水产养殖和其他动物系统中病原菌生物控制的新系统的制作方法
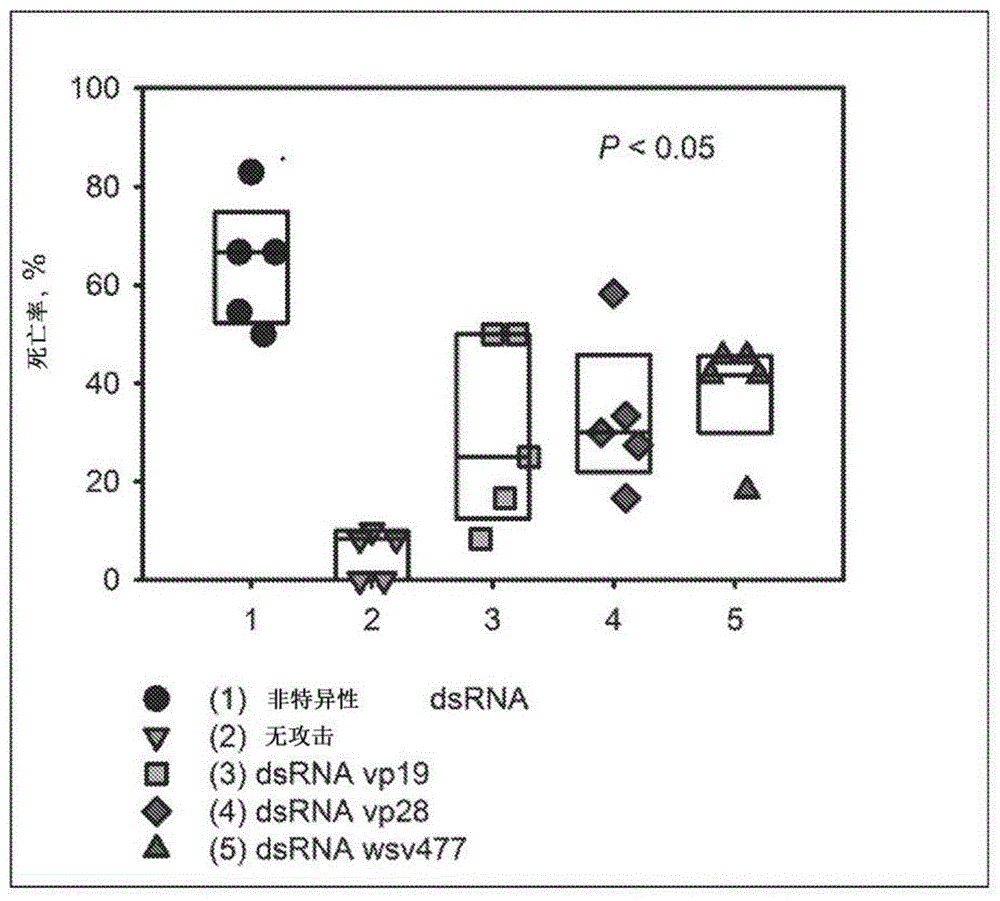
本申请要求2017年3月31日提交的美国临时申请no.62/480,138的权益和优先权。上述申请的整个说明书和附图在此通过引用整体并入。序列表本申请包含序列表,该序列表已经以ascii格式电子提交,并且其全部内容通过引用并入本文。通常,本发明的技术涉及用于动物系统中的疾病控制的新策略。具体地,本发明技术涉及用于在水生系统中生物控制水生病原体的新方法、系统和组合物。具体地,本发明可以包括用于在水产养殖系统中影响虾的疾病传播病原体的生物防治的新技术、系统和方法。
背景技术:
:水产养殖的发展使全球粮食生产发生了重大转变,远离传统的捕捞生产方法。主要由于人口增长以及传统捕捞渔业生产缺乏增长,水产养殖业迅速扩大,成为世界粮食生产生态系统的主要组成部分。水产养殖现在被视为在许多新兴经济体中发挥关键作用,因为它有助于增加粮食产量,同时有助于减少对鱼类资源的压力。正如联合国粮食及农业组织(unfao)在其2016年世界渔业和水产养殖状况报告中指出的那样,水产养殖是动物蛋白生产增长最快的领域,并且已经远远超过传统的捕捞渔业生产。例如unfao估计,水产养殖产量现在占人类消费的所有海产品的一半。全球人口的增加、海产品需求的增长以及捕捞渔业生产的限制将不可避免地导致水产养殖业的持续全球扩张及其相关的疾病出现和蔓延风险。尽管世界越来越依赖水产养殖作为粮食生产的主要来源,特别是在许多发展中经济体,但传统水产养殖系统存在若干技术和生物挑战,限制了其整体效益。水产养殖系统的一个主要缺点是水生动物通常被置于高密度生产系统中。这可导致拥挤和次优水质条件下的压力,从而易于传播疾病。特别是,水产养殖系统中的疾病暴发可能导致水生种群的大量损失,从而导致商业水产养殖中的巨大经济损失。实际上,据报道,在过去的20年中,这种疾病爆发使水产养殖业损失了数百亿美元。就虾类水产养殖而言,疾病问题尤其严重。据联合国粮食及农业组织称,尽管全球水产养殖虾产量有所增加,但由于广泛的虾病,主要产油国,特别是亚洲的产量,产量大幅下降。有下列几个原因。首先,与脊椎动物不同,虾缺乏许多适应性和先天免疫反应机制的关键组成部分,这些机制阻止了许多诱导或增强自然抗病性的传统方法。其次,大多数主要致病病毒引起非常低水平的持续性感染,这种感染可以在表观健康的虾群中发生在中度至极高的流行率。大多数虾病原体垂直传播,疾病是暴露于各种形式的环境或生理压力后大量病毒扩增的结果。压力源可能包括处理、产卵、水质差或温度或盐度的突然变化。虾病毒也可以水平传播。一旦病毒载量高且疾病明显,感染的水平传播伴随着疾病的传播。第三,虾通常同时或顺序感染多种病毒,或甚至同一病毒的不同菌株。这一事实对水产养殖系统中的诊断,检测和病原体排除提出了重大挑战。作为许多中的一个例子,白斑综合症(wss)是由白斑综合症病毒(wssv)引起的病毒性疾病。wssv是虾中的主要病原体,导致虾水产养殖的高死亡率和巨大的经济损失。wss病毒颗粒是一种非包裹的椭球形或杆状包膜颗粒,长约275nm并且宽约120nm。其环状双链dna由300kbp组成,覆盖约185个开放阅读框(orf)。wssv目前是全球甲壳类水产养殖贸易经济可持续性和增长的最重要障碍之一。自20世纪90年代初爆发以来,它造成了数十亿美元的损失。迄今为止,在亚洲、中部、南部和北美洲、欧洲和非洲等大多数主要虾产区都发现了wssv爆发。传统的预防和治疗虾类病原体(如wssv)的努力取得了有限的成功。例如,由于水产养殖系统的经济生产需求以及许多发展中国家缺乏技术专长和适当的水产养殖设施,减少环境和生理压力的努力受到限制。已经进行了其他尝试来创建和分离用于水产养殖的无病原体种群。然而,这种努力是缓慢的并且需要非常昂贵的专业知识和诊断能力。抗生素的大规模应用已经应用于虾水产养殖,特别是在生产周期期间,在幼虫和生长阶段。然而,在水产养殖中使用抗生素与环境和人类健康问题有关,包括细菌耐药性和疾病在水生环境中的持续存在。虾的可食用组织中抗生素残留物的积累也可能改变人体肠道菌群并导致食物中毒或过敏问题。最关键的是,抗生素对病毒无效。已经尝试了其他方法,例如应用免疫刺激剂或噬菌体治疗以靶向特定病原体,并且在商业和实际上取得了有限的成功。在水产养殖和其他动物系统中生物控制病原体的有效系统可以是:1)病原体(病毒)-特异性的,以便不杀死脱靶有机体;2)作用方式稳健或催化;3)在目标动物的整个发育过程中稳定且不易丢失;4)高效交付;5)管理简单,成本低;6)自我维持或再生。为此,调节病原体中基因表达的方法可能是解决上述问题的途径。通过增加表达或降低负责毒力的基因的表达来调节基因表达被认为有益于疾病的治疗。这对于已经鉴定出主要调节基因的那些疾病尤其重要。虽然大多数努力已经扩展到增强基因表达,但下调特定基因表达同样重要。由双链rna(dsrna)触发的天然存在的基因沉默机制(称为小干扰rna(sirna))已经成为在许多系统中抑制或敲低基因表达的非常重要的工具。rna干扰由dsrna触发,dsrna被rnase-iii样酶dicer切割成具有特征性5'和3'末端的21-25个核苷酸片段。这些sirna充当多蛋白复合物的指导,包括含有蛋白质argonaute2的paz/piwi结构域,其切割靶mrna。这些基因沉默机制是高度特异性和有效的,并且可能潜在地诱导整个有机体中基因表达的抑制。在过去的二十年中,基因表达技术也取得了巨大进步,包括非病毒和病毒载体的持续发展。基因表达的非病毒方法涉及使用质粒dna(pdna),其具有许多优点,包括易于使用和制备、稳定性和耐热性,以及无限大小。质粒不在哺乳动物宿主中复制并且不整合到宿主基因组中,但它们可以在宿主细胞中持续存在并且表达克隆的基因数周至数月。对使用抑制性rna分子重新产生兴趣的一个领域是传染病,特别是影响水产养殖种群的病原体。用于病原体生物控制的这种策略可以包括副转化生成和/或副转基因原理的应用。副转化生成通常指的是共生细菌或能够在足够时间内使宿主定殖以递送治疗性分子(例如dsrna)的细菌的系统,其被基因修饰并重新引入携带病原体的宿主或病原体易感群体中,例如水产养殖中的虾,它们表达效应分子。然而,激元发生有几个技术限制。例如,用于副转基因生成的细菌通常必须具有三个关键组分:实现所需效果的效应分子;显示或排泄效应分子的机制;和能够在宿主中存活足够长的细菌以产生预期量的效应分子,从而在宿主中实现所需的效果。因此,找到符合所有这些标准的这种合适的细菌是非常困难的。激元发生通常被理解为试图通过载体的共生体的转基因从载体群体中消除病原体的技术。该技术的目标是控制媒介传播的疾病。第一步是鉴定阻止载体物种传播病原体的蛋白。然后将编码这些蛋白的基因引入共生体中,以便它们可以在载体中表达。该策略的最后一步是将这些转基因共生体引入野外的载体群体中。成功进行副转化的命令的特征可包括:·共生细菌可以在体外容易地生长。·它们可以进行遗传修饰,例如通过含有所需基因的质粒转化。·工程共生体稳定且安全。·矢量和共生体之间的关联不能减弱。·现场交付易于处理。副转基因系统是可以在靶有机体中实现副转化的系统。在宿主可能拥有的许多有机体中(特别是在他们的消化系统中)鉴定对人类或动物无致病的合适的共生细菌对于转基因系统的成功是至关重要的。例如,所选择的细菌应该能够定殖各种虾类,以便它们可以部署在不同的物种和分离的菌株中。