整合四拷贝功能性F18菌毛操纵子基因的益生菌克隆株、构建方法及应用与流程
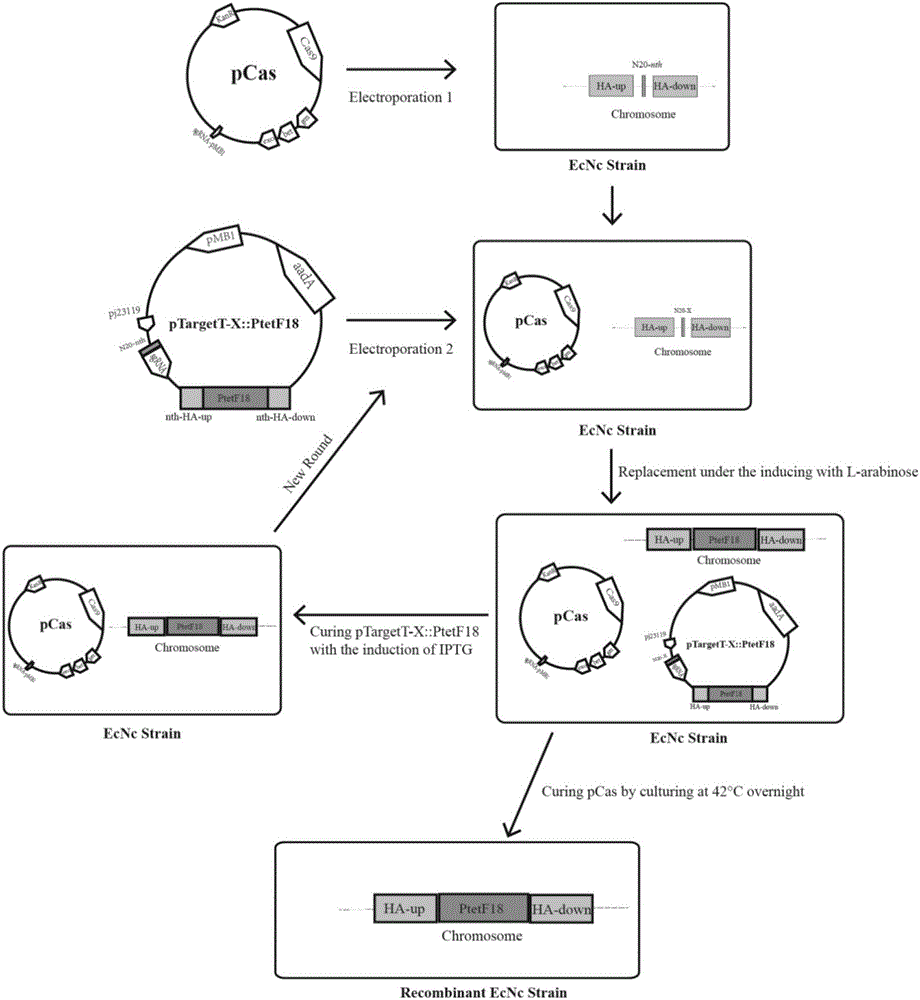
本发明涉及到生物技术应用领域,具体涉及到细菌的染色体整合、稳定遗传表达外源功能性f18菌毛蛋白。本发明利用crispr/cas9双质粒系统具有高效编辑细菌基因组(包括整合插入,敲除,点突变)的特性,且该crispr/cas9双质粒系统具有自身质粒去除简便和不残留抗性基因的机制。完成构建大肠杆菌益生菌nissle1917无质粒克隆菌ecnc表面稳定携带四拷贝f18菌毛操纵子基因并遗传表达f18菌毛,并不影响宿主菌ecnc的自身生物学性状,该重组整合菌株制备有望成为构建抗f18+菌株引起的仔猪断奶后腹泻或水肿病益生菌活疫苗候选株的新方法,实现多功能重组益生菌活疫苗候选株的制备。
背景技术:
:大肠杆菌nissle1917是一株无致病性,至今并未发现对宿主有已知的害处影响,能给予不同宿主健康益处,且该菌株作为在欧洲上市的名为mutaflor益生菌产品的主要活性成分,主要用于人类肠道健康调节。在临床应用上,益生菌nissle1917菌株作为基因工程的载体被加以改造,该益生菌已用于疫苗、肿瘤治疗、保健品和诊断制剂等方面产品的开发与研制。ecnc克隆株为大肠杆菌nissle1917原型野生株基因改造去除其胞内两个隐秘性质粒pmut1和pmut2,具备比亲本株携带更多(大)容量的同源或外源基因的特性,已在申请人实验室制备获得。为了实现微生物中同源或异源蛋白或化合物的过量表达生产,大多利用质粒的过表达,这种方法易于操作和调控的表达而被广泛使用,但这种方法存在遗传不稳定性。质粒的复制,抗生素抗性基因,过量表达以及其他异源基因等问题所带来的代谢负担会导致宿主细胞耗竭,产量损失甚至导致宿主细胞原始功能丧失。此外,为了维持细菌细胞中质粒的存在,须使用抗生素或其他选择性药剂,从而增加了整个生物加工的成本,同时也增加了耐药基因传播的几率。近年来,细菌染色体表达同源或异源基因在合成生物学和生物医学中越来越受青睐。在细菌染色体组整合同源或外源的dna基因分子,常用的分子生物学方法包括使用转座子、噬菌体、核酸内切酶以及重组酶系统,常规的分子生物学方法在细菌染色体整合基因上具有一定的优势,但也都存在着自身的局限性,如长片段基因难以整合、效率低、脱靶率高等。本次申请专利所使用的方法为crispr/cas9双质粒系统对益生菌nissle1917无质粒克隆株染色体进行外源大片段dna基因片段(f18菌毛操纵子编码基因长约5.6kb)的整合。断奶仔猪腹泻(pwd)和仔猪水肿病(ed)为养猪业临床上常见的细菌性疾病,主要病原菌为f4和f18菌毛阳性产肠毒素大肠杆菌etec,通过菌毛黏附、定植易感仔猪后感染发病,导致仔猪高死亡率,体重降低,生长缓慢,药物治疗费用昂贵等。临床上防治该疾病主要运用抗生素治疗,但随着养殖业生产中耐药菌的不断产生和累积,急需研究新的防控措施。本发明所构建的益生菌nissle1917无质粒克隆菌ecnc稳定携带遗传且在菌体表面表达和展示功能性f18菌毛,有望为断奶仔猪腹泻或水肿病的防控提供新思路和策略。技术实现要素:为了克服上述缺陷,本发明的目的在于提供整合四拷贝功能性f18菌毛操纵子基因的益生菌克隆株、整合四拷贝功能性f18菌毛操纵子基因表达展示在nissle1917益生菌克隆株细胞表面的构建方法及应用,利用益生菌nissle1917无质粒克隆菌ecnc菌株染色体中四个非必需基因x(yjcs,pcada,lacz,yien/trkd)内的位点,四拷贝整合并稳定表达f18菌毛,有望作为由f18+菌株引起的抗仔猪断奶后腹泻或水肿病的多功能重组益生菌活性疫苗候选株。为了实现上述发明目的,本发明所采用的技术方案为:整合四拷贝功能性f18菌毛操纵子基因的益生菌克隆株,其特征在于,该益生菌克隆株保藏于中国微生物菌种保藏管理委员会普通微生物中心cgmcc,保藏地址为北京市朝阳区北辰西路1号院3号中国科学院微生物研究所,保藏日期为2018.8.13,保藏编号为cgmccno.16253。整合四拷贝功能性f18菌毛操纵子基因表达展示在nissle1917益生菌克隆株细胞表面的构建方法,包括如下步骤:1)ptargett-x::ptetf18重组质粒的构建;x表示ecnc染色体上特定基因的靶标位点;2)通过整合四拷贝功能性f18菌毛操纵子基因的益生菌克隆株的构建,得到无抗性的四拷贝f18菌毛基因整合克隆株。