的小鼠的血液,腹股沟淋巴结(iln)和脾脏中维持较高数量的抗原特异性t细胞。(11c)当转移的t细胞缺乏ifnγ(11d)或给予抗-ifnγ中和抗体时,抗血小板剂没有益处。
说明性实施方式的描述
本文证明,膜结合和可溶性garp均由人癌细胞广泛表达,但较少被正常上皮细胞表达,并且garp的表达与癌症晚期和不良预后相关。此外,发现garp本身具有转化潜力,这使得正常的乳腺上皮细胞发生肿瘤。观察到癌细胞中的garp表达导致tgf-β活性增加,这可能是由于其能够将ltgf-β浓缩成顺式和反式,从而有助于癌症侵袭性和转移。肿瘤微环境中的garp表达促进了调节性t细胞的诱导,从而削弱了效应t细胞对癌症的功能。然而,通过阻断其结合tgf-β的能力来中和garp甚至在没有化学疗法的情况下导致抗癌活性。特别地,这里提供了新的抗体分子,4d3和5c5抗体,其可以有效地结合并中和garp。因此,实施方式的抗体可用于治疗癌症和增强免疫应答的方法(例如,与过继性t细胞疗法结合)。
虽然t细胞疗法具有通过识别和攻击肿瘤细胞来治疗癌症的潜力,但是肿瘤微环境可以通过诱导调节性t细胞来逃避免疫系统,这削弱了过继转移的效应t细胞控制癌症的能力。因此,本发明的实施方式通过提供治疗癌症的方法克服了与现有技术相关的挑战,所述方法包括t细胞疗法和抗血小板剂的组合。在该方法中,抗血小板剂可以加强肿瘤的过继性t细胞疗法,因为已经显示从活化的血小板分泌的可溶性因子抑制t细胞。例如,已经显示血小板分泌的潜伏tgfβ和garp可以导致癌细胞对过继性t细胞疗法的耐药性。因此,抗血小板因子如抗garp单克隆抗体(可以阻断tgfβ结合)可以与t细胞疗法联合使用以克服这种耐药性并治疗癌症。此外,其他免疫疗法如免疫检查点抑制剂可与t细胞疗法和抗血小板剂联合使用以增强免疫应答。
i.定义
“治疗”或“处理”指向对象给予或施加治疗剂或在对象上实施方法或疗法以获得疾病的治疗益处或健康相关的状况。例如,治疗可能包括给予药物上有效量的抑制garp信号传导的抗体。在另一个实例中,治疗可包括给予t细胞疗法和药学有效量的抗血小板剂(例如,抑制garp信号传导的抗体)。
“对象”和“患者”指人类或非人类,例如灵长类动物、哺乳动物、和脊椎动物。在特定的实施方式中,该对象是人。
本文所用术语“治疗益处”或“治疗有效”指就这种病情的医学治疗而言,促进或增强对象的健康状况的任何情况。这包括但不限于,在疾病的征兆或症状的频率或严重程度上的降低。例如,癌症的治疗可能涉及,例如,肿瘤尺寸的减小、肿瘤侵袭性的降低、癌症生长速率的降低,或转移的预防。癌症的治疗可能还涉及延长癌症患者的生存期。
“抗癌”试剂能对对象的癌细胞/肿瘤产生不利影响,例如通过杀死癌细胞、诱导癌细胞凋亡、降低癌细胞生长速率、降低转移的发生率或数量、降低肿瘤尺寸、抑制肿瘤生长、减少对肿瘤或癌细胞的血液供给、促进对癌细胞或肿瘤的免疫应答、阻止或抑制癌症发展、或延长患癌对象的寿命。
本文所述术语“抗体”以最广泛含义使用,并且特别涵盖单克隆抗体(包括全长单克隆抗体)、多克隆抗体、多特异性抗体(例如,双特异性抗体),和抗体片段,只要其显示所需的生物活性即可。
本文所用术语“单克隆抗体”指获自基本均一抗体群体的抗体,例如除了可能少量存在的突变如天然产生的突变外,群体包含的单独抗体是相同的。因此,定语“单克隆”表示所述抗体特征为不是离散抗体的混合物。在某些实施方式中,这类单克隆抗体通常包括含有结合靶标的多肽序列的抗体,其中结合靶标的多肽序列通过这样的过程获得,所述过程包括由多个多肽序列选择单个结合靶标的多肽序列。例如,选择过程可以是由多个克隆(如杂交瘤克隆、噬菌体克隆或重组dna克隆的库)选择独特的克隆。应当理解,可以进一步改变所选靶标结合序列,例如,改进其对所述靶标的亲和性,将所述靶标结合序列人源化,改进其在细胞培养中的生产,降低其体内的免疫原性,产生多特异性抗体等,并且含有经改变靶标结合序列的抗体也是本发明的单克隆抗体。不同于通常包含针对不同决定簇(表位)的不同抗体的多克隆抗体制剂,单克隆抗体制剂中的各单克隆抗体均针对抗原上的单一决定簇。除了它们的特异性之外,单克隆抗体制备的优点在于它们通常不被其他免疫球蛋白污染。
词组“药学或药理学上可接受的”指分子实体和组合物在恰当地给予动物,例如人时不产生不利、过敏性或其它不良反应。根据本公开,本领域技术人员已知包括抗体或其它活性成分的药物组合物的制备。另外,就动物(如人)给药而言,理解制备应该满足由fda官方生物学标准要求的无菌性、致热原性、常规安全和纯度标准。
本文使用的“药学上可接受的运载体”包含任何和所有水性溶剂(例如,水,酒精性/水性溶液,盐溶液,肠道外载剂,如氯化钠,林格氏右旋糖等),非水性溶剂(例如,丙二醇,聚乙二醇,植物油和可注射的有机酯,如乙酸乙酯),分散介质,包衣、表面活性剂、抗氧化剂、防腐剂(例如,抗菌剂或抗真菌剂,抗氧化剂,螯合剂和惰性气体),等渗剂,吸收延迟剂,盐,药物,药物稳定剂,凝胶,粘合剂,赋形剂,崩解剂,润滑剂、甜味剂,调味剂,染料,液体和营养补充剂,诸如此类的物质和其组合,此应为本领域普通技术人员已知。根据已知参数调节药物组合物中各种组分的ph和精确的浓度。
术语“单位剂量”或“剂量”是指适用于对象的物理上离散的单位,每个单位含有预定量的治疗组合物,其经计算以产生与其给予(即适当的路线和治疗方案)相关的上述讨论的所需响应。根据治疗次数和单位剂量,给药量取决于所需的效果。给予患者或对象的本发明实施方式的组合物的实际剂量可以由身体和生理因素来确定,例如体重,对象的年龄,健康和性别,所治疗疾病的类型,疾病渗透程度,先前或同时的治疗干预,患者的特发性疾病,给药途径以及特定治疗物质的效力,稳定性和毒性。例如,剂量还可包含每次给予约1μg/kg/体重至约1000mg/kg/体重(此范围包括间插剂量)或更多,以及其中可衍生的任何范围。在本文列举的数字衍生范围的非限定示例中,能给药的范围是约5μg/kg/体重至约100mg/kg/体重,约5μg/kg/体重至约500mg/kg/体重等。在任何事件中,负责给予的医师将确定组合物中活性成分的浓度,以及对于个体对象合适的剂量。
本文所用术语“接触”和“暴露”用于细胞时,描述治疗构建体和化疗或放疗试剂递送给靶细胞或放置成与所述靶细胞直接并置所用的方法。为了实现细胞杀死,例如,以有效杀死所述细胞或阻止其分裂的结合量将两种试剂递送给细胞。
术语“免疫检查点”是指免疫系统中的分子,例如蛋白质,其向其组分提供抑制信号以平衡免疫响应。已知的免疫检查点蛋白包括ctla-4,pd1及其配体pd-l1和pd-l2,以及lag-3,btla,b7h3,b7h4,tim3,kir。涉及lag3,btla,b7h3,b7h4,tim3和kir的途径在本领域中被认为构成类似于ctla-4和pd-1依赖途径的免疫检查点途径(参见例如ppardoll,2012.naturerevcancer12:252-264;mellman等,2011.nature480:480-489)。
“免疫检查点抑制剂”是指抑制免疫检查点蛋白功能的任何化合物。抑制包括功能减少和完全阻滞。特别地,免疫检查点蛋白是人免疫检查点蛋白。因此,免疫检查点蛋白抑制剂特别是人免疫检查点蛋白的抑制剂。
实施方式的抗体
在某些实施方式中,考虑了结合至少一部分garp蛋白并抑制garp信号传导和癌细胞增殖的抗体或其片段。如本文所用,术语“抗体”旨在广泛地指任何免疫结合剂,例如igg,igm,iga,igd,ige和遗传修饰的igg以及包含保留抗原结合活性的抗体cdr结构域的多肽。抗体可以选自嵌合抗体,亲和力成熟抗体,多克隆抗体,单克隆抗体,人源化抗体,人抗体,或抗原结合抗体片段或天然或合成配体。优选地,抗garp抗体是单克隆抗体或人源化抗体。
因此,通过已知方法并如本文所述,可以产生多克隆或单克隆抗体,抗体片段,以及结合结构域和cdr(包括任何前述的工程化形式),它们对garp蛋白、其各自的一个或多个表位、或任何前述的偶联物具有特异性,无论这些抗原或表位是从天然来源分离或是合成衍生物或是天然化合物的变体。
适用于本发明实施方式的抗体片段的实例包括但不限于:(i)fab片段,由vl、vh、cl和ch1结构域构成;(ii)“fd”片段,由vh和ch1结构域构成;(iii)“fv”片段,由单-抗体的vl和vh结构域构成;(iv)“dab”片段,由vh结构域构成;(v)分离的cdr区域;(vi)f(ab′)2片段,包含两个连接的fab片段的二价片段;(vii)单链fv分子(“scfv”),其中vh结构域和vl,结构域通过能使这两个结构域联合以形成结合结构域的肽接头连接;(viii)双特异性单链fv二聚体(参见美国专利号5,091,513)和(ix)通过基因融合构建的双抗体、多价或多特异性片段(美国专利公开号20050214860)。fv、scfv或双抗体分子可通过纳入连接vh和vl结构域的二硫桥来稳定。还可制备包含与ch3结构域相连的scfv的小体(minibody)(hu等,1996)。
在实施方式中还考虑了抗体样结合肽模拟物。liu等,(2003)描述了“抗体样结合肽模拟物”(abip),其是作为减少的抗体起作用的肽并且具有较长血清半衰期以及较简便合成方法的某些优点。
可以用抗原(例如garp细胞外结构域(ecd)蛋白)接种动物,以产生对garp蛋白特异的抗体。通常,抗原与另一分子结合或偶联以增强免疫应答。如本文所用,偶联物是与抗原结合的任何肽,多肽,蛋白质或非蛋白质物质,其用于在动物中引发免疫应答。响应于抗原接种在动物中产生的抗体包含多种不同的分子(多克隆抗体),其由多种单独的抗体产生b淋巴细胞制成。多克隆抗体是抗体种类的混合群体,每个抗体种类可以识别相同抗原上的不同表位。鉴于动物中多克隆抗体产生的正确条件,动物血清中的大多数抗体将识别动物已经免疫的抗原化合物上的集合表位。通过亲和纯化以仅选择识别目标抗原或表位的那些抗体从而进一步增强这种特异性。
单克隆抗体是单一种类的抗体,其中每个抗体分子识别相同的表位,因为所有产生抗体的细胞都来源于单个b淋巴细胞细胞系。产生单克隆抗体(mab)的方法通常以与制备多克隆抗体相同的细胞系开始。在一些实施方式中,啮齿动物如小鼠和大鼠用于产生单克隆抗体。在一些实施方式中,兔,绵羊或青蛙细胞用于产生单克隆抗体。大鼠的使用是众所周知的并且可以提供某些优点。常规使用小鼠(例如,balb/c小鼠)并且通常给出高百分比的稳定融合。
杂交瘤技术涉及将先前用garp抗原免疫的小鼠的单个b淋巴细胞与永生化骨髓瘤细胞(通常是小鼠骨髓瘤)融合。该技术提供了将单个抗体产生细胞繁殖无限代的方法,从而可以产生无限量的具有相同抗原或表位特异性的结构相同的抗体(单克隆抗体)。
血浆b细胞(cd45+cd5-cd19+)可以从新鲜制备的免疫兔的兔外周血单核细胞中分离,并进一步针对garp结合细胞选择。在富集产生抗体的b细胞后,可以分离总rna并合成cdna。可以扩增来自重链和轻链的抗体可变区的dna序列,构建到噬菌体展示fab表达载体中,并转化到大肠杆菌中。可以通过多轮富集淘选和测序来选择garp特异性结合fab。选择的garp结合命中可以在人胚胎肾(hek293)细胞(英杰公司(invitrogen))中使用哺乳动物表达载体系统表达为兔和兔/人嵌合形式的全长igg,并使用蛋白g树脂和快速蛋白液相色谱(fplc)分离单元纯化。
在一个实施方式中,抗体是嵌合抗体,例如,包含来自非人供体的抗原结合序列的抗体,所述非人供体被移植到异源非人、人或人源化序列(例如,框架和/或恒定域序列)。已经开发了用人源的类似结构域取代单克隆抗体的轻链和重链恒定结构域的方法,使外源抗体的可变区保持完整。或者,在针对人免疫球蛋白基因转基因的小鼠中产生“完全人”单克隆抗体。还开发了通过重组构建具有啮齿动物(例如小鼠)和人氨基酸序列的抗体可变结构域将单克隆抗体的可变结构域转化为更多人形式的方法。在“人源化”单克隆抗体中,只有高变cdr来自小鼠单克隆抗体,并且框架和恒定区来源于人氨基酸序列(参见美国专利号5,091,513和6,881,557)。据认为,用在人抗体的相应位置中发现的氨基酸序列替换抗体中具有啮齿动物特征的氨基酸序列将降低治疗使用期间不良免疫反应的可能性。还可对产生抗体的杂交瘤或其它细胞作出遗传突变或其它改变,从而可改变或不改变由杂交瘤产生的抗体的结合特异性。
用于在各种动物物种中产生多克隆抗体以及用于产生各种类型的单克隆抗体(包括人源化,嵌合和完全人)的方法是本领域熟知的并且是高度可预测的。例如,以下美国专利和专利申请提供了对这些方法的可能描述:美国专利申请号2004/0126828和2002/0172677;和美国专利号3,817,837;3,850,752;3,939,350;3,996,345;4,196,265;4,275,149;4,277,437;4,366,241;4469797;4,472,509;4606855;4703003;4742159;4767720;4,816,567;4867973;4,938,948;4,946,778;5021236;5164296;5196066;5,223,409;5,403,484;5420253;5,565,332;5,571,698;5,627,052;5656434;5770376;5789208;5,821,337;5844091;5858657;5861155;5871907;5,969,108;6,054,297;6165464;6365157;6406867;6709659;6709873;6753407;6814965;6849259;6861572;6875434;和6,891,024。本文及其中引用的所有专利,专利申请出版物和其他出版物均通过引用纳入到本申请中。
抗体可以从任何动物来源产生,包括鸟类和哺乳动物。优选地,抗体是绵羊,鼠(例如小鼠和大鼠),兔,山羊,豚鼠,骆驼,马或鸡抗体。此外,更新的技术允许从人组合抗体文库开发和筛选人抗体。例如,噬菌体抗体表达技术允许在没有动物免疫的情况下产生特异性抗体,如美国专利号4,522,588中所述,其通过引用并入本文。这些技术进一步描述于:marks(1992);stemmer(1994);gram等(1992);barbas等(1994);和schier等(1996)。
完全可以预期,无论动物种类,单克隆细胞系或其他抗体来源如何,针对garp抗体都能够中和或抵消garp的作用。某些动物物种可能不太适合产生治疗性抗体,因为它们更可能由于通过抗体的“fc”部分激活补体系统而引起过敏反应。然而,可以将完整抗体酶促消化成“fc”(补体结合)片段,并消化成具有结合结构域或cdr的抗体片段。去除fc部分降低了抗原抗体片段引发不希望的免疫应答的可能性,因此,没有fc的抗体可优先用于预防性或治疗性治疗。如上所述,抗体也可以构建成嵌合的或部分或完全人的,以减少或消除由于向动物给予已在其他物种中产生或具有其他物种序列的抗体而导致的不良免疫后果。
取代变体通常在蛋白质内的一个或多个位点含有一个氨基酸被另一个氨基酸替换,并且可以设计为调节多肽的一种或多种性质,有或没有其他功能或性质的丧失。取代可以是保守的,即,一个氨基酸被替换为具有相似形状和电荷的氨基酸。保守取代是本领域熟知的,包括例如:丙氨酸变为丝氨酸;精氨酸变为赖氨酸;天冬酰胺变为谷氨酰胺或组氨酸;天冬氨酸变为谷氨酸;半胱氨酸变为丝氨酸;谷氨酰胺变为天冬酰胺;谷氨酸变为天冬氨酸;甘氨酸变为脯氨酸;组氨酸变为天冬酰胺或谷氨酰胺;异亮氨酸变为亮氨酸或缬氨酸;亮氨酸变为缬氨酸或异亮氨酸;赖氨酸变为精氨酸;蛋氨酸变为亮氨酸或异亮氨酸;苯丙氨酸变为酪氨酸,亮氨酸或蛋氨酸;丝氨酸变为苏氨酸;苏氨酸变为丝氨酸;色氨酸变为酪氨酸;酪氨酸变为色氨酸或苯丙氨酸;和缬氨酸变为异亮氨酸或亮氨酸。或者,取代可以是非保守的,使得多肽的功能或活性受到影响。非保守变化通常涉及用化学上不相似的残基取代残基,例如用于非极性或不带电荷的氨基酸取代极性或带电氨基酸,反之亦然。
蛋白质可以是重组的,或在体外合成的。或者,可以从细菌中分离非重组或重组蛋白质。还预期含有这种变体的细菌可以在组合物和方法中实施。因此,不需要分离蛋白质。