此外,精心设计的副转基因系统还必须确保效应分子不会干扰任何关键的宿主过程,例如繁殖等。这些技术和生理挑战使得转基因系统的发展极为困难。重要的是,这些技术问题使得许多转基因系统既不是有效的也不适合作为有效的生物防治策略,特别是在像虾这样的复杂有机体中。这些困难还可能阻止许多原子转移系统被适当地扩大到对环境部署有效。通常,生物防治是指利用抑制疾病的微生物来消除、控制或预防所选病原体的感染、表达和/或传播。关于水产养殖和其他动物系统中病原体生物控制的上述问题可能代表了长期需要有效、经济的解决方案。尽管可以使用实现元素,但在某种程度上可能缺少实际的尝试来满足这一需求。这可能是由于本领域普通技术人员未能充分理解或理解所涉及问题和挑战的性质而导致的。由于缺乏了解,满足这些长期需求的尝试可能无法有效解决此处确定的一个或多个问题或挑战。这些尝试甚至可能背离了本发明技术所采取的技术方向,甚至可能导致在某种程度上认为本发明技术的成就是本领域技术人员所采取的方法的意外结果。如下面将更详细讨论的,本发明的技术克服了传统病原体控制系统的局限性,同时满足了真正有效的载体生物控制策略的目标。发明概述本发明的一个目的可以包括新颖的转基因生物防治策略。在该实施方案中,本发明技术包括用于敲除必需病毒基因的各种跨王国机制。这可以通过将工程微生物引入表达特定抑制性rna分子的宿主群体中来实现,所述抑制性rna分子可以下调和/或抑制选定的病毒和/或宿主基因。本发明的另一个目的可以包括用于水产养殖种群的新型超转基因生物防治策略。在该实施方案中,本发明的技术包括用于在水产养殖系统中生长的水生动物中敲除必需病毒基因的各种跨王国机制。这可以通过将工程微生物引入表达特定抑制性rna分子的水产养殖动物种群中来实现,所述抑制性rna分子可以下调和/或抑制选定的病毒和/或宿主基因。本发明的另一个具体目的可以提供一种新颖的准转基因系统,该系统可以抑制wssv在虾类水产养殖种群中的表达和繁殖以及其他病毒病原体。该系统可以包括引入经遗传修饰的细菌,所述细菌经转化以表达可以靶向和抑制一种或多种病原体病毒基因的选定dsrna。这样的靶可以包括但不限于wssv基因组的通常保守的区域,其编码:病毒衣壳蛋白19(vp19)和/或病毒衣壳蛋白19基因(vp28)和/或早期非结构基因477(wsv477)。本发明的另一个目的可以包括靶向多个必需的病毒特异性基因靶以进行沉默的方法,从而可以有选择地减少在水产养殖环境中生长的成年虾种群中的病毒病原体。在该实施方案中,本发明可以包括含有基因修饰的细菌的饲料的产生,所述细菌被配置为表达可以靶向和抑制一种或多种病原体病毒基因的选择的dsrna。在一个实施方案中,可以将这种处理过的饲料引入对病原体敏感或受病原体影响的群体。在这样的实施方案中,可以将含有被配置为表达可以靶向和抑制一个或多个wssv基因的选择的dsrna的基因修饰的细菌的饲料引入易受wssv感染或受wssv影响的虾类水产养殖种群。由于sirna可能具有催化活性,因此其潜在的有效性和安全性可能比水产养殖业目前采用的传统病毒控制方法更好。另外,在某些实施方案中,抑制性rna是非免疫原性的,使得它们可以被设计成是物种特异性的,从而不会损害和/或影响非靶标有机体。最后,由于基于细菌的dsrna递送系统可能是自我维持且持久的,因此可能需要使用更少的饲料。在某些情况下,可能只需要一次喂食。在某些情况下,此类干扰rna分子(例如dsrna)可能充当疫苗来免疫个体动物。因此,本发明的一个目的可以包括使用基因修饰的细菌来定殖和表达dsrna分子,所述dsrna分子在水生动物例如在水产养殖系统中生长的虾种群中提供个体或畜群免疫。本发明的另一个目的可以是产生可以表达一个或多个抑制性rna分子的基因修饰的共生和/或益生菌菌株。在某个实施方案中,可以对虾益生菌如枯草芽孢杆菌进行基因修饰,以表达针对必需wssv基因的一种或多种抑制性rna分子。本发明技术的本发明的另一个目的可以包括用于通过基因工程微生物的感染将抑制性rna分子引入目标宿主的系统和方法。在一个实施方案中,本发明可以提供可以在靶有机体体内表达一种或多种抑制性rna分子的基因工程微生物。此类靶有机体通常包括水生动物、水产养殖系统中的水生动物以及其他脊椎动物和无脊椎动物。本发明的另一个目的可以包括用于创建抑制性rna分子例如dsrna的方法和组合物,其可以启动通常是通过破坏多种动物系统中细胞内特定的mrna分子而抑制或敲除必需基因表达的生物学过程。另外的实施方案可以将抑制性rna分子引入靶有机体,其抑制多种动物系统中病原体毒力所必需的基因。这样的实施方案可以包括将抑制性rna分子引入可以靶向毒力基因、病毒外壳蛋白、真菌细胞壁基因、致病成分基因、物种特异性代谢基因等的靶有机体。本发明的其他目的可以包括用于抑制性rna分子的改进的递送系统,例如通过使用稳定因子,例如稳定蛋白等。本发明技术的另一个优选实施方案可以包括改进的系统,以促进抑制性rna分子在靶有机体体内的传播。再其他的实施方案可以包括基因修饰的微生物,其可以包括可以进一步共表达具有加工酶活性的某些蛋白质的基因构建体。此类共表达的蛋白质可包括可抑制和/或增强抑制性rna分子的翻译后加工和/或修饰的酶。其他类似的实施方案可以包括将微生物引入可以表达或什至过度表达可以增强抑制性rna分子的动员的各种基因和/或可以激活可以靶向病原性途径的次级下游宿主基因的基因。最后,本发明人描述了本发明的实施方案,包括用于由表达wssv特异性dsrna的rnaseiii缺陷型枯草芽孢杆菌菌株(bg322)喂虾的方案,以及虾wssv攻击实验。本发明人证明,在该实施方案中,当给与包含bg322的饲料时,该基因修饰的细菌能够在虾肠中存活并持续存在。本发明人进一步证实,与表达非特异性dsrna的细菌喂养的对虾相比,由表达wssv靶向dsrna的细菌喂养的对虾的wssv攻击实验的死亡率计数表明第一组对虾的死亡率降低了50%。另外,本发明人显示了由wssv特异性dsrna喂养的虾中病毒复制的3-4log减少。本发明人进一步证明了在由wssv-dsrna喂养的虾中提取的总rna中存在sirna的wssv特异性部分,其中在没有由非特异性dsrna喂养的虾或没有由任何dsrna喂养的虾中不存在该sirna。综上所述,在这些实施方案中,本发明人证明了通过肠小肠定殖细菌递送病毒特异性dsrna是用于免疫和/或治疗病毒虾感染的有效平台。附图简述结合在说明书中并构成说明书一部分的附图示出了本发明的一个或多个实施例,并且与说明书一起用于解释本发明的原理。附图仅出于说明本发明的一个或多个优选实施例的目的,并且不应被解释为限制本发明。在图中:图1:用表达对wssv特异的非特异性dslucdsrna或dsrna的bg322饲喂的虾在病毒攻击的最后一天的总死亡率:i)dsvp19(seqidno.3);ii)dsvp28(seqidno.2);iii)dswsv477(seqidno.1);图2:pad-dsrna质粒图谱;图3:pad-43-25(表达pgfp)质粒图;图4:在注射wssv后第10天,由bg322饲喂的虾中的wssv相对滴度表达为wssv特异性的非特异性dslucdsrna或dsrna(dsvp19,dsvp28和dswsv477)。将病毒拷贝数调整为log10比例,以图形方式表示所有组。图5:在bg322vp19喂养的wssv感染的虾中小rna的积累。右图:在15%聚丙烯酰胺-8m尿素凝胶上进行rna电泳,该凝胶经gelred核酸染色(thermofisherscientific)染色。