RNA信号连接器、靶mRNA翻译调控方法、逻辑门及应用与流程
本发明涉及基因工程和分子生物学领域,尤其涉及rna信号连接器、靶mrna翻译调控方法、逻辑门及其应用。
背景技术
感受胞外信号后调节相关基因表达是细胞的基本功能。细胞通过天然的信号网络实现复杂的生理功能,如细胞存活、行为及识别。近年来合成生物学发展迅速,而其重要目标之一就是在治疗及生物技术应用层面上重塑细胞信号网络。合成元件被广泛应用到构建各种新颖的基因调控线路中,例如遗传开关、数字化逻辑线路、人工信号通路及反馈回路。近期,本发明人构造了一类全新的基因调控装置——“crispr信号传导器”(参见本发明人的中国专利申请cn106318973a,名称“一种基于crispr-cas9的基因调控装置及基因调控方法”),其可感测和响应指定的细胞信号,并通crispr干扰或激活调节特定内源基因转录。该装置的作用是整合任何指定生物分子的相关信息和重塑细胞不同表型。
然而,与既往报道的遗传装置一样,该系统仍需要转入编码大蛋白(如dcas9)的外源性基因,这进一步增加了系统的复杂性。从应用的观点来看,使用精简的rna装置或许比使用含有编码大蛋白装置更加符合转基因递送技术的要求。
技术实现要素:
本发明的目的是提供结构简单而又有效的遗传装置及其应用。
因此,在第一方面,本发明提供一种rna信号连接器,该rna信号连接器为一段寡核苷酸序列,包括一个反义结构域和两个拷贝的适体结构域,该反义结构域与第一拷贝的适体结构域连接,第一拷贝的适体结构域通过一个接头序列与第二拷贝的适体结构域连接,该反义结构域与靶mrna序列互补,该适体结构域与作为输入信号的配体结合。
具体地,该反义结构域与该靶mrna的5’-utr或编码序列互补。
在一些实施方案中,该配体为茶碱、四环素、β-连环蛋白、vegf、opn或nf-kb,该适体结构域相应地为茶碱、四环素、β-连环蛋白、vegf、opn或nf-kb适体结构域。
进一步地,本发明提供一种rna信号连接器,该rna信号连接器为一段寡核苷酸序列,包括一个反义结构域和两个拷贝的适体结构域,还包括一个核酸开关,该反义结构域与该核酸开关连接,该核酸开关再通过第一接头序列与第一拷贝的适体结构域连接,第一拷贝的适体结构域通过第二接头序列与第二拷贝的适体结构域连接,该反义结构域与靶mrna序列互补,该适体结构域与作为输入信号的配体结合。
在一些实施方案中,该配体为茶碱、四环素、β-连环蛋白、vegf、opn或nf-kb,该核酸开关相应地为茶碱、四环素、β-连环蛋白、vegf、opn或nf-kb核酸开关,该适体结构域为eif4g适体结构域。
在具体的实施方案中,前述的rna信号连接器的cdna如下表1-17和序列表seqidno:1-40中的任一序列所示。下表中的序列代号与序列表的序列号对应,例如序列代号r1对应于序列号seqidno:1,以此类推。
表1.茶碱诱导的靶向并抑制海肾荧光素酶mrna翻译的信号连接器的cdna序列。每个序列由一个互补序列(粗体所示)、两个拷贝的茶碱适体和一个接头序列(下划线所示)组成。
表2.四环素诱导的靶向并抑制海肾荧光素酶mrna翻译的信号连接器的cdna序列。每个序列由一个互补序列(粗体所示)、两个拷贝的四环素适体和一个接头序列(下划线所示)组成。
表3.四环素诱导的靶向并抑制vegfmrna翻译的信号连接器的cdna序列。每个序列由一个互补序列(粗体所示)、两个拷贝的四环素适体和一个接头序列(下划线所示)组成。
表4.靶向并增强海肾荧光素酶mrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的eif4g适体和一个接头序列(下划线所示)组成
表5.茶碱诱导的靶向并增强海肾荧光素酶mrna翻译的信号连接器的cdna序列。每个序列由一个互补序列(粗体所示)、一个拷贝的茶碱核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表6.四环素诱导的靶向并增强海肾荧光素酶mrna翻译的信号连接器的cdna序列。每个序列由一个互补序列(粗体所示)、一个拷贝的四环素核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表7.β-连环蛋白诱导的靶向并抑制c-mycmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的β-连环蛋白适体和一个接头序列(下划线所示)组成