预期在组合物中,每ml含有约0.001mg至约10mg的总多肽,肽和/或蛋白质。因此,组合物中蛋白质的浓度可为约,至少约或至多约0.001,0.010,0.050,0.1,0.2,0.3,0.4,0.5,0.6,0.7,0.8,0.9,1.0,1.5,2.0,2.5,3.0,3.5,4.0,4.5,5.0,5.5,6.0,6.5,7.0,7.5,8.0,8.5,9.0,9.5,10.0mg/ml或更高(或其中可衍生的任何范围)。其中,约,至少约,或至多约1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99或100%可以是结合garp的抗体。
抗体或优选抗体的免疫部分可以与其他蛋白质化学偶联,或表达为含其他蛋白质的融合蛋白。为了本说明书和所附权利要求的目的,所有这些融合蛋白都包括在抗体或抗体的免疫部分的定义中。
实施方式提供针对garp,多肽和肽的抗体和抗体样分子,其与至少一种试剂连接以形成抗体偶联物或有效负载。为了增加抗体分子作为诊断剂或治疗剂的功效,通常与至少一个所需分子或部分连接或共价结合或复合。这种分子或部分可以是,但不限于,至少一种效应或报道分子。效应分子包含具有所需活性(例如细胞毒活性)的分子。已附着于抗体的效应分子的非限制性实例包括毒素,治疗性酶,抗生素,放射性标记的核苷酸等。相反,报告分子定义为可以使用测定法检测的任何部分。已与抗体偶联的报告分子的非限制性实例包括酶,放射性标记,半抗原,荧光标记,磷光分子,化学发光分子,发色团,发光分子,光亲和分子,有色颗粒或配体,例如生物素。
本领域已知几种方法用于抗体与其偶联物部分的连接或偶联。一些结合方法涉及使用金属鳌合复合物,例如采用与抗体结合的有机螯合剂如二乙烯三胺五乙酸酐(dtpa);乙二胺四乙酸;n-氯-对-甲苯磺酰胺;和/或四氯-3-6-二苯基甘脲-3。在偶联剂(例如戊二醛或高碘酸盐)存在时,单克隆抗体也可以与酶反应。在这些偶合剂的存在下或通过与异硫氰酸盐反应制得含荧光标记物的偶联物。
ii.t细胞疗法
本公开的某些实施方式涉及获得t细胞以及向对象给予t细胞作为靶向癌症细胞的免疫疗法。在过去的二十年中已经描述了几种用于功能性抗肿瘤效应t细胞的衍生,活化和扩增的基本方法。这些方法包括:自体细胞,如肿瘤浸润淋巴细胞(til);使用自体dc,淋巴细胞,人工抗原呈递细胞(apc)或涂有t细胞配体和活化抗体的珠子或通过捕获靶细胞膜分离的细胞离体活化的t细胞;天然表达抗宿主肿瘤t细胞受体(tcr)的同种异体细胞;和非肿瘤特异性自体或同种异体细胞,其经遗传重编程或“重定向”以表达肿瘤反应性tcr或嵌合tcr分子,其显示称为“t-体”的抗体样肿瘤识别能力。这些方法已经产生了许多用于t细胞制备和免疫的方案,其可以用于本公开的方法中。
t细胞制备
在一些实施方式中,t细胞衍生自血液、骨髓、淋巴或淋巴器官。在一些方面中,细胞是人细胞。细胞一般是原代细胞,如直接从对象分离和/或从对象分离并冷冻的那些细胞。在一些实施方式中,细胞包括t细胞或其他细胞类型的一个或多个亚组,如全t细胞群、cd4+细胞、cd8+细胞及其亚群,如由功能、活化状态、成熟、分化潜力、扩增、再循环、定位,和/或持续能力、抗原特异性、抗原受体类型、特定器官或区室中的存在、标志物或细胞因子分泌概况,和/或分化程度限定的那些。关于待治疗的对象,细胞可以是同种异体和/或自体的。某些方面,如对于现成的技术,细胞是多潜能和/或多能的,如干细胞,如诱导性多潜能干细胞(ipsc)。在一些实施方式中,该方法包括从对象分离细胞,对其进行制备、处理、培养和/或工程改造,如本文所述,并且在冷冻保存前或后将其回输给同一患者。
t细胞(例如,cd4+和/或cd8+t细胞的亚型和亚群)为原初t(tn)细胞、效应t细胞(teff)、记忆t细胞及其亚型,如干细胞记忆t(tscm)、中心记忆t(tcm)、效应记忆t(tem)、或终末分化的效应记忆t细胞、肿瘤-浸润性淋巴细胞(til)、不成熟t细胞、成熟t细胞、辅助t细胞、细胞毒性t细胞、粘膜相关的非变体t(mait)细胞、天然产生的和过继性调节t(treg)细胞、辅助t细胞,如th1细胞、th2细胞、th3细胞、th17细胞、th9细胞、th22细胞、滤泡辅助t细胞、α/βt细胞,和δ/γt细胞。
在一些实施方式中,在一种或多种t细胞群中富集或消耗这样的细胞,所述细胞对于特异性标志物如表面标志物呈阳性或对特异性标志物呈阴性。在一些情况中,此类标志物在某些t细胞群(例如,非记忆细胞)上不存在或以相对较低的水平表达,但在某些其它t细胞群(例如,记忆细胞)上存在或以相对较高的水平表达的那些。在一个实施方式中,对所述细胞(例如,cd8+细胞或cd3+细胞)富集(即,正选择)cd45ro、ccr7、cd28、cd27、cd44、cd127和/或cd62l呈阳性或表达高表面水平的cd45ro、ccr7、cd28、cd27、cd44、cd127和/或cd62l的细胞,和/或,消耗(例如,负选择)cd45ra阳性或表达高表面水平的cd45ra的细胞。在一些实施方式中,对细胞富集或消耗cd122、cd95、cd25、cd27和/或il7-rα(cd127)阳性细胞或表达高表面水平的cd122、cd95、cd25、cd27和/或il7-ra(cd127)的细胞。在一些实例中,对cd8+t细胞富集cd62l和cd45ro阳性(或cd45ra阴性)细胞。
在一些实施方式中,通过对非t细胞(例如b细胞、单核细胞或其它血液白细胞)上表达的标志物(例如cd14)进行负选择来从pbmc样品分离t细胞。某些方面,使用cd4+或cd8+选择步骤来分离cd4+辅助t细胞和cd8+细胞毒性t细胞。通过针对一种或多种原初、记忆和/或效应t细胞亚群上表达或以相对高表达的标志物进行正或负选择,可将此类cd4+和cd8+群进一步分选成亚群。
在一些实施方式中,在cd8+细胞中进一步富集或消除原初、中心记忆、效应记忆和/或中心记忆干细胞,例如通过基于相应亚群相关表面抗原的正选择或负选择。在一些实施方式中,富集中心记忆t(tcm)细胞以提高功效,如以改善长期存活、扩增和/或给予后的植入,某些方面,其在此类亚群中特别强势。参见terakura等.(2012)blood.1:72-82;wang等(2012)jimmunother.35(9):689-701。在一些实施方式中,将tcm-富集的cd8+t细胞与cd4+t细胞合并进一步增强功效。
在一些实施方式中,t细胞是自体的细胞。在该方法中,肿瘤样品获自患者,并且获得单细胞悬浮液。单细胞悬浮液可以任何合适的方式获得,例如,机械地(使用例如加利福尼亚州奥本的美天旎生物技术公司(miltenyibiotec,auburn,calif.)的gentlemacstm分解剂使肿瘤分解)或酶促地(例如,胶原酶或dna酶)。将肿瘤酶促消化物的单细胞悬浮液于白细胞介素-2(il-2)中培养。培养细胞直到铺满(例如,约2×106个淋巴细胞),例如,约5至约21天,优选约10至约14天。例如,细胞可以培养5天,5.5天,或5.8天至21天,21.5天或21.8天,例如10天,10.5天,或10.8天至14天,14.5天或14.8天。
培养的t细胞可以经汇集并快速扩增。快速扩增在约10至约14天,优选约14天的时间段内使抗原特异性t细胞的数量至少增加约50倍(例如,50倍、60倍、70倍、80倍、90倍、或100倍或更多)。具体而言,快速扩增在约10至约14天,优选约14天的时间段内产生至少约200倍(例如,200倍、300倍、400倍、500倍、600倍、700倍、800倍、900倍或更多)的增长。
扩增可以通过本领域已知的许多方法中的任何一种来完成。例如,在饲养淋巴细胞和白细胞介素-2(il-2)或白细胞介素-15(il-15),优选il-2存在下,使用非特异性t细胞受体刺激可以快速扩增t细胞。非特异性t细胞受体刺激可包括约30ng/ml的okt3,一种小鼠单克隆抗cd3抗体(可从
可以修饰自体t细胞以表达促进自体t细胞的生长和活化的t细胞生长因子。合适的t细胞生长因子包括,例如,白细胞介素(il)-2,il-7,il-15和il-12。修饰的合适方法是本领域已知的。参见例如sambrook等,《分子克隆:实验室手册》(molecularcloning:alaboratorymanual)(第3版),纽约冷泉港,冷泉港实验室出版社,2001;和ausubel等,《新编分子生物学实验指南》(currentprotocolsinmolecularbiology),纽约,格林出版联合公司和约翰韦利父子公司,1994。在特定方面,修饰的自体t细胞以高水平表达t细胞生长因子。t细胞生长因子编码序列,例如il-12的编码序列,在本领域中是容易获得的,启动子也是如此,其与t细胞生长因子编码序列的可操作连接促进高水平表达。
遗传工程改造的抗原受体
可以对t细胞进行基因工程改造以表达抗原受体,例如工程化的tcr和/或嵌合抗原受体(car)。例如,修饰自体t细胞以表达对癌抗原具有抗原特异性的t细胞受体(tcr)。合适的tcr包括,例如,对黑素瘤抗原具有抗原特异性的那些,例如gp100或mart-1。修饰的合适方法是本领域已知的。例如,参见sambrook和ausube1,同上。例如,使用heemskerk等,humgenether.19:496-510(2008)和johnson等,blood114:535-46(2009)中所述转导技术,t细胞可经转导以表达对癌症抗原具有抗原特异性的t细胞受体(tcr)。
在一些实施方式中,t细胞包括通过遗传工程改造导入的一种或多种核酸,其编码一种或多种抗原受体,和这类核酸的遗传工程改造的产物。在一些实施方式中,核酸是异源性的,即,通常不存在于细胞或从该细胞获得的样品中,如从另一个生物体或细胞获得的样品,其例如在待工程改造中的细胞和/或这类细胞来源的生物体中通常未发现。在一些实施方式中,核酸不是天然存在的,例如天然中不存在的核酸(例如嵌合的)。
在一些实施方式中,car含有特异性结合至抗原的胞外抗原-识别结构域。在一些实施方式中,抗原是在细胞表面上表达的蛋白质。在一些实施方式中,car是tcr-样car并且抗原是经处理的肽抗原,如胞内蛋白质的肽抗原,其像tcr那样在主要组织相容性复合物(mhc)分子中在细胞表面上识别。
示例性的抗原受体,包括car和重组tcr,以及用于工程改造和将受体导入细胞中的方法,包括例如,在国际专利申请公开号wo200014257、wo2013126726、wo2012/129514、wo2014031687、wo2013/166321、wo2013/071154、wo2013/123061,美国专利申请公开号us2002131960、us2013287748、us20130149337,美国专利号6,451,995、7,446,190、8,252,592、8,339,645、8,398,282、7,446,179、6,410,319、7,070,995、7,265,209、7,354,762、7,446,191、8,324,353、和8,479,118,和欧洲专利申请号ep2537416中所述的那些,和/或sadelain等,cancerdiscov.2013年4月;3(4):388-398;davila等,(2013)plosone8(4):e61338;turtle等,curr.opin.immunol.,2012年10月;24(5):633-39;wu等,cancer,2012年3月18(2):160-75中所述的那些。在一些方面中,遗传工程改造的抗原受体包括美国专利号7,446,190中所述的car,和国际专利申请公开号wo/2014055668a1中所述的那些。
在一些方面中,肿瘤抗原是人端粒酶逆转录酶(htert)、生存素、小鼠双微体2同源物(mdm2)、细胞色素p4501b1(cyp1b)、her2/neu、维尔姆氏(wilms’)肿瘤基因1(wt1)、活素(1ivin)、α胎甲球蛋白(afp)、癌胚抗原(cea)、粘蛋白16(muc16)、muc1、前列腺-特异性膜抗原(psma)、p53或细胞周期蛋白(d1)。例如,靶抗原是htert或生存素。在一些方面中,靶抗原是cd38。在其他方面中,靶抗原是cd33或tim-3。在其他方面中,其是cd26、cd30、cd53、cd92、cd148、cd150、cd200、cd261、cd262、或cd362。在一些实施方式中,工程改造的细胞可含有靶向一种或多种其他抗原的抗原。在一些实施方式中,一种或多种其他抗原是肿瘤抗原或癌症标志物。其他抗原包括:孤儿酪氨酸激酶受体ror1、tegfr、her2、l1-cam、cd19、cd20、cd22、间皮素、cea、和乙型肝炎表面抗原、抗叶酸受体、cd23、cd24、cd30、cd33、cd38、cd44、egfr、egp-2、egp-4、epha2、erbb2、3、或4、fbp、胎儿乙酰胆碱e受体、gd2、gd3、hmw-maa、il-22r-α、il-13r-α2、kdr、κ轻链、lewisy、l1-细胞粘附分子、mage-a1、间皮素、muc1、muc16、psca、nkg2d配体、ny-eso-1、mart-1、gp100、癌胚胎抗原、ror1、tag72、vegf-r2、癌胚抗原(cea)、前列腺特异性抗原、psma、her2/neu、雌激素受体、孕酮受体、肝配蛋白b2、cd123、c-met、gd-2,和magea3、ce7、维尔姆斯瘤(wt-1)、细胞周期蛋白如细胞周期蛋白a1(ccna1)和/或生物素化的分子,和/或通过hiv、hcv、hbv或其他病原体表达的分子。
嵌合抗原受体
在一些实施方式中,工程改造的抗原受体包括嵌合抗原受体(car),包括活化或刺激性car,共刺激性car(参见wo2014/055668),和/或抑制性car(icar,参见fedorov等,sci.transl.medicine,5(215)(2013年12月))。在一些方面中,car一般包括通过接头和/或跨膜结构域与一种或多种胞内信号转导组分连接的胞外抗原(或配体)结合结构域。此类分子通常模拟或近似通过天然抗原受体的信号、通过此类受体联合共刺激受体的信号、和/或通过单独共刺激受体的信号。
在一些实施方式中,构建car以使其具有针对特定抗原(或标志物或配体)的特异性,如在待通过过继治疗靶向的特定细胞类型中表达的抗原,例如癌症标志物,和/或意图用以诱导减弱应答的抗原,如在正常或非疾病细胞类型上表达的抗原。因此,car一般在其胞外部分中包合一种或多种抗原结合分子,例如一种或多种抗原-结合片段、结构域或部分,或一种或多种抗体可变结构域,和/或抗体分子。在一些实施方式中,所述car包括抗体分子的一个或多个抗原-结合部分,例如源自单克隆抗体(mab)的可变重(vh)链和可变轻(vl)链的单链抗体片段(scfv)。
在一些方面中,所述抗原特异性结合或识别组分连接至一种或多种跨膜和胞内信号转导结构域。在一些实施方式中,car包括与该car的胞外结构域融合的跨膜结构域。在一个实施方式中,使用天然关联car中结构域之一的跨膜结构域。在一些情况中,跨膜结构域经氨基酸取代修饰或选择,以避免这类结构域结合相同或不同表面膜蛋白的跨膜结构域,尽可能减少与受体复合物其他成员的相互作用。
在一些实施方式中,所述跨膜结构域源自天然或合成来源。当所述来源是天然来源时,某些方面,所述结构域源自任何膜结合或跨膜蛋白。跨膜区包括源自t-细胞受体α、β或ζ链,cd28,cd3ε,cd45,cd4,cd5,cds,cd9,cd16,cd22,cd33,cd37,cd64,cd80,cd86,cd134,cd137,cd154的那些(即包含至少它们的跨膜区域)。或者,在一些实施方式中,所述跨膜结构域是合成的。某些方面,合成跨膜结构域以疏水残基为主,例如亮氨酸和缬氨酸。某些方面,合成跨膜结构域的各末端会有苯丙氨酸、色氨酸和缬氨酸的三联体。