(l)takara14-30ssrna梯子;(1-3)bg322-dsvp19喂食被病毒感染的虾,(4)对照虾(无细菌,无wssv),(5)nsrna(dsluc)喂食被wssv感染的虾。左图:虾中vp19缺失的sirna的northern印迹检测:1-3,(3)用病毒感染的虾喂食bg322-dsvp19,(4)对照:虾(无bg322,无wssv),(5)特异性对照:bg322-dsluc喂食了被wssv感染的虾。箭头指示sirna级分中的vp19特异性条带;图6:使用dsrna选择性失活靶基因以启动目标宿主细胞中rna干扰反应的示意图;图7:说明虾的典型生命周期的示意图。发明详述本发明包括可以以不同方式组合的多个方面。提供以下描述以列出元件并描述本发明的一些实施例。这些元件与初始实施例一起列出,但是应当理解,它们可以以任何方式和任何数量组合以创建另外的实施例。各种描述的示例和优选实施例不应被解释为将本发明限制为仅明确描述的系统、技术和应用。此外,该描述应被理解为支持并涵盖具有任意数量的所公开元素的所有各种实施例、系统、技术、方法、设备的描述和权利要求,本申请或任何后续申请中具有任何数量的所公开元素的应用,具有每个公开元素的单独应用,以及具有所有元素的任何和所有各种排列和组合的应用。在一个实施方案中,本发明技术可以包括通过选择性灭活病原性,必需基因或其他基因来控制特定病原体的系统、方法和组合物。这种靶向基因失活可以通过抑制性rna分子(例如双链rna(dsrna)或小发夹rna(shrna))的表达和传递到可能发生病原体复制的目标宿主细胞来实现。如图6所示,在目标宿主中,dsrna可以通过酶dicer的作用加工成长度约为21个核苷酸的小干扰rna(sirna)。这些sirna可能进一步与ago和risc蛋白复合物相互作用,以结合目标病原体特异性mrna序列。最后,risc复合物可以切割病原体特异性mrna,沉默或敲低目标病原体或其他目标基因的表达,并阻断病原体的毒力、复制和/或增殖。另外的实施方案通常可以包括一个或多个靶基因的基因失活。例如,在一个优选的实施方案中,基因失活可以针对一种或多种对于毒力、外壳蛋白、代谢活性、感染途径和/或能量产生等必需的病原体基因。抑制性rna分子向靶动物/细胞/组织的递送可以通过跨王国递送系统来完成。在一个优选的实施方案中,抑制性rna分子的递送可以通过引入基因修饰的宿主特异性微生物例如肠或其他细菌来完成。由于细菌缺乏dicer/risc机制,因此细菌无法将dsrna加工成sirna,因此宿主必须将传递到目标宿主的dsrna加工成可能使目标病毒基因失活的sirna。这样的基因修饰的宿主特异性微生物通常可以称为益生菌或基因修饰的细菌,其可以包括:1)作为靶动物正常内部或外部细菌微生物组的一部分的微生物;2)经修饰能够在目标动物、组织、细胞或宿主环境中定殖的微生物;3)被目标宿主用作食物或能源的微生物;或4)经修饰可在特定动物、组织、细胞或宿主环境中定殖的微生物。在一个优选的实施方案中,一种或多种肠道细菌可以选自:酸微菌;放线菌;alphaprotcobactcria;厌氧蝇菌;bacilli;bactcroidia;β-变形菌;梭菌;δ-变形菌;epsilonproteobacteria;黄杆菌;fusobactcria;γ-变形菌;柔膜菌;opitutac;oscillatoriophycideae;phycisphaerae;planctomycetia;rubrobactcria;sphingobactcriia;集球藻;thermomicrohia;和疣微菌。在优选的实施方案中,本发明的技术可以包括对虾和其他水生生物的抑制性rna分子例如dsrna、shrna、sirna和微小rna的跨王国递送。在一个实施方案中,本发明可以包括一种dsrna介导的疾病控制系统,其可以被配置为灭活诸如虾的水生生物中的一个或多个病毒基因靶标。在一个优选的实施方案中,本发明可以包括一种dsrna介导的疾病控制系统,其可以被配置为灭活诸如水产养殖中生长的虾的水生生物中的一个或多个病毒基因靶标。虾特异性目标病原体的例子包括但不限于:白斑综合症病毒(wssv);传染性皮下及造血组织坏死病毒(ihhnv);黄头病毒(yhv);挑拉综合症病毒(tsv);肌肉坏死病毒(imnv)等。本发明的技术还可以包括用于将抑制性rna分子跨王国递送至其他水生脊椎动物的系统。在某些实施方案中,水生脊椎动物病原体的实例可以包括但不限于引起病毒性出血性败血病、传染性胰腺坏死、鲤鱼病毒血症、传染性造血性坏死病毒、河道鲶鱼病毒、草鱼出血性病毒、野病毒科(例如神经坏死病毒或条纹杰克神经坏死病毒)、传染性鲑鱼贫血病毒;以及寄生虫ceratomyxashasta、ichthyophthiriusmultifillius、cryptobiasalmositica、鲑鱼leopophtheirus、tetrahymena种、trichodina种和epistylus种、鲤鱼春季病毒血症(svc)和流行性造血坏死(ehnv)等。如上所述,在一个优选的实施方案中,一种或多种抑制性rna分子,在这种情况下为dsrna,可以通过转基因的肠细菌递送至虾的目标宿主/种群,所述转基因的肠细菌可以自然地或被配置为定居在虾的肠中。在这个实施方案中,一旦定居在宿主中,修饰细菌的垂直传播就可以传递到宿主的后代,从而自然地将病原性抗性复制到后代。在某些实施方案中,表达一种或多种抑制性rna分子的基因修饰的细菌可以在虾的整个生命周期中定居。例如,如图7所示,表达一种或多种抑制性rna分子的基因修饰的细菌可以在虾上定居,而该虾是:卵、无节幼体、原生动物,mysis,虾类幼虫阶段或成虫。在该实施方案中,定居的细菌可以表达抑制性的rna分子,例如dsrna/shrna,其可以由宿主的dicer/risc复合物进一步加工,从而使病原体特异性的mrna沉默/灭活必需的病原体基因。而且,这些已经永久和/或暂时成为宿主天然微生物组的一部分的定殖的肠道细菌可以将dsrna分子从肠道的最早幼体阶段持续地输送到成年阶段,从而在宿主的整个生命周期内提供必需病原体基因的mrna沉默/灭活。此外,由于肠道细菌载体可能是宿主微生物组中已经自然发生的一部分,因此其存在可能不会对有机体、环境或最终消费者构成任何风险。另外,在某些实施方案中,修饰的细菌还可以通过修饰的细菌在处理过的饲料、或含有基因修饰的细菌的饲料中的分布、或经基因修饰的细菌作为排泄的动物废物传播到环境中而水平地传播到宿主种群。这种特征可以允许将修饰的细菌一次性或至少定期地施用于宿主和/或宿主的环境,从而产生显着的商业优势。本发明的技术可以进一步包括控制靶标有机体中抑制性rna分子表达的水平和时间的方法和技术。在一个优选的实施方案中,一种或多种抑制性rna分子的表达可以在新型基因开关的控制下。这种基因转换可以由转换分子控制,转换分子可以是水溶性的食品级分子,可以添加到宿主有机体的环境中,例如水产养殖池或食品供应中。该开关分子的存在可以激活dsrna的产生。在没有dsrna的情况下,可能不会产生dsrna,或者只能以可忽略的水平发生。在一个优选的实施方案中,水产养殖虾或其他靶有机体可以被一种或多种病毒靶向产生dsrna的肠道细菌感染,而病原体基因失活dsrna的产生的时间和水平可以通过新型基因开关来调节。