步骤2)具体为:pcas质粒电转化入益生菌ecnc中以构建ecnc/pcas重组菌株:制备ecnc/pcas感受态细胞(在第一轮待插入yjcs位点时选择ecnc/pcas感受态细胞;而第二轮开始,制备插入了yjcs位点的ecnc/pcas感受态细胞以使得在第二轮插入cadap位点;而第三轮时,制备插入了yjcs、cadap位点的ecnc/pcas感受态细胞;依此类推),取ecnc/pcas感受态细胞(第一轮所用的感受态细胞为无插入的ecnc/pcas感受态细胞,第二轮,此感受态细胞为已经插入了yjcs位点的ecnc/pcas感受态细胞;第三轮,此感受态细胞为已经插入了yjcs,cadap的ecnc/pcas感受态细胞,依此类推)与一种ptargett-x::ptetf18重组质粒混合,对ecnc/pcas感受态细胞和ptargett-x::ptetf18重组质粒组成的混合物进行电转化,电击产物置于冰冻的lb培养基孵育,之后于30℃摇床培养复苏,涂布于含卡那霉素和壮观霉素的lb平板中于30℃培养过夜;挑取抗性平板上生长的可疑阳性克隆株病利用菌落pcr鉴定,若出现大于5600bp的条带,则证明整合成功,测序并确定是否获得了阳性单拷贝f18整合株,再去除抗性质粒;之后,进行下一次整合实验和去除抗性质粒操作,直至最终得到无抗性的四拷贝f18菌毛基因整合克隆株。步骤2)中,去除抗性质粒的步骤为:将含有pcas和ptargett-x::ptetf18质粒的ecnc菌株置于含卡那霉素和0.5mmiptg的lb培养基中30℃摇床中培养14-18小时,之后挑取少许菌液于含卡那霉素的lb平板上划线,长出的单菌落于含壮观霉素的lb平板上划线以验证其对壮观霉素的敏感性,以验证是否去除了ptargett-x::ptetf18质粒;去除ptargett-x::ptetf18质粒后,最后再进行另外一个温度敏高型质粒pcas的去除,即将该ecnc菌株置于无抗性的lb培养基中于42℃中传代,直到最终得到无抗性的四拷贝f18菌毛基因整合克隆株。此步骤2)中,每一轮的整合后,都要去除对应的ptargett-x::ptetf18质粒,而质粒pcas的去除操作在最后一轮的整合完成并去除最后一轮的ptargett-x::ptetf18抗性质粒后才进行。步骤1)具体为:利用pb032-x/pb033扩增引物从ptargetf质粒dna模板中反向pcr扩增含有靶标位点x的n20序列的ptargetf-x线性化片段,随后对所得的线性化片段环化得到第一环化产物,第一环化产物转化入感受态细胞dh5α中以构建ptargett-x重组质粒;xupsteamha-f/xupsteamha-r和xdownsteamha-f/xdownsteamha-r扩增引物分别用来从nissle1917基因组模板中扩增靶标位点的上游同源臂和下游同源臂;ptetfwd/ptetf18rev-x扩增引物用于从pbr322-f18dna模板中扩增带有ptet启动子的f18菌毛操纵子基因;ipcrptargetf-f(hindiii)/ipcrptargetf-r(psti)用来线性化ptargett-x重组质粒;使用multis一步克隆试剂盒将靶标位点上、下游同源臂、含ptet启动子的f18菌毛操纵子基因片段、线性化的ptargett-x片段一起环化得到第二环化产物;第二环化产物随后转化进感受态细胞dh5α中以构建ptargett-x::ptetf18系列重组质粒;x代表ecnc染色体上特定基因的靶标位点,特定基因包括yjcs,cadap,lacz,yien/trkd。上述四个片段的环化连接排列的顺序为:靶标位点上游同源臂片段、含ptet启动子的f18菌毛操纵子基因片段、靶标位点下游同源臂片段、线性化的ptargett-x片段,四片段进行了环化连接得到第二环化产物。本发明的另一个目的为提供整合四拷贝功能性f18菌毛操纵子基因的益生菌克隆株作为重组益生菌活疫苗候选株的应用,以及整合四拷贝功能性f18菌毛操纵子基因的益生菌克隆株作为断奶仔猪腹泻和仔猪水肿病的预防治疗用生物制剂的应用。步骤2)中,完成一轮整合插入实验后,使用iptg诱导切割ptargett-x::ptetf18质粒复制起始位点pmb1,消除ptargett-x::ptetf18质粒,同时消除壮观霉素sp+抗性,而后进行下一轮(下一位点)的整合插入试验。当上述四个位点yjcs,cadap,lacz,yien/trkd都已整合重组完毕后,去除最后一轮的ptargett-x::ptetf18质粒后,最终于42℃传代培养消除pcas质粒(温度敏感性质粒),直至该重组菌丢失卡那霉素kan+抗性,筛选出无抗性四拷贝整合f18菌毛操纵子基因的重组菌株。一种细菌染色体基因组插入功能性大片段外源基因的生物学技术,对宿主菌基因组无任何抗性遗传修饰。其外源基因片段于益生菌nissle1917无质粒克隆株ecnc染色体基因组的四个非必需基因x(yjcs,pcada,lacz,yien/trkd)内的位点内插入的生物学技术,是基于使用crispr/cas9双质粒系统的基因编辑技术。