一种杨树抗旱基因及其应用的制作方法
本发明属于分子生物学技术领域,特别涉及一种杨树抗旱基因及其应用。
背景技术:
森林覆盖世界陆地总面积的30%左右,是地球陆地生态系统的主要组成部分,也是重要的可再生自然资源,对人类生活、自然界的碳循环及维持生态平衡都具有不可替代的地位和作用。在自然条件下,林木植物在生长发育过程中经常会遭受到许多非生物逆境如干旱、水渍、盐渍、低温、高温等伤害,对林木植物的生长发育、木材产量和材质等产生不利影响。其中干旱是环境胁迫中最普遍的逆境因子。全球性干旱问题日趋严重,干旱和半干旱地区面积不断增加,已经对世界范围的林业生产及生态环境造成了不利影响。受全球气候变暖的影响,干旱事件的频次和强度显著增加,已经对森林生态系统的健康,分布,结构,组成和生态多样性造成了巨大的影响。据不完全统计,近50多年来超过百分之七十的林木树种在水源供给不足后生长会特别容易受到侵害。研究人员预测,由于全球气候变暖和大面积森林的砍伐,全球性干旱问题及其对森林的影响还会更加严重,全球多数森林生态系统将面临着“干旱致死”的潜在风险。
林木植物在长期的进化和演变过程中,逐渐形成了一套感受和传导水分胁迫信号的系统,并形成一系列生理和发育的机制来应对环境中的缺水胁迫,最大限度地减轻干旱造成的伤害。林木是多年生乔木树种具有高大的主干和强大根系,对于干旱胁迫的响应和适应具有极其复杂的调控过程,包含形态,生理、生化和分子水平多层面的调控机制。尤其是强大的根系对于提高杨树的耐旱能力具有重要的作用。近年来,植物响应水分胁迫的研究在基因表达调控、蛋白修饰和相互作用、信号传递网络等方面取得了许多新的发现和突破。主要集中在bzip、nac、myb、wrky等几个大转录因子基因家族在干旱胁迫下的调控作用,而在其他转录因子家族中关于调控抗干旱的研究,尤其是调控根的生长和发育的转录因子则相对较少。
参考文献
(1)jiab,zhoug,wangf,etal.partitioningrootandmicrobialcontributionstosoilrespirationinleymuschinensispopulations[j].soilbiology&biochemistry,2006,38(4):653-660.
(2)sluiter,a.,hames,b.,ruiz,r.,scarlata,c.,sluiter,j.,templeton,d.,andcrocker,d.(2008)determinationofstructuralcarbohydratesandlignininbiomass.nationalrenewableenergylaboratory,golden,co.
(3)chenty,wangb,wuyy,etal.structuralvariationsofligninmacromoleculefromdifferentgrowthyearsoftriploidofpopulustomentosa,carr[j].internationaljournalofbiologicalmacromolecules,2017,101.
(4)wen,j.-l.;sun,s.-l.;xue,b.-l.;sun,r.-c.recentadvancesincharacterizationofligninpolymerbysolution-statenuclearmagneticresonance(nmr)methodology.materials2013,6,359-391.
技术实现要素:
为了解决现有技术中的不足,本发明提供了一种新的杨树抗旱基因pdnfy-b21,特异性的调控根的生长和发育,转基因杨树可以显著提高杨树的耐旱性。
较为具体地,本发明第一方面提供了一种杨树蛋白,所述蛋白的氨基酸序列如seqidno.6所示。
本发明第二方面提供了一种杨树基因,所所述基因的核酸序列编码如seqidno.6所示的氨基酸序列。
在一些实施方式中,所述基因的核酸序列如seqidno.5所示。
本发明第三方面提供了一种基因工程表达载体,所述表达载体包含如本发明第二方面所述的杨树基因,优选,所述基因工程表达载体的基础载体为pbi121质粒。
本发明第四方面提供了一种重组农杆菌,所述重组农杆菌包含如本发明第三方面所述的表达载体,优选,所述农杆菌为gv3101。
本发明第五方面提供了一种制备转基因杨树的方法,所述方法为将本发明第四方面所述的重组农杆菌转化到杨树当中,形成所述转基因杨树。
在一些实施方式中,所述方法包括如下步骤:
(1)菌液活化:将本发明第四方面所述的重组农杆菌的菌液在含利福平和卡那霉素的yeb液体中培养,形成第一混合物;将所述第一混合物离心,弃上清,形成第一沉淀;用添加乙酰丁香酮的wpm培养液重悬所述第一沉淀,形成第二混合物;
(2)农杆菌侵染:将无菌杨树组培苗幼嫩的叶片放入所述第二混合物中浸染;
(3)共培养:吸干浸染后叶片上的液体成分,将所述叶片置入wpm共培养基,暗培养,;
(4)选择培养:
将所述共培养的叶片转移到wpm选择培养基上,在光照下培养,形成抗性不定芽;
(5)筛选生根培养:
将所述抗性不定芽转入wpm生根培养基中培养,形成生根杨树幼苗。