表8.β-连环蛋白诱导的靶向并增强海肾荧光素酶mrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、一个拷贝的β-连环蛋白核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表9.vegf诱导的靶向并增强opnmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、一个拷贝的vegf核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表10.opn诱导的靶向并增强vegfmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、一个拷贝的opn核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表11.vegf诱导的靶向并抑制opnmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的vegf适体和一个接头序列(下划线所示)组成
表12.茶碱诱导的靶向并抑制c-mycmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的茶碱适体和一个接头序列(下划线所示)组成
表13.茶碱诱导的靶向并抑制bcl2mrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的茶碱适体和一个接头序列(下划线所示)组成
表14.nf-kb诱导的靶向并增强baxmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、一个拷贝的nf-kb核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表15.nf-kb诱导的靶向并增强p21mrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、一个拷贝的nf-kb核酸开关(斜体所示)、两个拷贝的eif4g适体和两个接头序列(下划线所示)组成
表16.nf-kb诱导的靶向并抑制bcl2mrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的nf-kb适体和一个接头序列(下划线所示)组成
表17.nf-kb诱导的靶向并抑制c-mycmrna翻译的信号连接器的cdna序列。该序列由一个互补序列(粗体所示)、两个拷贝的nf-kb适体和一个接头序列(下划线所示)组成
在第二方面,本发明提供一种调控细胞中的靶mrna的翻译的方法。该方法包括将第一方面的rna信号连接器导入该细胞中,并在配体存在下使该rna信号连接器抑制或激活该靶mrna的翻译。
具体地,该配体以浓度依赖性方式使该rna信号连接器抑制或激活该靶mrna的翻译。
在第三方面,本发明提供一种逻辑门。该逻辑门由2个第一方面的rna信号连接器组成。
具体地,该逻辑门为非门、与门、与非门、或门、或非门、异或门和异或非门。
在第四方面,本发明提供第一方面的rna信号连接器或第三方面的逻辑门重新连接细胞信号通路并创建反馈回路的用途。
在第五方面,本发明提供第一方面的rna信号连接器或第三方面的逻辑门沉默存活基因的表达并抑制癌细胞的生长的用途。
在第六方面,本发明提供第一方面的rna信号连接器或第三方面的逻辑门将致癌信号转化为抑癌信号的用途。
本发明的有益效果是:本发明的rna信号连接器是一种遗传装置,为一段寡核苷酸序列,包括反义结构域和适体结构域,任选包括核酸开关,不含编码大蛋白(如dcas9)的外源性基因,结构简单。通过提供细胞外源性或内源性的配体作为输入信号,该rna信号连接器可以调控细胞中的靶mrna的翻译。该rna信号连接器还可以用于构建生物逻辑门,建立输入和输出信号之间的联系,在重新连接细胞信号通路并创建反馈回路、沉默存活基因的表达并抑制癌细胞的生长以及将致癌信号转化为抑癌信号等方面具有潜在的应用。
附图说明
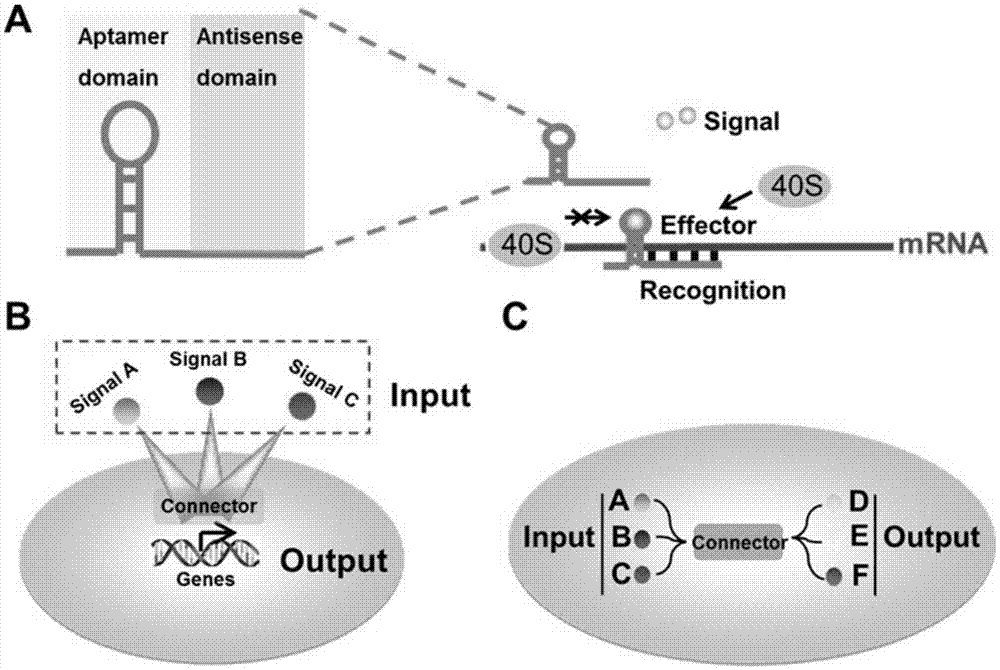
图1显示信号连接器的设计和构造,用于构造信号节点之间的连接。(a)信号连接器的结构域组成。该装置使用反义域识别目标基因的mrna和配体域来响应不同的信号,进而控制目标基因的翻译。(b)通过这种模块化的方法,可以利用特定外源信号控制细胞基因表达。(c)信号连接器可用于构建细胞输入与输出之间的联系。