本发明的技术可以包括用于产生宿主特异性细菌,尤其是可以作为抑制性rna分子的合适递送载体的宿主特异性肠道细菌的方法和技术。作为示例性模型,虾可以用作目标宿主。然而,如本领域普通技术人员可以理解的,这样的方法和技术可以应用于多种不同的有机体。在该优选的实施方案中,可以从野外捕获来自各种幼虫和成年虾物种的一种或多种虾肠样品,并对其特征进行鉴定以鉴定相关的细菌基因组。从这个最初的基因组特征,可以建立一个居住在幼虫肠道中的靶细菌及其物种特异性相对丰度的列表。该表征中的其他步骤可以包括比较和对比来自由上述幼虫变态的成虫的肠道细菌的基因组,以确定哪些细菌种类从幼虫到成年阶段持续存在以及以什么比例存在。在某些实施方案中,宿主特异性细菌可包括以下特征中的一个或多个:1)虾的幼体阶段和成年阶段的肠道菌群中的优势细菌;2)可在虾外部培养,例如在发酵罐中;3)对非目标有机体无已知的不利环境或健康影响;4)能够被基因工程改造以在虾的至少一个但优选地所有生命阶段中稳定地表达和递送足量的dsrna以抑制靶基因复制。细菌rna酶iii可能降解抑制性rna分子,例如dsrna。这样,在一个实施方案中,本发明的技术可包括修饰先前鉴定的宿主特异性细菌或益生菌,以具有降低的rna酶iii表达或失活的rna酶iii功能或活性。rna酶iii表达和/或活性的降低或失活可以抑制或降低rna酶iii介导的dsrna加工成较小的rna种类。在一个优选的实施方案中,可以对先前鉴定的宿主特异性细菌进行基因修饰以在rna酶iii缺陷型的背景中有效表达抑制性rna分子。在该优选实施方案中,可以通过同源重组或其他合适的方法敲除宿主特异性细菌的rna酶iii基因。本发明技术的另一个实施方案可以包括促进宿主特异性细菌基因过表达的系统和方法,所述宿主特异性细菌基因已知增强抑制性rna分子的稳定和/或动员。在这个优选的实施方案中,已知一种或多种稳定dsrna的基因可以过表达以延长其寿命并促进其在宿主有机体/细胞/组织内的移动。在另一个优选的实施方案中,可以剔除调节或抑制已知稳定dsrna的基因的一个或多个基因,从而导致其上调,从而延长dsrna的寿命,从而促进其在宿主有机体内的移动,从而增强病毒基因的抑制作用。另外的实施方案也可以过表达基因或靶基因敲除,其可能导致膜囊泡运输的上调,以促进dsrna动员和递送至宿主有机体。每个前述系统可以体现在遗传构建体中,该遗传构建体可以包括转录调节元件,例如启动子、终止子、共激活子和共阻遏物,以及可以在原核和真核系统中调节的其他控制元件。这样的系统可以允许控制系统内表达的抑制性rna分子或其他蛋白质的类型、时间和数量。另外的实施方案可以包括可以通过另外的外部和/或环境因素,例如特定蛋白质或化合物的存在,例如响应病原体产生的应激相关蛋白质或什至由蛋白质产生的蛋白质和其他前体化合物的存在而诱导的遗传构建体。当前发明技术的另外的优化程序可以包括在实验室环境中使用对多种食物靶标和表达dsrna的微生物的密度不同的竞争研究将修饰的微生物引入虾幼虫中,以确定抑制病毒复制的有效剂量。可以通过原位rna杂交来分析虾的幼虫组织,以确定dsrna对基因表达影响最大的细胞位置,从而更好地选择其他靶基因用于dsrna介导的基因抑制。如上所述,在一个实施方案中,本发明包括新的准转基因系统,其可以进一步包括用于实施基于rnai的策略的新方法,其中天然虾共生细菌用表达dsrna的质粒转化,靶向减少或消除此类病原体的传播的必需基因。这样的实施方案可以特别应用于水产养殖环境中的水生生物。如上所述,水生生物,例如虾,具有天然的抗病毒防御机制rna干扰(rnai)。简而言之,通过利用exo-sirnarnai途径,虾可以识别病毒复制过程中产生的病毒长双链(ds)rna,并使用称为dicer2的rnaseiii家族酶将其消化为21bp的短干扰rna(sirna)片段,并将其用作效应子,以鉴定、切割和灭活复制的病毒基因组。因此,根据本发明的一个方面,提供了一种控制病原体感染的虾的方法,该方法包括向虾种群施用表达异源核酸序列的基因修饰的细菌,所述异源核酸序列特异性地下调虾的至少一种必需病原体基因产物的表达,其中所述至少一种必需病原体抗性基因的表达下调可以防止虾病原体的复制和/或致病性。在一个实施方案中,本发明包括用于病原体载体生物控制的新型超转基因系统的产生。本发明可以具体地包括被配置成将一种或多种抑制性rna分子递送至病原体/疾病传播有机体的副转基因系统。在一个实施方案中,本发明可以包括一种或多种基因工程微生物,其被配置为将一种或多种抑制性rna分子递送至水产养殖系统中的水生生物。在一个优选的实施方案中,本发明可以包括一种或多种经遗传工程改造的肠道细菌,其被配置成将一种或多种dsrna分子递送至水产养殖系统中的水生生物。本发明的其他实施方案包括与靶宿主生物体(在这种情况下为虾)的一种或多种肠道细菌的产生,所述肠道细菌可以与目标宿主有机体内共生或充当本文定义的益生菌。这些肠道细菌可能会在虾发育的所有阶段持续存在于肠道中。本发明的另一个实施方案包括一种或多种肠道细菌的产生,所述肠道细菌可以是内共生的,或如本文所定义的益生菌,其被进一步基因修饰或转化以产生一种或多种dsrna(双链)分子。这些dsrna分子可以对应于一种或多种病原体基因。此外,dsrna分子可能会产生rna介导的选择性病毒基因的下调或抑制。这种rna介导的选择性病毒基因的下调或抑制可能是通过干扰rna过程完成的,如此处一般所述。另一个实施方案包括与靶宿主生物体一起产生一种或多种肠道细菌,其可以与目标宿主有机体内共生,或如本文所述作为益生菌,并且可以定植于目标宿主的肠道,并且还进一步配置为连续递送与白斑综合症病毒(wssv)或本文鉴定的其他水生病原体的一种或多种病原体基因相对应的dsrna分子,并可以进一步引起干扰rna介导的反应,从而导致靶病毒基因的抑制。在一个优选的实施方案中,本发明可以包括用于对虾中的wssv感染进行生物防治的方法和组合物。在该优选实施方案中,可以向虾种群施用表达异源核酸序列的基因修饰的细菌,所述异源核酸序列特异性下调至少一种必需wssv基因产物的表达,其中必需wssv基因表达的下调可以防止虾中wssv的复制和/或致病性。在该优选实施方案中,异源核酸序列表达rna双链体,其包含有义区和反义区,其中反义区包括与靶基因编码的信使rna序列互补的多个连续核苷酸。在一个实施方案中,编码sirna的多核苷酸包含至少一种核苷酸序列,其被配置为产生靶向一种或多种必需wssv基因的hprna。在该优选实施方案中,此类hprna可以抑制wssv中靶基因的表达,包括但不限于:病毒衣壳蛋白19(vp19)\病毒衣壳蛋白19(vp28)和早期非结构基因(wsv477)。在此实施例中,表达rna双链体或hprna的异源核酸序列可以选自:seqidno.1、seqidno.2和seqidno.3。在该实施方案中,seqidno.1被配置为靶向并抑制早期非结构基因(wsv477)的表达(seqid.no9和10)。seqidno.1被配置为靶向并抑制病毒衣壳蛋白19(vp28)的表达(seqid.no7和7),而seqidno.3被配置为靶向并抑制病毒衣壳蛋白19(vp19)的表达(seqid.no5和6)。应当注意,dna序列的鉴定还包括其编码的相应rna序列。因此,如本领域普通技术人员所理解的,对包括dna的seqidno.的引用还具体包括其表达的rna的序列。