car一般包括至少一种或多种胞内信号转导组分。在一些实施方式中,所述car包括tcr复合物的胞内组分,例如介导t-细胞活化和细胞毒性的tcrcd3+链,例如,cd3ζ链。因此,在一些方面中,所述抗原结合分子连接至一种或多种细胞信号转导模块。在一些实施方式中,细胞信号传导模块包括cd3跨膜结构域、cd3胞内信号传导结构域,和/或其他cd跨膜结构域。在一些实施方式中,所述car还包括一种或多种其它分子的部分,例如fc受体γ、cd8、cd4、cd25或cd16。例如,在一些方面中,所述car包括cd3-ζ(cd3-zeta)或fc受体γ和cd8、cd4、cd25或cd16之间的嵌合分子。
t细胞受体(tcr)
在一些实施方式中,基因工程化改造的抗原受体包括:重组t细胞受体(tcr)和/或从天然存在的t细胞克隆的tcr。“t细胞受体”或“tcr”是指含有可变α和β链(也分别称为tcrα和tcrβ)或可变γ和δ链(也分别称为tcrγ和tcrδ)并且能够特异性结合至与mhc受体结合的抗原肽的分子。在一些实施方式中,tcr是αβ形式。一般而言,以αβ和γδ形式存在的tcr一般在结构上相似,但是表达它们的t细胞具有不同的解剖学位置或功能。tcr可存在于细胞表面上或以可溶形式存在。一般而言,tcr存在于t细胞(或t淋巴细胞)的表面上,其中其一般负责识别与主要组织相容性复合物(mhc)分子结合的抗原。在一些实施方式中,tcr也可含有恒定结构域、跨膜结构域和/或短胞质尾(参见例如,janeway等,《免疫学:健康和疾病中的免疫系统》(immunobiology:theimmunesysteminhealthanddisease),第3版,当代生物学出版社(currentbiologypublications),p.4:33,1997)。例如,在一些方面中,tcr的各链可具有一个n-末端免疫球蛋白可变结构域,一个免疫球蛋白恒定结构域,跨膜结构域,和在c-末端处的短胞质尾。在一些实施方式中,tcr与参与介导信号传导的cd3复合物的非变异蛋白质结合。除非另外说明,术语“tcr”应理解为包括其功能性tcr片段。该术语也包括完整或前场tcr,包括αβ形式或γδ形式的tcr。
因此,出于本文的目的,引用tcr包括任何tcr或功能性片段,如结合至在mhc分子中结合的特异性抗原性肽,即mhc-肽复合物的tcr的抗原结合部分。可互换使用的tcr的“抗原结合部分”或“抗原结合片段”是指含有tcr的结构域的部分但结合全长tcr结合的抗原(例如,mhc-肽复合物)的分子。在一些情况中,抗原结合部分含有tcr的可变结构域,如tcr的可变α链和可变β链,其足以形成用于结合至特异性mhc-肽复合物的结合位点,如一般而言各链含有3个互补决定区的情况。
在一些实施方式中,tcr链的可变结构域结合形成环,或与免疫球蛋白类似的互补决定区(cdr),其通过形成tcr分子的结合位点赋予抗原识别并确定肽特异性。一般而言,像免疫球蛋白那样,cdr由构架区(fr)分开(参见,例如,jores等,proc.nat′lacad.sci.u.s.a.87:9138,1990;chothia等,emboj.7:3745,1988;还参见lefranc等,dev.comp.immunol.27:55,2003)。在一些实施方式中,cdr3是负责识别经加工的抗原的主要cdr,不过也已经显示α链的cdr1与抗原性肽的n-末端部分相互作用,而β链的cdr1与肽的c-末端部分相互作用。cdr2被认为识别mhc分子。在一些实施方式中,β-链的可变区可含有其他超变(hv4)区。
在一些实施方式中,tcr链含有恒定结构域。例如,像免疫球蛋白那样,tcr链(例如,α-链,β-链)的胞外部分可含有2个免疫球蛋白结构域,n-末端处的可变结构域(例如,vα或vβ;一般是氨基酸1至116,基于kabat编号,kabat等,《免疫感兴趣的蛋白质序列》(sequencesofproteinsofimmunologicalinterest),美国国立卫生研究院公共健康服务部(usdept.healthandhumanservices,publichealthservicenationalinstitutesofhealth),1991,第5版),和一个与细胞膜相邻的恒定区(例如,α-链恒定结构域或cα,一般是氨基酸117至259,基于kabat,β-链恒定结构域或cβ,一般是氨基酸117至295,基于kabat)。例如,在一些情况中,由两条链形成的tcr的胞外部分含有两个接近膜的恒定结构域,和两个远离膜的含有cdr的可变结构域。tcr结构域的恒定结构域含有短连接序列,其中半胱氨酸残基形成二硫键,组成两条链间的连接。在一些实施方式中,tcr在α和β链各自可具有额外的半胱氨酸残基,使得tcr在恒定结构域中含有两个二硫键。
在一些实施方式中,tcr链可含有跨膜结构域。在一些实施方式中,跨膜结构域带正电。在一些情况中,tcr链含有胞质尾。在一些情况中,该结构允许tcr与其他分子如cd3结合。例如,含有具有跨膜区的恒定结构域的tcr可锚定细胞膜中的蛋白质并与cd3信号传导器或复合物的非变异亚基结合。
一般而言,cd3是多蛋白质复合物,其可具有哺乳动物中的3种不同链(γ、6、和ε)和ζ-链。例如,在哺乳动物中,复合物可含有cd3γ链、cd3δ链、两条cd3ε链、和cd3ζ链的同二聚体。cd3γ、cd3δ和cd3ε链是免疫球蛋白超家族的高度相关细胞表面蛋白,其包含单个免疫球蛋白结构域。cd3γ、cd3δ和cd3ε链的跨膜区带负电,所述特征使这些链结合带正电的t细胞受体链。cd3γ、cd3δ和cd3ε链的胞内尾各自含有称为基于免疫受体酪氨酸活化基序或itam的单个保守基序,而各cd3ζ链具有三个。一般而言,itam涉及tcr复合物的信号传导能力。这些辅助分子具有带负电的跨膜区并在将信号从tcr传播到细胞中起作用。cd3-和ζ-链与tcr一起形成称为t细胞受体复合物的物质。
在一些实施方式中,tcr可以是2条链α和β(或任选的γ和δ)的异二聚体,或者其可以是单链tcr构建体。在一些实施方式中,tcr是含有,如通过一个或多个二硫键连接的2条单独链(α和β链或γ和δ链)。在一些实施方式中,针对靶抗原(例如,癌症抗原)的tcr经鉴定并导入细胞中。在一些实施方式中,可从多种来源获得编码tcr的核酸,如通过对公开可得的tcrdna序列的聚合酶链反应(pcr)扩增。在一些实施方式中,从生物来源,如来自细胞如来自t细胞(例如,细胞毒性t细胞)、t-细胞杂交瘤或其他公众可得的来源获得tcr。在一些实施方式中,可从体内分离的细胞获得t-细胞。在一些实施方式中,可从患者分离高亲和性t细胞克隆,并分离tcr。在一些实施方式中,t-细胞可以是培养的t-细胞杂交瘤或克隆。在一些实施方式中,针对目标抗原的tcr克隆已在用人免疫系统基因(例如,人白细胞抗原系统,或hla)工程改造的转基因小鼠中产生。参见例如,肿瘤抗原(参见例如,parkhurst等,(2009)clincancerres.15:169-180和cohen等.(2005)jimmunol.175:5799-5808)。在一些实施方式中,使用噬菌体展示分离针对目标抗原的tcr(参见例如,varela-rohena等,(2008)natmed.14:1390-1395和li(2005)natbiotechnol.23:349-354)。在一些实施方式中,tcr或其抗原结合部分可从已知的tcr序列中合成生成。
治疗方法
本实施方式的某些方面可用于预防或治疗与garp信号传导相关的疾病或病症。可以通过任何合适的药物减少garp的信号传导以防止癌细胞增殖。优选地,这些物质是抗garp抗体。
在某些实施方式中,本文提供了用于治疗或延迟个体癌症进展的方法,包括向个体给予有效量的抗血小板剂和t细胞疗法。预期用于治疗的癌症的实例包括肺癌,头颈癌,乳腺癌,胰腺癌,前列腺癌,肾癌,骨癌,睾丸癌,宫颈癌,胃肠癌,淋巴瘤,肺中的肿瘤前病变,结肠癌症,黑色素瘤和膀胱癌。
在一些实施方式中,所述个体患有对一种或多种抗癌疗法具有耐药性(已被证明具有耐药性)的癌症。在一些实施方式中,对抗癌疗法的耐药性包括癌症或难治性癌症的复发。复发可以指治疗后在原始部位或新部位再次出现的癌症。在一些实施方式中,对抗癌疗法的耐药性包括在用抗癌疗法治疗期间癌症的进展。在一些实施方式中,癌症处于早期或晚期。
在本公开内容的方法的一些实施方式中,个体中活化的cd4和/或cd8t细胞的特征在于相对于给予组合之前的产生γ-ifn的cd4和/或cd8t细胞和/或增强的细胞溶解活性。γ-ifn可以通过本领域已知的任何方法测量,包括例如涉及细胞固定,透化和用抗γ-ifn抗体染色的细胞内细胞因子染色(ics)。细胞溶解活性可以通过本领域已知的任何方法测量,例如,使用混合效应细胞和靶细胞的细胞杀伤试验。
t细胞疗法可以在相对于抗血小板剂之前,期间,之后或以各种组合给予。该给予可以以间隔方式进行,其范围从同时到数分钟到数天到数周。在t细胞疗法与抗血小板剂是分开给予患者的实施方式中,一般会确保每次递送的期间不会显著超时,从而使得两种化合物仍然能够对患者发挥有利的联合作用。在这类情况中,可以考虑将抗体疗法和抗癌疗法提供给患者,互相间隔在约12到24或72小时内,更优选地,互相间隔在约6到12小时内。在一些情况中,可能需要显著延长治疗时间,其中各给药之间间隔数天(2、3、4、5、6或7)到数周(1、2、3、4、5、6、7或8)。
在一些实施方式中,可以在t细胞治疗前给予对象非清髓性淋巴细胞消耗化疗。非清髓性淋巴细胞消耗化疗可以是任何合适的此类疗法,其可以通过任何合适的途径给予。例如,非清髓性淋巴细胞消耗化疗可以包括给予环磷酰胺和氟达拉滨,特别是如果癌症是黑素瘤(其可以是转移性的)。给予环磷酰胺和氟达拉滨的示例性途径是静脉注射。同样的,可以给予任何合适剂量的环磷酰胺和氟达拉滨。在具体方面中,给予约60mg/kg环磷酰胺2天,此后给予约25mg/m2氟达拉滨5天。
在某些实施方式中,向对象给予促进自体t细胞生长和活化的t细胞生长因子伴随着自体t细胞或在自体t细胞之后。t细胞生长因子可以是任何合适的生长因子,其促进自体t细胞的生长和活化。合适的t细胞生长因子的示例包括白介素(il)-2、il-7、il-15和il-12,其可以单独或在各种组合物中使用,如il-2和il-7,il-2和il-15,il-7和il-15,il-2,il-7和il-15,il-12和il-7,il-12和il-15,或il-12和il2。il-12是一种优选的t细胞生长因子。
t细胞疗法和抗血小板剂可以通过相同的给药途径或通过不同的给药途径给药。在一些实施方式中,t细胞疗法和/或抗血小板剂通过静脉内,肌肉内,皮下,局部,口服,透皮,腹膜内,眼内,通过植入,吸入,鞘内,心室内或鼻内给药。可以给予有效量的t细胞疗法和抗血小板剂用于预防或治疗疾病。t细胞疗法和抗血小板剂的适当剂量可根据待治疗的疾病类型,疾病的严重程度和病程,个体的临床状况,个体的临床病史和对治疗的响应,以及主治医生的自由裁量来确定。
针对离散的,固体的,可接近的肿瘤特别考虑肿瘤内注射或注射到肿瘤脉管系统中。局部,区域或全身给药也可能是适当的。对于>4cm的肿瘤,给予的体积将约为4-10ml(特别是10ml),而对于<4cm的肿瘤,将使用约1-3ml的体积(特别是3ml)。以单剂量递送的多次注射包含约0.1至约0.5ml体积。
药物组合物
当进行含有抑制性抗体的治疗组合物的临床应用时,制备适合于预期应用的药物或治疗组合物通常是有益的。在某些实施方式中,药物组合物可包含,例如,至少约0.1%的活性化合物。在其它实施方式中,所述活性化合物可包含约2%-约75%的单元重量或例如,约25%-约60%,以及从中衍生的任何范围。
本文还提供了包含t细胞疗法,抗血小板剂和药学上可接受的运载体的药物组合物和制剂。
本发明实施方式的治疗组合物有利地以可注射组合物的形式作为液体溶液或悬浮液给药;也可以制备适于在注射前溶解或悬浮在液体中的固体形式。这些制剂也可以经乳化。
活性化合物可以配制用于肠胃外给药,例如配制用于通过静脉内,肌肉内,皮下或甚至腹膜内途径注射。通常,这种组合物可以制成液体溶液或悬浮液;也可以制备适合用于在注射前加入液体制备溶液或悬浮液的固体形式;并且,制剂也可以经乳化。
适于注射使用的药物形式包括无菌水溶液或分散液;包括芝麻油,花生油或丙二醇水溶液制剂;和用于临时制备无菌可注射溶液或分散液的无菌粉末。在所有情况下,剂型都必须无菌,并应必须是易被注射的流体。它也应该在制造和储存条件下稳定,并且必须在保存过程中能够抵抗微生物如细菌和真菌的污染作用。
蛋白质组合物可以中性或盐形式配置。药学上可接受的盐包括酸加成盐(用蛋白质的游离氨基基团形成),其用无机酸(例如盐酸或磷酸)或有机酸(例如乙酸、草酸、酒石酸、扁桃酸等)形成。用游离羧基形成的盐还可源自无机碱例如钠、钾、铵、钙、或铁的氢氧化物,和有机碱例如异丙胺、三甲胺、组氨酸、普鲁卡因等。
药物组合物可包括包含水、乙醇、多元醇(例如甘油、丙二醇和液体聚乙二醇等)及其合适的混合物以及植物油的溶剂或分散介质。可通过使用包衣如卵磷脂、如果是分散体则保持所需粒度以及使用表面活性剂,来维持合适的流动性。可以通过各种抗菌剂和抗真菌剂,例如,对羟基苯甲酸酯类、氯丁醇、苯酚、山梨酸、硫柳汞等带来对微生物作用的阻碍。在很多情况中,优选包括等张剂,例如糖类或氯化钠。可通过将延迟吸收的试剂例如单硬脂酸铝和明胶用于组合物中来延长可注射组合物的吸收。
本文所述的药物组合物和制剂可以通过将具有所需纯度的活性成分(例如抗体或多肽)与一种或多种任选的药学上可接受的运载体(《雷明顿药物科学》(remington′spharmaceuticalsciences),第22版,2012)混合,以冻干制剂或水溶液形式制备。在所用剂量和浓度下,药学上可接受的运载体通常是对受者无毒的,包括但不限于:缓冲剂如磷酸盐、柠檬酸盐和其它有机酸缓冲剂;抗氧化剂,包括抗坏血酸和甲硫氨酸;防腐剂(如十八烷基二甲基苄基氯化铵;氯化六烃季铵;苯扎氯铵、苄索氯铵;苯酚、丁基或苄基醇;对羟基苯甲酸烷酯,如对羟基苯甲酸甲酯或丙酯;邻苯二酚;间苯二酚;环己醇;3-戊醇;和间甲酚);低分子量(小于约10个残基)的多肽;蛋白质,如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,如聚乙烯吡咯烷酮;氨基酸,如甘氨酸、谷胺酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖、二糖和其它糖,包括葡萄糖、甘露糖或糊精;螯合剂,如edta;糖,如蔗糖、甘露醇、海藻糖或山梨糖醇;形成盐的抗衡离子,如钠;金属络合物(如zn-蛋白质络合物);和/或非离子型表面活性剂,例如聚乙二醇(peg)。本文的示例性药学上可接受的载体还包括间隙药物分散剂,例如可溶性中性活性透明质酸酶糖蛋白(shasegp),例如人可溶性ph-20透明质酸酶糖蛋白,例如rhuph20(
抗血小板剂
本方法的实施方式涉及抗血小板剂。短语“抗血小板剂”是指抑制血小板的活化,聚集和/或粘附的任何化合物,并且旨在包括具有该活性的化合物的所有药学上可接受的盐,前药例如酯和溶剂化物形式,包括水合物,具有一个或多个手性中心的化合物可以作为外消旋体,外消旋混合物和作为单独的非对映异构体或对映异构体出现,包括所有这些异构形式及其混合物,意图包括任何结晶多晶型物,共晶体和无定形形式。
可用于本发明口服剂型的抗血小板剂的非限制性实例包括二磷酸腺苷(adp)拮抗剂或p2yi2拮抗剂,磷酸二酯酶(pde)抑制剂,腺苷再摄取抑制剂,维生素k拮抗剂,肝素,肝素类似物,直接凝血酶抑制剂,糖蛋白iib/iiia抑制剂,抗凝血酶,以及药学上可接受的盐,异构体,对映体,多晶型晶体形式,包括无定形形式,溶剂化物,水合物,共晶体,复合物,活性代谢物,活性衍生物和修饰,其前药等。
adp拮抗剂或p2y12拮抗剂阻断血小板细胞膜上的adp受体。该p2yi2受体在血小板聚集中很重要,血小板通过纤维蛋白交联。阻断该受体通过阻断糖蛋白iib/iiia途径的活化来抑制血小板聚集。在一个示例性实施方式中,抗血小板剂是adp拮抗剂或p2yi2拮抗剂。在另一个示例性实施方式中,抗血小板剂是噻吩并吡啶。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂是噻吩并吡啶。