本发明第六方面提供了一种用于检测转基因杨树的pcr引物对,所述引物对包括第一上游引物和第一下游引物,
所述第一上游引物序列为:
5′-agtggattgatgtgatatctccactga-3′,如seqidno.9所示;
所述第一下游引物序列为:
5′-ttatgatagggttgctaaaagtttagcactgt-3′,如seqidno.4所示。
本发明第七方面提供了一种用于检测或监测杨树抗旱性的荧光pcr引物对,所述引物对包括第二上游引物和第二下游引物,
所述第二上游引物序列为:
5'-gatgatttgctttgggctatggctac-3',如seqidno.1所示;
所述第二下游引物序列为:
5'-tccagtcttcgcagatcccttggt-3',如seqidno.2所示。
本发明第八方面提供了一种用于检测或监测杨树抗旱性的试剂盒,所述试剂盒包括如本发明第七方面所述的pcr引物对。
本发明第九方面提供了一种本发明第一方面所述蛋白,如本发明第二方面所述的基因,如本发明第三方面所述的基因工程表达载体,如本发明第四方面所述的重组农杆菌,或如本发明第五方面所述的制备转基因杨树的方法在改良杨树性状或改良所述杨树性状的育种中的用途,所述杨树性状包括:
(a)抗干旱性;
(b)二氧化碳同化率;
(c)生长速度;
(d)根的呼吸速率;
(e)根导水率;
(f)根的生长;(g)根的木质素浓度和结构;
优选,所述杨树为84k杨树或黑杨ne19。
与现有技术相比,本发明具有以下优点:
本发明是国内外首次发现了杨树基因pdnfy-b21序列的抗旱功能,它是广泛存在的核转录因子,前人研究显示nf-yb转录因子对于提高植物的抗旱性有着重要作用,目前在林木植物中过量表达可提高林木植物抗旱性的功能没有相关报道。本发明实验结果表明转基因杨树pdnfy-b21的表达量与受体对照(非转基因植株)相比均有很大程度的提高,其抗旱性也因此得到增强。因此本发明提出可利用pdnfy-b21的过量表达来增强植物抗旱性以便用于林业的抗旱品种选育,由此减缓干旱逆境对林业产量和品质造成的伤害。
附图说明
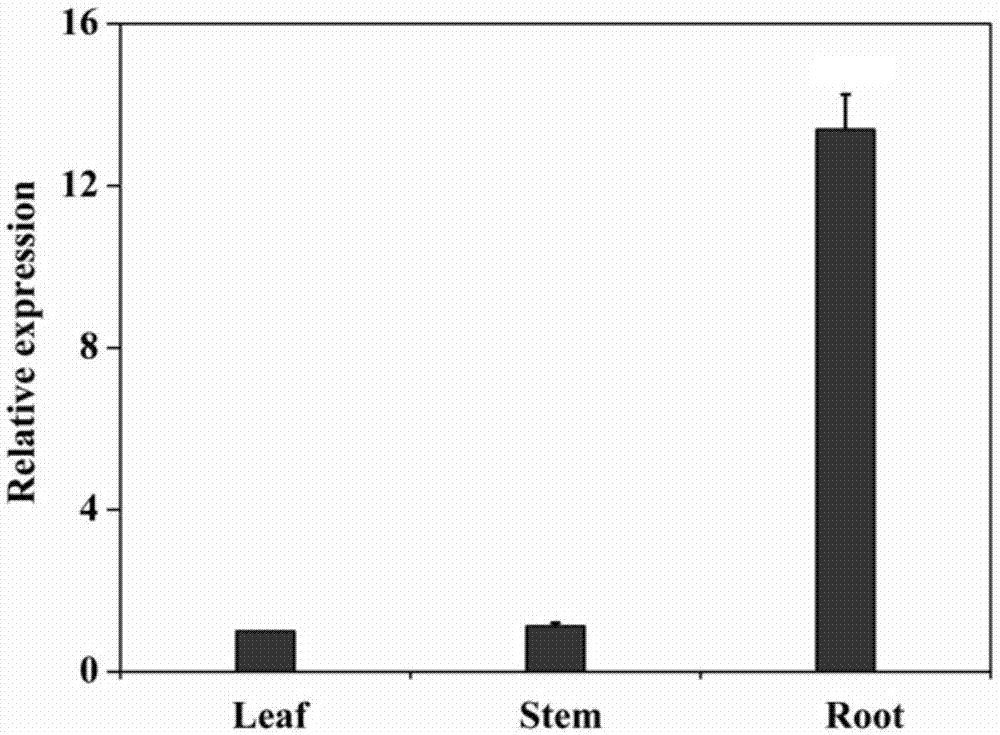
图1为pdnfy-b21基因在黑杨ne19中组织特异性表达柱状图,其中,以叶中的表达量为基准,数值为1;
图2为pdnfy-b21基因在黑杨ne19中根组织中干旱胁迫下表达柱状图,其中,以0d表达量为基准,数值为1;
图3为pdnfy-b21基因的核酸碱基序列示意图;
图4为pdnfy-b21基因的蛋白质氨基酸序列示意图;
图5为pbi121-35s:pdnfyb21:gus载体构建流程意图;
图6为pdnfy-b21基因转基因植株筛选照片;
图7为pdnfy-b21基因转基因植株dna片段电泳图;
图8为pdnfy-b21基因转基因植株与野生型叶片gus染色照片对照;
图9为pdnfy-b21基因转基因植株与野生型pdnfy-b21基因表达量对照柱状图,其中,以tw的表达量为基准,数值为1;
图10为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8抗旱效果对照照片;
图11为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下净二氧化碳同化速率对照曲线图;
图12为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下茎高对照曲线图;
图13为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根的大小(根长,根径和根的体积)对照柱状图(每簇从左到右依次为野生型、#6、#8),其中,分别以wt的量为基准,数值为1;
图14为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根密度对照柱状图;
图15为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根冠比对照柱状图(每簇从左到右依次为野生型、#6、#8);
图16为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根电导率对照柱状图;
图17为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下生物量对照柱状图(每簇从左到右依次为野生型、#6、#8);
图18为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根系呼吸速率对照曲线图;
图19为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根导水率对照曲线图;
图20为野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在干旱胁迫环境下根木质素含量对照柱状图;
图21为基因野生型植株与pdnfy-b21转基因植株oxpdnfy-b21#6、#8在水势0.12mpa环境下根木质素的浓度和结构图。
具体实施方式
