超长粒水稻的培育方法与流程
本发明属于水稻分子育种技术领域。具体涉及一种超长粒水稻的培育方法。本发明利用图位克隆的方法分离克隆了一个新的水稻粒长基因gl3.3,该基因与已知粒长基因gs3聚合能够形成超长粒长的水稻材料。利用gs3和gl3.3的kasp分子标记引物能够有效的在育种中选择并聚合二者的优势等位基因,选择具有gs3gl3.3基因型组合的超长粒的家系,可以显著提高稻米品质的育种效率。
背景技术:
水稻是我国的主要粮食作物,而水稻的外观品质是决定水稻商品价值的重要指标。长久以来,我国水稻育种家与种业公司一直将产量作为育种的终极目标,而水稻品质的改良则比较滞后(孔宪琴等,2013)。我国大部分审定品种的稻米品质较为一般,无法满足国民对品质优良稻米的需求,加工成商品米后价格较低,一定程度上加深了“稻强米弱”的现象,导致我国稻米进口量的上升(孔宪琴等,2013;于红燕和刘世义,2016)。因粳稻品质普遍优于籼稻,最近几年,常规粳稻品种在我国水稻主要种植地区种植面积逐年扩大。我国品质优良的籼稻品种非常少,因此籼稻特别是杂交籼稻的外观品质的遗传改良迫在眉睫。
稻米的外观品质主要由粒形和堊白决定,粒形修长、垩白率低的籼稻受到消费者的青睐(bao,2014)。传统育种中改良稻米的外观品质,通常是将产量高的品系与品质优的品系进行杂交,然后不断的同产量高的品系自交从而得到产量品质都优良的品系。但是,由于粒长、堊白等外观品质都属于数量性状,由多个基因调控,需要在较大的群体中进行选择,并且性状考察也比较耗时耗力,实际改良过程难度较大。通过分子育种的方法,利用已克隆粒形、堊白基因的分子标记,将有利的等位基因聚合在一起,只需鉴定基因型,无需鉴定表型,节省了人力物力,是改良稻米外观品质的有效方法。
分子育种具有许多优势,但是该方法首先要有稳定可靠的基因资源和基因型组合。迄今为止,研究人员利用不同的群体和方法,克隆一些稻米粒形和堊白的主效基因。品种测序和关联分析的研究结果都表明gw5和gs3分别是水稻粒宽和粒长最主要和主效的基因(takanokai等,2009;duan等,2017;zhou等,2017)。而其它的粒形主效基因大多属于稀有变异,如粒宽基因gw2、粒长基因gl3、种子大小基因gs2(song等,2007;qi等,2012;hu等,2015)。还有一些重要而非主效的粒形基因,如种子大小基因gs5、长宽比基因gw8和长宽比基因gw7等(li等,2011;wang等,2012;wang等,2015)。而堊白基因目前只有chalk5被克隆(li等,2014)。这些基因为稻米外观品质的改良提供了宝贵的基因资源。虽然克隆的粒形基因已经不少,但是可供用来改良外观品质的组合很少。wang等(2012)利用将gs3和gw8无功能的等位型gs3和gw8聚合到短圆粒材料hjz74中,成功将该材料改良成修长的粒形。wang等(2014)后来通过直接研究具有修长粒形的材料台风优55和台风优208,发现它们因为就有gs3和gw7的基因型组合而表现为修长的粒形。
因此,从现有材料中直接挖掘有效的基因型组合是更为合理有效的方法。申请人利用自然存在的超长粒材料与短粒材料进行杂交,然后解析造成超长粒形的原因,最后总结出有效的基因型组合用来改良水稻粒长。通过开发出有效基因型组合的分子标记,为稻米的粒形改良提供有效的技术支撑。
技术实现要素:
本发明的目的是提供一种培育超长粒水稻的方法,通过试验检测两个粒长基因的功能标记,可以在苗期就准确预测水稻植株的粒长水平,从而加快超长粒水稻家系的选择进度。
本发明鉴定了两个粒长的基因,并提供了功能性的分子标记或称标记引物。借助这些分子标记辅助选择技术可以有目的地将柱头外露基因精确导入和聚合,从而高效率快速选育具有超长粒形的家系,节约人力物力,保持水稻杂交制种的稳产和高产。
本发明通过以下技术方案实现:
申请人通过利用基因gs3(登陆号os03g0407400)的功能性分子标记引物kasp-gs3通过pcr扩增,然后经过荧光定量pcr读带以检测该基因的基因型,所述分子标记引物的dna序列如下所示:
左端引物序列1(kasp-gs3f1):gaaggtgaccaagttcatgctacgctgcctccagatgctgc,
左端引物序列2(kasp-gs3f2):gaaggtcggagtcaacggattacgctgcctccagatgctga,
右端引物序列(sf28r):gtagtacaaaaagaaacagc;
申请人又通过另一个基因gl3.3(登陆号os03g0841800)的功能性分子标记引物kasp-gl3.3通过pcr扩增,然后通过荧光定量pcr读带以检测该基因的基因型,所述分子标记引物的dna序列如下所示:
左端引物序列1(kasp-gl3.3f1):gaaggtgaccaagttcatgctttttggggttgtatatcagg,
左端引物序列2(kasp-gl3.3f2):gaaggtcggagtcaacggattttttggggttgtatatcaga,
右端引物序列(kasp-gl3.3r):cacatagagaaatttttac;
申请人通过验证,证明上述两个分子标记引物(即,kasp-gs3和kasp-gl3.3)可以应用于选育超长粒形水稻家系的遗传改良。
申请人提供了一种筛选水稻粒长主效基因gs3的分子标记方法,其步骤包括:利用粒长主效基因gs3的分子标记引物(kasp-gs3)扩增待检水稻基因组dna,并利用荧光定量pcr检测所得的扩增产物,带型聚类于x轴,读带基因型为allelex(5016),证明水稻粒长主效基因gs3的增效等位基因gs3的存在。
与此同时,申请人提供了另一种筛选水稻粒长基因gl3.3的分子标记方法,其步骤包括:利用粒长基因gw5的分子标记(即,kasp-gl3.3)扩增待检水稻基因组dna,并利用荧光定量pcr检测所得的扩增产物,带型聚类于x轴,读带基因型为allelex(5016),证明水稻长粒主效基因gl3.3的增效等位基因gl3.3的存在。
申请人提供了一种利用分子标记引物在选育超长粒水稻的辅助选择中的应用,所述的应用步骤包括:
(1)将鉴定为性状优良的短粒和中长粒骨干材料分别与具有高超长粒基因型组合gs3gl3.3的栽培稻“南洋占”杂交,分别得到f1a和f1b;
(2)以步骤(1)中得到的f1a与短粒骨干材料连续回交5代,每代利用分子标记引物kasp-gs3和分子标记引物kasp-gl3.3鉴定真杂种,直到获得近等基因系bc5f1a;以步骤(1)中得到的f1b与中长粒骨干材料连续回交5代,每代利用分子标记引物kasp-gl3.3鉴定真杂种,直到获得近等基因系bc5f1b;