crispr/cas9双质粒系统中的一个重组质粒ptargett-x::ptetf18构建的生物学技术,基于使用高保真pcr扩增dna聚合酶,质粒环化,线性化和多片段拼接克隆dna重组技术将crispr/cas9系统质粒ptargett质粒构建成重组质粒ptargett-x::ptetf18。保留ptargett质粒的基本特性,包括壮观霉素抗性筛选标签aada基因,复制起始位点pmb1,调控转录的合成启动子pj23119和sgrna特征序列等特性。重组质粒同时携带能介导cas9蛋白切割靶标位点的n20-x-sgrna与两侧带有插入位点同源臂的f18操纵子序列。ecnc/pcas重组菌导入重组质粒ptargett-x::ptetf18后,在l-阿拉伯糖诱导pcas质粒上的gam,bet和exo三种活性酶的作用和抗性选择压力的存在下,实施同源重组而获得整合了f18操纵子基因的ecnc整合重组菌株。使用iptg诱导切割ptargett-x::ptetf18质粒复制起始位点pmb1,消除ptargett-x::ptetf18质粒,同时消除壮观霉素sp+抗性,而后进行下一轮(下一位点)的整合插入试验。当上述四个位点都已整合重组完毕后,去除最后一轮的ptargett-x::ptetf18质粒后,最终于42℃传代培养消除pcas质粒(温度敏感性质粒),直至该重组菌丢失卡那霉素kan+抗性,筛选出无抗性四拷贝整合f18菌毛操纵子基因的重组菌株。相对于现有技术,本发明的有益效果为:1、去除nissle1917益生菌原有质粒,不引入新的外源质粒,无抗性基因的存在。2、该重组菌不存在需使用抗生素才能长期维持外源基因的表达。3、经多次传代,仍稳定持久遗传表达外源菌毛蛋白(f18菌毛)。4、本活疫苗候选株的成功构建不会影响该益生菌宿主菌已知的如生长性能等生物学特性。5、相对于亲本株,该重组菌能够有效提高对猪肠道传代上皮细胞(ipec-1)的粘附性能。6、口服免疫小鼠能产生免疫血清,即抗f18菌毛的igg的抗体。7、口服免疫后小鼠的血清能显著减低f18+菌株(如强毒菌株8516)对猪肠道传代细胞系ipec-j2的粘附。附图说明图1为整合四拷贝f18菌毛编码操纵子基因的益生菌ecnc重组菌株的构建过程示意图;图2为四拷贝整合重组菌mcf18is抗f18的免疫印迹图;m:蛋白标准标记物,ecnc:消除了两个隐性质粒的大肠杆菌nissle1917,mcf18is:f18多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18;图2所用一抗为抗f18菌毛鼠源多抗血清;图3为四拷贝整合重组菌mcf18is体外粘附实验结果图;ecnc:消除了两个隐性质粒的大肠杆菌nissle1917,mcf18is:f18多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18;该图表明相对于ecnc菌株,四拷贝整合菌mcf18is对猪源肠道传代上皮细胞的粘附能力的百分比变化情况,所用的细胞为ipec-1细胞;图4为间接elisa抗体滴度检测结果图;ecnc:消除了两个隐性质粒的大肠杆菌nissle1917,mcf18is:f18多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18;分别将ecnc和四拷贝整合重组菌mcf18is口服免疫8周龄balb/c小鼠各两次,时隔两周,收集并分离所得到的免疫血清进行间接elisa试验,检测验证免疫小鼠的抗f18的抗体滴度变化;图5为体外细胞粘附抑制实验结果;ecnc:消除了两个隐性质粒的大肠杆菌nissle1917mcf18is:f18多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18;分别将ecnc和四拷贝整合重组菌mcf18is口服免疫8周龄balb/c小鼠各两次,时隔两周,所得免疫血清测试验证其对f18+etec对猪源肠道细胞的抑制粘附活性;该图表明相对于ecnc菌株,口服整合重组菌的小鼠所得的免疫血清对etec粘附猪源肠道上皮细胞抑制百分比变化情况,所用的细胞为ipec-j2传代细胞,所用etec细菌为f18阳性猪场田间分离病原性大肠杆菌菌株8516;本发明的益生菌克隆株保藏于中国微生物菌种保藏管理委员会普通微生物中心cgmcc,保藏地址为北京市朝阳区北辰西路1号院3号中国科学院微生物研究所,保藏日期为2018.8.13,保藏编号为cgmccno.16253;分类命名为大肠埃希式菌escherichiacoli。