例如,在要求保护的情况下,异源抑制性多核苷酸可以选自:seqidno.1、seqidno2和/或seqidno.3,这种要求可以包括抑制rna分子的序列,如本领域普通技术人员可以在不进行过多实验的情况下容易地确定的那样。在一个优选的实施方案中,与基因病原体的其他菌株的序列比较时,由靶病原体基因编码的信使rna序列可以包括位于高于平均同源性的区域中的基因,或者换句话说,完全或部分位于病原体基因组的最保守区域中的基因。在一个特定的实施方案中,seqidno1、seqidno2和seqidno.3一般对应于seqidno.5-10中高度保守的结构和/或非结构蛋白编码区。自然地,这样的序列是示例性的,因为它们可以可替代地在一个或多个不同的基因编码区段上是冗余的或重叠的。本发明可以进一步包括一种或多种用于调节多种病原体基因的载体,其中包含一种或多种dsrna的载体可以对应于一种或多种选择的病原体基因,例如在seqidno.6、8、10中鉴定的wssv基因。如在图2-3中大体上所示,该实施方案可以包括质粒表达系统的使用。在一些实施方案中,该质粒可以具有一个或多个表达盒,包括:至少一个基因抑制盒,其包含与启动子序列可操作地连接的多核苷酸,其中所述多核苷酸编码干扰rna分子,例如dsrna,或随后将产生干扰rna分子的分子,例如dsrna,其通过rna干扰降低靶病原体基因的表达。本发明的优选实施方式包括用于调节多个宿主基因的载体,其中包含一个或多个dsrna的载体可以对应于一个或多个选择宿主基因。该实施方案可以包括质粒表达系统的使用。在一些实施方案中,该质粒可以具有一个或多个表达盒,包括:至少一个基因抑制盒,其包含与启动子序列可操作地连接的多核苷酸,其中所述多核苷酸编码干扰rna分子,例如dsrna,或随后将产生干扰rna分子的分子,例如dsrna,其通过rna干扰减少目标宿主基因的表达。本发明的另一个实施方案包括用于调节宿主和病原体基因的载体,其中所述载体包含一个或多个dsrna,其可以对应于一个或多个选择的宿主和病原体基因。该实施方案可以包括质粒表达系统的使用。在一些实施方案中,该质粒可具有一个或多个表达盒,包括:至少一个基因抑制盒,其包含可操作地连接至启动子序列的第一多核苷酸,其中所述多核苷酸编码干扰rna分子,例如dsrna,或随后会产生干扰rna分子(例如dsrna)的分子,该分子会通过rna干扰降低目标宿主基因的表达。该基因盒可以进一步包含可操作地连接至启动子序列的第二多核苷酸,其中该多核苷酸编码干扰rna分子,例如dsrna,或随后将产生干扰rna分子的分子,例如dsrna,其通过rna干扰降低靶病原体基因的表达。本发明还包括用于抑制宿主中病毒或细菌基因表达的载体,其中所述载体包含至少一个基因抑制盒,所述基因抑制盒包含与启动子序列可操作地连接的多核苷酸,其中多核苷酸编码通过rna干扰减少宿主中靶病原体基因表达的sirna分子。在一个实施方案中,编码sirna的多核苷酸包含seqidno.2、seqidno.2和/或seqidno.3的核苷酸序列。同样,本发明的载体可以包括多个基因抑制盒,其中每个基因抑制盒包含编码靶向相同mrna序列或不同mrna序列的sirna分子(例如dsrna)的多核苷酸。例如,每个基因抑制盒可编码靶向两个或更多个不同基因的mrna序列的dsrna分子。此外,本发明的每种载体可以包括多个基因促进盒和多个基因抑制盒。用于基因抑制盒的合适启动子的实例包括但不限于t7启动子、bla启动子、u6启动子、polii启动子、ell启动子和cmv启动子等。任选地,基因促进盒和基因抑制盒的每个启动子序列可以是诱导性的和/或组织特异性的。本发明的另一个方面可以包括提供基因工程肠细菌的新颖方法,其可以被配置为定居在动物肠道中并防止病毒病原体逃逸到肠道周围的上皮中。更具体地说,本发明的一个目的可以是将基因工程肠细菌引入虾肠,并进一步配置以产生和分泌dsrna。这些dsrna分子可能会被周围的上皮细胞吸收,从而产生强大的rnai级联反应,从而阻止病毒复制和/或抑制病原体水平,从而使大量病毒粒子无法从虾肠周围的上皮细胞迁移。在另外的实施方案中,靶标上皮细胞可以通过内吞、囊泡运输、吞噬作用和/或其他主动或被动多核苷酸转运过程摄取由位于肠中的转化的超转基因细菌分泌的dsrna。在其一些实施方案中,本发明涉及分离的核酸剂,更具体地但非排他地涉及其在控制病原体感染的动物例如虾中的用途。根据本发明的一些实施方案,必需基因可以包括选自由一种或多种对毒力、外壳蛋白、代谢活性、感染途径和/或能量产生等必需靶病原体基因组成的组的基因。靶基因可包括负责致病性或引起宿主疾病状况的能力的一种或多种基因。此类靶基因的实例还可包括一种或多种毒力因子。此类毒力因子的示例可能包括但不限于:·粘附因子:该组可能包括有助于细菌病原体粘附于某些细胞的基因;·入侵因素:该组可能包括允许细菌入侵宿主细胞的表面成分基因;·胶囊:这一类可能包括结构性胶囊的基因,可以保护细菌免受调理作用和吞噬作用。·内毒素:这一类可能包括几种类型的有毒脂多糖的基因,这些基因可以引发免疫反应。·外毒素:这一类可能包括由致病细菌产生和/或分泌的几种蛋白质毒素和酶的基因。主要类别包括细胞毒素、神经毒素和肠毒素。·铁载体:这一类可能包括几种类型的铁结合因子的基因,这些因子可使某些细菌与宿主竞争铁,后者与血红蛋白、转铁蛋白和乳铁蛋白结合。·宿主转化因子:该组可能包括改变宿主代谢以致病菌受益的基因,包括但不限于逃避宿主防御。·结构性:这组基因可能包括编码病毒复制所需的病毒capsis的基因。·非结构病毒:这组基因可能包括编码病毒复制所需的非结构病毒蛋白质的基因。实例可以包括负责病毒基因组整合和/或复制的基因。本发明的一个优选实施方案可以包括一种分离的核酸试剂,其包含表达特异性下调至少一种病原体基因表达的核酸序列的多核苷酸。在一个优选的实施方案中,该分离的核酸剂可包含表达dsrna序列的多核苷酸,该dsrna序列通过sirna过程特异性下调至少一种动物病原体的表达。在本发明的一些实施方案中,本发明的另一个实施方案可以包括核酸构建体,其包含编码分离的核酸试剂例如dsrna的核酸序列或可以形成dsrna的核酸试剂。根据本发明的替代实施方案,核酸序列直接对应于病原体基因,而在替代实施方案中,核酸序列对应于一个或多个病原体基因或与之重叠。根据本发明的一些实施方案,dsrna核酸序列直接对应于病原体基因,而在替代实施方案中,dsrna核酸序列对应于一个或多个病原体基因或与之重叠。如上所述,本发明的技术可以应用于植物和动物的多种有机体。例如,本发明可以包括dsrna介导的疾病控制系统,其可以被配置成使任何合适的宿主有机体中的一个或多个病原体基因靶失活。在一个特定的实施方案中,一种或多种抑制性rna分子,在这种情况下为dsrna,可以被递送至脊椎动物和无脊椎动物的目标宿主/种群。例如,家禽,例如:鸡、火鸡、鸭和鹅(参见表4)、蜜蜂(参见图5)以及哺乳动物(参见图6),包括人类,都是通过基因修饰的肠道细菌而产生的,这些细菌可能是天然的,也被配置为在宿主的肠道中定殖。在该示例性实施例中,定居的细菌可以表达抑制性的rna分子,例如dsrna/shrna,可以由宿主的dicer/risc复合物进一步加工,从而使病原体特异性的mrna沉默/灭活必需的病原体基因。下表4-6列出了本发明可能靶向的一些家禽特异性病原体的实例。这些已定殖的肠道细菌已经成为宿主自然微生物组的一部分,可以在发育的各个阶段不断地通过肠道传递dsrna分子,从而在宿主的整个生命周期中提供必需病原体特异性基因的mrna沉默/灭活。