(3)将bc5f1a自交得到bc5f2a,利用分子标记引物kasp-gs3和分子标记引物kasp-gl3.3鉴定出基因型为纯合基因型组合gs3gl3.3的单株,即为改良后的短粒骨干材料;将bc5f1b自交得到bc5f2b,利用分子标记引物kasp-gl3.3鉴定基因型为纯合基因型组合gl3.3的单株,
其中:
所述的分子标记引物kasp-gs3的核苷酸序列如序列表seqidno:5至seqidno:7所示所示;
所述的分子标记引物kasp-gl3.3的核苷酸序列如序列表seqidno:8至seqidno:10所示。
本发明的优点在于:
(1)本发明发现了一个新的粒长基因gl3.3,该基因能够提高水稻的粒长。并且gl3.3与已克隆粒长基因gs3存在上位性互作,二者的聚合能够形成超长的水稻粒形。前人虽然克隆一些粒长基因,但是对于这些基因的聚合效应没有进行研究。并且一般两个基因的聚合效应会小于两个基因单独效应的累加,而gl3.3和gs3的聚合效应大于两个基因单独效应的累加。
(2)本发明鉴定的粒长主效基因经过了遗传转化验证。利用gl3.3的转基因材料,申请人验证了该基因能够有效的提高水稻的粒长。gl3.3通过增加颖壳纵向细胞的长度来提高水稻的粒长而不影响细胞的宽度。因此gl3.3是一种特殊的类型,与之前发现的粒形基因都不相同,能够更有效的用于粒长的改良。
(3)本发明提供了粒长主效基因gl3.3的功能性分子标记。功能性的分子标记检测水稻植株中的粒长基因位点,仅在苗期就能快速鉴定出长粒的纯合基因型单株,及时淘汰短粒单株,不仅节约生产成本,而且大大提高了选择效率,极大地缩短水稻品种的育种周期。而前人研究大多都没有提供基因的功能性分子标记。
附图说明
序列表seqidno:1是本发明的粒长基因gl3.3的编码区(cds)的核苷酸序列及其对应的氨基酸序列。
序列表seqidno:2是粒长基因gl3.3编码的蛋白质序列。
序列表seqidno:3是粒长基因gs3的编码区(cds)的核苷酸序列及其对应的氨基酸序列。
序列表seqidno:4是粒长基因gs3编码的蛋白质序列。
序列表seqidno:5是左端引物序列1(即分子标记引物kasp-gs3的左端引物序列1)。
序列表seqidno:6是左端引物序列2(即分子标记引物kasp-gs3的左端引物序列2)。
序列表seqidno:7是右端引物序列(即分子标记引物kasp-gs3的右端引物序列)。
序列表seqidno:8是左端引物序列1(即分子标记引物kasp-gl3.3的左端引物1)。
序列表seqidno:9是左端引物序列2(即分子标记引物kasp-gl3.3的左端引物2)。
序列表seqidno:10是右端引物序列(即分子标记引物kasp-gl3.3的右端引物)。
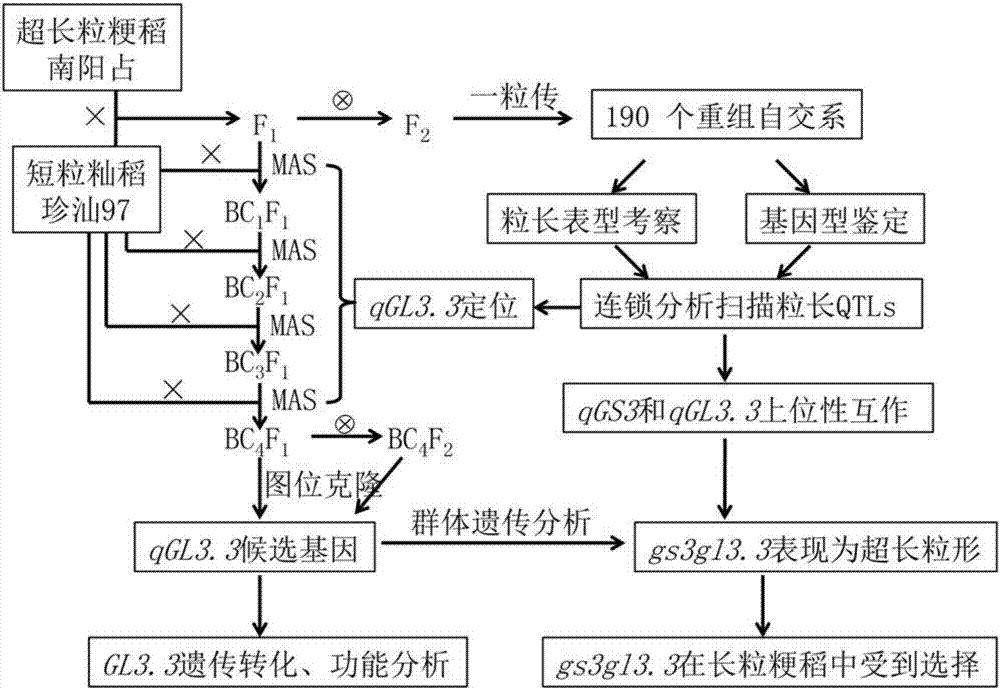
图1:是本发明的总体技术路线图。
图2:本发明粒长基因gl3.3的图位克隆。附图标记说明:图2中的a图是双亲和近等基因系的种子的外观形态;图2中的b图是重组自交系中粒形qtl初定位的结果;图2中的c图和图2中的d图是gl3.3精细定位过程;图2中的e图是候选基因在双亲间的等位差异。
图3:本发明的粒长基因gl3.3的遗传转化验证过程。附图标记说明:图3中的a图至e图是水稻品种珍汕97(zs97)和两种用crispr系统敲除材料的种子后代的表型差异:其中,图3中的a图是zs97与敲除gl3.3基因的突变体材料的水稻种子;图2中的b图是zs97与与两种突变体材料的核苷酸序列的差异比较;图3中的c图是转基因水稻种子粒长的表型差异;图3中的d图是转基因种子粒宽的表型差异;图3中的e图是转基因水稻种子的千粒重表型差异;图3中f图至图3中的i图为gl3.3基因互补对水稻品种表型差异的检测,其中:图3中的f图是水稻品种“南阳占”的阳性和阴性材料的种子表型的差异;图3中的g图是“南阳占”的阳性和阴性材料的种子的粒长;图3中的h图是“南阳占”的阳性和阴性材料的种子的粒宽;图3中的i图是“南阳占”的阳性和阴性材料的种子的千粒重的表型差异。
图4:本发明的粒长基因gl3.3的表达模式与功能验证结果。附图标记说明:图4中的a图是gl3.3亚细胞定位;图4中的b图是各种组织的gus染色;图4中的c图是近等基因系中gl3.3在各个时期的表达量;图4中的d图是近等基因系的颖壳;图4中e是近等基因系纵向细胞扫描电镜图;图4中f和g分别是近等基因系纵向细胞数目和细胞长度的比较。
图5:本发明的粒长基因gl3.3的群体遗传学分析。附图标记说明:图5中a图和:图5中的b图分别是gl3.3在4726种栽培稻中的等位变异和单倍型划分;图5中的c图和图5中的d图是gl3.3不同等位型在不同亚群中的表型差异;图5中e的图是为gl3.3不同等位型的核酸多态性差异;图5中的f图和g图是gl3.3不同等位型的延伸性单倍型纯合性分析。
图6:本发明的粒长基因gl3.3的上位性效应测试。附图标记说明:图6中的a图为gs3和gl3.3在粳稻中组成的四种基因型组合的粒长分布;图6中的b图为gs3和gl3.3在重组自交系群体中的互作分析;图6中的c图是重组自交系中四种基因型组合家系的种子;图6中的d图至图6中的m图为gl3.3近等基因系各个农艺性状的表型比较。图中所有的p值都基于两尾t测验。
图7:本发明的粒长基因功能性kasp标记鉴定方法。附图标记说明:图7中的a图是kasp-gs3扩增产物荧光定量pcr读带结果;图7中的b图是kasp-gl3.3扩增产物荧光定量pcr读带结果。其中:x轴和y轴分别代表两种荧光信号的强弱。
图8:本发明的gl3.3基因在栽培稻中粒长改良的分子标记辅助选择方法应用的流程图。
具体实施方式
本发明的总体技术路线:
本发明按照以下总体技术路线来进行:
1、基因精细定位;
2、基因功能验证和效应分析(见图1)。
总体技术步骤1,基因的精细定位是按照遗传学上常用的图位克隆方法实施(具体参考实施例1)。
(1)材料的选择。选择目标性状差异明显的双亲构建遗传群体更有可能定位效应大的qtl。初步定位的群体一般为双亲杂交产生的f2群体或者重组自交系(ril)群体(方宣钧等,2000;yanoandsasaki,1997)。所述的ril群体由于接近纯系,能够重复多代多环境考察表型,能够有效而稳定的检测和验证qtl。