具体实施方式以下通过具体实施例对本发明作进一步详细说明。著名益生素大肠杆菌nissle1917(ecnc)为非致病性的大肠杆菌益生菌,已被证实该益生菌能给予不同宿主健康益处,并且大肠杆菌nissle1917是一株无致病性,能给予不同宿主健康益处,并无发现对宿主有已知的不益影响,并且该菌株作为在欧洲上市的名为mutaflor益生菌产品的主要活性成分,主要用于人类肠道健康调节。著名益生素大肠杆菌nissle1917(ecnc)保藏于中国微生物菌种保藏管理委员会普通微生物中心cgmcc,保藏地址为北京市朝阳区北辰西路1号院3号中国科学院微生物研究所,保藏日期为2018.7.2,保藏编号为cgmccno.16045。本试验利用益生菌nissle1917无质粒克隆株ecnc为疫苗载体构建稳定表达f18菌毛的重组益生菌,以此作为日常饲料添加剂免疫仔猪以防控仔猪断奶后腹泻(pwd)。该f18菌毛整合重组益生菌能够特异性的结合到猪肠道细胞的f18菌毛受体上,优先占据受体位点,并诱导抗f18的粘膜免疫,从而阻断病原f18+etec对仔猪肠道的粘附,有效预防仔猪断奶后腹泻或水肿病产生。1、ptargett-x::ptetf18重组质粒的构建所用的引物及含有pam的n20序列如表2,表3所示。表2中序列从上至下依次为seqidno.1-44,表3中序列从上至下依次为seqidno.45-48。用超高保真dna聚合酶(vazyme,p501)进行pcr扩增反应(反应体系见表1)。本申请书上,x代表ecnc染色体上特定基因的靶标位点,特定基因包括yjcs,cadap,lacz,yien/trkd。表1质粒构建时的pcr扩增反应体系pcr循环参数为:95℃预变性2min;95℃变性10sec;退火温度一般为55℃(若扩增不出来温度升高或降低1~3℃)退火时间为30sec;72℃延伸25sec/kb(根据不同pcr产物长度而定);35个循环,72℃最后延伸10min。pcr反应结束后,产物通过1.0%~1.5%浓度(视扩增出来的产物片段长度而定)的琼脂糖凝胶电泳后切胶回收纯化对应的pcr产物进行下一步的试验。首先,利用pb032-x/pb033扩增引物从ptargetf质粒dna模板中反向pcr扩增含有靶标位点x的n20序列的ptargetf-x线性化片段,随后所得的线性化片段利用ⅱ一步克隆试剂盒(vazyme,c112)环化pcr产物,转化进感受态细胞dh5α中以构建ptargett-x重组质粒。xupsteamha-f/xupsteamha-r和xdownsteamha-f/xdownsteamha-r扩增引物分别用来从nissle1917基因组模板中扩增靶标位点的上游同源臂和下游同源臂。ptetfwd/ptetf18rev-x扩增引物用于从pbr322-f18质粒的dna模板中扩增带有ptet启动子的f18菌毛操纵子基因。ipcrptargetf-f(hindiii)/ipcrptargetf-r(psti)用来线性化ptargett-x重组质粒。使用multis一步克隆试剂盒(vazyme,c113)将上述四段pcr产物片段进行环化,四段pcr产物片段包括靶标位点上下游同源臂,含ptet启动子的f18菌毛操纵子基因片段,线性化的ptargett-x片段(即ptargett-x重组质粒的线性化产物)。随后转化进感受态细胞dh5α中以构建ptargett-x::ptetf18系列重组质粒。2、四拷贝f18菌毛益生菌ecnc整合株的构建整合重组菌株构建的过程参见附图1注解。pcas质粒电转化进ecnc中构建ecnc/pcas重组菌株。制备ecnc/pcas感受态时,10mm终浓度的l-阿拉伯糖用于后期λ-red同源重组的诱导(即在制备ecnc/pcas感受态时,加入10mm终浓度的l-阿拉伯糖)。取50μl感受态细胞与约100ngptargett-x::ptetf18系列dna(每一次整合插入实验仅加入一种ptargett-x::ptetf18重组质粒)混合;混合物放于电极间隙为0.1cm的电极杯(bio-rad)中1.8kv进行电转化,电击产物置于1.5ml指形管并加入1ml冰冻的lb培养基孵育约5min,之后于30℃摇床复苏1小时,随后涂布于含卡那霉素(50mg/liter)和壮观霉素(50mg/liter)的lb平板中于30℃培养过夜。挑取抗性平板上生长的可疑阳性克隆株利用菌落pcr鉴定(鉴定引物为第一对引物yzxptetf18up-f/yzxptetf18up-r和第二对引物yzxptetf18down-f/yzxptetf18down-r,上述两对引物的pcr结果若均有阳性条带,则证明整合成功;外加一对鉴定引物yzxptetf18up-f/yzxptetf18down-r,该对引物pcr产物若只有约600bp,则证明整合失败,若出现大于5600bp的条带,则证明整合成功,随后选取阳性整合重组菌株送测序再次鉴定。