在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂选自磺吡酮,噻氯匹定,氯吡格雷,普拉格雷,r-99224(普拉格雷的活性代谢物,由sankyo提供),r-1381727,r-125690(礼来(lilly)),c-1330-7,c-50547(千年制药公司(millenniumpharmaceuticals)),ins-48821,ins-48824,ins-446056,ins-46060,ins-49162,ins-49266,ins-50589(激励制药公司(inspirepharmaceuticals))和sch-572423(先灵葆雅(scheringplough))。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂是盐酸噻氯匹定(ticlidtm)。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂选自磺吡酮,噻氯匹定,azd6140,氯吡格雷,普拉格雷及其混合物。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂是氯吡格雷。在另一个示例性实施方式中,氯吡格雷的治疗有效量为约50mg至约100mg。在另一个示例性实施方式中,氯吡格雷的治疗有效量为约65mg至约80mg。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂选自氯吡格雷硫酸氢盐(plavixtm),氯吡格雷硫酸氢盐,氯吡格雷氢溴酸盐,氯吡格雷甲磺酸盐,坎格雷洛四钠(ar-09931mx),arl67085,ar-c66096ar-c126532和azd-6140(阿斯利康)。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂是普拉格雷。在另一个示例性实施方式中,普拉格雷的治疗有效量为约1mg至约20mg。在另一个示例性实施方式中,氯吡格雷的治疗有效量为约4mg至约11mg。在另一个示例性实施方式中,adp拮抗剂或p2yi2拮抗剂选自氯吡格雷,噻氯匹定,磺吡酮,azd6140,普拉格雷及其混合物。
在某些实施方式中,抗血小板剂是氯吡格雷或其药学上可接受的盐,溶剂化物,多晶型物,共晶体,水合物,对映体或前药。在另一个实施方式中,氯吡格雷或其药学上可接受的盐,溶剂化物,多晶型物,共晶体,水合物,对映体或前药是粉末。
pde抑制剂是阻断磷酸二酯酶(pde)的五种亚型中的一种或多种的药物,防止由相应的一种或多种pde亚型使细胞内第二信使,环磷酸腺苷(camp)和环磷酸鸟苷(cgmp)失活。在一个示例性实施方式中,抗血小板剂是pde抑制剂。在一个示例性实施方式中,抗血小板剂是选择性camppde抑制剂。在一个示例性实施方式中,pde抑制剂是西洛他唑(pletaltm)。
腺苷再摄取抑制剂阻止腺苷细胞再摄取到血小板,红细胞和内皮细胞中,导致腺苷的细胞外浓度增加。这些化合物抑制血小板聚集并引起血管舒张。在一个示例性实施方式中,抗血小板剂是腺苷再摄取抑制剂。在一个示例性实施方式中,腺苷再摄取抑制剂是双嘧达莫(persantinetm)。
给予维生素k抑制剂以阻止血栓形成(血管中的不当血液凝固)。这对于易患者的深静脉血栓形成,肺栓塞,心肌梗塞和中风的一级和二级预防是有用的。在一个示例性实施方式中,抗血小板剂是维生素k抑制剂。在一个示例性实施方式中,维生素k抑制剂选自醋硝香豆素,氯茚二酮,双香豆素(双香豆素),二苯茚二酮,二醋酸乙酯,苯丙香豆素,苯茚二酮,噻氯香豆素和华法林。
肝素是一种生物物质,通常由猪肠制成。它通过激活抗凝血酶iii来起作用,抗凝血酶iii阻止凝血酶凝结血液。在一个示例性实施方式中,抗血小板剂是肝素或肝素的前药。在一个示例性实施方式中,抗血小板剂是肝素类似物或肝素类似物的前药。在一个示例性实施方式中,肝素类似物选自抗凝血酶iii,贝米肝素,达肝素,达那肝素,依诺肝素,磺达肝素(皮下),那屈肝素,帕肝素,瑞维肝素,舒洛地希和亭扎肝素。
直接凝血酶抑制剂(dti)是一类通过直接抑制凝血酶起抗凝血剂(延迟血液凝固)作用的药物。在一个示例性实施方式中,抗血小板剂是dti。在另一个示例性实施方式中,dti是单价的。在另一个示例性实施方式中,dti是二价的。在一个示例性实施方式中,dti选自水蛭素,比伐卢定(iv),来匹卢定,地西卢定,阿加曲班(iv),达比加群,达比加群酯(口服制剂),美拉加林,希美加群(口服制剂但有肝脏并发症)及其前药。
在一个示例性实施方式中,抗血小板剂选自阿莫地平,贝前列素,卡巴匹林钙,氯吡格雷,去纤维蛋白多核苷酸,双三唑,依前列醇,吲哚布芬,伊洛前列素,皮可酰胺,利伐沙班(口服fxa抑制剂)曲前列环素,三氟醋柳酸或其前药。
在某些实施方式中,抗血小板剂是结合至少一部分garp蛋白的抗体或其片段。如本文所用,术语“抗体”旨在广泛地指任何免疫结合剂,例如igg,igm,iga,igd,ige和遗传修饰的igg以及包含保留抗原结合活性的抗体cdr结构域的多肽。抗体可以选自嵌合抗体,亲和力成熟抗体,多克隆抗体,单克隆抗体,人源化抗体,人抗体,或抗原结合抗体片段或天然或合成配体。优选地,抗garp抗体是单克隆抗体或人源化抗体。因此,通过已知方法并如本文所述,可以产生多克隆或单克隆抗体,抗体片段,以及结合结构域和cdr(包括任何前述的工程化形式),其对garp蛋白,其各自的一个或多个表位,或任何前述的偶联物具有特异性,无论这些抗原或表位是从天然来源分离或是合成衍生物或是天然化合物的变体。
适用于本发明实施方式的抗体片段的实例包括但不限于:(i)fab片段,由vl、vh、cl和ch1结构域构成;(ii)“fd”片段,由vh和ch1结构域构成;(iii)“fv”片段,由单一抗体的vl和vh结构域构成;(iv)“dab”片段,由vh结构域构成;(v)分离的cdr区域;(vi)f(ab′)2片段,包含两个连接的fab片段的二价片段;(vii)单链fv分子(“scfv”),其中vh结构域和vl结构域通过能使这两个结构域联合以形成结合结构域的肽接头连接;(viii)双特异性单链fv二聚体(参见美国专利号5,091,513)和(ix)通过基因融合构建的双抗体、多价或多特异性片段(美国专利公开号20050214860)。fv、scfv或双抗体分子可通过纳入连接vh和vl结构域的二硫桥来稳定。还可制备包含与ch3结构域相连的scfv的小体(minibody)(hu等,1996)。
其他疗法
在某些实施方式中,本发明实施方式的组合物和方法包括抗体或针对garp的抗体片段,以抑制其在癌细胞增殖中的活性,与第二种或另外的疗法联合。此类疗法可用于治疗与garp介导的细胞增殖相关的任何疾病。例如,该疾病可以是癌症。
在某些实施方式中,本发明实施方式的组合物和方法涉及t细胞疗法和抗血小板剂与至少一种其他疗法联合。该其他疗法可以是放射疗法,手术(例如,乳房肿瘤切除术和乳房切除术),化疗,基因疗法,dna疗法,病毒疗法,rna疗法,免疫疗法,骨髓移植,纳米疗法,单克隆抗体疗法或前述组合。该其他疗法可以为辅助或新辅助疗法的形式。
该方法和组合物包括提高治疗或保护效果的组合疗法,和/或增加另一抗癌或抗过度增殖疗法的治疗效果。治疗和预防方法和组合物可以有效实现所需效果的联合量提供,例如以实现癌细胞杀伤和/或细胞过度增殖的抑制。该过程可包括使细胞与抗体或抗体片段和第二疗法接触。组织、肿瘤、或细胞可以与包括一种或多种试剂(即,抗体或抗体片段或抗癌试剂)的一种或多种组合物或一种或多种药物制剂接触,或通过用两种或多种不同的组合物或制剂接触该组织、肿瘤、和/或细胞,其中一种组合物提供1)抗体或抗体片段;2)抗癌试剂,或3)抗体或抗体片段和抗癌试剂两者。此外,设想这样的组合疗法可以与化疗、放疗、外科疗法或免疫疗法联用。
本文所用术语“接触”和“暴露”用于细胞时,描述治疗构建体和化疗或放疗试剂递送给靶细胞或放置成与所述靶细胞直接并置所用的方法。为了实现细胞杀死,例如,以有效杀死所述细胞或阻止其分裂的结合量将两种试剂递送给细胞。
抑制性抗体可以在相对于抗血小板剂之前,期间,之后或以各种组合给予。该给予可以以间隔方式进行,其范围从同时到数分钟到数天到数周。在抗体或抗体片段与抗癌剂是分开给予患者的实施方式中,一般会确保每次递送的期间不会显著超时,从而使得两种化合物仍然能够对患者发挥有利的联合作用。在这类情况中,可以考虑将抗体疗法和抗癌疗法提供给患者,互相间隔在约12到24或72小时内,更优选地,互相间隔在约6到12小时内。在一些情况中,可能需要显著延长治疗时间,其中各给药之间间隔数天(2、3、4、5、6或7)到数周(1、2、3、4、5、6、7或8)。
在某些实施方式中,一个治疗疗程将会持续1到90天,或更多(该范围包括间隔天数)。考虑将一种试剂在第一天到第九十天(该范围包括间隔天数)中的任意一天或它们的任意组合给予患者,并且将另一种试剂在第一天到第九十天(该范围包括间隔天数)中的任意一天或它们的任意组合给予患者。在一天(24小时)内,可以一次或多次给予该患者该试剂。此外,一个治疗疗程后,预计将会有一段不进行抗癌治疗的时间。这一段时间可能持续1到7天,和/或1到5周,和/或1到12个月或更久(该范围包括间隔天数),这取决于患者的状况,例如其预后、力量、健康状况等。预期在需要时重复该治疗循环。
在一些实施方式中,其他疗法是给予小分子酶抑制剂或抗转移剂。在一些实施方式中,其他疗法是给予副作用限制剂(例如,旨在减轻治疗副作用的发生和/或严重性的药剂,例如抗恶心剂等)。在一些实施方式中,其他疗法是放射疗法。在一些实施方式中,其他疗法是手术。在一些实施方式中,其他疗法是放射疗法和手术的组合。在一些实施方式中,其他疗法是γ辐射。在一些实施方式中,其他疗法是靶向pi3k/akt/mtor途径的疗法,hsp90抑制剂,微管蛋白抑制剂,凋亡抑制剂和/或化学预防剂。其他疗法可以是本领域已知的一种或多种化疗剂。
可以采用各种组合。对于下面的例子,抗体疗法,或t细胞疗法和抗血小板剂是“a”,抗癌疗法是“b”:
a/b/ab/a/bb/b/aa/a/ba/b/bb/a/aa/b/b/bb/a/b/b
b/b/b/ab/b/a/ba/a/b/ba/b/a/ba/b/b/ab/b/a/a
b/a/b/ab/a/a/ba/a/a/bb/a/a/aa/b/a/aa/a/b/a
将该实施方式的任何化合物或疗法给予患者需要遵循给予该化合物的一般方案,(必要时)考虑试剂的毒性。因此,在一些实施方式中,存在监测因组合疗法产生的毒性的步骤。
化疗
根据本实施方式可以使用各种化疗剂。术语“化疗”指使用药物治疗癌症。“化疗剂”指代在癌症治疗中给予的化合物或组合物。这些试剂或药物根据其在细胞内的活性模式分类,例如,其是否受影响细胞周期和在什么阶段影响细胞周期。或者,可以根据试剂直接交联dna的能力、插入dna的能力、或通过其影响核酸合成诱导染色体和有丝分裂的畸变的能力进行表征。
化疗剂的实例包括烷化剂,例如噻替哌和环磷酰胺;烷基磺酸盐如白消安、英丙舒凡和哌泊舒凡;吖丙啶如苄替哌、卡波醌、美妥替哌、和乌瑞替哌;乙烯亚胺和甲基三聚氰胺包括六甲蜜胺、三乙撑蜜胺、三亚乙基磷酰胺、三亚乙基硫代磷酰胺和三甲基邻三聚氰胺(trimethylomelamine);乙酰基蛋白(特别是膨松素(bullatacin)和布鲁啉酮(bullatacinone));喜树碱(包括拓扑替康和伊立替康);苔藓抑素;卡利他汀;cc-1065(包括其阿多来新、卡折来新和比折来新合成类似物);念珠藻素(特别是自念珠藻环肽1和自念珠藻环肽8);多拉司他汀(dolastatin);多卡霉素(duocarmycin)(包含合成类似物、kw-2189和cbi-tmi);艾榴塞洛素;氮芥,例如苯丁酸氮芥、萘氮芥、氯磷酰胺、雌莫司汀、异环磷酰胺、双氯乙基甲胺、盐酸氧氮芥、美法仑、新恩比兴(novembichin)、苯芥胆甾醇、泼尼氮芥(prednimustine)、曲磷胺、和乌拉莫司汀;亚硝基脲,例如卡莫司汀、吡葡亚硝脲、福莫司汀(foremustine)、洛莫司丁、尼莫司汀、雷莫司汀;抗生素,例如烯二炔抗生素(例如,卡奇霉素、特别是卡奇霉素γ1i和卡奇霉素ωi1;达内霉素(dynemicin),包含达内霉素(dynemicina);二膦酸盐,例如氯屈膦酸(clodronate);埃斯培拉霉素(esperamicin);以及新抑癌蛋白(neocarzinostatin)发色团和相关的色蛋白烯二炔抗生素发色团、阿柔比星(aclacinomysins)、放线菌素、安曲霉素(authrarnycin)、偶氮丝氨酸、博来霉素、放线菌素c、卡拉比星(carabicin)、洋红霉素、嗜癌菌素、色霉素、放线菌素d、柔红霉素、地托比星、6-重氮基-5-氧代-l-正亮氨酸、多柔比星(包括吗啉代-多柔比星、氰基吗啉代-多柔比星、2-吡咯啉-多柔比星和脱氧多柔比星)、表柔比星、依索比星、伊达比星、马赛罗霉素、丝裂霉素例如丝裂霉素c、霉酚酸、诺拉霉素、橄榄霉素、培洛霉素、甲基丝裂霉素(potfiromycin)、嘌呤霉素、三铁阿霉素、罗多比星、链黑霉素、链佐星、块菌素、乌苯美司、净司他丁、佐柔比星;抗代谢剂,例如氨甲喋呤和5-氟尿嘧啶(5-fu);叶酸类似物,例如二甲叶酸、氨蝶并蝶呤、三甲曲沙;嘌呤类似物,例如氟达拉滨、6-巯基嘌呤、硫咪嘌呤、硫鸟嘌呤;嘧啶类似物,例如安西他宾、阿扎胞苷、6-氮杂尿苷、卡莫氟、阿糖胞苷、二脱氧尿苷、去氧氟尿苷、依诺他宾、氮尿苷;雄激素,例如卡鲁睾酮、丙酸屈他雄酮、环硫雄醇、美雄烷、睾内酪;抗肾上腺,例如米托坦、曲洛司坦;叶酸补充剂,例如亚叶酸(frolinicacid);醋葡醛内酯;醛磷酰胺糖苷;氨基乙酰丙酸;二氢嘧啶脱氢酶灭活剂;安吖啶;阿莫司汀(bestrabucil);比生群;乙茎去氮氨蝶呤(edatraxate);defofamine;秋水仙胺;亚丝醌;依氟鸟氨酸碱盐酸盐(elfornithine);依利醋铵;大环内酯;依托格鲁;硝酸镓;羟基脲;香菇菌多糖;氯尼达明;类美坦西醇,例如美登素和安丝菌素;米托胍腙;米托蒽醌;莫哌达醇;二胺硝吖啶;喷司他丁;蛋氨氮芥;吡柔比星;洛索蒽醌;鬼臼酸;2-乙基酰肼;丙卡巴肼;psk多糖复合物;雷佐生;根霉素;西佐喃;锗螺胺;细交链孢菌酮酸;三亚胺醌;2,2′,2″-三氯三乙胺;单端孢菌毒素(特别是t-2毒素、疣孢漆斑霉素a、露湿漆斑霉素a和蛇形菌素(anguidine));氨基钾酸酯;长春地辛;达卡巴嗪;盐酸甘露醇氮芥;二溴甘露醇;二溴卫矛醇;胍血生;卡胞核嘧啶(gacytosine);阿糖胞苷(“ara-c”);环磷酰胺;紫杉烷,例如紫杉醇和多西他赛吉西他滨;6-硫鸟嘌呤;巯基嘌呤;铂配合物,例如顺铂、奥沙利铂和卡铂;长春碱;铂;依托泊甙(vp-16);异环磷酰胺;米托蒽醌;长春新碱(vancristine);长春瑞滨;诺肖林;替尼泊苷;依达曲沙;道诺霉素;氨基蝶呤;希罗达(xeoloda);伊班膦酸钠;伊立替康(例如,cpt-11);拓扑异构酶抑制剂rfs2000;二氟甲基鸟氨酸(dmfo);视黄醇,例如视黄酸;卡培他滨(capecitabine);卡铂,丙卡嗪,上古霉素,吉西比宁,纳维伦,法奈尔蛋白,蛋白质坦斯费酶抑制剂,转氨酶抑制剂和上述任何一个的药学可接受盐、酸或衍生物。
放疗
造成dna损伤并已广泛使用的其他因素包括通常称为γ射线、x射线和/或将放射性同位素直接递送给肿瘤细胞。也考虑其它形式的dna破坏因素,如微波质子束肤辐射(美国专利5,760,395和4,870,287)和紫外辐照。所有这些因素很有可能会对dna、dna前体、dna复制和修复、染色体装配和维持造成较宽范围的损伤。x-射线的剂量范围从延长期间(3至4周)50至200伦琴的每天剂量到2000至6000伦琴的单次剂量。放射性同位素的剂量范围差异很大,取决于所述同位素的半衰期、发射辐射的强度和类型、以及肿瘤细胞的摄入。
免疫疗法
技术人员将理解,其他免疫疗法可以与实施方式的方法组合使用或与其结合使用。在治疗癌症的情况中,免疫疗法一般依赖于使用免疫效应细胞和分子靶向并破坏癌细胞。利妥昔单抗