(2)构建近等基因系。为了实现对qtl的精细定位,需要通过与亲本之间进行回交,构建近等基因系(nil),从而消除背景的影响,使目标qtl的效应呈现为单一孟德尔因子的变化(tuinstraetal.,1997)。一般回交到bc3代或者bc4代,背景比较纯净,即可选择目标区段杂合的单株进行自交获得大量的bc3f2或bc4f2用于筛选重组单株,实现qtl的精细定位。
(3)利用初定位时双侧标记筛选重组单株,然后通过在该区间加密标记,采用染色体步移的方法来缩小定位区间。如果无法根据当代表型确认重组某些组单株的基因型,则需要对这些单株进行后代测验来确认它们的基因型。后代测验即种植某个重组单株的自交f2群体,然后考察群体的表型是否发生分离。如发生分离则确认该重组单株基因型为杂合型;如未发生分离则确认该重组单株的基因型为纯合型。利用上述方法可将qtl定位到足够小的区间,通常只包含一个候选基因。如果含有多个候选基因,可以结合生物信息学的方法和表达量分析方法进行排除。另外,通过对双亲中候选基因进行测序来寻找等位变异也是确认和验证候选基因的有效方法。
总体技术步骤2,即基因的功能验证采用常规遗传转化的方法实施(具体参考实施例2)。
遗传转化的准备工作分为两步同时进行:第一步是受体细胞的培养;第二步是目的基因的制备。
图位克隆过程中的遗传转化,一般选择受体亲本作为转化对象。在水稻中,粳稻的转化效率远高于籼稻,所以在很多情况下申请人都是选择粳稻材料作为转化对象。首先对受体材料即糙米按常规方法进行组织培养,待诱导分化得到愈伤组织后进行常规继代培养。与此同时构建表达载体和将目的片段克隆到表达载体上。接下来选择优质的愈伤组织进行预培养,这样就可以利用纯化后的载体进行农杆菌转化或其他转化。转化后的细胞首先要通过筛选,选择阳性的进行植株再生培养。对再生植株进行表型鉴定和基因型鉴定,直到获得后代分离出的单拷贝的阳性和阴性植株。通过对阳性阴性材料的随机区组种植和表型考察确定目的基因的效应。
实施例1:粒长主效基因gl3.3的定位
1.重组自交系群体的构建
利用超长粒粳稻品种南阳占(nyz)和短粒籼稻品种珍汕97(zs97)进行杂交获得f1(双亲种子形态参见图2中的a图),然后再进行自交,共获得190株f2群体。这190个单株的f2群体,可以按照“一粒传”的方法(方宣钧等,2000),进行不断自交直到f7代(图1)。
2.基因型鉴定和遗传图谱构建
基因型的鉴定方法为常规ssr分析方法。具体步骤,首选是筛选多态性ssr引物。本发明共筛选了574对ssr引物,其中rm系列的引物为452对,mrg系列的引物为118对。rm系列引物在水稻品种南阳占和珍汕97间有多态性的标记为185对,mrg系列引物在珍汕97为54对。由于多态性标记比较充足,遗传连锁图谱主要由rm系列引物构建,mrg系列引物主要用来填补缺失(gap)。利用ssr分析获得190个标记的基因型后,申请人利用mapmaker/exp3.0软件的“kosambi”作图函数绘制遗传连锁图。最终获得的遗传图谱全长1362.1cm,标记间的平均距离为7.77cm(王令强,2007)。
3.粒长qtl的初步定位
将190份rils材料于2001年和2002年5月到10月连续两年种植于湖北省武汉市华中农业大学试验田。每个家系种植2行,每行10株,株行距为16.5cm×26.4cm。每个家系考察中间的8株的粒长,8株的均值作为该家系的粒长表型值。qtl的鉴定基于基因型、表型和遗传连锁图,利用winqtlcart2.5软件(zeng,1994)的复合区间作图法,以2cm为步长在全基因组检测粒长相关的qtl。qtl检测的阈值为lod>2.5,即认为该区间存在影响粒长的qtl。qtl扫描的峰形图利用r软件绘制(ihakaandgentleman,1996)。总共检测到6个粒长的qtl,其中位于第三染色体的qgs3和qgl3.3效应最为显著(见图2中的b图)。
4.gl3.3的精细定位
为了精细定位gl3.3,申请人构建了zs97背景的bc4代近等基因系(nil)。nil-nyz具有显著高于nil-zs97的粒长(图2中的a图和图6中的g图)。在5950株bc4f2单株中,申请人鉴定到了187个在标记d3-3和rm442间发生交换的重组单株。利用这些重组单株,申请人将gl3.3定位到d3-8和d3-9之间,约为15kb的区间(见图2中的c图至图2中的e图)。这个区间包含两个开放阅读框,os03g0841800和os03g0841900。通过双亲比较测序发现,后者在双亲间无功能性变异。而os03g0841800在第三外显子后有一个g到a的变异,该变异导致南阳占(nyz)中该基因的转录产物缺少第三和第四外显子(见图2中的e图)。因此,申请人初步确认os03g0841800为gl3.3的候选基因
实施例2:gl3.3的功能验证和效应分析
1、gl3.3的功能验证
os03g0841800编码一个6281kb的开放阅读框(orf),该orf包含12个外显子和11个内含子,在粳稻“日本晴”参考基因组中注释为gsk5(kawaharaetal.,2013)。为了进一步确认os03g0841800是否就是gl3.3,申请人利用crispr/cas9系统(shanetal.,2013)在短粒籼稻珍汕97中敲除了gsk5基因。申请人获得了两种导致该基因转录提前终止的突变体(图3中的a图至图3中的e图)。两种突变型植株相比野生型zs97(非转基因)都具有显著提高的粒长和粒重(见图3中的c图至e图3中的图)。另一方面,申请人在“nyz”中互补转化了“zs97”中的gl3.3的等位型基因(hieietal.,1994)。分析结果表明,转基因阴性植株具有比阳性植株显著更长的粒形和更高的千粒重(见图3中的f图至图3中的i图)。这些结果表明gsk5就是gl3.3并且该基因为一个负调控水稻粒长和千粒重的基因。
2.gl3.3的表达模式和功能