为使本发明的目的、技术方案和优点更加清楚,下面将结合附图对本发明实施方式作进一步地详细描述。
本发明中所用的术语“转基因杨树”是指含有导入的基因并能够稳定地增强所导入的基因表达并产生具有特定的生物学性状的杨树。
通常按照常规条件如sambrook等人,分子克隆:实验室手册(newyork:coldspringharborlaboratorypress,1989),或draper等人(blackwell科学出版社,1988)所述的条件,或参考所用试剂说明书所建议的条件。
本发明中发现的杨树抗旱转录因子蛋白pdnfy-b21特异性地调控杨树根的生长和发育从而提高杨树的耐旱能力,目前为止,包括在杨树中都还未有任何相关功能的报道,是一个未知功能的蛋白和基因。
本发明的目的是在于提供了一种杨树(populus)抗旱基因pdnfy-b21,该基因的核苷酸序列和蛋白序列为图3和图4所示。通过将杨树抗旱基因pdnfy-b21在84k杨中的超表达,发现此基因可显著提高杨树的抗旱性,干旱胁迫40天后转基因杨树能正常生长,并且木材产量基本不受影响,为干旱逆境下提高木材的产量和材质提供保证。
本发明的另外一个目的是在于提供了一种杨树抗旱基因pdnfy-b21在林木抗旱育种中的应用,提高了林木对干旱的耐受能力,保证了林木的木材产量和材质免受或少受干旱胁迫的不利影响。
为了实现上述发明目的,本发明采用以下技术措施:
一种杨树抗旱基因pdnfy-b21,其制备步骤是:
一种黑杨ne19[p.nigra×(p.deltoides×p.nigra)]抗旱基因pdnfy-b21,获得:为了揭示nfy-b家族基因在杨树中是否能够响应干旱胁迫,本申请的发明人根据毛果杨的ptnfy-b21(potri.016g085000.1)基因序列设计了如seqidno.1、seqidno.2所示一对针对黑杨ne19的命名为pdnfy-b21基因的荧光定量pcr引物,验证了pdnfy-b21基因在杨树中的根特异性表达以及干旱应激高表达,进一步设计基因编码序列的如seqidno.3、seqidno.4所示两侧引物。以黑杨ne19根的cdna第一链为模板利用pcr技术克隆得到该目的基因的cds序列并进行测序,获得杨树pdnfy-b21基因序列。
一种杨树抗旱基因pdnfy-b21在杨树抗旱育种中的应用,其应用过程是:
(1)使用xbai和smai酶对空载体pbi121质粒进行双酶切,另外设计加有xbai和smai酶酶切位点的引物(seqidno.7、seqidno.8所示),通过pcr反应将杨树抗旱基因pdnfy-b21构建到pbi121表达载体上,命名为pbi121-pdnf-yb21。
(2)将步骤(1)中制备的载体pbi121-pdnf-yb21转入根癌农杆菌gv3101(invitrogen),再通过农杆菌介导的叶盘转化法将pdnfy-b21基因转化进84k杨。
(3)筛选阳性植株。
我们通过叶盘转化法对84k杨的叶片进行侵染,在卡纳霉素筛选压力下获得了可能为转pdnfy-b21基因的阳性植株16株,进一步对阳性植株的叶片dna提取后进行pcr鉴定发现有9个株系出现了约600bp大小的特异性条带,而非转基因株系中则没有。gus染色发现pcr鉴定的9个株系的叶片都呈蓝色。最终确认该9个植株为基因已入的阳性植株。通过荧光定量pcr对插入的pdnfy-b21基因的表达量进行检测,发现9个阳性株系并不相同,其中oxpdnfy-b21#6,oxpdnfy-b21#8的表达量最高,因此在后续实验中,选取这两个株系做为实验材料。
克隆本发明中所述的抗旱蛋白基因pdnfy-b21的方法是本领域中所常采用的方法。提取杨树叶片dna是常用的分子生物学技术,提取mrna的方法也有多种成熟的技术,试剂盒(trizolreagent)可从商业途径获得(invitrogen公司),而构建cdna文库也是常用的分子生物学技术。构建本发明中所述的载体构建和将基因转染入杨树所用到的酶切、连接、叶盘转化法等方法也是本领域中常用技术。其中所涉及的质粒,转染用媒体(如根癌农杆菌gv3101和所用试剂成分如蔗糖、植物激素等)可从商业途径获得。
实施例1:杨树抗旱基因pdnfy-b21的筛选:
近年来,植物响应干旱胁迫的研究在基因表达调控、蛋白修饰和相互作用、信号传递网络等方面取得了许多新的发现和突破。主要集中在bzip、nac、myb、wrky等几个大转录因子基因家族在旱胁迫下的调控作用,而在其他转录因子家族中关于调控抗干旱的研究,尤其是调控根的生长和发育的转录因子则相对较少。本发明中发现的杨树抗旱转录因子蛋白pdnfy-b21特异性地调控杨树根的生长和发育从而提高杨树的耐旱能力,目前为止,包括在杨树中都还未有任何相关功能的报道,是一个未知功能的蛋白和基因。
转录因子在植物响应和调控水分胁迫方面发挥着重要的作用。为了揭示nfy-b家族基因在杨树中是否能够响应干旱胁迫,本申请的发明人根据毛果杨的ptnfy-b21(potri.016g085000.1)基因序列设计了如下所示一对针对黑杨ne19的命名为pdnfy-b21基因的荧光定量pcr引物。进行如下两个测试。
上游引物序列为:
5'-gatgatttgctttgggctatggctac-3',如seqidno.1所示;