此外,由于肠道细菌载体可能是家禽或蜜蜂微生物组中已经天然存在的一部分,因此它的存在不会对有机体、环境或最终消费者构成任何风险,也不会允许通过将作为废物排泄到环境中的改良细菌分布垂直传播给子代和水平传播到整个宿主。在某些实施方案中,益生菌可以被配置为表达一种或多种干扰rna分子。在该优选实施方案中,受试者,例如人,可以服用治疗或有效量的益生菌,其可以充当疫苗或抗生素或抗病毒药物。此类益生菌的产生和递送是众所周知的,并且将在本领域普通技术人员的技术范围内。如本文所用,术语“水产养殖”包括在受控条件下的水生生物的养殖。如本文所用,术语“水生生物”和/或“水生动物”包括在淡水或盐水中生长的有机体。水生生物/动物包括脊椎动物、无脊椎动物、节肢动物、鱼类、软体动物,包括虾(例如对虾、penaeusesculentu、penaeussetiferus、南美蓝对虾、西方白对虾、日本对虾、南美白对虾、斑节对虾、中国对虾、阿兹特库斯对虾、桃红对虾、印度对虾、和墨吉对虾、加利福尼亚对虾、熊虾、草虾、丰年虾、淡水虾等)、螃蟹、牡蛎、扇贝、虾蛤、软骨鱼(例如鲷鱼、鳟鱼、鲈鱼、条纹鲈、罗非鱼、鲶鱼、鲑鱼、鲤鱼、鲶鱼、黄尾鱼、鲤鱼斑马鱼、红鼓等)、甲壳类动物等。虾还包括在水产养殖中养殖的虾。术语“益生菌”是指可以在宿主中定植足够长的时间以研究治疗或有效量的干扰rna分子的微生物,例如细菌。益生菌可以包括内共生细菌,或可以永久性地暂时定居于动物(例如水生生物)的天然菌群。益生菌有机体还可以包括藻类和真菌,例如酵母。细菌载体的具体例子包括细菌(例如球菌和棒状杆菌)、丝状藻类和碎屑。在宿主的所有生命周期中可以是内源的可转化细菌载体细胞的具体实施方案可以包括本文列出的所有那些。另外的实施方案可以包括一种或多种选自本文列出的实例的细菌菌株。自然地,这样的列表不是排他的,并且仅是副转基因细菌菌株的某些优选实施方案的示例。如本文所用,短语“宿主”是指携带致病性病原体的动物、易受致病性病原体感染的动物、成员携带致病性病原体的动物种群或成员容易感染致病性病原体的动物种群。这些宿主包括表4-6和其他地方列出的宿主。如本文所用,“病原体”是指致病因子。这些包括表4-6和其他地方中包含的病原体。如本文所用,“疫苗”是指导致主动和被动免疫的组合物。多核苷酸及其表达的基因产物都在本文中称为疫苗。包括经配置以表达干扰细菌的经处理细菌的饲料也可以是疫苗。将经处理的饲料喂养给动物可以是疫苗接种。如本文所用,短语“饲料”是指作为饲料方案的一部分引入的动物可消耗材料,或者在水生动物的情况下直接应用于水中。“处理的饲料”是指经配置以表达干扰细菌的经处理的细菌处理的饲料。如本文所用,术语“控制”和/或“生物控制”是指减少和/或调节病原体/疾病的进程和/或传播。如本文所用,术语“核酸”是指核糖核苷酸或脱氧核糖核苷酸的聚合物。通常,“核酸或”核酸剂”聚合物以单链或双链形式存在,但也已知形成包含三条或更多链的结构。术语“核酸”包括天然存在的核酸聚合物以及包含已知核苷酸类似物或修饰的骨架残基或键的核酸,所述核酸类似物或修饰的骨架残基或键是合成的、天然存在的和非天然存在的,其具有与参考核酸相似的结合特性,并且以与参考核苷酸相似的方式代谢。示例性的类似物包括但不限于硫代磷酸酯、氨基磷酸酯、甲基膦酸酯、手性甲基膦酸酯、2-o-甲基核糖核苷酸和肽核酸(pna)。“dna”、“rna”、“多核苷酸”、“多核苷酸序列”、“寡核苷酸”、“核苷酸”、“核酸”、“核酸分子”、“核酸序列”、“核酸片段”和“分离的核酸片段”在本文可互换使用。如本文所用,术语“分离的”、“纯化的”或“生物纯的”是指基本上或基本上不含通常以其天然状态或在生产该材料时通常伴随该材料的组分的材料。在一个示例性的实施方案中,使用分析化学技术例如聚丙烯酰胺凝胶电泳或高效液相色谱法测定纯度和均质性。基本上纯化了制剂中存在的主要种类的核酸或特定细菌。在一个示例性的实施方案中,术语“纯化的”表示核酸或蛋白质在电泳凝胶中产生基本上一个条带。通常,分离的核酸或蛋白质具有表示为范围的纯度水平。组分的纯度范围的下限为约60%、约70%或约80%,纯度范围的上限为约70%、约80%、约90%或大于约90%。当涉及例如细胞或核酸、蛋白质或载体时,术语“重组”表示该细胞、有机体、核酸、蛋白质或载体已通过引入异源核酸或蛋白质或天然核酸或蛋白质的改变而被修饰,或者该细胞源自如此修饰的细胞。因此,例如,重组细胞可以表达在细胞的天然形式内未发现的基因(非重组或野生型),或表达否则以异常方式表达、过表达、表达不足或根本不表达的天然基因。术语“基因修饰”、“生物转化”、“转基因”、“转化”和“转染”在含义上与“重组”相似。“转化”、“转基因”和“转染”是指多核苷酸向宿主有机体的基因组或细胞中的转移。多核苷酸的这种转移可导致多核苷酸的遗传稳定遗传或使多核苷酸保留在染色体外(未整合到细胞的染色体中)。遗传稳定的遗传可能潜在地要求转基因有机体或细胞在一段时间内经历一种或多种条件,这些条件需要转录部分或全部转移的多核苷酸才能使转基因有机体或细胞生存和/或生长。转化到细胞中但未整合到宿主染色体中的多核苷酸仍作为表达载体保留在细胞内。为了使表达载体保留在细胞或细胞的后代中,可能需要在一定的生长或环境条件下使细胞生长。此外,为了发生表达,可能需要将有机体或细胞保持在一定条件下。包含重组多核苷酸的宿主有机体或细胞可被称为“转基因”或“转化的”有机体或细胞,或简称为“转化体”,以及重组有机体或细胞。遗传改变的有机体是指其遗传物质发生任何变化的任何有机体,无论是在细胞核还是细胞质(细胞器)中。这样,遗传改变的有机体可以是重组或转化的有机体。与野生型有机体相比,转基因有机体还可以是经历了一种或多种诱变剂的有机体或经历一种或多种诱变剂的有机体的后代,并且其dna的变化是由一种或多种诱变剂引起的(即不经历诱变剂的有机体)。同样,已经繁殖出将突变整合到其遗传物质中的有机体是遗传改变的有机体。术语“载体”是指可以将dna、rna、蛋白质或多肽引入宿主的某些方式。待引入宿主的多核苷酸、蛋白质和多肽本质上可以是治疗性或预防性的;可以编码或为抗原;本质上可以是监管的。载体种类繁多,包括病毒、质粒、噬菌体、粘粒和细菌。“表达载体”是能够在选择的宿主细胞或有机体中复制的核酸。表达载体可以复制为自主结构,或者可以全部或部分整合到宿主细胞染色体或细胞器的核酸中,或者用作将外来dna传递到细胞的穿梭物,从而与宿主细胞基因组一起复制。因此,表达载体是能够在选定的宿主细胞、细胞器或有机体中复制的多核苷酸,例如质粒、病毒、人工染色体、核酸片段,并将其表达载体上的某些基因(包括目的基因)转录并翻译成细胞、细胞器或有机体中的多肽或蛋白质;或本领域已知的任何合适的构建体,其包含“表达盒”。相反,如本文实施例中所述,“盒”是含有本发明表达载体的一部分的多核苷酸。盒的使用有助于表达载体的组装。表达载体是复制子,例如质粒、噬菌体、病毒、嵌合病毒或粘粒,并且其包含与表达控制序列可操作连接的所需多核苷酸序列。当表达控制序列控制和调节该多核苷酸序列的转录和/或翻译时,该多核苷酸序列“可操作地连接至一个或多个表达控制序列”(例如,启动子和任选的增强子)。如本文所用,短语“基因产物”是指rna分子或蛋白质。本发明利用分子生物学领域的常规技术。