为了定位gl3.3在细胞中表达的位置,申请人构建了camv35s启动子驱动的gl3.3-黄色荧光蛋白融合(yfp)表达载体(hajdukiewiczetal.,1994)。转化水稻原生质体(xueetal.,2008)的结果表明gl3.3-yfp定位在细胞核,这表明gl3.3在细胞核内表达(图4中的a图)。申请人利用荧光定量pcr(rt-pcr)(livaketal.,2001)和常规gus染色方法来分析gl3.3基因的时空表达特性。gus染色结果表明,gl3.3基因在被试验的水稻品种的胚乳、幼苗、茎和花器官中大量表达(图4中的b图)。nil-nyz和nil-zs97的rt-pcr,结果表明gl3.3基因的mrna在水稻品种的幼苗、幼穗、胚乳、茎和根中表达(图4中的c图)。因此推定gl3.3基因是一个在水稻各个组织时期中组成型表达的基因。由于nil-nyz的颖壳在受精前就比nil-zs97长,所以申请人利用扫描电镜分析了两个nils的颖壳外表面来解析颖壳长度差异的具体原因(见图4中的d图至图4中的g图)。发现nil-nyz的纵向细胞数目显著少于nil-zs97的纵向细胞数目;而nil-nyz的纵向细胞长度要显著大于nil-zs97纵向细胞的长度。因此,nil-nyz颖壳纵向细胞数目~10%的降低和细胞长度~20%的增加导致了整体颖壳长度的增加。
3.gl3.3的等位型与人工选择
由于gl3.3是一个负调控粒长的基因,因此南阳占(nyz)含有一种突变型的gl3.3。在4726份栽培稻材料中,nyz所含有的这种等位型的比例为1%,并且这种等位型只出现在粳稻中,因此申请人称其为gl3.3jap(图5中的a图,图5中的b图)。在粳稻中,具有gl3.3jap这种等位型的品种相比野生型的品种具有显著较长的粒形(图5中的d图)。通过进一步分析gl3.3的自然变异,申请人发现了另一种功能性的变异:即,在第6外显子上一个由c到a的转换,导致氨基酸序列从组氨酸到谷氨酰胺的变化。这种等位型在4726份栽培稻中的比例为5%,并且主要存在于澳洲稻中,因此申请人将其称为gl3.3aus(图5中的a图,图5中的b图)。在澳洲稻中,具有这种等位型的品种相比野生型的品种具有显著较长的粒形(图5中的c图)。利用dnasp软件(rozas等,2003)计算两种突变型和野生型的核酸多态性,申请人发现两种突变型都具有显著较低的多态性(图5中的e图),可能在育种的过程中受到过人工选择。申请人进一步研究在澳洲稻和粳稻中gl3.3野生型和突变型的延伸性单倍型纯合性,发现野生型的连锁不平衡衰退很快,而突变型具有衰退的比较慢,表明突变型受到了人工选择(图5中的f图,图5中的g图)。
4.gl3.3与gs3上位性互作形成超长粒形
虽然gl3.3aus和gl3.3jap都影响水稻粒长的自然变异,但是相比gl3.3在作图群体中的加性效应(a=0.35mm),gl3.3aus的效应偏小(a=0.24mm)而gl3.3jap的效应显著偏大(a=1.36mm)(见图5中的c图,图5中的d图)。由于gl3.3在粳稻中的效应过大,超过了已知的任何粒形的主效基因,申请人推测可能gl3.3在粳稻中和某个粒长基因存在远程的连锁不平衡。通过分析粒形主效基因gs3的基因型,申请人发现92%具有gl3.3jap等位型的粳稻品种也具有gs3等位型,表明gs3在粳稻中与gl3.3jap远程连锁(见图6中的a图)。前人研究表明gs3在育种过程中受到人工选择(takanokai等,2009),因此gl3.3jap与gs3一起在超长粒粳稻品种中受到选择。其实考虑了gs3的效应,gl3.3在粳稻中的效应(a=0.45mm)依然大于作图群体中的效应(图6中的a图),因此申请人怀疑gl3.3可能与gs3存在互作。通过重新分析重组自交系群体中gs3和gl3.3的效应,申请人发现这两个基因存在明显的上位性互作(图6中的b图,图6中的c图)。这种互作使得gl3.3在gs3无功能的背景下效应的粳稻,即gs3gl3.3会形成一种超长的粒形。gl3.3在“nyz”互补转基因材料中的效应大于“zs97”敲除转基因材料的效应也进一步说明了这一点(见图3)。
实施例3:gl3.3在水稻粒长改良中的应用
本发明定位到的gl3.3为一个稀有且主效的粒长基因,并且能够和gs3互作而发挥更强的功能形成超长粒表型,能有效地提高现有栽培稻品种的粒长性状。可以在育种过程中利用gl3.3的分子标记进行辅助选择能够减少种植面积、简化育种流程和节约人力物力,以提高育种效率。因此将水稻种质材料(例如本实验中的品种“南阳占”)中具有gl3.3无功能等位型(gl3.3aus和gl3.3jap)的材料用于水稻粒长性状的改良,具有十分广泛的应用前景。
本实施例的技术路线可参见图7所述的方法,图7是申请人设计的利用gl3.3优势等位基因改良水稻粒长的总体技术流程。该技术流程包括:首先选择生产上的短粒或者中长粒骨干水稻品种材料,即简称的“骨干材料”(这些“骨干材料”都是可以得到的,包括在生产上具有优良性状而粒长有待改良的常规稻品种,以及不育系和恢复系,鉴定上述骨干材料的方法都是现有报道的方法,不存在探索性劳动,获得不育系和恢复系的方法也都是现有报道的方法和本领域的公知常识),与具备优势基因型组合gs3gl3.3的栽培稻材料(例如“南阳占”,但不限于此)进行杂交,可分别得到短粒骨干材料杂交种f1代(或称f1a)和中长粒骨干材料f1代(或称f1b)。将f1a与短粒骨干材料连续回交5代(这是领域的公知方法)即可获得近等基因系,每代利用分子标记引物kasp-gs3和kasp-gl3.3选择这两个位点的真杂种单株(图7),直到得到bc5f1a。相对应的,f1b与中长粒骨干材料连续回交5代获得近等基因系,每代都利用分子标记引物kasp-gs3和kasp-gl3.3选择这两个位点的真杂种单株,直到得到bc5f1b。短粒骨干材料和中长粒骨干材料的近等基因系bc5f1a和bc5f1b分别自交,获得bc5f2a和bc5f2b。再利用分子标记引物kasp-gs3和kasp-gl3.3鉴定出bc5f2a中具有纯合gs3gl3.3基因型组合的单株,即为柱头外露率改良后的短粒骨干水稻材料。再利用分子标记引物kasp-gs3和kasp-gl3.3鉴定出bc5f2b中具有纯合gl3.3基因型组合的单株,即为柱头外露率改良后的中长粒水稻骨干材料。改良后的水稻短粒骨干材料和水稻中长粒骨干材料粒形已经成为超长粒,故可称之为水稻超长粒骨干材料。
主要参考文献
1.方宣钧等编.作物dna标记辅助育种[m].北京:科学出版社,2000;
2.孔宪琴等,我国稻米产业形势与企业发展策略[j].中国稻米,2013,19(6):32-34;
3.王令强.分子标记遗传图谱构建和稻米品质性状的遗传分析[d].华中农业大学,2007;
4.于红燕等,我国水稻产业发展现状、趋势及对策[j].农村经济与科技,2016,27(9):7-9;
5.duanp,xuj,zengd,etal.naturalvariationinthepromoterofgse5contributestograinsizediversityinrice[j].molecularplant,2017,10(5):685-694;