下游引物序列为:
5'-tccagtcttcgcagatcccttggt-3',如seqidno.2所示。
1.组织特异性测试
取长势一致的一年生的黑杨ne19茎段(~15cm)扦插在盆中,每10天每盆浇水1l,生长3个月后,将其从盆中拔出,用水轻轻把根的土清洗干净,取其根,茎,叶,立即放入液氮中,然后放-80℃冰箱为pdnfy-21基因的荧光定量分析备用。
然后使用seqidno.1和seqidno.2所示的引物对pdnfy-b21在根、茎、叶中进行荧光定量表达分析,发现pdnfy-b21特异性在根中表达,结果参见图1。
2.时间特异性测试
为了排除生长期和生长环境因素对干旱胁迫效果的影响,取五组杨树苗分别依次在加入霍格兰(hoagland's)营养液(无peg6000)培养后再加入含有30%peg6000(w/v)(sigma)的霍格兰(hoagland's)营养液进行培养,总共20天,都在第20天做测试。
具体方案为:取长势一致的一年生的黑杨ne19茎段(~15cm)扦插在盆中,生长3个月后开始试验,将试验用杨树苗分成5组,从盆中拔出杨树苗,用水轻轻把根的土清洗干净,放在加入霍格兰(hoagland's)营养液(无peg6000)里依次培养20,15,10,5,0天,然后放在加入30%peg6000(w/v)(sigma)的霍格兰(hoagland's)营养液里依次培养0,5,10,15,20天。在20天后将五组杨树苗的根分别快速取下(peg6000模拟干旱胁迫,第一组相当于未做干旱胁迫的对照组),立即放入液氮中,然后放-80℃冰箱为荧光定量分析备用。
使用seqidno.1和seqidno.2所示的引物,通过荧光定量表达分析pdnfy-b21的相对表达量,结果参见图2。从图2中可以看出,相对于未做干旱胁迫的对照组来说,pdnfy-b21在干旱胁迫处理5-15天中pdnfy-b21的表达逐渐增高,在处理20天后,pdnfy-b21的表达量回落,仍略高于前述对照。
图2所示结果可以说明pdnfy-b21在一定程度上是干旱胁迫诱导提高表达的,这种机制用于杨树遇到干旱胁迫下自我保护。胁迫超过20天后表达有所下降,这可能是因为本实验检测的是rna的表达,而蛋白的表达具有时间累积性,此时pdnfy-b21的蛋白的累积已经达到有效抵抗干旱的效果,或者pdnfy-b21的蛋白参与的信号通路和/或代谢调控网络已经通过代偿等机制对干旱胁迫有了一定的适应。也可能是pdnfy-b21基因的表达已经修复了干旱胁迫的损伤。
基于上述发现,本发明接下来针对pdnfy-b21基因进行了多方面的研究。
实施例2:杨树抗旱基因pdnfy-b21的克隆:
根据公布的毛果杨基因组数据寻找与黑杨ne19的pdnfy-b21序列同源的毛果杨基因序列ptnfy-b21(potri.016g085000.1),设计基因编码序列的两侧引物,正向引物:5′-atggcggcagaggcaccggc-3′(seqidno.3);反向引物:5′-ttatgatagggttgctaaaagtttagcactgt-3′(seqidno.4)。
1、提取杨树mrna
rna的提取(使用trizoltmkit提取rna)。
液氮研磨100mg黑杨ne19根,茎,叶(重量比1:1:1)混合样本。
a.加1mltrizol,室温(22-25℃,下同)放置5min。
b.加入200μl氯仿,剧烈振荡30s,室温放置2min。
c.13000rpm,离心15min,4℃,取上清转移至新管中,加入500μl异丙醇,混匀后室温放置15min。
d.13000rpm,离心15min,4℃,去上清,加入1ml70%(无水酒精与h2o的体积比)乙醇。
e.7500rpm,离心7min,4℃,去上清,空气中干燥。
f.用depc处理过的去离子水溶解rna,放到-80℃超低温冰箱中保存备用。
2、cdna第一链的反转录,采用天根公司的tianscriptiicdna试剂盒,操作参照所用试剂盒说明进行操作。
3、以cdna为模板进行pcr扩增,pcr扩增产物经测序得到了一种杨树抗旱基因pdnfy-b21(经生工生物工程(上海)股份有限公司测序),其核苷酸编码序列为图3所示(seqidno.5),该核苷酸序列编码的蛋白序列如图4(seqidno.6)所示。
pcr扩增杨树基因pdnfy-b21的50μl反应体系如下:
反应的时间和温度如下:
94℃3min
94℃30s
59℃30s
72℃1min,34cycles
72℃10min
实施例3:pdnfy-b21表达载体的构建及农杆菌的转化:
其他pcr步骤与实施例2相同,区别在于pcr引物包含xbai和smai酶酶切位点,具体为:
正向引物:
5′-tctagaatggcggcagaggcaccggc(seqidno.7);
反向引物:
5′-cccgggttatgatagggttgctaaaagtttagcactgt-3′(seqidno.8)。