抗体-药物偶联物已成为癌症治疗学发展的突破性方法。癌症是世界上死亡的主要原因之一。抗体-药物偶联物(adc)包含与细胞杀伤药物共价连接的单克隆抗体(mab)。这种方法将mab对其抗原靶标的高特异性与高效细胞毒性药物相结合,从而产生“武装”mab,将有效载荷(药物)递送至富含抗原水平的肿瘤细胞(carter等,2008;teicher2014;leal等,2014)。靶向递送药物还使其在正常组织中的暴露最小化,导致毒性降低和治疗指数改善。fda批准的两种adc药物,2011年批准的
在免疫疗法的一方面中,肿瘤细胞必须携带一些易于靶向的标志物,即,不存在于大多数其它细胞上。存在许多肿瘤标志物并且这些标记物中的任一种适用于在本实施方式的内容中靶向。常见的肿瘤标志物包括cd20,癌胚抗原,酪氨酸酶(p97),gp68,tag-72,hmfg,唾液酸lewis抗原,muca,mucb,plap,层粘连蛋白受体,erbb和p155。免疫疗法的另一方面是将抗癌作用与免疫刺激作用结合。免疫刺激分子同样存在包括例如细胞因子,例如il-2,il-4,il-12,gm-csf,γ-ifn,趋化因子,例如mip-1,mcp-1,il-8,和生长因子,例如flt3配体。
现今在研或使用的免疫疗法的示例是:免疫佐剂,例如牛分枝杆菌(mycobacteriumbovis),镰状疟原虫(plasmodiumfalciparum),二硝基氯苯和芬香族化合物(美国专利5,801,005和5,739,169;hui和hashimoto,1998;christodoulides等,1998),细胞因子疗法,例如干扰素,和,il-1,gm-csf和tnf(bukowski等,1998;davidson等,1998;hellstrand等,1998);基因疗法,例如tnf,il-1,il-2,p53(qin等,1998;austin-ward和villaseca,1998;美国专利5,830,880和5,846,945)和单克隆抗体,例如抗-cd20,抗-神经节苷脂gm2,和抗-p85(hanibuchi等,1998;美国专利5,824,311)。考虑一种或多种抗癌症疗法可以用于本文所述的抗体疗法。
在一些实施方式中,免疫疗法可以是免疫检查点抑制剂。免疫检查点是免疫系统中的分子,其发出信号(例如,共刺激分子)或拒绝信号。可能被免疫检查点的阻断靶向的抑制性检查点分子包括腺苷a2a受体(a2ar),b7-h3(也称为cd276),b和t淋巴细胞衰减剂(btla),细胞毒性t淋巴细胞相关蛋白4(ctla-4,也称为cd152),吲哚胺2,3-双加氧酶(ido),杀伤细胞免疫球蛋白(kir),淋巴细胞活化基因-3(lag3),程序性死亡1(pd-1),t细胞免疫球蛋白结构域和粘蛋白结构域3(tim-3)和t细胞活化的v结构域ig抑制剂(vista)。特别地,免疫检查点抑制剂靶向pd-1轴(axis)和/或ctla-4。
免疫检查点抑制剂可以是药物,例如小分子,重组形式的配体或受体,或特别是抗体,例如人抗体(例如,国际专利公开wo2015016718;pardoll,natrevcancer,12(4):252-64,2012;两者均通过引用纳入本文)。可以使用已知的免疫检查点蛋白质抑制剂或其类似物,特别是可以使用嵌合,人源化或人形式的抗体。如技术人员所知,替代和/或等同名称可用于本公开中提及的某些抗体。这些替代和/或等同名称在本发明的上下文中是可互换的。例如,已知兰姆珠单抗还有替代和等同名称mk-3475和彭美罗珠单抗。
在一些实施方式中,pd-1结合拮抗剂是抑制pd-1与其配体结合伴侣结合的分子。在一个具体方面,pd-1配体结合伴侣是pdl1和/或pdl2。在另一个实施方式中,pdl1结合拮抗剂是抑制pdl1与其结合伴侣结合的分子。在一个具体方面,pdl1结合伴侣是pd-1和/或b7-1。在另一个实施方式中,pdl2结合拮抗剂是抑制pdl2与其结合伴侣结合的分子。在一个具体方面,pdl2结合伴侣是pd-1。拮抗剂可以是抗体,其抗原结合片段,免疫粘附素,融合蛋白或寡肽。示例性抗体描述于美国专利号us8735553,us8354509和us8008449中,所有这些专利均通过引用纳入本文。用于本文提供的方法的其他pd-1轴拮抗剂是本领域已知的,例如美国专利申请号us20140294898,us2014022021和us20110008369中所述,所有这些文献均通过引用纳入本文。
在一些实施方式中,pd-1结合拮抗剂是抗pd-1抗体(例如,人抗体,人源化抗体或嵌合抗体)。在一些实施方式中,抗pd-1抗体选自尼莫单抗,彭美罗珠单抗和ct-011。在一些实施方式中,pd-1结合拮抗剂是免疫粘附素(例如,包含融合至恒定区(例如,免疫球蛋白序列的fc区)的pdl1或pdl2的细胞外或pd-1结合部分的免疫粘附素。在一些实施方式中,pd-1结合拮抗剂是amp-224。尼莫单抗,也称为mdx-1106-04,mdx-1106,ono-4538,bms-936558和
可以在本文提供的方法中靶向的另一免疫检查点是细胞毒性t淋巴细胞相关蛋白4(ctla-4),也称为cd152。人ctla-4的完整cdna序列具有genbank登录号l15006。ctla-4存在于t细胞表面,并且当与抗原呈递细胞表面上的cd80或cd86结合时充当“关闭”开关。ctla4是免疫球蛋白超家族的成员,其在辅助t细胞的表面上表达并向t细胞传递抑制信号。ctla4类似于t细胞共刺激蛋白cd28,并且两种分子分别与抗原呈递细胞上的cd80和cd86(也称为b7-1和b7-2)结合。ctla4向t细胞传递抑制信号,而cd28传递刺激信号。细胞内ctla4也存在于调节性t细胞中,并且可能对其功能很重要。通过t细胞受体和cd28的t细胞活化导致ctla-4的表达增加,ctla-4是b7分子的抑制性受体。
在一些实施方式中,免疫检查点抑制剂是抗-ctla-4抗体(例如,人抗体,人源化抗体或嵌合抗体),其抗原结合片段,免疫粘附素,融合蛋白或寡肽。
可以使用本领域熟知的方法产生适用于本发明方法的抗人ctla-4抗体(或由其衍生的vh和/或vl结构域)。或者,可以使用本领域公认的抗ctla-4抗体。例如,公开于us8,119,129,wo01/14424,wo98/42752;wo00/37504(cp675,206,也称为特姆单抗;以前的替西木单抗),美国专利号6,207,156;hurwitz等,(1998)procnatlacadsciusa95(17):10067-10071;camacho等,(2004)jclinoncology22(145):摘要号2505(抗体cp-675206);和mokyr等,(1998)cancerres58:5301-5304的抗-ctla-4抗体可用于本文公开的方法中。上述参考文献的各自的启示都通过引用纳入本文。也可以使用与任何这些本领域公认的抗体竞争结合ctla-4的抗体。例如,人源化ctla-4抗体描述于国际专利申请号wo2001014424,wo2000037504和美国专利号us8017114中;全部通过引用纳入本文。
示例性的抗-ctla-4抗体是伊匹单抗(也称为10d1,mdx-010,mdx-101和
用于调节ctla-4的其他分子包括ctla-4配体和受体,如美国专利号us5844905,us5885796和国际专利申请号wo1995001994和wo1998042752中所述;所有这些都通过引用纳入本文,和免疫粘附,如美国专利号us8329867中所述,该专利通过引用纳入本文。
手术
约60%的癌症患者会经历一些类型的外科手术,包括预防性、诊断性或分期、治愈性和缓解性外科手术。治愈性手术包括切除,其中全部或部分癌组织被物理移除,切除和/或破坏,并且可以与其他疗法结合使用,例如本实施方式的治疗,化疗,放射疗法,激素疗法,基因疗法,免疫疗法和/或替代疗法。肿瘤切除是指物理去除至少部分肿瘤。除了肿瘤切除术,外科手术治疗包括激光手术、冷冻手术、电外科手术和显微镜控制手术(莫氏(mohs)手术)。
所有癌细胞、组织或肿瘤的部分或全部切割后,可能会在体内形成空腔。治疗可通过灌注、直接注射或局部区域应用其他抗癌治疗来实现。这样的治疗可以例如每1、2、3、4、5、6或7天或每1、2、3、4和5周或每1、2、3、4、5、6、7、8、9、10、11或12个月重复。这些治疗也可用不同的剂量。
其他物质
考虑到其它物质可与本实施方式某些方面联用以提高治疗的治疗效率。这些其他物质包括影响细胞表面受体和gap连接上调的物质、细胞抑制和分化物质、细胞粘附抑制剂、增加所述过度增殖性细胞对凋亡诱导物的敏感性的物质、或其它生物试剂。通过提高gap接头的数量将提高对相邻过度增殖细胞群的抗-过度增殖作用。在其他实施方式中,细胞抑制或分化试剂可与本实施方式的某些方面联用以改进治疗的抗过度增殖效力。考虑细胞粘附抑制剂来改善本实施方式的效力。细胞粘附抑制剂的示例为粘着斑激酶(fak)抑制剂和洛伐他汀。还可以预料到增加过度增殖细胞对凋亡的敏感性的其它药剂,如抗体c225可与本实施方式的某些方面联用以提高治疗效力。
制品或试剂盒
在实施方式的各个方面,设想了一种试剂盒,其含有治疗剂和/或其他治疗剂和递送剂。在一些实施方式中,本发明实施方式涉及用于制备和/或给予实施方式的疗法的试剂盒。试剂盒可包含一个或多个密封的小瓶,其含有本发明实施方式的任何药物组合物。试剂盒可包括,例如,至少一种garp抗体以及制备,配制和/或给予实施方式的组分或进行本发明方法的一个或多个步骤的试剂。在一些实施方式中,试剂盒还可包含合适的容器,其是不与试剂盒的组分反应的容器,例如ep管,测定板,注射器,瓶或管。容器可以由可消毒的材料制成,例如塑料或玻璃。
在一些实施方式中,本文还提供了包含过继性t细胞和抗血小板剂(例如,抗garp抗体)的制品或试剂盒。所述制品或试剂盒可进一步包含包装说明书,所述包装说明书包含将过继性t细胞与抗血小板剂联合使用以治疗或延迟个体癌症进展或增强患有癌症的个体的免疫功能的说明书。本文所述的任何过继性t细胞和/或抗血小板剂可包括在制品或试剂盒中。在一些实施方式中,过继性t细胞和抗血小板剂在相同的容器或分开的容器中。合适的容器包括例如,大瓶、小瓶、袋和注射器。容器可以由多种材料形成,例如玻璃,塑料(例如聚氯乙烯或聚烯烃),或金属合金(例如不锈钢或哈氏合金)。在一些实施方式中,该容器装有制剂且贴有或附有标签,标签上可记载使用指南。制品或试剂盒还可包括从商业和使用者立场来看可能需要的其他材料,包括其他缓冲剂、稀释剂、填充剂、针头、注射器和具有使用说明书的插页。在一些实施方式中,制品还包括一种或多种其他物质(例如化疗剂和抗肿瘤剂)。一种或多种物质的合适容器包括例如,大瓶、小瓶、袋和注射器。
试剂盒可以进一步包括说明书,其概述了本文所述方法的程序步骤,并且将遵循与本文所述或本领域普通技术人员已知的基本相同的程序。指令信息可以在包含机器可读指令的计算机可读介质中,当使用计算机执行时,所述机器可读指令导致显示递送药学有效量的治疗剂的真实或虚拟程序。
iv.实施例
包括以下实施例以说明本发明的优选实施方式。本领域的普通技术人员应理解,根据代表本发明人所发现技术的实施例中公开的技术能很好地实施本发明,从而可将其视为实施本发明的优选模式。但是,本领域技术人员根据本公开应理解,在不偏离本发明精神和范围的前提下,可对所公开的具体实施方式进行许多变化,同时仍能获得相同或类似的结果。
实施例1-癌细胞中garp的表达
最近的研究,包括癌症基因组图谱(tcga)项目,已经表明lrrc32在多达30%的具有许多人类癌症类型的患者中被扩增,包括卵巢癌,肺癌,乳腺癌和头颈癌(图1a)。为了检查garp蛋白表达,对来自存档的人肿瘤的人肿瘤微阵列进行免疫组织化学(ihc),随后确定garp表达是否具有任何预后意义。通过对用人garp稳定转染的前b白血病细胞系的染色确定抗人garp抗体的特异性(图1b)。鉴于lrrc32在人乳腺癌中被扩增(szepetowski等,1992),首先在使用ihc在乳腺癌患者中评估garp表达。结果由临床病理学家以双盲方式读取和评分。患者匹配的未受累乳腺组织与原发性乳腺癌(n=16)的ihc分析表明,16名患者中的9名患者的癌组织中garp表达显著增加(图1c)。通过rt-pcr,28.5%的乳腺癌患者(n=42)与正常乳腺组织相比,garpmrna表达增加≥2倍。然后对癌症样本进行ihc,包括55个结肠癌样本,55个相邻的正常组织和11个相应的淋巴结以及相邻的正常组织(图1d)。正常上皮样本显示没有明显的garp阳性(图1d和1e)。然而,原发性癌症(结肠和肺)和淋巴结(ln)转移组织对garp有不同的染色(对同种型对照抗体均为阴性)(图1e)。与正常组织中不可检测的水平(定义为0)相比,原发癌中garp阳性细胞的百分比为26.1%(p=8.6×10-9),ln转移中为25%(p=0.008)。在0至4的等级上,garp强度评分在0和3之间,在原发性结肠癌中平均为0.78(p=1.1×10-8),在ln转移中平均为1.18(p=0.003)(图1e)。类似地,在肺和前列腺的原发性癌症中发现显著增加的garp水平(图1e)。更重要的是,无论肿瘤的病理分级或疾病的淋巴结状态如何,garp水平与结肠癌和肺癌患者的总体存活率成反比(图1f)。高garp表达也与前列腺癌中的高gleason评分相关(p=0.035)(图1f)。这些结果首次表明garp在人类癌症中广泛表达,并且表达水平与疾病侵袭性相关。
体外生物化学研究已经确定,garp也以可溶形式存在,其与来自treg细胞的潜伏tgf-β1以复合物形式分泌(gauthy等,2013)。还已经表明,garp依赖于内质网中的分子伴侣grp94用于折叠和细胞表面表达(zhang等,2015)。为了确定garp分泌是否是treg细胞特异性事件或garp内在现象,在具有和不具有grp94的小鼠前b细胞中表达n末端血凝素(ha)标记的garp,然后在细胞裂解物和条件培养基中分析garp表达。仅在gp96+(wt)细胞中观察到三个garp条带:分子量分别为约75,45和30kda(图2a)。45和30kda片段似乎是全长细胞表面garp(75kda)的翻译后切割产物,因为它们与pngasef相比对endoh更具抗性,并且在wt中发现,但在grp94ko细胞中未发现。如果是这样,30kda的n-末端garp片段应该从细胞表面释放到培养基中。实际上,通过质谱法对培养基中的30kda蛋白质进行凝胶提取和测序证实它来自garp的n-末端片段(图2b)。
接下来确定在癌症患者的血清中是否存在可溶性garp(sgarp)以及sgarp的血清水平是否具有任何预后意义。从雄性正常对照(n=7)和前列腺癌患者(n=48)收集血清,并通过elisa分析garp。发现sgarp存在于来自正常个体和前列腺癌患者的血清中(图2c)。进一步分析显示,较高的garp水平与增加的前列腺癌特异性抗原(psa)水平和转移相关(图2d)。此外,使用garp-tgf-β1夹心elisa在前列腺癌患者和正常对照的血清中评估sgarp-tgf-β1复合物的存在。如所预测的,癌症患者的血清比正常对象含有更高水平的可溶性garp和tgf-β1复合物(图2e)。为了深入了解可溶性garp的功能,制备了融合蛋白,其由与igg的fc结构域(garp-fc)连接的garp的n-末端细胞外结构域组成。该构建体在中国仓鼠卵巢(cho)细胞中表达。然后从条件培养基中纯化garp融合蛋白。如通过活性tgf-βelisa测量的,在garp-fc和活性tgf-β1之间发现直接关联(图2f),表明存在garp-fc-tgf-β1复合物。该发现与先前报道的sgarp的免疫抑制性质一致(hahn等,2013)。
实施例2-garp和tgf-β