6.hajdukiewiczp,svabz,maligap.thesmall,versatileppzpfamilyofagrobacteriumbinaryvectorsforplanttransformation.[j].plantmolecularbiology,1994,25(6):989-94;
7.hieiy,ohtas,komarit,etal.efficienttransformationofrice(oryzasativa,l.)mediatedbyagrobacterium,andsequenceanalysisoftheboundariesofthet-dna[j].plantjournal,1994,6(2):271;
8.huj,wangy,fangy,etal.ararealleleofgs2enhancesgrainsizeandgrainyieldinrice[j].molecularplant,2015,8(10):1455;
9.kawaharay,delbm,hamiltonjp,etal.improvementoftheoryzasativanipponbarereferencegenomeusingnextgenerationsequenceandopticalmapdata[j].rice,2013,6(1):4;
10.livakkj,schmittgentd.analysisofrelativegeneexpressiondatausingreal-timequantitativepcrandthe2(-deltadeltac(t))method[j].methods,2001,25(4):402-408;
11.liy,fanc,xingy,etal.naturalvariationings5playsanimportantroleinregulatinggrainsizeandyieldinrice.[j].naturegenetics,2011,43(12):1266-9;
12.liy,fanc,xingy,etal.chalk5encodesavacuolarh(+)-translocatingpyrophosphataseinfluencinggrainchalkinessinrice.[j].naturegenetics,2014,46(6):398;
13.qip,linys,songxj,etal.thenovelquantitativetraitlocusgl3.1controlsricegrainsizeandyieldbyregulatingcyclin-t1;3[j].cellresearch,2012,22(12):1666-80;
14.rossihaka,robertgentleman.r:alanguagefordataanalysisandgraphics[j].journalofcomputationalandgraphicalstatistics,1996,5(3):299-314;
15.rozasj,sánchezdelbarriojc,messeguerx,etal.dnasp,dnapolymorphismanalysesbythecoalescentandothermethods.[j].bioinformatics,2003,19(18):2496-7;
16.shanq,wangy,lij,etal.targetedgenomemodificationofcropplantsusingacrispr-cassystem.[j].naturebiotechnology,2013,31(8):686;
17.songxj,huangw,shim,etal.aqtlforricegrainwidthandweightencodesapreviouslyunknownring-typee3ubiquitinligase.[j].naturegenetics,2007,39(5):623-30;
takanokain,jiangh,kubot,etal.evolutionaryhistoryofgs3,ageneconferringgrainlengthinrice[j].genetics,2009,182(4):1323;
18.tuinstram,ejetag,goldsbroughp.heterogeneousinbredfamily(hif)analysis:amethodfordevelopingnear-isogeniclinesthatdifferatquantitativetraitloci[j].theorapplgenet,1997,95:1005-1011;
19.wangs,wuk,yuanq,etal.controlofgrainsize,shapeandqualitybyosspl16inrice[j].naturegenetics,2012,44(8):950-954;
20.wangs,lis,liuq,etal.theosspl16—gw7regulatorymoduledeterminesgrainshapeandsimultaneouslyimprovesriceyieldandgrainquality[j].naturegenetics,2015,47(8):949-54;
21.xuew,xingy,wengx,zhaoy,etal.naturalvariationinghd7isanimportantregulatorofheadingdateandyieldpotentialinrice[j].naturegenetics,2008,40:761-767;
22.yanom,sasakit.geneticandmoleculardissectionofquantitativetraitsinrice[j].plantmolbiol,1997,35:145-153;
23.zhouh,lip,xiew,etal.genome-wideassociationanalysesrevealthegeneticbasisofstigmaexsertioninrice[j].molecularplant,2017,10(4):634-644;