以seqidno.7和seqidno.8所示序列为引物,通过pcr扩增得到的杨树抗旱基因pdnfy-b21扩增产物,通过使用xbai和smai酶对上述pcr扩增产物与空载体pbi121质粒(参见图5)分别进行双酶切;通过dna连接酶连接反应将杨树抗旱基因pdnfy-b21构建到pbi121表达载体上,得到重组载体pbi121-pdnf-yb21,并将重组载体转化到感受态农杆菌细胞gv3101得到含有基因pdnfy-b21的重组农杆菌(参考sambrook等人,分子克隆:实验室手册(newyork:coldspringharborlaboratorypress,1989),卡那霉素筛选,其插入片段经载体引物(35s启动子序列引物)与杨树pdnfy-b21基因下游引物pcr鉴定后,用正确重组了pdnfy-b21的农杆菌转化杨树。
实施例4:84k杨的抗旱基因pdnfy-b21转化
一、杨树培养基配制
wpm重悬液:wpm+30g/l蔗糖+100mol/las;
wpm共培养基:wpm+30g/l蔗糖+100μmol/las+2.0mg/l6-ba+0.3mg/lnaa+5.5-6.0g/l琼脂;
wpm选择培养基:wpm+30g/l蔗糖+100mg/lkana+2.0mg/l6-ba+0.3mg/lnaa+400mg/lcef+5.5-6.0g/l琼脂;
wpm筛选生根培养基:wpm+30g/l蔗糖+150mg/lkana+0.05mg/liba+0.05mg/lnaa+400mg/lcef+5.5-6.0g/l琼脂;在除琼脂以外的成分混合好之后,通过酸度计调ph为5.80~5.85,加入琼脂,121℃高温高压蒸汽灭菌20min。
其中,wpm为:(woodyplantmedium);as为:乙酰丁香酮;6-ba为:6-苄氨基腺嘌呤;naa为:萘乙酸;kana为:卡那霉素;cef为头孢霉素;iba为吲哚丁酸。
二、转化步骤
(1)菌液活化:从-80℃冰箱中取出含有基因pdnfy-b21的重组农杆菌,取200μl菌液接种于20mlyeb液体培养基中(含100mg/l利福平+100mg/l卡那霉素),28℃摇床中培养48h。待菌液呈金黄色时,取5ml菌液接种于500mlyeb液体培养基中(含100mg/l利福平+100mg/l卡那霉素),置于28℃摇床中培养至菌液od600=0.6~0.8。将菌液在18℃下12,000r/min离心2min,弃上清。用添加乙酰丁香酮(100μmol/l)的wpm液体培养基(即wpm重悬液)重悬后即可用于转化。
(2)农杆菌侵染
选取无菌84k杨组培苗幼嫩的叶片,在无菌条件下,将其切成0.5×0.5cm2的小块,将其放入经wpm重悬液重悬好的菌液中浸染10-15min,且浸泡过程中缓慢的摇晃。
(3)共培养
用无菌的滤纸将浸染过的叶片表面的菌液吸干,将叶片平铺回wpm共培养基上,25℃±2℃,暗培养2d,形成经过共培养的外植体。
(4)选择培养
将经过共培养的外植体转移到wpm选择培养基上,在25℃、光照为2000-10000lux条件下培养2-3周其间每10d更换一次培养基。
(5)筛选生根培养
当抗性不定芽长至约2cm时,切下并转入wpm生根培养基中,10d左右即生根。
参见图6。
(6)转基因84k杨的移栽
待生根幼苗长(30天左右)至约8cm时取出幼苗,轻轻洗净根部琼脂,移栽至温室土壤中培养,并用保鲜膜罩上,人工模拟自然条件光照(光照16小时,暗8小时),10天后揭膜。温室条件:相对湿度~45%,恒温20-24℃,光照周期为8h黑暗16h光照培养。按照上述方法制备了9个转基因株,分别标号为#1、#2、#3、#4、#5、#6、#8、#12、#16。
实施例5:转基因杨树的鉴定
(1)pcr鉴定
i.用于pcr的野生型84k杨和转pdnfy-b21基因84k杨总dna的提取
采用改良的ctab法提取杨树dna(saghaimaroofetal.1984),方法如下:
a.取~0.6g新鲜叶片于10ml离心管中,在液氮中研磨成粉;立即加入3ml1%ctab和90μlβ-巯基乙醇(提前预热),65℃水浴45min(前30min每10min混匀一次),取出放至室温;
b.加入与ctab等体积的氯仿:异戊醇(v/v为24:1),剧烈震荡后平放10min;16℃,10000r/min,离心10min;取上清于另以离心管中,重复8次;
c.取上清2ml到10ml离心管中,加入等体积预冷的异丙醇,轻轻晃动至出现絮状沉淀;用枪头小心吸出絮状沉淀于1.5ml离心管中,用75%乙醇水溶液漂洗两次;100%乙醇漂洗一次,在37℃烘箱中烘干;加入50μlddh2o,1μlrnase,37℃处理1.5h;-20℃保存。
ii.以转pdnfy-b21基因84k杨与未转基因植株的基因组dna为模板,在35s启动子序列及pdnfy-b21基因序列上设计引物,进行pcr反应,目的片段大小约为600bp。设计的引物如下表:
引物名称引物序列
35s上游引物5’agtggattgatgtgatatctccactga3’(seqidno.9)
pdnfy-b21下游引物5′-ttatgatagggttgctaaaagtttagcactgt-3′(seqidno.4)
检测结果显示,转化杨树均能够扩增出预期大小的电泳条带,与作为阳性的重组质粒电泳条带的大小一致,而阴性对照非转基因植株的dna则没有,表明转基因杨树基因组中已经含有外源基因dna片段结果如图7示。