图2显示信号连接器有效地抑制或激活基因表达。(a)信号连接器的设计,阻止目标基因的翻译。该装置被设计用来靶向目标mrna的5'-utr。在没有配体的情况下,对40s核糖体亚基进行扫描,直到起始密码子开始翻译。在配体的存在中,配体-适体复合物扰乱了核糖体的扫描并阻断了翻译的起始。(b)在与靶mrna的蛋白质编码区结合时,它阻断核糖体介导的翻译延伸。(c)不同的信号连接器被设计成针对海肾荧光素酶信使mrna的不同区域。(d)信号连接器仅在1mm茶碱存在下抑制相对荧光素酶活性。pgpu6/gfp/neo空载作为阴性对照。(e)信号连接器(r1、r2和r3)在0、250、500、750或1000μm的茶碱中生长。这种茶碱的添加以剂量依赖性的方式抑制了海肾酶信使rna的翻译。(f)在hek293细胞中,通过四种不同的对四环素应答的信号连接器,来抑制荧光素酶。(g)用于增强目标基因翻译的信号连接器的设计。信号连接器的翻译激活是由于激活了eif4g的起始因子复合物的形成。在没有茶碱/四环素的情况下,反义域不能与目标基因的信使rna结合。在茶碱/四环素的存在下,配体茎形成,反义域会与它的目标结合。(h)通过信号连接器增加了相对荧光酶的活性。(i)添加了茶碱/四环素增加了对海肾荧光素酶的翻译。pgpu6/gfp/neo,空载。nc,阴性对照,含有两个重复元件,在人类基因组中没有靶点。报告的数据来自至少三个实验的平均值±sd。
图3显示基于信号集成的逻辑门是由信号连接器构造的。(a)在每一个逻辑门中,信号连接器集成输入信号的茶碱和四环素,并产生荧光素酶的输出。使用信号连接器执行非门(b)、与门(c)、与非门(d)、或门(e)、或非门(f)、异或门(g)和异或非门(h)函数。
图4显示信号连接器有效地重新连接并创建信号通路和反馈回路。(a)信号连接器设计用来重新连接信号通路的机制。(b)在hek293细胞中使用实时qpcr对c-myc信使rna的相对表达水平进行了评估。在对ltd4刺激反应的细胞中增加了c-mycmrna的水平。(c)原癌基因的柱状图。(d)利用信号连接的机制创建信号通路。(e)响应β-连环蛋白(β-catenin)的信号连接器的设计。该装置释放反义区,仅在β-连环蛋白的存在下靶向mrna。(f)在对ltd4刺激反应的hek293细胞中对海肾荧光素酶的相对活性进行了评估。(g)设计的模型和实验结果,说明了信号连接器在构造opn-vegf正反馈回路中的假定作用。(h)设计的模型和实验结果,说明了信号连接器在构造opn-vegf负反馈回路中的假定作用。nc,阴性对照,含有两个重复元件,在人类基因组中没有靶点。报告的数据来自至少三个实验的平均值±sd。实验至少重复三次。
图5显示信号连接器特别抑制了存活基因的表达,抑制了目标癌细胞的细胞生长。(a)信号连接器的机制,目的是选择性地杀死癌细胞,这些细胞通过激活的tertp和配体来控制细胞的存活。(b)遗传和门的示意图。htert的启动子和配体(1000μm的茶碱)是线路的两个输入。(c)两种不同的信号连接器被设计用来靶向人类c-myc信使rna和bcl2mrna的5-utrs。(d和e)对膀胱癌细胞(t24和5637)和正常成纤维细胞的目标蛋白表达进行定量分析。nc,有两个重复元件的阴性对照,在人类基因组中没有目标。(f)细胞的生长是用cck-8测定的时间间隔来测量的。方差分析用于比较细胞生长曲线。报告的值是三个独立实验的平均值±sd。(g)细胞的凋亡是用hoechst染色测定。报告的值是三个独立实验的平均值±sd。
图6显示信号连接器在对致癌信号的反应中同时激活和抑制细胞基因,并将致瘤信号转化为抗肿瘤的途径。(a)癌基因信号nf-kb被重定向到激活两个肿瘤抑制因子bax和p21,并通过癌细胞中的信号连接器抑制两个致癌基因bcl2和c-myc。(b)在细胞接种后的48小时内,以t24细胞为标准,在t24细胞中测定出bax、bcl2、c-myc和p21的相对表达水平。报告的值是三个独立实验的平均值±sd。(c和d)注射后20天,在信号连接器组中形成的肿瘤相对于阴性对照组而言,要小得多。(e)在注射t24细胞后,每隔5天就计算一次肿瘤体积。柱条表示标准偏差(sd)。(f)肿瘤重量变化。**p<0.01。
具体实施方式
以下通过实施例的方式对本发明进行进一步的详细描述。应当理解,这些描述仅出于说明本发明的目的,并不意在以任何方式限制本发明。
除非另外定义,本文使用的所有技术和科学术语与本公开所属领域技术人员通常理解的含义相同。如果有冲突,以本说明的定义为准。本文中使用的材料、方法和示例仅作说明用途,无意作为限制,除非特别指明。