24.zengzb.zengzb.precisemappingofquantitativetraitloci.genetics136:1457-1468[j].genetics,1994,136(4):1457-1468。
序列表
<110>华中农业大学
<120>超长粒水稻的培育方法
<141>2018-02-28
<160>10
<170>siposequencelisting1.0
<210>1
<211>1275
<212>dna
<213>水稻(oryzasativa)
<220>
<221>gene
<222>(1)..(1275)
<220>
<221>cds
<222>(1)..(1275)
<400>1
atggcctattctggactaaggcatggcggcgttgggagctcctctcgg48
metalatyrserglyleuarghisglyglyvalglyserserserarg
151015
ccgggacatggattcaagggcccggctagttcggttgaatgtctagga96
proglyhisglyphelysglyproalaserservalglucysleugly
202530
agggagatgctggaaatgcaattgagggattccaaaccagatgttggt144
argglumetleuglumetglnleuargaspserlysproaspvalgly
354045
gatgaaaagaacactgaacgagatgtagtcgacggctctagcgctgaa192
aspglulysasnthrgluargaspvalvalaspglyserseralaglu
505560
gctggtcacataatagctactacgatccgtggccgaaatggtctacct240
alaglyhisileilealathrthrileargglyargasnglyleupro
65707580
aaacagtctgtcacttatattgctgagcatgtggtcggaactggttct288
lysglnservalthrtyrilealagluhisvalvalglythrglyser
859095
tttggggttgtatatcaggccaaatgtcgagaaacaggagaaattgtt336
pheglyvalvaltyrglnalalyscysarggluthrglygluileval
100105110
gccattaaaaaggttcttcaagataaacgttacaagaacagggaattg384
alailelyslysvalleuglnasplysargtyrlysasnarggluleu
115120125
caaattatgcatatgctggatcatcctaatattgttggtcttaagcat432
glnilemethismetleuasphisproasnilevalglyleulyshis
130135140
tacttcttctcaaccactgaaagggatgaactttacctcaatcttgtc480
tyrphepheserthrthrgluargaspgluleutyrleuasnleuval
145150155160
cttgagtatgttccagagacagtaaaccgaattgcgagacagtacagc528
leuglutyrvalprogluthrvalasnargilealaargglntyrser
165170175
agaatgaatcaacgggtgcccctcatttatgtcaaattatacacttat576
argmetasnglnargvalproleuiletyrvallysleutyrthrtyr
180185190
cagatatgccgagcacttgcttacatacataactgtgttggcatctgc624
glnilecysargalaleualatyrilehisasncysvalglyilecys
195200205
caccgtgatataaaaccacagaatgttcttgttaacccacatacacac672
hisargaspilelysproglnasnvalleuvalasnprohisthrhis
210215220
cagctcaaaatatgtgattttggcagtgcaaaagttttggtcaaagga720
glnleulysilecysasppheglyseralalysvalleuvallysgly
225230235240
gagccaaatatctcctacatatgctcaagatattacagggcaccagag768
gluproasnilesertyrilecysserargtyrtyrargalaproglu
245250255
ctcatatttggtgcaactgaatatactactgccattgatttgtggtct816
leuilepheglyalathrglutyrthrthralaileaspleutrpser
260265270
acaggctgtgtcatggcagagcttctgcttggacagcctctgttccct864
thrglycysvalmetalagluleuleuleuglyglnproleuphepro
275280285
ggggaaagtggagttgatcagctggtcgagattatcaaggttttgggt912
glygluserglyvalaspglnleuvalgluileilelysvalleugly
290295300
actccaacaagggaagagatcaagtgcatgaatccgaactacacggag960
thrprothrargglugluilelyscysmetasnproasntyrthrglu
305310315320
ttcaaattccctcaaattaaggctcatccatggcacaaggttttccaa1008
phelyspheproglnilelysalahisprotrphislysvalphegln
325330335
aaaaggcttccacctgaagcagtggaccttgtaagcaggtttctgcaa1056
lysargleuproproglualavalaspleuvalserargpheleugln
340345350