验证用的pcr反应体系为常规pcr反应体系。pcr循环参数为:95℃预变性2min;95℃变性10sec;退火温度一般为55℃(若扩增不出来温度升高或降低1~3℃)退火时间为30sec;72℃延伸25sec/kb(根据不同pcr产物长度而定);25个循环,72℃最后延伸10min。pcr反应结束后,产物通过1.0%~1.5%浓度(视扩增出来的产物片段长度而定)的琼脂糖凝胶电泳进行扩增产物大小判断。在获得阳性整合株后,进一步去除抗性质粒:将含有pcas和ptargett-x::ptetf18质粒的ecnc菌株(即前述的阳性整合重组菌株)置于约5ml含卡那霉素(50mg/liter)和iptg(0.5mm)的lb培养基中30℃摇床中培养14小时以上,之后挑取少许菌液于含卡那霉素(50mg/liter)的lb平板上划线,长出的单菌落于壮观霉素(50mg/liter)lb平板上划线以验证其对壮观霉素的敏感性,测试验证证明去除了ptargett-x::ptetf18质粒。而后进行下一轮(下一位点)的整合插入试验,一共进行四轮整合插入试验(四轮整合插入试验分别针对yjcs,cadap,lacz,yien/trkd四个位点的先后顺序进行整合插入。当上述四个位点都已整合重组完毕后,去除ptargett-x::ptetf18质粒之后,最终再进行另外一个质粒pcas的去除,将该重组克隆株置于无抗性的lb培养基中于42℃中传代,直到最终得到无抗性的四拷贝f18菌毛基因整合克隆株。表2中引物从上至下为seqidno.1-44。表3中引物从上至下为seqidno.45-48。表2本试验所用引物备注:x代表ecnc染色体上的靶标位点,如yjcs,cadap,lacz,yien/trkd。表3本试验所用的ecnc染色体上靶标位点序列外加pam三碱基targetedlocusregionn20+pamyjcsggatatgtggggtaacgacgcggpcadagttcgcagtggaagtaccgttgglaczctggggaatgaatcaggccacggyien/trkdgagggtgagccataatgaagtgg备注:pam三碱基为上述四个序列的最后三个碱基。图2为四拷贝整合重组菌mcf18is抗f18的免疫印迹图。通过玻板凝集试验及westernblot测试验证,f18菌毛能够于重组益生菌ecnc上功能性表达展示。在标准免疫印迹试验中,ecnc重组整合菌株(通过煮沸全菌所制备的蛋白样品)中表达的f18菌毛的亚单位fedc能被小鼠抗f18多抗血清(1:500稀释)抗体来检测并识别。将ecnc菌株和8516菌株的煮沸全菌蛋白样品分别用作阴性对照和f18的阳性对照。多重整合菌株所表达的f18菌毛亚单位fedc的目标蛋白条带(23kda)能够被抗f18多抗血清识别。m,sizemarker;ecnc:消除了两个隐性质粒的escherichiacolinissle1917;mcf18is,多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18;8516:f18阳性参考菌株。图3为四拷贝整合重组菌mcf18is体外粘附实验结果图。该图表明相对于ecnc菌株,四拷贝整合菌mcf18is对猪源肠道传代上皮细胞的粘附能力的百分比变化情况,所用的细胞为ipec-1细胞。从图3可知,重组益生菌对猪源肠道传代上皮细胞ipec-1的粘附能力有显著性的提高,进一步证明该重组菌能够表面功能性表达并展示f18菌毛。图3中,ecnc的粘附指数设定为100%。数据表示为三次重复实验的平均值的标准误差(sem)。通过体外细胞粘附试验,结果表明,相对于ecnc菌株(100±9.82%),f18多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18,对ipec-1细胞的粘附性能提高了78.96±26.69%,差异极显(**,p=0.0069)。ecnc:消除了两个隐秘性质粒的escherichiacolinissle1917;mcf18is,多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18。图4为间接elisa抗体滴度检测结果图。分别将ecnc和四拷贝整合重组菌mcf18is口服免疫8周龄balb/c小鼠各两次,时隔两周,收集并分离所得到的免疫血清进行间接elisa试验,检测验证免疫小鼠的抗f18的抗体滴度变化。用balb/c小鼠模型做动物实验能获得抗f18菌毛igg的抗体滴度,因而能够检测出重组菌所表达的f18免疫抗原性(其免疫原性优异)。