i.材料与方法
1.质粒构建
设计了针对海肾荧光素酶/c-myc/opn/vegf/bcl2/bax/p21mrna信号连接器的cdna序列,并用在线软件对其rna二级结构进行了分析。筛选出表达反义区并保持适体二级结构的rna,并合成相应的cdna序列分别插入到pgpu6/和bamhi/bbsⅰ限制性位点的gfp/neo载体中。采用类似的方法,设计、合成了以c-myc/bcl2mrna为目标的核糖体侧翼信号连接器的cdna序列,分别在sali/bamhi的限制位点上被插入到htert-neo-bam载体上。为了构建质粒pcdna3-0-vegf和pcdna3-opn,将表达缺失n端信号肽的opn/vegf截短型的cdna序列分别插入到pcdna3.0和bamhi/ecori中。为了构建真核表达质粒shrna-nc和shrna-vegf,将合成的shrna序列分别插入到用bamhi/bbs消化的pgpu6/gfp/neo中。
2.细胞株与细胞培养
t24、5637和hek-293细胞是由本实验室从美国模式培养物保藏中心(atcc)购买,并在含10%胎牛血清(invitrogen)的dmem培养基中在添加5%co2存在下培养。从表皮中提取的正常人类原发性成纤维细胞在同一培养基中进行原代培养。如下所述从这些细胞系产生稳定的细胞系。所有细胞系都经验证不含支原体。
稳定表达海肾荧光素酶的hek-293细胞是通过用pcdna3/rluc/neo转染细胞并用g418选择阳性克隆获得的。具体地讲,在6孔板中进行稳定选择,该板中每孔接种约2×105个hek-293细胞,使用lipofectamine2000transfectionreagent(invitrogen)按照生产商说明书转染2mg的线性化质粒。转染后48小时,用胰蛋白酶消化细胞单层,转移到t25烧瓶或100mm直径的培养皿中。通过在含有50mgg418/ml的完全培养基中生长,选择稳定转染子的混合集落。将这些多克隆细胞系进行扩增,然后通过荧光素酶报道基因测定法进行验证。
在转染pgpu6/gfp/puro载体后,选择稳定表达信号连接器或阴性对照的hek-293细胞。共表达多个信号连接器的hek-293/t24细胞是通过稳定转染单一pgpu6/gfp/puro载体构建的,该载体由单一u6启动子驱动而同时产生这些装置。具体地讲,将细胞接种在六孔板中,使用lipofectamine2000transfectionreagent(invitrogen)按照生产商说明书转染2mg的线性化质粒。转染后,将细胞在补加有4mg/ml的嘌呤霉素的培养基中生长大约14天,以选择稳定细胞系的混合集落。通过gfp表达验证这些多克隆细胞。使用倒置式荧光显微镜直接观察培养板中的荧光细胞。
3.荧光素酶报告实验
将稳定表达海肾荧光素酶的hek-293细胞接种于6孔板(5x105/孔)中。转染后48小时,弃去培养基,将细胞在500ml裂解缓冲液(analyticalluminescencelaboratories)中进行裂解。用海肾荧光素酶分析系统(promega)根据生产商说明书测定海肾荧光素酶活性。针对细胞提取物中蛋白质浓度(bio-rad)的变化校正海肾荧光素酶活性。一式两份进行测定,实验重复了三次。
4.细胞核和细胞质rna分部分析
细胞核和细胞质rna用cytoplasmicandnuclearrnapurificationkit(norgen,belmont,ca)根据生产商说明书进行分离。
5.体外翻译反应
将纯化的配体(茶碱或eif4g)与1mg信号连接器和1mg海肾荧光素酶mrna温育,然后将该混合物再与thermoscientific1-stephumancoupledivtkit(waltham,ma,usa)的成分按生产商的说明书进行温育。如上所述计算体外翻译的海肾荧光素酶的活性。
6.elisa法测定vegf/opn浓度
用信号连接器或对照稳定转染hek293细胞。然后通过elisa测定法测量vegf/opn蛋白的浓度。简单地讲,每个样品收获106个细胞,重悬于200ml的裂解缓冲液中。离心收集裂解液的上清液,通过酶标仪(bio-rad,hercules,ca)测量od值,并使用标准的校准曲线换算成蛋白质浓度。
7.蛋白印迹分析