tactcaccaaatcttcggtgcaccgctatggaagcctgcatgcaccct1104
tyrserproasnleuargcysthralametglualacysmethispro
355360365
ttctttgatgagttgagagatccaaacactcgtctacctaatgggcgt1152
phepheaspgluleuargaspproasnthrargleuproasnglyarg
370375380
cctcttcctcctctcttcaacttcagaacacaagagctaaatggcatc1200
proleuproproleupheasnpheargthrglngluleuasnglyile
385390395400
cctccagaagccatcgaacgtctggttcccgagcatgcgagaaggcag1248
proproglualailegluargleuvalprogluhisalaargarggln
405410415
agtttgttcatggcgctgcgcacctag1275
serleuphemetalaleuargthr
420
<210>2
<211>424
<212>prt
<213>水稻(oryzasativa)
<400>2
metalatyrserglyleuarghisglyglyvalglyserserserarg
151015
proglyhisglyphelysglyproalaserservalglucysleugly
202530
argglumetleuglumetglnleuargaspserlysproaspvalgly
354045
aspglulysasnthrgluargaspvalvalaspglyserseralaglu
505560
alaglyhisileilealathrthrileargglyargasnglyleupro
65707580
lysglnservalthrtyrilealagluhisvalvalglythrglyser
859095
pheglyvalvaltyrglnalalyscysarggluthrglygluileval
100105110
alailelyslysvalleuglnasplysargtyrlysasnarggluleu
115120125
glnilemethismetleuasphisproasnilevalglyleulyshis
130135140
tyrphepheserthrthrgluargaspgluleutyrleuasnleuval
145150155160
leuglutyrvalprogluthrvalasnargilealaargglntyrser
165170175
argmetasnglnargvalproleuiletyrvallysleutyrthrtyr
180185190
glnilecysargalaleualatyrilehisasncysvalglyilecys
195200205
hisargaspilelysproglnasnvalleuvalasnprohisthrhis
210215220
glnleulysilecysasppheglyseralalysvalleuvallysgly
225230235240
gluproasnilesertyrilecysserargtyrtyrargalaproglu
245250255
leuilepheglyalathrglutyrthrthralaileaspleutrpser
260265270
thrglycysvalmetalagluleuleuleuglyglnproleuphepro
275280285
glygluserglyvalaspglnleuvalgluileilelysvalleugly
290295300
thrprothrargglugluilelyscysmetasnproasntyrthrglu
305310315320
phelyspheproglnilelysalahisprotrphislysvalphegln
325330335
lysargleuproproglualavalaspleuvalserargpheleugln
340345350
tyrserproasnleuargcysthralametglualacysmethispro
355360365
phepheaspgluleuargaspproasnthrargleuproasnglyarg
370375380
proleuproproleupheasnpheargthrglngluleuasnglyile
385390395400
proproglualailegluargleuvalprogluhisalaargarggln
405410415
serleuphemetalaleuargthr
420
<210>3
<211>699
<212>dna
<213>水稻(oryzasativa)
<220>
<221>gene
<222>(1)..(699)
<220>
<221>cds
<222>(1)..(699)
<400>3
atggcaatggcggcggcgccccggcccaagtcgccgccggcgccgccc48
metalametalaalaalaproargprolysserproproalapropro
151015
gacccatgcggccgccaccgcctccagctcgccgtcgacgcgctccac96
aspprocysglyarghisargleuglnleualavalaspalaleuhis
202530
cgcgagatcggattcctcgagggtgaaataaattcaatcgaagggatc144
arggluileglypheleugluglygluileasnserilegluglyile
354045
cacgctgcctccagatgctgcagagaggttgacgaattcatcggaaga192
hisalaalaserargcyscysarggluvalaspglupheileglyarg
505560
actcctgatccattcataacgatttcatcggagaagcgaagtcatgat240
thrproasppropheilethrileserserglulysargserhisasp