其中,m代表dnamarkers;pc代表带有pbi121-pdnf-yb21质粒的阳性对照(实施例3中的pbi121重组质粒);wt代表野生型84k杨树样本;#1、#2、#3、#4、#5、#6、#8、#12、#16分别代表相应转基因植株。
(2)gus染色检测
x-gluc是gus基因检测的底物,gus(β-葡萄糖苷酶)可将x-gluc水解为蓝色物质,此物质不溶于植物细胞组织,因此gus活性部位就会显示蓝色,可实现植物gus融合蛋白的快速检测。
本实验构建的表达载体将pdnfy-b21基因与载体上的gus序列一起设计为融合蛋白,因此可使用gus染色间接反映转基因植物pdnfy-b21基因的表达情况。实验的具体操作如下:
a.gus染液的配制:
50mm磷酸钠缓冲液+10mmna2edta,0.5mmk4[fe(cn)6]·3h2o+0.5mmk3[fe(cn)6],0.1%tritonx-100,和1mg/mlx-gluc,ph7.0(所述浓度指相应成分整个溶液中的浓度)。
b.剪取杨树叶片放进干净管中,并加gus染液,使叶片完全浸没于gus染色液中,于37℃黑暗条件下染色12小时,然后用75%酒精脱色20min。最后转移到固定液(水合氯醛:水:甘油;8:3:1;w/v/v)中进行固定2h后进行观测和拍照。
gus染色结果显示,转基因杨树的叶有蓝色出现,而酒精脱色后的野生型杨树材料则是无色透明的,结果如图8示。
其中,wt代表野生型;#1、#2、#3、#4、#5、#6、#8、#12、#16分别代表相应转基因植株。
(3)荧光定量检测
提取转pdnfy-b21基因84k杨与未转基因植株的叶片rna,检测各株系间nfy-b21基因的表达量差异。
a.目标基因的荧光定量检测
根据实施例2所测序得到的pdnfy-b21基因的编码序列(seqidno.5),使用primer5软件设计荧光定量引物。
将各组样品反转录好的cdna稀释至100ng/μl,做为模板。使用天根公司的fastfireqpcrpremix(sybrgreen)试剂盒进行荧光定量反应。20μl反应体系包括:
荧光定量pcr反应条件为95℃15min,95℃10s,60℃30s,40个循环。以actin基因为内参基因,每个样品技术重复4次,用2-δδct法计算荧光定量的结果。
荧光定量pcr结果显示9个阳性株系的pdnfy-b21的表达量并不相同,其中oxpdnfy-b21#6,oxpdnfy-b21#8的表达量最高,因此在后续实验中,选取这两个株系做为实验材料。结果如图9示。wt代表野生型;#1、#2、#3、#4、#5、#6、#8、#12、#16分别代表相应转基因植株。
实施例6转基因杨树性能测试
一、杨树植株抗旱性实验
长期干旱处理条件
在组培苗移入土中培养1个月后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤含水势为-1.82mpa做为干旱环境,进行40天长期干旱处理。使用psyprowaterpotentialsystem(wescor,utah,usa)测量土壤盆里的土壤的水势。每天下午4:00-5:00浇水维持土壤水势-1.82mpa(模拟缺水条件,干旱状态),保持各盆在相应的土壤水势条件下40天。由图10可见(其中三株杨树苗从左到右分别依次代表野生型、#6、#8,保留塑料盆的一部分是为了指示照片的相对尺寸),干旱40天后野生型84k杨叶片大多开始变黄,而转化植株oxpdnfy-b21#6、#8有发达的根系无明显影响。因此说明转基因杨树抗旱性均明显增强。
二、杨树二氧化碳同化测试
在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,土壤水势采用psyprowaterpotentialsystem(wescor,utah,usa)测定,每隔5天进行二氧化碳同化试验,采用光合仪li-cor6400(lincoln,ne,usa)仪测定二氧化碳同化率。同结果参见图11。
由图11可见,oxpdnfy-b21#6、#8两株系二氧化碳同化速度几乎没有差别,二者都明显快于野生型亲本对照植株。
这说明杨树转入pdnfy-b21基因后,能够显著地增加二氧化碳同化速速,这将使该转基因杨树生长更快,杨树苗能够更快成材。该转基因杨树生长加快有利于更快地降低大气中二氧化碳的温室效应。
三、杨树茎高测试
在组培苗移入土中培养30天个后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,每隔10天进行茎高试验。结果参见图12。
由图12可见,oxpdnfy-b21#6、#8两株系茎高差别不明显,二者都明显高于野生型亲本对照植株。转基因杨树在旱胁迫环境下条件下四十天内一直在生长,而野生型在第10天就生长迟滞。这说明,在该干旱胁迫环境下,转基因杨树能够持续生长。这也与前述二氧化碳同化效果是对应的。具有优势。
四、杨树根大小测试
根常位于地表下面,吸收土壤里面的水分及溶解其中的无机盐,并且具有支持、繁殖、贮存合成有机物质的作用。在旱胁迫下,根水分的吸收和运输具有重要的作用。