将细胞用pbs洗涤,并在ripa缓冲液(50mmtris-hclph7.2,150mmnacl,1%np40,0.1%sds,0.5%doc,1mmpmsf,25mmmgcl2,补充有磷酸酶抑制剂混合物)中裂解。使用bca蛋白测定法测定蛋白质浓度。将等量的全蛋白质提取物电泳到sds-聚丙烯酰胺凝胶上,然后转移到pvdf膜(millipore,billerica,ma)。用5%干牛奶封闭样品,然后与第一抗体(abcam,cambridge,ma)温育过夜。然后,将样品与辣根过氧化物酶缀合的第二抗体(amersham,piscataway,nj)温育,用supersignal化学发光试剂(piercechemicalco.)显影免疫印迹。使用imagej分析软件(美国国立卫生研究院)对蛋白质条带进行定量,通过将每种蛋白质的量针对在同一提取样品中检测到的gapdh水平进行归一化,生成柱状图。每个实验重复三次。
8.细胞增殖试验
用0.25%胰蛋白酶处理细胞(15min,37℃),计算细胞数,然后在0、24、48和72小时用电子细胞计数器(beckmancoulter)进行分析。试验至少独立地重复三次。
9.细胞凋亡测定
使用hoechst33258染色试剂盒(life,eugene,or)观察信号连接器诱导的凋亡细胞。简单地讲,将经处理的细胞在4%多聚甲醛中固定10分钟,在pbs中洗涤两次。然后,将细胞用0.5ml的hoechst33258染色液染色5分钟,用荧光显微镜在350nm波长下拍照。每个测定重复三次。
10.rna提取与实时定量pcr
使用trizol(invitrogen)根据建议的方案从细胞分离总rna。用revertaidtmfirststrandcdnasynthesiskit(fermentas,hanover,md)在2ml体积中从总rna合成cdna链。在abiprism7000fluorescentquantitativepcrsystem(appliedbiosystems,fostercity,ca)上,用all-in-onetmqpcrmix(genecopoieainc,rockville,md)在20ml反应体积中进行实时定量pcr。pcr循环参数为:95℃15min,然后94℃15s,55℃30s,72℃30s,40个循环。用2-δδct方法测定相对表达倍数变化。
11.裸鼠肿瘤形成实验
所有涉及动物的实验都得到了机构审查委员会的批准。4周龄的雌性balb/c裸鼠是在科学院动物中心获得的。将稳定表达信号连接器或阴性对照的107个t24细胞重悬于100mlpbs中,并皮下注射到3只4周大的雌性balb/c裸鼠的左右腋窝。每5天进行一次肿瘤生长检查,肿瘤体积用以下公式计算:0.5*长度*宽度2。注射后20天,将小鼠实施安乐死,并对每个肿瘤的皮下重量进行测量。
12.数据分析
没有使用统计方法预先确定样本容量。调查人员在实验和结果评估过程中盲目地分配。统计分析采用studentt-测验或方差分析,p<0.05被认为具有统计学意义。所有的统计测试都是使用spss17.0版本软件(spss,chicago,il)来完成的。
ii.结果
1.基于rna的信号连接器的设计和构造
既往研究已经证明,将rna核酸适体插入到信使rna(mrna)的5'非翻译区(5'-utr),在配体的存在下可降低翻译起始的速率,该过程是通过阻断核糖体结合所产生的。我们推测:设计的一段反向互补rna序列可用于与靶mrna结合,并且当存在特异性信号时将借助rna适体反式调节mrna的翻译。为了测试这一点,我们设计了含有识别指定mrna的反义结构域(18-22nt大小、优选20nt大小的反义rna)和适体域的rna装置以控制翻译(图1a)。我们将信号连接器和翻译活化结构域偶联以增强翻译,或者单独利用适体结构域以抑制翻译。原则上,我们称为“信号连接器”的这些模块化装置,可以响应任何指定的信号分子(图1b)并控制任何指定靶mrna的翻译,从而将指定的内源信号(输入)连接到特异性细胞信号(输出)(图1c)。
2.信号连接器有效抑制靶基因表达
为了测试这种方法能否抑制翻译起始(图2a)和延伸(图2b),我们设计了12个与海肾荧光素酶报道基因mrna序列不同区域互补的信号连接器,其结合在5'-utr或编码序列(图2c)。这些信号连接体均含有两个片段:与靶mrna序列互补20nt的反义rna序列和两个茶碱适体拷贝(jenisonetal.,1994)。12个连接器分别稳定转染到表达海肾荧光素酶的hek293细胞中,在1000mm茶碱作用下有11个可引起荧光素酶表达的显著降低(图2d)。缺乏茶碱的诱导下,含有信号连接器的细胞中荧光素酶活性相对水平在没有明显改变。