公开本发明中一般使用方法的基本文献包括greenandsambrook,4thed.2012,coldspringharborlaboratory;kriegler,genetransferandexpression:alaboratorymanual(1993);和ausubeletal.,eds.,currentprotocolsinmolecularbiology,1994-current,johnwiley&sons。除非另有说明,否则根据常规用法使用技术术语。分子生物学中常用术语的定义可以在例如benjaminlewin,genesix,牛津大学出版社出版,2007(isbn0763740632);krebs,etal.(eds.),theencyclopediaofmolecularbiology,blackwellscienceltd.出版,1994(isbn0-632-02182-9);和roberta.meyers(ed.),molecularbiologyandbiotechnology:acomprehensivedeskreference,vchpublishers,inc.出版,1995(isbn1-56081-569-8)中找到。本教导考虑根据所选宿主物种靶向同系物和直系同源物。同源序列包括直向同源序列和旁系同源序列。术语“旁系同源的”涉及导致旁系同源基因的物种的基因组内的基因复制。术语“直系同源”由于祖先关系而涉及不同有机体中的同源基因。因此,直系同源物是在给定两个物种(kooninev和galperinmy(sequence-evolution-function:computationalapproachesincomparativegenomics.boston:kluweracademic;2003.chapter2,evolutionaryconceptingeneticsandgenomics.availablefrom:ncbi(dot)nlm(dot)nih(dot)gov/books/nbk20255)的最后一个共同祖先中源自单个祖先基因的进化对应物,因此具有相同功能的可能性很高。因此,直系同源物通常起着与另一物种的原始物种相似的作用。同源性(例如,同源性百分比,序列同一性+序列相似性)可以使用任何计算成对序列比对的同源性比较软件来确定。如本文所用,在两个核酸或多肽序列的上下文中的“序列同一性”或“同一性”包括提及两个序列中比对时相同的残基。当使用序列同一性百分比来表示蛋白质时,会认识到不相同的残基位置通常会因保守的氨基酸取代而有所不同,其中氨基酸残基被具有相似化学性质(例如电荷或疏水性)的其他氨基酸残基取代,因此不会改变分子的功能特性。如果序列在保守取代中不同,则可以向上调整序列同一性百分比以校正取代的保守性质。通过这种保守取代而不同的序列应具有“序列相似性”或“相似性”。进行该调整的手段是本领域技术人员众所周知的。通常,这涉及将保守取代计为部分错配而不是全部错配,从而增加序列同一性百分比。因此,例如,在相同氨基酸的评分为1,非保守取代的评分为零的情况下,保守取代的评分为零至1。计算保守取代的得分,例如,根据henikoffs和henikoffjg.的算法[aminoacidsubstitutionmatricesfromproteinblocks.proc.natl.acad.sci.u.s.a.1992,89(22):10915-9]。根据一个具体的实施方案,同源序列与本文提供的序列至少是本文提供的序列的60%、65%、70%、75%、80%、85%、90%、95%或甚至与本文提供的序列(核酸或氨基酸序列)相同。在50%-99%之间的seqidnos1-6的同源序列可以包括在本发明的某些实施方案中。宿主的病原体抗性基因产物的表达下调可以例如通过直接检测基因转录物(例如通过pcr)、通过检测由基因编码的多肽(例如通过蛋白质印迹或免疫沉淀)、通过检测由基因编码的多肽的生物活性(例如,催化活性、配体结合等)、或通过监测宿主的变化(例如,降低宿主的死亡率等)来监测。另外或可选择地,与未用本发明的试剂处理的野生型(即对照)宿主相比,可通过测量宿主中的病原体水平(例如病毒水平、细菌水平等)来监测病原体抗性基因产物的表达下调。如上所述,根据本发明的一些方面,提供了一种分离的核酸试剂,其包含特异性下调至少一种宿主病原体抗性基因产物表达的核酸序列。根据一个实施方案,所述试剂是多核苷酸试剂,例如rna沉默剂。在一个优选的实施方案中,所述试剂是被配置为诱导rna干扰的多核苷酸试剂,例如dsrna。如本文所用,术语“干扰rna分子”或“干扰rna”是指能够抑制或“沉默”靶基因表达的rna。在某些实施方案中,rna沉默剂能够通过转录后沉默机制防止mrna分子的完全加工(例如,完全翻译和/或表达)。rna沉默剂包括非编码rna分子,例如包含成对链的rna双链体,以及可从其产生此类小的非编码rna的前体rna。示例性的rna沉默剂包括dsrna,例如sirna、mirna和shrna。在一实施方案中,rna沉默剂能够诱导rna干扰。在另一个实施方案中,rna沉默剂能够介导翻译阻遏。在本发明的一些实施方案中,核酸试剂是双链rna(dsrna)。如本文所用,术语“dsrna”涉及通过碱基配对结合在一起的两条反平行多核糖核酸链。实例包括seqidno:1-5。两条链的长度可以相同或不同,只要两条链之间具有足够的序列同源性,即可形成双链结构,且双链结构在整个长度上至少互补60%、70%、80%、90%、95%或100%。根据本发明的一个实施方案,dsrna分子没有突出端。根据本发明的另一个实施方案,dsrna分子包含突出端。根据其他实施方案,链被排列成使得在链的末端至少有1、2或3个碱基不对齐(即,相对链中不存在互补碱基),使得当链为双链时,双链体的一个或两个末端出现1、2或3个残基的突出端退火。应当注意,可以根据编码靶基因转录本的dna的核酸序列来定义dsrna,并且应当理解,与基因的编码序列相对应的dsrna序列包含该基因编码的rna互补序列,或转录为rna的基因的其他序列。抑制性rna序列与靶基因转录物的部分可以大于90%相同,或甚至100%相同。备选地,rna的双链体区可以在功能上定义为能够在严格条件下(例如400mmnacl,40mmpipesph6.4、1mmedta,60℃杂交12磅/小时;然后洗涤)与一部分靶基因转录物杂交的核苷酸序列。与靶基因转录物互补的双链核苷酸序列的长度可以是至少约18、19、21、25、50、100、200、300、400、491、500、550、600、650、700、750、800、900、1000或更多个碱基。在本发明的一些实施方案中,双链核苷酸序列的长度为约18至约530个或更长的核苷酸。本教导涉及各种长度的dsrna,其中较短的版本,即x较短或等于50bp(例如17-50),被称为sirna或mirna。更长的51-600dsrna分子在本文中称为dsrna,可以对其进行进一步加工以生成sirna分子。根据一些实施方案,dsrna的核酸序列的长度大于15个碱基对。根据其他实施方案,dsrna的核酸序列的长度为19-25个碱基对、长度为30-100个碱基对、长度为100-250个碱基对或长度为100-500个碱基对。根据其他实施方案,dsrna的长度为500-800个碱基对、长度为700-800个碱基对、长度为300-600个碱基对、长度为350-500个碱基对或长度为400-450个碱基对。在一些实施方案中,dsrna的长度为400个碱基对。在一些实施方案中,dsrna的长度为750个碱基对。