相对于免疫了ecnc菌株的小鼠血清的抗f18抗体滴度(0.809±0.512),免疫了四拷贝整合重组菌mcf18is的小鼠血清样品抗f18抗体水平(2.092±0.292)上升了不少。ecnc:消除了两个隐秘性质粒的escherichiacolinissle1917;mcf18is,多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18。图5为体外细胞粘附抑制实验结果。分别将ecnc和四拷贝整合重组菌mcf18is口服免疫8周龄balb/c小鼠各两次,时隔两周,所得免疫血清测试验证其对f18+etec对猪源肠道细胞的抑制粘附活性。该图表明相对于ecnc菌株,口服整合重组菌的小鼠所得的免疫血清对etec粘附猪源肠道上皮细胞抑制百分比变化情况,所用的细胞为ipec-j2传代细胞,所用etec细菌为f18阳性猪场田间分离大肠杆菌菌株8516。体外细胞抑制试验证明,相对于免疫了ecnc菌株小鼠血清抑制细菌粘附的能力(100±7.07%),免疫小鼠血清能显著抑制f18+野生菌(如猪场田间分离株8516)对猪源肠道传代上皮细胞ipec-j2的粘附下降(32.54±6.63%),差异极显(**,p=0.0032)。ecnc:消除了两个隐秘性质粒的escherichiacolinissle1917;mcf18is,多重整合菌株,ecncδyjcs::ptetf18δpcada::ptetf18δlacz::ptetf18δyien/trkd::ptetf18。本发明建立在申请人实验室制备获得的大肠杆菌nissle1917无质粒克隆株ecnc(去除了细菌内的两个隐秘性质粒pmut1和pmut2)细菌染色体基因组序列信息清晰的基础上,选择其基因组内四个非必需基因x(yjcs,pcada,lacz,yien/trkd)内的位点作为插入f18菌毛操纵子基因的位点,使用crispr/cas9双质粒系统构建含大肠杆菌f18菌毛操纵子基因,其两侧带有插入位点x同源臂的crispr/cas9重组质粒ptargett-x::ptetf18。进一步将重组质粒ptargett-x::ptetf18转化入表达cas9蛋白的重组nissle1917/pcas宿主菌,鉴于pcas质粒上表达cas9蛋白,结合重组质粒ptargett-x::ptetf18上的利用sgrna靶向指导cas9切割酶特异性切割靶标位点和同源重组酶(gam、exo、bet)作用,以导入的同源片段为模板实施同源重组,完成f18菌毛操纵子基因整合进ecnc基因组的靶向位点。而后使用iptg诱导启动pcas质粒上iptg诱导依赖性启动子(ptrc),从而启动ptrc启动子下游的靶向于ptargett-x::ptetf18质粒上的pmb1复制子的sgrna(sgrnapmb1),高效切割ptargett-x::ptetf18质粒上复制起始位点(pmb1),从而消除ptargett-x::ptetf18质粒并同时消除壮观霉素抗生素筛选标志。经计划的四轮不同位点整合重组反应,相继完成f18菌毛操纵子基因整合进ecnc基因组的四个非必需位点中。最后在42℃培养,消除温度敏感性pcas质粒,最终获得不含任何抗生素抗性基因且表达f18菌毛的重组nissle1917大肠杆菌无质粒克隆株。本方法所构建的重组菌株通过无抗性多代(至少30代)培养后,westernblot检测验证能稳定遗传表达f18菌毛,生长周期曲线与亲本株一致,且该方法不会影响宿主菌已知的如生长速度等生物学特性。通过体外细胞粘附实验该重组菌能够有效提高重组菌对肠道传代细胞(ipec-1)的粘附性能,经两轮口服免疫8周龄balb/c小鼠动物实验后,所得免疫血清具有抗f18菌毛蛋白的igg滴度,且其血清能显著减低f18+菌株(如猪场田间分离株8516)对猪肠道传代细胞系ipec-j2的粘附,该重组整合菌株制备有望成为构建抗f18+菌株引起的仔猪断奶后腹泻或水肿病益生菌活疫苗候选株的方法。以上显示描述了本发明的基本原理、主要特征以及优点。本行业的技术人员应该了解,本发明不受上述实施例的限制,上述实施例和说明书中描述的只是说本发明的原理,在不脱离本发明精神和范围的前提下,本发明还会有各种变化和改进,这些变化和改进都落进要求保护本发明范围内。本发明要求保护范围由所附的权利要求书及其等效物界定。