此外,抑制活性似乎与结合位点距mrna5'帽的距离成反比相关,表明靶基因的表达在翻译的早期可更有效地被抑制。然后我们观察到荧光素酶活性的抑制效果与茶碱呈剂量依赖性(图2e),因此我们推测信号连接器的效果受到招募到每个mrna靶标的配体数量的影响。为了测试这种可能性,我们在装置的3'端引入一个或三个茶碱适体,再检测荧光素酶表达水平,并将它们的效果与2个信号连接器的效果进行比较。具有三个适体的装置对荧光素酶表达的抑制作用类似于2个信号连接器,这可能是由于饱和效应。然而,仅含有一个适体的装置仅能非常弱地降低报告基因表达。
我们还使用大分子成分(核糖体、trna、氨基酰基-trna合成酶,起始因子、延伸因子和终止因子)、纯化的配体(茶碱)、体外转录的海肾荧光素酶mrna以及体外转录的rna信号连接器(图2c中使用的r1)或阴性对照进行体外翻译反应。体外数据提示,所观察到的信号连接器的沉默效应的确由配体-适体复合物引起,这提示了路障机制。
然后,我们使用简单的数学模型以便更好地理解各种输入参数和输出之间的关系,该方程描述了剂量或浓度效应关系和最大效应,这是许多生物现象的关键特征。基于这个方程和我们观察到的实验结果,我们将使用2个信号连接器且在足够量的配体存在下进行后续的基因敲低实验。
为了证明这种方法具有模块化设计的特性,我们构建了几个靶向人类血管内皮生长因子(vegf)基因的信号连接器,并用四环素配体替代茶碱适体(mulleretal.,2006)。相关实验数据支持信号连接器对不同适体结构域的模块性(图2f)。如前期预测,这些装置(r13-r15和r17-r20)仅在100μm四环素条件下发挥有效的沉默效应。含有信号连接器细胞的vegfmrna表达水平在缺乏或存在四环素的情况下没有明显变化,也就是表明信号连接器主要起翻译抑制作用,不影响mrna水平(图2)。
这些结果表明信号连接器可以用作基因开关来下调靶基因的表达。
3.信号连接器有效激活内源基因表达
然后我们使用真核翻译起始因子4g(eif4g)相应的rna适体(miyakawaetal.,2006),确定信号连接器是否也可以通过促进起始因子复合物的形成来增强目标基因的翻译(图2g)。eif4g能够募集核糖体40s亚基并激活mrna翻译(moore,2005)。我们选择海肾荧光素酶基因作为靶基因,荧光素酶报告基因检测的结果表明,相对于对照组,具有两个适体的特异性信号连接器可诱导荧光素酶蛋白活性增加15倍(图2h)。从该装置中消除一个适体拷贝后,诱导激活效率显著降低,而添加另一拷贝的适体而表达增加很少(图2)。
我们还构建了一个简单的数学模型来阐明各种输入参数和输出之间的关系,方程揭示了该关系是非线性和可饱和的(图2)。基于用于基因激活的新方程的预测和观察到的结果,我们在足够量的配体存在下,用设计的2个信号连接器进行基因表达激活实验。为了实现翻译起始的动态调节,我们使用识别茶碱的一个适体和识别eif4g以调节基因表达的两个适体的组合,其中反义结构域被设计为与茶碱适体结合(图2g)。茶碱结合稳定适体,并导致允许反义结构域与靶基因的mrna结合。荧光素酶报告基因测定的结果表明,添加茶碱以剂量依赖性方式增加荧光素酶的活性(图2i)。
这些结果表明信号连接体可以用作基因开关来上调内源性靶基因的表达
4.使用信号连接器构建所有基本类型的逻辑门
在电子电路的构建中,逻辑运算和数字系统可以通过使用逻辑门来实现,包括非门、与非门、或门、异或门和异或非门。生物细胞信息处理的许多方面与电子电路的信号集成类似。于是我们提出一个问题,信号连接器是否可以用来构造复杂的可编程逻辑门电路。由于信号连接器强大的基因调控能力,我们尝试构建了各种逻辑门,通过稳定地转染这些装置(图3a),产生输出信号以响应多个输入信号。我们在稳定表达5'端戴帽及不戴帽的海肾荧光素酶mrna的hek293细胞中构建了所有基本类型的双输入布尔逻辑门,通过使用配体识别外源性的茶碱或四环素信号。在表达5'端不戴帽的荧光素酶mrna的构建中,将编码海肾荧光素酶的开放阅读框架(orf)置于初级orf的下游。主要的orf在末端包含一个终止密码子,并且有足够长时间阻止翻译重新启动。
首先,我们构建了两个非门,每个都在低输入时产生高输出。如图2d和f所示,mrna的反义rna目标序列的位置对于信号连接器的抑制效率很重要。我们使用了两个信号连接器(r1和r13),在1000μm的茶碱或100μm四环素的存在的情况下,最大限度地抑制了5'端戴帽的海肾荧光素酶mrna的翻译。如图3b所示,在没有输入信号的情况下,每个门都显示出高荧光素的输出。