65707580
cattctcaccacttcttgaagaagtttcgctgtttgtgcagagcaagt288
hisserhishispheleulyslyspheargcysleucysargalaser
859095
gcgtgctgcctcagctacctctcctggatctgctgctgcagcagcgcc336
alacyscysleusertyrleusertrpilecyscyscysserserala
100105110
gccggcggctgctcatcctcctcctcctcctccttcaacctcaagagg384
alaglyglycysserserserserserserserpheasnleulysarg
115120125
ccgagctgctgctgcaactgcaactgcaactgctgctcctcctcctcc432
prosercyscyscysasncysasncysasncyscysserserserser
130135140
tcctcatgtggggcggcgttaacgaagagtccgtgtcgctgccgccgc480
sersercysglyalaalaleuthrlysserprocysargcysargarg
145150155160
cgcagctgctgctgccgtcgctgctgctgcggcggcgtcggcgtccgc528
argsercyscyscysargargcyscyscysglyglyvalglyvalarg
165170175
gcgtgcgcgagctgcagctgctccccgccgtgcgcgtgctgcgcgccg576
alacysalasercyssercysserproprocysalacyscysalapro
180185190
ccgtgcgcgggatgctcgtgccgctgcacctgcccgtgcccgtgcccc624
procysalaglycyssercysargcysthrcysprocysprocyspro
195200205
ggcggctgctcctgcgcgtgcccggcgtgcaggtgctgctgcggcgtc672
glyglycyssercysalacysproalacysargcyscyscysglyval
210215220
cctcgttgctgccccccctgcttgtga699
proargcyscysproprocysleu
225230
<210>4
<211>232
<212>prt
<213>水稻(oryzasativa)
<400>4
metalametalaalaalaproargprolysserproproalapropro
151015
aspprocysglyarghisargleuglnleualavalaspalaleuhis
202530
arggluileglypheleugluglygluileasnserilegluglyile
354045
hisalaalaserargcyscysarggluvalaspglupheileglyarg
505560
thrproasppropheilethrileserserglulysargserhisasp
65707580
hisserhishispheleulyslyspheargcysleucysargalaser
859095
alacyscysleusertyrleusertrpilecyscyscysserserala
100105110
alaglyglycysserserserserserserserpheasnleulysarg
115120125
prosercyscyscysasncysasncysasncyscysserserserser
130135140
sersercysglyalaalaleuthrlysserprocysargcysargarg
145150155160
argsercyscyscysargargcyscyscysglyglyvalglyvalarg
165170175
alacysalasercyssercysserproprocysalacyscysalapro
180185190
procysalaglycyssercysargcysthrcysprocysprocyspro
195200205
glyglycyssercysalacysproalacysargcyscyscysglyval
210215220
proargcyscysproprocysleu
225230
<210>5
<211>41
<212>dna
<213>水稻(oryzasativa)
<220>
<221>primer_bind
<222>(1)..(41)
<400>5
gaaggtgaccaagttcatgctacgctgcctccagatgctgc41
<210>6
<211>41
<212>dna
<213>水稻(oryzasativa)
<220>
<221>primer_bind
<222>(1)..(41)
<400>6
gaaggtcggagtcaacggattacgctgcctccagatgctga41
<210>7
<211>20
<212>dna
<213>水稻(oryzasativa)
<220>
<221>primer_bind
<222>(1)..(20)
<400>7
gtagtacaaaaagaaacagc20
<210>8
<211>41
<212>dna
<213>水稻(oryzasativa)
<220>
<221>primer_bind
<222>(1)..(41)
<400>8
gaaggtgaccaagttcatgctttttggggttgtatatcagg41
<210>9
<211>41
<212>dna
<213>水稻(oryzasativa)
<220>
<221>primer_bind
<222>(1)..(41)
<400>9
gaaggtcggagtcaacggattttttggggttgtatatcaga41
<210>10
<211>19
<212>dna
<213>水稻(oryzasativa)
<220>
<221>primer_bind
<222>(1)..(19)
<400>10
cacatagagaaatttttac19
- 还没有人留言评论。精彩留言会获得点赞!