在正常鼠乳腺上皮细胞中强化的garp表达上调tgf-β表达并驱动肿瘤发生。在正常鼠乳腺上皮细胞(nmumg)细胞中,tgf-β发挥生长抑制响应和上皮-间充质细胞转化(emt)响应(xie等,2003)。因此,nmumg细胞已被广泛用于研究tgf-β信号传导和生物学(xu等,2009)。鉴于garp调节tgf-β的生物利用度,nmumg细胞用于生物测定以研究膜结合的garp和可溶性garp对上皮细胞的影响。发现稳定的garp表达诱导smad-2/3磷酸化和波形蛋白的表达,但下调e-钙粘蛋白,与经典的tgf-β信号传导增加一致(图3a)。此外,用可溶性garp-fc刺激的nmumg细胞在24小时内从其典型的多边形和扁平上皮细胞形态变为纺锤形形态(图3b),伴随波形蛋白的时间和剂量依赖性上调(图3c和3d)。如所预期的,与用空载体(ev)转导的细胞相比,稳定表达garp或garp-fc的nmumg细胞具有更高的活性tgf-β1表达(图3e)以及可溶性garp(图3f)。进行体外“划痕”测定以测量表达garp的细胞的迁移性质。通过表达garp的细胞,间隙的闭合率(通过刮擦培养板产生)显著增加,表明获得的迁移能力增加(图3g和3h)。还检查了强制性garp表达是否使nmumg细胞在体内建立肿瘤。为此,将表达garp的nmumg细胞或ev对照细胞注射到雌性免疫缺陷nod-ragl-/-小鼠的第四乳腺脂肪垫中,其中,所有这些细胞也被工程化以共表达荧光素酶。通过生物发光的体内成像,发现生物活性物质仅在接受garp+或garp-fc+nmumg的小鼠中形成,但在接受ev转导细胞的小鼠中不形成(图3i)。通过组织学证实了表达garp的细胞的肿瘤形成(图3j)。总之,这些结果表明garp通过上调tgf-β具有转化特性,将garp鉴定为潜在的新型致癌基因。
沉默garp延迟肿瘤生长。正常小鼠乳腺上皮细胞系(nmumg*)的一种变体,其中rna结合蛋白hnrnpe1被rna干扰敲减,最近被描述为能够在裸鼠中形成肿瘤(howley等,2015)。有趣的是,发现这些细胞表达了显著水平的内源性garp(图4a-4c),提高了tgf-β生物合成的可能性,除了沉默tgf-β介导的翻译抑制复合物,在该模型中驱动乳腺癌。为了测试该假设,在nmumg*细胞中进行garp的短发夹rna(shrna)敲减(kd)(图4a-4c)。如通过mtt测定所确定的,garp沉默不影响nmumg*细胞的体外增殖(图4d)。值得注意的是,nmumg*细胞中单独的garp沉默显著减弱了它们在体内的生长(图4e)。此外,这些garpkd细胞转移至肺和肝的能力受到损害(图4f和4g)。
鼠乳腺癌细胞中的garp上调促进tgf-β活化,肿瘤生长,转移和免疫耐受。lrrc32最初在乳腺癌中被描述为经常被扩增的基因(ollendorff等,1994),并且已显示tgf-β信号传导促进乳腺癌侵袭和转移(massague,2008;padua等,2008;siegel等,2003)。然而,癌症中tgf-β生物学研究不足的方面是tgf-β通过调节宿主免疫应答的癌症-外在作用(li和flavell,2008)。因此,在balb/c小鼠的高侵袭性和转移性4t1乳腺癌模型中检查了garp对同基因免疫充足环境中癌症生长和转移的影响(pulaski和ostrand-rosenberg,2001)。与nmumg系统类似,4p1细胞中garp或garp-fc的过表达导致活性tgf-β的产生增加(图5a和5b)。tgf-β抑制肿瘤特异性免疫的关键机制之一是通过foxp3+treg的诱导。为此,在多克隆t细胞活化剂存在3天下,用来自4t1-garp,4t1-garp-fc和空载体(ev)对照细胞的条件培养基体外培养纯化的原初cd4+t细胞。与来自对照细胞的培养基相比,来自表达garp的细胞的条件培养基在诱导treg分化上的效率高2-3倍(图5c)。将4t1-ev,4t1-garp和4t1-garp-fc细胞原位注射到6-8周龄雌性balb/c小鼠的第四右乳腺脂肪垫中。发现表达garp的细胞更具攻击性,如原发性肿瘤的生长动力学增加(图5d和5e)和肺转移增加(图5f)所示。还发现这种侵袭性与肿瘤微环境中增强的tgf-β信号传导相关,如癌细胞中增加的p-smad-2/3(图5g和5h),以及致耐受性treg细胞的扩增(图5i和5j)所确定。
实施例3-黑素瘤研究
4t1肿瘤模型的研究提出了garp是否对肿瘤特异性t细胞的功能发挥抑制作用的问题。为了解决这种可能性,使用具有确定抗原特异性的b16黑素瘤模型以及cd8+t细胞受体(tcr)转基因小鼠(pmel),其具有对黑素瘤相关抗原gp100特异的t细胞(muranski等,2008;overwijk等,2003)。用或不用garp-fc制备b16-f1细胞,然后皮下注射到c57bl/6小鼠中。然后在离体活化的pmel细胞的过继细胞转移(act)之前用环磷酰胺(cy)对荷瘤小鼠进行淋巴细胞清除(rubinstein等,2015)(图6a)。发现b16细胞表达garp-fc导致对act的抗性增加(图6b和6c),这与受体小鼠中抗原特异性pmel细胞数量减少有关,特别是在肿瘤生长的前四周,此时肿瘤表面积小于100mm2(图6d和6e)。类似地,pmelcd8+t细胞在响应抗原刺激时产生ifnγ的能力在携带garp-fc+b16黑素瘤的小鼠中也受损(图6f和6g)。
实施例4-garp作为癌症中的新治疗靶标
本文描述的研究已经证明garp在多种人类癌症中异常表达,并且鼠肿瘤中的garp表达与tgf-β生物利用度,癌症侵袭性和t细胞耐受性的增加相关。接下来使用基于抗体的策略确定garp是否可以作为癌症中的新治疗靶标。为了产生抗garp单克隆抗体(mab),用重组人garp免疫小鼠,然后用稳定表达人garp的经照射的全骨髓瘤sp2/0细胞进行加强,目的是产生针对garp的单克隆抗体,其是构象特异性的。如通过流式细胞术测定的,产生超过20种特异性识别人garp的mab(图7a)。所有这些克隆都对人garp具有特异性,除了一个克隆4d3以外,其对小鼠garp具有低水平的交叉反应性。4d3是唯一能够阻断外源人ltgfβ-1(hultgfβ-1)与表面garp结合的克隆(图7b)。为了检查garp抗体是否在体内具有任何直接的抗肿瘤活性,用表达人garp的4t1肿瘤细胞原位接种balb/c小鼠。然后用4d3(igg1)或第二抗garpmab5c5(igg2a)或用同种型对照抗体(iso)处理小鼠,有或没有单剂量的cy。选择该方案是因为先前已显示tgf-β中和抗体1d11能够增强cy控制4t1的能力(chen等,2014)。发现4d3和5c5处理均适度抑制原发性肿瘤生长(图7c)。然而,更重要的是,与同种型处理组相比,抗-garpmab处理组中向肺的肿瘤转移显著降低,而不管是否伴随化疗处理(图7d)。值得注意的是,用阻断ltgf-β与garp结合的4d3处理也与脾treg和gr1+cd11b+髓源抑制细胞(mdsc)的显著减少有关(图7e)。
血小板不仅在细胞内产生和储存高水平的tgfβ,而且是迄今为止已知的组成型表达tgfβ的细胞表面对接受体garp的唯一细胞实体。因此,血小板可通过活性分泌以及garp介导的从其他细胞或细胞外基质的捕获而促成tgfβ的全身水平。研究了血小板在多大程度上以及如何对生理性tgfβ库起作用。从野生型(wt)小鼠获得基线血清,然后给予血小板消除抗体。将这些小鼠依次放血并通过elisa定量血清tgfβ。血小板的消耗导致活性和总tgfβ的完全丧失,一旦血小板计数恢复,其有效地反弹(图8a)。这些实验证明血小板对循环tgfβ水平起主要作用。
通过关注血小板garp在活性tgfβ产生中的作用,实验性地解决了癌症免疫中血小板衍生tgfβ的生物学。除血小板特异性hsp90b1ko小鼠外,还产生了两个额外的小鼠模型:一个选择性缺失血小板中的garp(pf4-cre-lrrc32flox/flox,或plt-garpko),另一个小鼠具有血小板限制性敲除tgfβ1(pf4-cre-tgfblflox/flox或plt-tgfβ1ko)。由于gp96也是garp的专性伴侣,因此来自plt-gp96ko小鼠和plt-garpko小鼠的血小板均不表达细胞表面garp-tgfβ复合物。然而,与wt血小板相比,来自plt-tgfβ1ko小鼠的血小板表达相似水平的表面garp-tgfβ1复合物(图8b-8d),表明可以在没有自分泌tgfβ1的情况下形成garp-tgfβ1复合物。
然后在wt和敲除小鼠的血浆和血清中测量活性和潜伏tgfβ的水平(图8e,f)。在wt小鼠中,与血浆相比,活性tgfβ在血清中升高,表明血小板和/或凝血级联在tgfβ活化中的作用(图8e)。重要的是,plt-gp96ko和plt-garpko小鼠的血清中具有非常少的活性tgfβ,证实了血小板固有的garp在将潜伏tgfβ转化为活性形式中的重要性。相反,plt-tgfβ1ko小鼠中活性tgfβ的血清水平与wt小鼠的血清水平相当(图8e),表明血小板能够以反式方式激活来自非血小板来源的tgfβ。值得注意的是,血清中的总潜伏tgfβ水平仅在plt-tgfβ1ko小鼠中降低,而在plt-gp96ko或plt-garpko小鼠中不降低(图8f)。总之,这些数据表明血小板固有的garp是tgfβ全身激活中最重要的机制。该实验还明确地证实,活性tgfβ的血清而非血浆水平仅反映血小板活化。
假设血小板特异性garp在抗肿瘤t细胞免疫中起着关键的负面作用。通过比较wt,plt-tgfβ1ko和plt-garpko受体小鼠中黑素瘤的过继性t细胞疗法的功效来解决该假设(图9)。在wt或ko小鼠中建立b16-fl黑素瘤,然后在第9天用环磷酰胺(cy)淋巴细胞清除,并在第10天输注离体活化的pmelt细胞(图9a)。与wt小鼠相比,在plt-garpko小鼠中更高效地控制肿瘤(图9a)。这与plt-garpko小鼠的外周血中pmel细胞增强的持久性(图9b)和功能性相关(图9c)。形成鲜明对比的是,plt-tgfβ1ko小鼠,其血小板表达garp并且仍然能够激活tgfβ,对肿瘤没有改善的控制(图9d)。接下来在mc38结肠癌系统中研究了这些发现的一般性,因为这种可移植肿瘤在同系小鼠中的生长经历了cd4和cd8介导的免疫压力。与wt小鼠相比,plt-garpko小鼠中mc38的生长显著减少(图10a-10c)。带有mc38的plt-garpko小鼠具有降低的活性tgfβ的血清水平(10d)。更重要的是,mc38肿瘤切片中p-smad2/3(p-smad2/3)的染色显示plt-garpko小鼠中mc38细胞中tgfβ信号传导的显著减弱(图10e和10f)。这与plt-garpko小鼠中全身性骨髓衍生的抑制细胞(图10g)和肿瘤浸润性调节性t细胞的减少有关(图10h)。总之,这表明血小板是肿瘤微环境中tgfβ活性的主要来源,并且它们通过garp-tgfβ对抗肿瘤免疫发挥有效的免疫抑制作用。
为了确定血小板对抗肿瘤免疫抑制作用的临床相关性,在药理学上解决血小板对免疫治疗的影响。在第0天皮下注射后在c57bl/6小鼠中建立b16-f1黑素瘤,然后在第7天用cy进行淋巴细胞清除,并在第8天输注离体引发的pmel细胞,以及抗血小板(ap)剂:阿司匹林和氯吡格雷。阿司匹林和氯吡格雷分别通过阻断环氧酶和adp受体来抑制血小板活化。单独的cy不能控制肿瘤,并且额外的ap在该模型中也没有抗肿瘤作用(图11a,左图)。用t细胞加cy有效控制黑素瘤大约一个月,但大多数小鼠最终复发。相反,抗血小板剂加过继性t细胞转移对b16-f1非常有效,大多数小鼠有超过3个月的无复发存活(图11a,右图)。作为进一步的证据,抗原特异性t细胞在同时接受抗血小板治疗和act的小鼠的血液,腹股沟淋巴结(iln)和脾脏中维持较高的数量(图11b)。重要的是,当转移的t细胞缺乏ifnγ时(图11c)或给予抗-ifnγ中和抗体时(图11d),抗血小板剂没有给予益处,证明抗血小板剂的作用是免疫介导的。
实施例5-材料和方法
细胞系和小鼠。前b细胞系(70z/3)是brianseed(哈佛大学)的礼物(randow和seed,2001)。4t1小鼠乳腺上皮细胞癌细胞系,野生型(wt)正常小鼠乳腺上皮细胞(nmumg)和具有hnrnpe1沉默的nmumg*亚系如先前所述(hussey等,2011)。b16-f1和293ft细胞系购自atcc。
6-8周龄雌性balb/c,c57bl/6j,nod-rag-/-,nsg种对(nodscidgamma)和pmel1t细胞受体(tcr)转基因(tg)小鼠购自杰克逊实验室公司(jacksonlaboratory)(美国缅因州巴港)。所有涉及小鼠的动物实验均由南卡罗来纳州医科大学的研究动物护理和使用委员会批准,并遵循既定的指南。共同饲养对照和处理的小鼠,并且在所有实验中使用6-8周龄雌性周龄匹配的小鼠。
组织微阵列和人血清。所有人肿瘤微阵列(tma)均由福尔马林固定的石蜡包埋组织制成。结肠,肺和两种乳腺癌中一种的tma是从南卡罗来纳医科大学(musc;南卡罗来纳州查尔斯顿)收集的标本开发的。这些tma中的每个患者样本在载玻片上以两个核心表示,并且每个核心直径为1mm。乳腺癌和前列腺癌的tma购自茵姆基因公司(imgenex,inc)(加利福尼亚州圣迭戈)。这些患者标本可以直径为2毫米的单个核心获得。临床和人口统计学信息来自musc的hollings癌症中心的癌症登记处或由商业来源提供。该研究得到了musc的研究审查委员会(irb)的批准。
免疫组织化学(ihc)。本研究中使用的小鼠抗人garp抗体(alx-804-867-c100,恩佐生物科学公司(enzolifesciences))首先通过western印迹在未转染和hgarp转染的人胚胎肾(hek)-293细胞中进行测试,并通过ihc使用hgarp-转染和对照载体转染的小鼠前b白血病细胞70z/3进行测试。两项分析均证明抗体和稀释液的特异性为1∶250(结肠癌)至1∶60(所有其他癌症)。
将tma载玻片在62℃下烘烤2小时,然后在二甲苯中脱蜡并再水合。然后通过在蒸汽锅中在柠檬酸盐缓冲液(ph=6.0)中煮沸30分钟来进行抗原修复。将载玻片在3%h2o2的dh2o中孵育7分钟,并且在室温下用2%正常马血清阻断非特异性结合2小时。将样品与抗h-garp抗体在4℃下孵育16小时,然后使用二抗(vectastainabc试剂盒)孵育并使用dab底物(vectorlabssk-4100)进行显色。染色对细胞质和细胞膜具有特异性,采用负核染色。
对于小鼠ihc,分离原发性肿瘤和肺。对于新鲜冷冻切片,将肿瘤组织置于oct培养基中,或对于固定切片,置于4%多聚甲醛中固定过夜。对于肿瘤和肺的苏木精和曙红(h&e)分析,将固定的组织在70%乙醇中孵育过夜,然后进行石蜡包埋,然后切割用于h&e染色。对于新鲜冷冻肿瘤切片上的p-smad-2/3,用4%多聚甲醛固定5μm切片,然后用3%h2o2孵育。为了使非特异性染色最小化,将切片与合适的动物血清在室温下孵育20分钟,然后在4℃下与一级抗-p-smad-2/3(ep823y;阿柏堪穆公司(abcam))一起孵育过夜。遵循抗大鼠vectastainabckil(载体实验室公司(vectorlabs))的标准方案。
garp和psmad-2/3的染色强度由经过委员会认证的病理学家(s.s.)评分,样品类型不知情(0:阴性;1:微弱;2:中度;3:强烈但强度低于4;和4:激烈)。还计算了tma中每个患者样品的阳性细胞百分比;在tma中,其中标本重复出现,两个核心的平均值用作代表值。实施斯氏t检验以比较分类变量,如正常与癌症或不同的疾病阶段或类别。使用x-tile软件进行garp与存活相关性的kaplan-meier分析(camp等,2004)。使用卡方检验测试群体特征在低和高garp表达者之间的统计学显著差异。
免疫荧光分析。将新鲜冷冻的肿瘤冷冻切片(5μm)风干,在丙酮中固定10分钟,然后与藻红蛋白偶联的抗cd31抗体(1∶50)一起孵育。通过在olympus荧光显微镜上成像后使用imagejv1.34软件程序(nih)计算cd31染色的面积来确定血管密度。
通过慢病毒表达的短发夹rna进行garp敲减。表达靶向小鼠garp转录物的短发夹rna(shrna)的慢病毒载体购自西格玛奥德里奇公司(sigma-aldrich)(密苏里州圣路易斯)。如先前所述(hong等,2013;wu等,2012),在hek293ft细胞中产生亲嗜性garpshrna和对照乱序慢病毒shrna颗粒。为了敲减nmumg*细胞中的garp,用靶向garp和乱序对照的慢病毒上清液转导细胞。使用抗小鼠garp抗体(ebioscience)通过rt-pcr(appliedbiosystemsstep-oneplus)和流式细胞术(bdverse)评估敲减效率。