然后,我们构建了一个只有在输入信号都很高的情况下才产生高输出的与门。如图2i所示,由于激活效率与目标距mrna5’端的距离成反比,我们使用了两个信号连接器(r25和r29),在配体的存在中,各自对5’端无帽的海肾荧光素酶mrna仅产生微小的翻译激活作用。如图3c所示,虽然每一个单独的信号(1000μm的茶碱或100μm四环素)并没有显著地刺激目标荧光素酶基因的表达,但这两种信号输入都存在的情况下,通过两个装置之间的协同作用产生了强大的激活作用。
我们还构建了一个与非门,如果任何输入都很低,它就会显示出较高的输出值。我们使用两个信号连接器(r12和r16),在配体的存在下,对5端’戴帽的海肾荧光素酶信使mrna进行最低限度的抑制。如图3所示,虽然引入单个信号(1000μm的茶碱或100μm四环素)并没有显著抑制目标荧光素酶基因的表达,但在两种输入信号存在的情况下,协同作用地诱导了强大的抑制作用。
接下来,我们构造了一个或门,如果其中一个或两个输入都很高,那么它就会产生高输出。在此建造过程中,我们仍然使用了两个信号连接器(r22和r26),每一个信号连接器都在配体的存在下,强烈激活了5’端无帽的海肾荧光酶mrna的翻译。如图3e所示,荧光素酶信号可以由这两个信号中的任何一个(1000μm的茶碱或100μm四环素)产生。
我们还构建了一个或非门。我们使用了两个信号连接器(r1和r13)最大限度地抑制了海肾荧光素酶mrna的翻译。由于单个信号的引入明显抑制了目标基因的表达,所以只有当两个信号(1000μm的茶碱或100μm四环素)都没有出现时,荧光素酶才能产生。(图3f)
我们还构建了一个异或门。我们设计了两个信号连接器(r22和r27),目标是在5’端无帽的海肾荧光素酶mrna的两个不同区域,同时它们彼此的rna序列是互补的。结果表明,每一种装置在相应的配体中都能强烈激活荧光素酶的表达。与此相反,由于两个装置的反义域(图3g)的特殊碱基配对,这两种装置同时引入并没有显著激活目标荧光素酶基因的表达。
最后,我们构建了一个异或非门。使用相似的设计策略,我们使用了两个信号连接器(r1和r14),它们在相应的配体中,强烈抑制5’端戴帽的海肾荧光酶mrna的翻译。当1000μm的茶碱或100μm四环素同时存在或同时不存在时,异或非门可以产生一个高荧光素酶信号的输出(图3h)。
这些结果表明,信号连接器可以将输入信号与所需的细胞输出信号建立逻辑连接。
5.信号连接器能够有效地重新连接信号通路并创建反馈回路
在真核细胞中,信号蛋白经常激活转录因子来启动下游基因的转录。因为在理论上,信号连接器可以将转录因子与抑制下游基因翻译联系起来,我们通过重新连接原有的细胞信号通路来改造分子网络(图4a)。β-连环蛋白蛋白是一种多功能蛋白质,通常在癌细胞的细胞核中聚集,激活致癌基因c-myc基因的转录(he等人,1998)。我们合成了一种信号连接器,其中包含了一个β-连环蛋白配体(culler等人,2010),同时通过反义序列定位在c-mycmrna5′utr(表s7)内的区域。我们研究了在hek293细胞中利用白细胞三烯d4(ltd4)刺激β-连环蛋白通路的效果。稳定表达了信号连接器或阴性对照后,两种细胞系均表现出c-mycmrna(图4b)的表达,而与阴性对照组的细胞相比,稳定表达信号连接器的细胞中c-myc蛋白的表达能力明显下降(图4c)。这些结果表明,我们的信号连接器可以通过在转录因子和下游基因的mrna之间建立一个负向连接,从而有效地重新连接信号通路。
我们还测试了信号连接器是否可以促使特定的调控因子激活下游目标基因(图4),从而创造出一种新的信号传导通路。特定的信号连接器使用一个配体域来识别β-连环蛋白信号和其他两个配体域来形成翻译起始因子复合物。在没有β-连环蛋白信号的情况下,β-连环蛋白适体的茎可“关闭”反义结构域。在有β-连环蛋白信号的情况下,这个信号连接器可以与目标海肾荧光酶mrna(图4e)相互作用。通过在hek293t细胞中稳定转染信号连接器或阴性对照,研究了白三烯d4(ltd4)的作用。荧光素酶测定结果表明,在表达信号连接器的细胞中,荧光素酶性明显升高(图4f),而在表达阴性对照的细胞中,其活性不受ltd4影响。这些结果表明,β-连环蛋白信号可以在设计的信号连接器的帮助下,有效地激活海肾荧光素酶的表达。