在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,40天后进行根的大小(根长,根径,根体积)和每个株系根的个数测定试验。同结果参见图13(以野生型为基准,记录#6、#8的倍数)和14。
由图13和14可见,oxpdnfy-b21#6、#8两株系根的大小和每个株系根的个数几乎没有差别,二者都明显高于野生型亲本对照植株,说明在干旱的条件下转基因株系具有更发大的根系,因此在干旱条件下,转基因株系能够正常生长。
五、杨树根冠比测试
根冠比是指植物地下部分与地上部分的鲜重或干重的比值。它的大小反映了植物地下部分与地上部分的相关性,也是衡量植物抗旱性的一个重要指标。
在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,每隔10天进行根冠比试验。结果参见图15。
由图15可见,oxpdnfy-b21#6、#8两株系根冠比几乎没有差别,二者都明显高于野生型亲本对照植株,说明在干旱的条件下转基因株系具有更发大的根系和根冠比,因此在干旱条件下,转基因株系能够正常生长。
六、杨树根电导率测试
植物的质膜是活细胞与环境之间的界面和屏障干旱等不良环境,都会使质膜受到不同程度的损伤。测定质膜透性变化的最常用的方法是测定组织外渗液电导率的变化,是衡量植物抗胁迫能力强弱的重要指标之一。
在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,每隔10天采用dds-307(雷磁-dds-307a,上海)电导率仪测定根的电导率仪。同结果参见图16。
由图16可见,oxpdnfy-b21#6、#8两株系根的电导率几乎没有差别,二者都明显低于野生型亲本对照植株,说明在干旱的条件下转基因株系受到干旱的胁迫的影响要低于对照植株。
七、杨树生物量测试
在组培苗移入土中培养30天个后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤含水势为-1.82mpa,进行生物量试验。结果参见图17。
由图17可见,oxpdnfy-b21#6、#8两株系生物量几乎没有差别,二者都明显多于野生型亲本对照植株。
八、杨树根系呼吸速率测试
根的呼吸与其对矿质元素的吸收以及与其生长和分化有密切关系,其呼吸速率与合成代谢、生长速度都呈正相关。
在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,每隔5天进行根呼吸速率试验,采用光合仪li-8100a(lincoln,ne,usa)仪测定土壤的呼吸速率,通过排除法计算根系的呼吸速率(文献1)。同结果参见图18。
由图18可见,oxpdnfy-b21#6、#8两株系根的呼吸速率几乎没有差别,二者都明显高于野生型亲本对照植株。
这说明杨树转入pdnfy-b21基因后,能够显著地提高根的呼吸速率,这将使该转基因杨树生长更快,具有更发达的根系,从而提高耐旱性。
九、杨树根导水率测试
植物根系的导水率在一定程度上决定了植物的生物和发育,尤其是在水分匮乏的条件下,根系的导水率直接决定了植物的生长状态。
在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-1.82mpa40天,每隔5天进行根导水率测定试验,采用高压导水率仪hpfm-gen3(dynamax,usa)测定根的导水率,同结果参见图19。
由图19可见,oxpdnfy-b21#6、#8两株系根的导水率几乎没有差别,二者都明显高于野生型亲本对照植株。这说明杨树转入pdnfy-b21基因后,能够显著地提高根的导水率,这将使该转基因杨树具有更发达的根系,从而提高耐旱性。
十、杨树根木质素浓度和结构测试
根的木质素浓度的测实验按照参考文献(1)进行,木质素结构测试按照参考文献(2,3,4)进行,同结果参见图20和21。
根的生长和发育与木质素的浓度和结构密切相关。
由图20可见,在组培苗移入土中培养30天后,分别取野生型(亲本)、oxpdnfy-b21#6、#8株系多株,控制土壤水势-0.12mpa(水分充足)40天,取其杨树的根对其木质素的浓度和结构进行分析。由图21可见,oxpdnfy-b21#6、#8两株系根的木质素浓度和结构(s/g)几乎没有差别,二者的木质素浓度和木质素s/g的比例都明显高于野生型亲本对照植株。
通过上述对照试验数据可知,pdnfy-b21基因能够显著地增强杨树的抗旱性,提高杨树生长速度。
前述多方面的对照数据相辅相成,可以说明,当杨树转入pdnfy-b21基因后,能够显著地抵抗干旱胁迫,正常或接近正常地生长。这将有利于提高干旱地区杨树的种植,改善环境,收获树木材料,减缓温室效应。
此基因过量表达可显著提高杨树的抗旱性,在正常水分充足的条件下,与未转基因的植株相比,转pdnfy-b21杨树的根具有更高的木质素浓度和木质素s/g比例。在干旱胁迫40天,与未转基因的植株相比,转pdnfy-b21杨树能正常生长,并且木材产量和材质基本不受影响,具有较高的光合作用,根的呼吸速率,根的导水率,茎的生长速率,和发的根系,根冠比,和生物量,为干旱逆境下提高杨树的产量提供保证。
序列表
<110>北京林业大学
<120>一种杨树抗旱基因及其应用
<130>m1cncn181008
<160>11