生成garp表达载体。通过pcr扩增garp并将其亚克隆在migr1逆转录病毒载体中的bglii和hpai位点之间。通过将garp序列的细胞外结构域与编码鼠igg2a恒定区的fc部分的序列连接,产生用于表达重组garp-fc融合蛋白的cdna构建体。通过pcr从phcmv1载体扩增fc序列,并使用pcr从migr1逆转录病毒载体扩增garp。将两个片段连接并克隆到migr1逆转录病毒表达载体中。将亲嗜性garp和garp-fc逆转录病毒颗粒包装到pheonix-亲嗜性细胞中。前b细胞,4t1细胞和nmumg*细胞的病毒繁殖和转导基于已建立的方案(wu等,2012;zhang等,2015)。通过在转导后48小时在杀稻瘟菌素存在下培养至少72小时来稳定地选择细胞。
garp-fc的纯化。为了纯化garp-fc蛋白garp-fc,使用lipofectamine2000(英杰公司)根据制造商的说明将migr1载体转染到中国仓鼠卵巢(cho)细胞中。通过杀稻瘟素(5μg/ml)选择稳定转染的克隆,并使用抗小鼠garp和抗小鼠fc抗体在还原条件下通过sds-page和western印迹定量蛋白质表达。通过蛋白a亲和层析(ge医疗公司)从细胞培养上清液中纯化重组garp-fc。
抗garp抗体的产生和表征。用重组人garp(r&d系统公司,明尼苏达州明尼阿波利斯)和弗氏完全佐剂免疫4只balb/c小鼠,然后用稳定表达人garp的sp2/0细胞加强2-3次。来自具有高抗garp抗体滴度的小鼠的脾b细胞在聚乙二醇存在下与sp2/0细胞融合。在hat培养基中选择杂交瘤并通过有限稀释测定克隆。使用用空载体(70z/3-ev)稳定转导的70z/3细胞和人garp(70z/3-garp)的过表达,通过elisa和流式细胞术筛选和测定抗体的特异性。
蛋白质提取,免疫沉淀和western印迹分析。必要时通过胰蛋白酶-edta收获细胞,在pbs中洗涤,并在蛋白酶抑制剂混合物(西格玛奥德里奇公司)存在下在放射免疫沉淀测定(ripa)裂解缓冲液中在冰上裂解。通过bradford测定法(bio-rad)定量无核蛋白质裂解物,并使用抗小鼠garp(af6229;r&d系统公司),抗小鼠vimentin(d21h3;细胞信号传导公司),抗小鼠e-钙粘蛋白(24e10;细胞信号传导公司)和抗小鼠p-smad-2/3(ep823y;阿柏堪穆公司)在还原条件下通过sds-page和western印迹分析等量的裂解物。
细胞增殖和体外伤口愈合测定。将nmumg细胞(4×105)在无血清dmem(corningcellgro)中饥饿过夜。饥饿的细胞在指定的时间与garp-fc在2%fbsdmem中培养。为了测量细胞增殖,将2.5×104个细胞接种在完全培养基(dmem,10%fcs,1%青霉素-链霉素)中的96孔板中并孵育过夜。用3-[4,5-二甲基噻唑-2-基]-2,5-二苯基溴化四唑鎓(mtt)测定增殖,将其在指定时间加入细胞中并在37℃下再孵育3小时。然后除去培养基并通过摇动与100μl的dmso混合15分钟。然后使用读板器测量570nm处的吸光度。通过伤口愈合测定法测量细胞迁移:在100%汇合时,使用1ml移液管尖端制造两个平行的伤口。在24,48和72小时后评估迁移,并使用imagej软件(nih)测量伤口闭合的定量。
4t1肿瘤模型和garp抗体疗法。用5×105个细胞(4t1ev,4t1garp或4t1garp-fc)皮下(s.q.)接种在6-8周龄雌性balb/c小鼠的第四乳腺脂肪垫中。用数字游标卡尺每周监测肿瘤生长三次,并使用下式计算肿瘤体积:肿瘤体积(mm3)=[(宽度)2×长度]/2。在garp抗体疗法实验中,从肿瘤接种后3天开始,将抗garp抗体或多克隆同种型对照抗体(0.1mg/小鼠,在0.1mlpbs中;每周三次)腹膜内(i.p.)给予小鼠。对于用环磷酰胺(cy)和抗体的组合疗法,除了抗体处理之外,在肿瘤接种后3天用一次注射cy(4mg/小鼠)处理小鼠。在终点,处死小鼠并分离原发性肿瘤,引流ln,脾,肺和肝。通过胶原酶d(西格玛公司(sigma))消化然后通过histopaque-1083(西格玛公司)介导的密度分离来分离肿瘤浸润的淋巴细胞。
b16-f1肿瘤模型和过继性t细胞疗法(act)。使用2.5×105个细胞皮下接种三组(b16ev和b16garp-fc;每组n=5-7)6-8周龄雌性c57bl/6j小鼠的右腹侧,并且当指定时,在过继性t细胞治疗前一天用一次腹膜内注射cy(4mg/小鼠)处理。为了获得gp100特异性t细胞,用hgp100(25-33表位,1μg/ml,美国肽公司(americanpeptidecompany))和小鼠il-12(10ng/ml,shenandoa)刺激来自pmeltcr转基因雌性小鼠的脾细胞3天。在注射cy后一天,通过每个受体小鼠尾静脉注射2×106个活化的pmelt细胞进行act。用游标卡尺每周监测原发性肿瘤生长3次。在act后2、3、4和5周监测外周过继转移的pmel细胞。在37℃下在hgp100和布雷菲德菌素a(bfa)存在下刺激pmel细胞3小时评估离体pmelifn-γ产生并通过流式细胞术分析。
nmumg肿瘤模型。使用5×105个细胞(nmumg*-ev,garp敲减nmumg*)皮下接种于雌性nod-rag-1-/-(n=5每组;6-8周龄)小鼠的第四左乳腺脂肪垫中。称重动物并每周测量肿瘤。在终点,收获原发性肿瘤,肺和肝脏。在另一个实验中,用5×105个细胞(nmumg-garp-luc,nmumg-garp-fc-luc或nmumg-luc细胞)皮下接种于雌性nod-rag-1-/-小鼠(每组n=4-5;6-8周龄)的第四左乳腺脂肪垫中。如下每周评估体内荧光素酶成像:给小鼠腹膜内注射d-荧光素(帕金埃尔默公司(perkinelmer)),剂量为每只小鼠150mg/kg并麻醉。然后使用xenogenivis成像系统获取生物发光图像。使用livingimage软件(xenogen)将生物发光信号定量为在所定义的感兴趣区域中的光子通量(光子/s/cm2)。
tgf-β1,garp和garp-tgf-β1分析。根据制造商的方案,使用tgf-β1和garpelisa试剂盒(白乐津公司(biolegend),加利福尼亚州圣迭戈)在人和小鼠血清中测量活性tgf-β1,总tgf-β1和可溶性garp。为了通过elisa测量garp-tgf-β1复合物,根据制造商的说明书(白乐津公司,加利福尼亚州圣迭戈),用tgf-β1捕获抗体包被96孔板。将样品在室温下孵育2小时,然后与我们实验室中开发的抗hgarp检测抗体一起再孵育2小时。
对于mfb-f11功能测定,将mfb-f11细胞(来自斯坦福大学的tonywyss-coray的礼物)在含有10%fbs和1%青霉素/链霉素的dmem中培养。每孔接种2×104个细胞并孵育过夜。在加入稀释的血清或肿瘤上清液之前,将细胞血清饥饿2-3小时。将稀释的血清或肿瘤上清液样品孵育24小时,然后使用quanti-blue培养基(invivogen,加利福尼亚州圣迭戈)进行分析(tesseur等,2006)。
统计学分析。在tma中,样品一式两份点样,两个核心的平均值被用作代表值。实施斯氏t检验以比较分类变量,如正常与癌症或不同的疾病阶段或类别。使用x-tile软件进行garp与存活相关性的kaplan-meier分析(camp等,2004)。使用卡方检验测试群体特征在低和高garp表达者之间的统计学显著差异。使用双因素方差分析(anova)进行肿瘤曲线分析;用graphpadprism使用双尾学生t检验分析所有其他实验。所有数据表示为平均值±sem。p值<0.05被认为具有统计学显著性。
实施例6-针对garp的抗体(lrrc32)
表1.garp单克隆抗体克隆
***
根据本公开,本文公开和权利要求的所有方法无需过多实验即可进行和执行。虽然以优选实施方式的形式描述了本发明的组合物和方法,但本领域技术人员显然了解,可将各种改变施用于本文所述方法以及本文所述方法的各个步骤或各步骤的顺序,而并不背离本发明的概念、精神和范围。更具体说,显然某些化学和生理相关试剂可代替本文所述的试剂,同时实现相同或类似的结果。认为本领域技术人员了解的所有这些类似的取代物或改进都在所附权利要求所定义的本发明的精神、范围和概念内。
参考文献
下列参考文献都特定通过引用纳入本文,它们对本文陈述的内容提供示例性、程序上或其他细节上的补充。
ahn,y.o.,lee,j.c.,sung,m.w.,和heo,d.s.(2009).在头颈部鳞状细胞癌细胞系中膜结合tgf-β1的存在及其被il-2激活的免疫细胞衍生的ifn-γ调节(presenceofmembrane-boundtgf-betalanditsregulationbyil-2-activatedimmunecell-derivedifn-gammainheadandnecksquamouscellcarcinomacelllines).journalofimmunology182,6114-6120.
akhurst,r.j.,和hata,a.(2012).靶向疾病中tgfβ信号传导途径(targetingthetgfbetasignallingpathwayindisease).naturereviewsdrugdiscovery11,790-811.
baker,k.,raut,p.,和jass,j.r.(2008).结肠直肠癌细胞表达功能性细胞表面结合的tgfβ(colorectalcancercellsexpressfunctionalcellsurface-boundtgfbeta).internationaljournalofcancerjournalinternationalducancer122,1695-1700.
beyer,m.,和schultze,j.l.(2006).癌症中的调节性t细胞(regulatorytcellsincancer).blood108,804-811.
bhowmick,n.a.,ghiassi,m.,bakin,a.,aakre,m.,lundquist,c.a.,engel,m.e.,arteaga,c.l.,和moses,h.l.(2001).转化生长因子-β1通过rhoa依赖性机制介导上皮细胞向间充质细胞转分化(transforminggrowthfactor-betalmediatesepithelialtomesenchymaltransdifferentiationthrougharhoa-dependentmechanism).molecularbiologyofthecell12,27-36.
camp,r.l.,dolled-filhart,m.,和rimm,d.l.(2004).x-tile:用于生物标记评估和基于结果的切割点优化的新型生物信息学工具(x-tile:anewbio-informaticstoolforbiomarkerassessmentandoutcome-basedcut-pointoptimization).clinicalcancerresearch:anofficialjournaloftheamericanassociationforcancerresearch10,7252-7259.
carrillo-galvez,a.b.,cobo,m.,cuevas-ocana,s.,gutierrez-guerrero,a.,sanchez-gilabert,a.,bongarzone,p.,garcia-perez,a.,munoz,p.,benabdellah,k.,toscano,m.g.等(2015).间充质基质细胞表面表达garp/lrrc32:对其生物学和免疫调节能力的影响(mesenchymalstromalcellsexpressgarp/lrrc32ontheirsurface:effectsontheirbiologyandimmunomodulatorycapacity).stemcells33,183-195.
chen,x.,yang,y.,zhou,q.,weiss,j.m.,howard,o.z.,mcpherson,j.m.,wakefield,l.m.,和oppenheim,j.j.(2014).在乳腺癌小鼠模型中使用抗tgfβ抗体和环磷酰胺进行有效化学免疫疗法(effectivechemoimmunotherapywithanti-tgfbetaantibodyandcyclophosphamideinamousemodelofbreastcancer).plosone9,e85398.
cuende,j.,lienart,s.,dedobbeleer,o.,vanderwoning,b.,deboeck,g.,stockis,j.,huygens,c.,colau,d.,somja,j.,delvenne,p.等(2015).针对garp/tgf-β1复合物的单克隆抗体在体内抑制人调节性t细胞的免疫抑制活性(monoclonalantibodiesagainstgarp/tgf-betalcomplexesinhibittheimmunosuppressiveactivityofhumanregulatorytcellsinvivo).sciencetranslationalmedicine7,284ra256.
derynck,r.,akhurst,r.j.,和balmain,a.(2001).肿瘤抑制和癌症进展中的tgf-β信号传导(tgf-betasignalingintumorsuppressionandcancerprogression).naturegenetics29,117-129.
edwards,j.p.,fujii,h.,zhou,a.x.,creemers,j.,unutmaz,d.,和shevach,e.m.(2013).调节小鼠t细胞上garp/潜伏tgf-β1复合物的表达及其在调节性t细胞和th17分化中的作用(regulationoftheexpressionofgarp/latenttgf-betalcomplexesonmousetcellsandtheirroleinregulatorytcellandth17differentiation).journalofimmunology190,5506-5515.
flavell,r.a.,sanjabi,s.,wrzesinski,s.h.,和licona-limon,p.(2010).tgfβ在肿瘤环境中的免疫细胞极化(thepolarizationofimmunecellsinthetumourenvironmentbytgfbeta).naturereviewsimmunology10,554-567.
gauthy,e.,cuende,j.,stockis,j.,huygens,c.,lethe,b.,collet,j.f.,bommer,g.,coulie,p.g.,和lucas,s.(2013).garp受mirna调节,并控制人调节性t细胞产生潜伏tgf-β1(garpisregulatedbymirnasandcontrolslatenttgf-betalproductionbyhumanregulatorytcells).plosone8,e76186.
hahn,s.a.,stahl,h.f.,becker,c.,correll,a.,schneider,f.j.,tuettenberg,a.,和jonuleit,h.(2013).可溶性garp对人cd4(+)t细胞具有有效的抗炎和免疫调节作用(solublegarphaspotentantiinflammatoryandimmunomodulatoryimpactonhumancd4(+)tcells).blood122,1182-1191.