序列表<110>扬州大学<120>整合四拷贝功能性f18菌毛操纵子基因的益生菌克隆株、构建方法及应用<130>xhx2018112604<141>2018-11-26<160>48<170>siposequencelisting1.0<210>1<211>60<212>dna<213>escherichiacoli<400>1agtcctaggtataatactagtggatatgtggggtaacgacggttttagagctagaaatag60<210>2<211>60<212>dna<213>escherichiacoli<400>2agtcctaggtataatactagtgttcgcagtggaagtaccgtgttttagagctagaaatag60<210>3<211>60<212>dna<213>escherichiacoli<400>3agtcctaggtataatactagtctggggaatgaatcaggccagttttagagctagaaatag60<210>4<211>60<212>dna<213>escherichiacoli<400>4agtcctaggtataatactagtgagggtgagccataatgaaggttttagagctagaaatag60<210>5<211>36<212>dna<213>escherichiacoli<400>5actagtattatacctaggactgagctagctgtcaag36<210>6<211>40<212>dna<213>escherichiacoli<400>6ttctctagagtcgacctgcagctcgttccgctcacacttc40<210>7<211>42<212>dna<213>escherichiacoli<400>7ataatggtttcttagacgtcatcctggccgcgcacctgatac42<210>8<211>42<212>dna<213>escherichiacoli<400>8cagggtaatagatctaagcttacattaatttggggttacgat42<210>9<211>23<212>dna<213>escherichiacoli<400>9gggtgaccaggtcgcgaagaatc23<210>10<211>25<212>dna<213>escherichiacoli<400>10gacgtctaagaaaccattattatca25<210>11<211>47<212>dna<213>escherichiacoli<400>11tcttcgcgacctggtcacccttagaattactgtatctcgaaaacaat47<210>12<211>42<212>dna<213>escherichiacoli<400>12ttctctagagtcgacctgcagatcgaaaacaatgcgcgtctg42<210>13<211>39<212>dna<213>escherichiacoli<400>13ataatggtttcttagacgtccaccatgtagctgcggtct39<210>14<211>21<212>dna<213>escherichiacoli<400>14actgcgaacaaaattgttggt21<210>15<211>42<212>dna<213>escherichiacoli<400>15cagggtaatagatctaagcttatgtaggcttcgttaaaggtt42<210>16<211>47<212>dna<213>escherichiacoli<400>16ccaacaattttgttcgcagtttagaattactgtatctcgaaaacaat47<210>17<211>47<212>dna<213>escherichiacoli<400>17ccaacaattttgttcgcagtttagaattactgtatctcgaaaacaat47<210>18<211>40<212>dna<213>escherichiacoli<400>18ataatggtttcttagacgtcttcattccccagcgaccaga40<210>19<211>22<212>dna<213>escherichiacoli<400>19gcgctaatcacgacgcgctgta22<210>20<211>43<212>dna<213>escherichiacoli<400>20cagggtaatagatctaagcttcggataaacggaactggaaata43<210>21<211>47<212>dna<213>escherichiacoli<400>21cagcgcgtcgtgattagcgcttagaattactgtatctcgaaaacaat47<210>22<211>42<212>dna<213>escherichiacoli<400>22ttctctagagtcgacctgcagaaccacagacgaatcagcatg42<210>23<211>41<212>dna<213>escherichiacoli<400>23ataatggtttcttagacgtcagaatttcccgcctgagcagt41<210>24<211>21<212>dna<213>escherichiacoli<400>24gtttcttgctggtggctaaaa21<210>25<211>42<212>dna<213>escherichiacoli<400>25cagggtaatagatctaagcttaaagctgccgccgattagccc42<210>26<211>47<212>dna<213>escherichiacoli<400>26ttttagccaccagcaagaaattagaattactgtatctcgaaaacaat47<210>27<211>22<212>dna<213>escherichiacoli<400>27aagcttagatctattaccctgt22<210>28<211>21<212>dna<213>escherichiacoli<400>28ctgcaggtcgactctagagaa21<210>29<211>20<212>dna<213>escherichiacoli<400>29ataggttaatgtcatgataa20<210>30<211>20<212>dna<213>escherichiacoli<400>30cctcttcagcacaaatgcct20<210>31<211>22<212>dna<213>escherichiacoli<400>31tatttcatgtattaatttgctg22<210>32<211>21<212>dna<213>escherichiacoli<400>32attgttttcgagatacagtaa21<210>33<211>21<212>dna<213>escherichiacoli<400>33aactgtgataaactaccgcat21<210>34<211>20<212>dna<213>escherichiacoli<400>34cccatccgtgaacttcatcg20<210>35<211>19<212>dna<213>escherichiacoli<400>35cgtttacctgcattgcctt19<210>36<211>20<212>dna<213>escherichiacoli<400>36cttaccatagggccgatcca20<210>37<211>21<212>dna<213>escherichiacoli<400>37tagatttcatacacggtgcct21<210>38<211>21<212>dna<213>escherichiacoli<400>38cagcgatttccaagttaccac21<210>39<211>21<212>dna<213>escherichiacoli<400>39taccgttagccagagttgtcc21<210>40<211>21<212>dna<213>escherichiacoli<400>40cttaccatagggccgatccaa21<210>41<211>20<212>dna<213>escherichiacoli<400>41gatcgggcagcgtaatttca20<210>42<211>20<212>dna<213>escherichiacoli<400>42gctccaagttatatcagctg20<210>43<211>21<212>dna<213>escherichiacoli<400>43atacattttcagcacctagcg21<210>44<211>21<212>dna<213>escherichiacoli<400>44tatagatttgccctgactcca21<210>45<211>23<212>dna<213>escherichiacoli<400>45ggatatgtggggtaacgacgcgg23<210>46<211>23<212>dna<213>escherichiacoli<400>46gttcgcagtggaagtaccgttgg23<210>47<211>23<212>dna<213>escherichiacoli<400>47ctggggaatgaatcaggccacgg23<210>48<211>23<212>dna<213>escherichiacoli<400>48gagggtgagccataatgaagtgg23当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1