<170>siposequencelisting1.0
<210>1
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>1
gatgatttgctttgggctatggctac26
<210>2
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>2
tccagtcttcgcagatcccttggt24
<210>3
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>3
atggcggcagaggcaccggc20
<210>4
<211>32
<212>dna
<213>人工序列(artificialsequence)
<400>4
ttatgatagggttgctaaaagtttagcactgt32
<210>5
<211>546
<212>dna
<213>杨树(populus)
<400>5
atggcggcagaggcaccggcgagtccagggggaggaagccatgagagcggagaccaaagt60
cctcgctctaattctaatgtacgtgaacaagacaggttcttaccgatcgcaaatatcagt120
aggattatgaagaaagcgctacctgctaatggaaagattgctaaggatgctaaagagact180
gttcaagaatgtgtttctgagttcattagctttatcactagcgaagcaagtgataagtgt240
cagcgagaaaagaggaagacgataaatggtgatgatttgctttgggctatggctacgtta300
gggtttgaggattatattgatcctcttaagatttacctgtctcgatacagagagatggag360
ggtgataccaagggatctgcgaagactggagatacatctgctaaaaaggatattcaccct420
ggtccaaatgcgcaggtgaaatggagctttatgttgctgagttactgccatttcttattt480
ttagtcaatcaacgtacctccttgtctttattttacagtgctaaacttttagcaacccta540
tcataa546
<210>6
<211>181
<212>prt
<213>杨树(populus)
<400>6
metalaalaglualaproalaserproglyglyglyserhisgluser
151015
glyaspglnserproargserasnserasnvalarggluglnasparg
202530
pheleuproilealaasnileserargilemetlyslysalaleupro
354045
alaasnglylysilealalysaspalalysgluthrvalglnglucys
505560
valserglupheileserpheilethrserglualaserasplyscys
65707580
glnargglulysarglysthrileasnglyaspaspleuleutrpala
859095
metalathrleuglyphegluasptyrileaspproleulysiletyr
100105110
leuserargtyrargglumetgluglyaspthrlysglyseralalys
115120125
thrglyaspthrseralalyslysaspilehisproglyproasnala
130135140
glnvallystrpserphemetleuleusertyrcyshispheleuphe
145150155160
leuvalasnglnargthrserleuserleuphetyrseralalysleu
165170175
leualathrleuser
180
<210>7
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>7
tctagaatggcggcagaggcaccggc26
<210>8
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>8
cccgggttatgatagggttgctaaaagtttagcactgt38
<210>9
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>9
agtggattgatgtgatatctccactga27
<210>10
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>10
gtcctcttccagccatctc19
<210>11
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>11
ttcggtcagcaataccagg19
- 还没有人留言评论。精彩留言会获得点赞!