hong,f.,liu,b.,chiosis,g.,gewirth,d.t.,和li,z.(2013).α1结构域的α7螺旋区域是整合素与内质网伴侣gp96的结合中的关键:癌症转移的潜在治疗靶标(alpha7helixregionofalphaidomainiscrucialforintegrinbindingtoendoplasmicreticulumchaperonegp96:apotentialtherapeutictargetforcancermetastasis).thejournalofbiologicalchemistry288,18243-18248.
howley,b.v.,hussey,g.s.,link,l.a.,和howe,p.h.(2015).tgfβ通过rna结合蛋白hnrnpe1对抑制素βa的翻译调节增强了上皮-间充质转化细胞的侵袭性(translationalregulationofinhibinbetaabytgfbetaviatherna-bindingproteinhnrnpe1enhancestheinvasivenessofepithelial-to-mesenchymaltransitionedcells).oncogene.
hussey,g.s.,chaudhury,a.,dawson,a.e.,lindner,d.j.,knudsen,c.r.,wilce,m.c.,merrick,w.c.,和howe,p.h.(2011).鉴定在翻译延伸步骤调节肿瘤发生的mrnp复合物(identificationofanmrnpcomplexregulatingtumorigenesisatthetranslationalelongationstep).molecularcell41,419-431.
huygens,c.,lienart,s.,dedobbeleer,o.,stockis,j.,gauthy,e.,coulie,p.g.,和lucas,s.(2015).溶酶体相关跨膜蛋白4b(laptm4b)降低人类调节性t细胞中转化生长因子β1(tgf-β1)的产生(lysosomal-associatedtransmembraneprotein4b(laptm4b)decreasestransforminggrowthfactorbetal(tgf-betal)productioninhumanregulatorytcells).thejournalofbiologicalchemistry290,20105-20116.
kehrl,j.h.,wakefield,l.m.,roberts,a.b.,jakowlew,s.,alvarez-mon,m.,derynck,r.,sporn,m.b.,和fauci,a.s.(1986).人t淋巴细胞产生转化生长因子β及其在调节t细胞生长中的潜在作用(productionoftransforminggrowthfactorbetabyhumantlymphocytesanditspotentialroleintheregulationoftcellgrowth).thejournalofexperimentalmedicine163,1037-1050.
kopp,h.g.,placke,t.,和salih,h.r.(2009).血小板衍生的转化生长因子-β下调nkg2d,从而抑制自然杀伤细胞的抗肿瘤反应性(platelet-derivedtransforminggrowthfactor-betadown-regulatesnkg2dtherebyinhibitingnaturalkillercellantitumorreactivity).cancerresearch69,7775-7783.
li,m.o.,和flavell,r.a.(2008).tgf-β:所有t细胞交易的主人(tgf-beta:amasterofalltcelltrades).cell134,392-404.
li,y.,kim,b.g.,qian,s.,letterio,j.j.,fung,j.j.,lu,l.,和lin,f.(2015).肝星状细胞通过来自细胞表面结合的潜伏tgf-β1/garp复合物的活性tgf-β1抑制t细胞(hepaticstellatecellsinhibittcellsthroughactivetgf-betalfromacellsurface-boundlatenttgf-beta1/garpcomplex).joumalofimmunology195,2648-2656.
lyons,r.m.,gentry,l.e.,purchio,a.f.,和moses,h.l.(1990).纤溶酶激活潜伏重组转化生长因子β1的机制(mechanismofactivationoflatentrecombinanttransforminggrowthfactorbeta1byplasmin).jcellbiol110,1361-1367.
massague,j.(2008).癌症中的tgfβ(tgfbetaincancer).cell134,215-230.
muranski,p.,boni,a.,antony,p.a.,cassard,l.,irvine,k.r.,kaiser,a.,paulos,c.m.,palmer,d.c.,touloukian,c.e.,ptak,k.等(2008).肿瘤特异性th17极化细胞根除大的黑素瘤(tumor-specificth17-polarizedcellseradicatelargeestablishedmelanoma).blood112,362-373.
oft,m.,heider,k.h.,和beug,h.(1998).tgfβ信号传导是癌细胞侵袭和转移所必需的(tgfbetasignalingisnecessaryforcarcinomacellinvasivenessandmetastasis).currbiol8,1243-1252.
ollendorff,v.,noguchi,t.,delapeyriere,o.,和birnbaum,d.(1994).garp基因编码富含亮氨酸重复序列蛋白家族的新成员(thegarpgeneencodesanewmemberofthefamilyofleucine-richrepeat-containingproteins).cellgrowth&differentiation:themolecularbiologyjournaloftheamericanassociationforcancerresearch5,213-219.
overwijk,w.w.,theoret,m.r.,finkelstein,s.e.,surman,d.r.,dejong,l.a.,vyth-dreese,f.a.,dellemijn,t.a.,antony,p.a.,spiess,p.j.,palmer,d.c.等(2003).逆转自身反应性cd8+t细胞的功能耐受状态后的肿瘤消退和自身免疫(tumorregressionandautoimmunityafterreversalofafunctionallytolerantstateofself-reactivecd8+tcells).thejournalofexperimentalmedicine198,569-580.
padua,d.,zhang,x.h.,wang,q.,nadal,c.,gerald,w.l.,gomis,r.r.,和massague,j.(2008).tgfβ通过血管生成素样4引发乳腺肿瘤的肺转移(tgfbetaprimesbreasttumorsforlungmetastasisseedingthroughangiopoietin-like4).cell133,66-77.
pertovaara,l.,kaipainen,a.,mustonen,t.,orpana,a.,ferrara,n.,saksela,o.,和alitalo,k.(1994).在成纤维细胞和上皮细胞中响应转化生长因子-β诱导血管内皮生长因子(vascularendothelialgrowthfactorisinducedinresponsetotransforminggrowthfactor-betainfibroblasticandepithelialcells).jbiolchem269,6271-6274.
pulaski,b.a.,和ostrand-rosenberg,s.(2001).小鼠4t1乳腺肿瘤模型(mouse4t1breasttumormodel).《新编免疫学方案》(currentprotocolsinimmunology),johnecoligan[等]编,第20章,单元2022.
randow,f.,和seed,b.(2001).内质网伴侣蛋白gp96是先天免疫所必需的,但不是细胞活力所必需的(endoplasmicreticulumchaperonegp96isrequiredforinnateimmunitybutnotcellviability).naturecellbiology3,891-896.
roberts,a.b.,sporn,m.b.,assoian,r.k.,smith,j.m.,roche,n.s.,wakefield,l.m.,heine,u.i.,liotta,l.a.,falanga,v.,kehrl,j.h.等(1986).β型转化生长因子:体内纤维化和血管生成的快速诱导和体外胶原形成的刺激(trahsforminggrowthfactortypebeta:rapidinductionoffibrosisandangiogenesisinvivoandstimulationofcollagenformationinvitro).proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica83,4167-4171.
rubinstein,m.p.,su,e.w.,suriano,s.,cloud,c.a.,andrijauskaite,k.,kesarwani,p.,schwartz,k.m.,williams,k.m.,johnson,c.b.,li,m.等(2015).白细胞介素-12增强鼠和人cd8(+)t细胞的功能和抗肿瘤活性(interleukin-12enhancesthefunctionandanti-tumoractivityinmurineandhumancd8(+)tcells).cancerimmunology,immunotherapy:cii64,539-549.
sato,y.,和rifkin,d.b.(1989).周细胞和平滑肌细胞抑制内皮细胞运动:共培养期间纤溶酶激活潜伏转化生长因子-β1样分子(inhibitionofendothelialcellmovementbypericytesandsmoothmusclecells:activationofalatenttransforminggrowthfactor-beta1-likemoleculebyplasminduringco-culture).jcellbiol109,309-315.
siegel,p.m.,shu,w.,cardiff,r.d.,muller,w.j.,和massague,j.(2003).转化生长因子β信号传导损害neu诱导的乳腺肿瘤发生,同时促进肺转移(transforminggrowthfactorbetasignalingimpairsneu-inducedmammarytumorigenesiswhilepromotingpulmonarymetastasis).proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica100,8430-8435.
stockis,j.,colau,d.,coulie,p.g.,和lucas,s.(2009).膜蛋白garp是活化的人treg表面上潜伏tgf-β的受体(membraneproteingarpisareceptorforlatenttgf-betaonthesurfaceofactivatedhumantreg).europeanjournalofimmunology39,3315-3322.
sunderkotter,c.,goebeler,m.,schulze-osthoff,k.,bhardwaj,r.,和sorg,c.(1991).巨噬细胞衍生的血管生成因子(macrophage-derivedangiogenesisfactors).pharmacolther51,195-216.
szepetowski,p.,ollendorff,v.,grosgeorge,j.,courseaux,a.,bimbaum,d.,theillet,c.,和gaudray,p.(1992).人乳腺癌中11q13.5-q14的dna扩增(dnaamplificationat11q13.5-q14inhumanbreastcancer).oncogene7,2513-2517.
tesseur,i.,zou,k.,berber,e.,zhang,h.,和wyss-coray,t.(2006).用于测量生物活性tgf-β的高灵敏度和特异性生物测定(highlysensitiveandspecificbioassayformeasuringbioactivetgf-beta).bmccellbiology7,15.
tran,d.q.(2012).tgf-β:foxp3(+)调节性t细胞的剑,魔杖和屏障(tgf-beta:thesword,thewand,andtheshieldoffoxp3(+)regulatorytcells).jmolcellbiol4,29-37.
tran,d.q.,andersson,j.,wang,r.,ramsey,h.,unutmaz,d.,和shevach,e.m.(2009).garp(lrrc32)对血小板上潜伏tgf-β的表面表达和活化的foxp3+调节性t细胞至关重要(garp(lrrc32)isessentialforthesurfaceexpressionoflatenttgf-betaonplateletsandactivatedfoxp3+regulatorytcells).proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica106,13445-13450.
wang,r.,kozhaya,l.,mercer,f.,khaitan,a.,fujii,h.,和unutmaz,d.(2009).garp的表达选择性地鉴定活化的人foxp3+调节性t细胞(expressionofgarpselectivelyidentifiesactivatedhumanfoxp3+regulatorytcells).proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica106,13439-13444.
wang,r.,wan,q.,kozhaya,l.,fujii,h.,和unutmaz,d.(2008).鉴定调节性t细胞特异性细胞表面分子,其介导抑制信号并诱导foxp3表达(identificationofaregulatorytcellspecificcellsurfacemoleculethatmediatessuppressivesignalsandinducesfoxp3expression).plosone3,e2705.
wang,r.,zhu,j.,dong,x.,shi,m.,lu,c.,和springer,t.a.(2012).garp调节tgfβ的生物利用度和活化(garpregulatesthebioavailabilityandactivationoftgfbeta).molecularbiologyofthecell23,1129-1139.
wu,s.,hong,f.,gewirth,d.,guo,b.,liu,b.,和li,z.(2012).分子伴侣gp96/grp94通过其c末端疏水结构域与toll样受体和整联蛋白相互作用(themolecularchaperonegp96/grp94interactswithtoll-likereceptorsandintegrinsviaitsc-terminalhydrophobicdomain).thejournalofbiologicalchemistry287,6735-6742.
xie,l.,law,b.k.,aakre,m.e.,edgerton,m.,shyr,y.,bhowmick,n.a.,和moses,h.l.(2003).在小鼠乳腺上皮细胞系中转化生长因子β调节的基因表达(transforminggrowthfactorbeta-regulatedgeneexpressioninamousemammaryglandepithelialcellline).breastcancerresearch:bcr5,r187-198.
xu,j.,lamouille,s.,和derynck,r.(2009).tgf-β诱导的上皮细胞向间充质细胞转化(tgf-beta-inducedepithelialtomesenchymaltransition).cellresearch19,156-172.
yu,q.,和stamenkovic,i.(2000).细胞表面定位的基质金属蛋白酶-9蛋白水解活化tgf-β并促进肿瘤侵袭和血管生成(cellsurface-localizedmatrixmetalloproteinase-9proteolyticallyactivatestgf-betaandpromotestumorinvasionandangiogenesis).genes&development14,163-176.
zhang,y.,wu,b.x.,metelli,a.,thaxton,j.e.,hong,f.,rachidi,s.,ansa-addo,e.,sun,s.,vasu,c.,yang,y.等(2015).gp96是garp伴侣并控制调节性t细胞功能(gp96isagarpchaperoneandcontrolsregulatorytcellfunctiohs).thejournalofclinicalinvestigation125,859-869.
序列表
<110>li,zihai(z·李)
<120>通过靶向糖蛋白a重复优势蛋白(garp)治疗和诊断癌症以及提供有效免疫疗法的方法
<130>mesc.p0096wo
<150>62315069
<151>2016-03-30
<150>62315121
<151>2016-03-30
<160>17
<170>patentinversion3.5
<210>1
<211>9
<212>prt
<213>智人(homosapiens)
<400>1
glytyrserilethrserasptyrala
15
<210>2
<211>7
<212>prt
<213>智人(homosapiens)
<400>2
ilesertyrserglyserthr
15
<210>3
<211>18
<212>prt
<213>智人(homosapiens)
<400>3
alalysserglyglyasptyrtyrglysersersertyrtrptyrphe
151015
aspval
<210>4
<211>126
<212>prt
<213>智人(homosapiens)
<400>4
leuaspvalglnleuglngluserglyproglyleuvallysproser
151015
glnserleuserleuthrcysthrvalthrglytyrserilethrser
202530
asptyralatrpasntrpileargglnpheproglyasnlysleuglu
354045
trpmetglytyrilesertyrserglyserthrsertyrthrproser
505560
leulysserargileserilethrargaspthrserlysasnhisphe
65707580
pheleuglnserasnservalthrthrgluaspthralathrtyrtyr
859095
cysalalysserglyglyasptyrtyrglysersersertyrtrptyr
100105110
pheaspvaltrpglyalaglythrthrvalthrvalserser
115120125
<210>5
<211>12
<212>prt
<213>智人(homosapiens)
<400>5
glnserleuleuasnserargserglnlysasntyr
1510
<210>6
<211>3
<212>prt
<213>智人(homosapiens)
<400>6
argalaser
1
<210>7
<211>9
<212>prt
<213>智人(homosapiens)
<400>7
glnasnasphissertyrprophethr
15
<210>8
<211>113
<212>prt
<213>智人(homosapiens)
<400>8
aspileglnmetthrglnserproserserleuservalseralagly
151015
glulysvalthrmetsercyslysserserglnserleuleuasnser
202530
argserglnlysasntyrleualatrptyrglnglnlysproglygln
354045
proprolysleuleuiletyrargalaserthrargglyserglyval
505560
proaspargphethrglyserglyserglythraspphethrleuthr
65707580
ileserservalglnalagluaspleualavaltyrtyrcysglnasn
859095
asphissertyrprophethrpheglyserglythrlysleugluile
100105110
lys
<210>9
<211>8
<212>prt
<213>智人(homosapiens)
<400>9
glyphethrpheserasntyrval
15
<210>10
<211>8
<212>prt
<213>智人(homosapiens)
<400>10
ileserserglyglysertyrthr
15
<210>11
<211>13
<212>prt
<213>智人(homosapiens)
<400>11
alaargglytyraspasnglyasptyrvalmetasptyr
1510
<210>12
<211>120
<212>prt
<213>智人(homosapiens)
<400>12
gluvallysleuvalgluserglyglyglyservallysproglygly
151015
serleulysleusercysalaalaserglyphethrpheserasntyr
202530
valmetsertrpvalargglnthrproglulysargleuglutrpval
354045
alathrileserserglyglysertyrthrtyrtyrproaspserval
505560
lysglyargleuthrileserargaspasnalalysasnthrleutyr
65707580
leuglnmetserserleuargsergluaspthralamettyrtyrcys
859095
alaargglytyraspasnglyasptyrvalmetasptyrtrpglygln
100105110
glythrservalthrvalserser
115120
<210>13
<211>10
<212>prt
<213>智人(homosapiens)
<400>13
gluservalaspthrtyrglyasnserphe
1510
<210>14
<211>3
<212>prt
<213>智人(homosapiens)
<400>14
argalaser
1
<210>15
<211>9
<212>prt
<213>智人(homosapiens)
<400>15
glnglnthrasngluhisproprothr
15
<210>16
<211>111
<212>prt
<213>智人(homosapiens)
<400>16
aspilevalleuthrglnserproalaserleualavalserleugly
151015
glnargalathrilesercysargalasergluservalaspthrtyr
202530
glyasnserphemethistrptyrglnglnileproglyglnpropro
354045
lysvalleuiletyrargalaserasnleugluserglyileproala
505560
argpheserglyserglyserargthraspphethrleuthrileasn
65707580
provalglualaglyaspvalalathrtyrtyrcysglnglnthrasn
859095
gluhisproprothrpheglyglyglythrlysleugluilelys
100105110
<210>17
<211>12
<212>prt
<213>智人(homosapiens)
<400>17
leuargthrleuserleugluasnserleuthrarg
1510
- 还没有人留言评论。精彩留言会获得点赞!