接下来,我们测试了信号连接器将反馈回路纳入基因-基因相互作用网络的能力。正反馈回路可以放大从发送方接收到的信号,并将系统从初始状态跃迁。我们用信号连接器设计了一个反馈回路,其中骨蛋白(opn)和vegf是彼此的激活物(图4g)。opn和vegf是分泌蛋白质的细胞因子,调节细胞活动和血管生成(ferrara等人,2003;lyle等人,2014),并且他们的rna配体在之前的文献中已经提出(ng等人,2006;mi等人,2009)。我们将eif4g配体的两个拷贝插入opn或vegf核酸开关(riboswitch)的3'端,构造信号连接器识别vegf或opn。结果表明,在膀胱癌中,opn和vegf是相互独立的。我们将opn或vegf过表达质粒,转染进入表达信号连接器的t24细胞,并发现相应过表达质粒的转染有效地提高了受调控基因的表达水平(图4g)。我们还研究了信号连接器是否可以用于在opn和vegf(图4h)之间构造负反馈循环,使系统更加稳定。因为opn可以通过信号连接器诱发vegf的表达,我们只需要证明vegf还可以通过类似的方法抑制opn的表达。我们将vegf配体的两个拷贝插入到反义rna的3'端,构造调控opn的信号连接器。利用稳定地表达这个信号连接器的t24细胞,我们证明了vegf的瞬态表达可以减少opn的浓度,而vegf基因敲除后又增加了opn的水平(图4h)。
这些结果表明,信号连接器是构造调控回路和基因-基因网络的有效工具。
6.信号连接器能沉默存活基因表达并抑制癌细胞生长
为了检查信号连接器是否可以用来识别细胞,重新编程细胞状态和行为,我们使用了人类端粒酶逆转录酶(htert)启动子来驱动沉默存活基因的信号连接器的表达(gao等人,2014),并选择膀胱癌细胞作为靶细胞(图5)。htert
启动子(htertp)在超过85%的人类癌症中是非常活跃的,但是在大多数正常的细胞中是不活跃的(takakura等人,1999)。因此,我们构建了一个逻辑与门,其中激活的htertp和配体必须联合起来抑制存活基因(图5b)。如前所述产生抑制人类c-myc基因(sardi等人,1998)和bcl2基因(kunze等人,2012)的信号连接器,并稳定地转染胱癌细胞或正常的成纤维上皮细胞(图5c)。在膀胱癌细胞系中,与缺乏配体(图5d)相比,相应的装置能够显示出在存在茶碱的情况下基因表达显著下降。这些装置并没有对成纤维上皮细胞产生抑制作用,无论配体是否存在(图5e)。这些细胞系的生长曲线也表明,该线路有效地抑制了靶标膀胱癌细胞的增殖,而不影响成纤维上皮细胞(图5f)。此外,我们还研究了这些装置是否可以诱导癌细胞凋亡。用信号连接器处理的膀胱癌细胞表现出较强的蓝色荧光,显示典型的凋亡特征。相反,信号连接器在正常细胞中没有这样的效果(图5g)。
这些结果表明,基于信号连接器的与门线路可以抑制目标细胞株的基因表达。
7.通过信号连接器将致癌信号转化为抑癌信号
在人类细胞中成功应用信号连接媒介的转译控制,为同时开启或关闭多基因转译程序打开了一种途径,在该程序中,一些基因被激活,其他基因被抑制。我们假设这些装置应该有可能改变致癌途径的输出,并通过同步激活和抑制内源性基因来控制癌细胞的命运。
nf-kb是一种促进癌症发生发展的致癌信号。nf-kb通过激活cyclind1、c-fos和c-jun等下游靶基因来促进细胞增殖(li等人,2015)。因此,我们试图通过使用信号连接器将nf-kb信号从增殖通路重新连接到静息/死亡通路。我们构建了4个信号连接器识别nf-kb(p65)(urster等人,2008)以激活bax(r37)和p21(r38),并抑制两个致癌基因bcl2(r39)和c-myc(r40)。人膀胱癌t24细胞中表达高水平的nf-kb信号(图6a)。两个拷贝的eif4g配体被插入到nf-kb核酸开关的3'端,以构建激活bax或p21的信号连接器,而两个nf-kb的配体被连接到反义rna3'端,以构建抑制bcl2和c-myc的信号连接器。结果表明,在稳定表达信号连接器的t24细胞中可以同时增强bax和p21的蛋白质表达水平,减少bcl2和c-myc(图6b)。最后,我们确定了信号连接器能否同时抑制体内肿瘤的生长。将稳定表达信号连接器或其阴性对照的t24细胞注射到雄性裸鼠中。在注射后20天,我们发现在信号连接器组中形成的肿瘤比阴性对照组的肿瘤小得多(图6c-e)。另外,与实验结束时的阴性对照组相比,信号连接器组的平均肿瘤重量明显较低(图6f)。
这些结果表明,信号连接可以通过重定向致癌信号通路来抑制体内肿瘤的生长。
以上应用了具体实例对本发明进行阐述,只是用于帮助理解本发明,并不用以限制本发明。对于本发明所属技术领域的技术人员,依据本发明的思想,还可以做出若干简单推演、变形或替换。
- 还没有人留言评论。精彩留言会获得点赞!