术语“sirna”是指诱导rna干扰(rnai)途径的小的抑制性rna双链体(通常在17-30个碱基对之间,但也更长,例如31-50bp)。通常,化学合成sirna为21聚体,在末端具有19bp的双链体中心区域和对称的2个碱基的3'突出端,尽管最近已描述了化学合成的25-30个碱基长的rna双链体与相同位置的21mers相比,其效能可提高100倍之多。理论上,使用更长的rna触发rnai所获得的增强的效力据认为是由dicer提供底物(27mer)而不是产物(21mer)产生的,这提高了sirna双链体进入risc的速度或效率。已发现3'突出端的位置会影响sirna的效力,反义链上具有3'突出端的不对称双链体通常比有义链上具有3'突出端的不对称双链体更有效(rose等人,2005年)。这可以归因于不对称链加载到risc中,因为靶向反义转录物时观察到相反的功效模式。在某些实施方案中,dsrna可以来自2个来源,一种来自基因转录物,该基因转录物是由dna的相反链上的相反基因启动子产生的;2)来自单个基因启动子产生的但具有内部互补性的折返发夹结构。例如,可以连接双链干扰rna(例如,sirna)的链以形成发夹或茎环结构(例如,shrna)。因此,如上所述,rna沉默剂也可以是短发夹rna(shrna)。如本文所用,术语“shrna”是指具有茎-环结构的rna试剂,其包含互补序列的第一和第二区域,区域的互补程度和方向足以使区域之间发生碱基配对,第一和第二区域由环区域连接,该环是由于环区域内核苷酸(或核苷酸类似物)之间缺乏碱基配对而导致的。环中核苷酸的数目是并包括3至23、或5至15、或7至13、或4至9、或9至11之间的数字。环中的某些核苷酸可参与与环中其他核苷酸的碱基对相互作用。可用于形成环的寡核苷酸序列的实例包括5'-uucaagaga-3'(brummelkamp,t.r.etal.(2002)science296:550,seqidno.302)和5'-uuuguguag-3'(castanotto,d.etal.(2002)rna8:1454,seqidno.303)。本领域技术人员将认识到,所得的单链寡核苷酸形成茎环或发夹结构,其包含能够与rnai机制相互作用的双链区。如本文所用,短语“microrna”(在本文中也可互换地称为“mirna”或“mir”)或其前体是指充当转录后调节子的microrna(mirna)分子。通常,mirna分子是长度约为20至22个核苷酸的rna分子,其可以被加载到risc复合物中并指导另一个rna分子的切割,其中另一个rna分子包含基本上与mirna分子的核苷酸序列互补的核苷酸序列。通常,mirna分子是通过蛋白质(例如dcl蛋白)从“pre-mirna”或本文中使用的pre-mirna分子加工而成的,并加载到risc复合物中,在其中它可以指导靶rna的切割分子。pre-microrna分子通常是从pri-microrna分子(原始转录本)加工而成的。pre-microrna两侧的单链rna片段对于将pri-mirna加工成pre-mirna非常重要。切割位点似乎是由距茎-ssrna连接点的距离决定的(hanetal.2006,cell125,887-901,887-901)。如本文所用,“pre-mirna”分子是约100至约200个核苷酸,优选约100至约130个核苷酸的rna分子,其可以采用包含不完善的双链rna茎和单链rna环的二级结构(也称为“发夹”),并在双链rna茎中进一步包含mirna的核苷酸序列(及其互补序列)。根据一个具体的实施方案,mirna及其补体位于距mirna双链rna茎的自由端约10至约20个核苷酸处。单链环区的长度和序列不是关键性的,并且可以有很大的变化,例如,长度介于30至50个核苷酸之间。mirna及其补体之间的互补性不一定是完美的,并且可以容忍大约1-3个未配对核苷酸的凸起。rna分子采用的二级结构可以通过本领域常规的计算机算法例如mfold来预测。双链rna的特定链源于pre-mirna,它通过dcl活性释放并加载到risc复合体上,取决于5'端的互补程度,从而将在5'末端参与切割的dsrna茎不同链的核苷酸之间的氢键作用最少的链加载到risc复合物中,并确定靶rna分子降解的序列特异性。但是,如果凭经验从特定的合成pre-mirna分子中获得的mirna分子不起作用(因为risc复合体上载有“错误的”链),则可以很明显地看出,可以通过交换mirna分子及其在pre-mirna分子dsrna茎的相应链上的补体的位置来解决此问题。如本领域中已知的,涉及两个氢键的a和u之间或涉及两个氢键的g和u之间的结合不如涉及三个氢键的g和c之间的结合强。天然存在的mirna分子可以包含在其天然存在的pre-mirna分子中,通过将通常从这种现有的pre-mirna分子加工而来的mirna分子的核苷酸序列交换为另一个感兴趣的mirna的核苷酸序列,它们也可以被引入到现有的pre-mirna分子支架中。pre-mirna的支架也可以是完全合成的。同样,合成的mirna分子可以包含在现有的pre-mirna分子支架或合成的pre-mirna支架中,并从中进行加工。某些pre-mirna支架可能比其他支架更可取,因为它们可以正确地加工成设计的microrna的效率,特别是当表达为嵌合基因时,除pre-microrna外,其他dna区(例如未翻译的前导序列或转录终止区和聚腺苷酸化区)也被整合到了初级转录物中。根据本教导,dsrna分子可以是天然存在的或合成的。dsrna可以是长和短dsrna分子的混合物,例如dsrna、sirna、sirna+dsrna、sirna+mirna或它们的组合。在一个优选的实施方案中,设计一种或多种核酸试剂以特异性地靶向目标靶基因(例如病原体非结构基因)。应当理解,核酸试剂可用于下调一个或多个靶基因(例如,如上文详细描述的)。如果靶向多个靶基因,则使用包含用于靶向多个靶基因的多种核酸试剂的异源组合物。或者,将多种核酸试剂分开配制。根据一个具体的实施方案,使用了针对单个靶的许多不同的核酸剂分子,其可以分别或同时(即,共同配制)使用。例如,为了使目的mrna的表达沉默,可以如下选择适合用于本发明某些实施方案的dsrna的合成。首先,扫描包括3'utr和5'utr的mrna序列。其次,使用任何序列比对软件,例如可从ncbi获得的blast软件,将mrna序列与合适的基因组数据库进行比较。滤出与其他编码序列具有显着同源性的mrna序列中的推定区域。选择合格的靶序列作为dsrna合成的模板。优选的序列是与基因组中的其他基因几乎没有同源性以减少“脱靶”效应的序列。应当理解,本发明的一些实施方案的rna沉默剂不必限于仅包含rna的那些分子,而是还涵盖化学修饰的核苷酸和非核苷酸。根据一个实施方案,dsrna特异性地靶向选自seqidno1-4或其同源物变体的基因。另外,术语amplicon是指一段dna或rna。在某些实施方案中,dsrna分子的表达不需要转录dsrna的顺式作用调节序列(例如,异源)。如本文所用,术语“异源的”是指外源的、非天然存在于宿主的天然细胞中或dsrna被馈送到宿主的细胞中(例如通过整合位置或在单元格中非自然找到)。所述核酸剂可以进一步包含在包含其他调控元件的核酸构建体中。例如,从表达盒的转录,调节区(例如启动子、增强子、沉默子、前导子、内含子和聚腺苷酸化)可用于调节rna链(或多个链)的转录。因此,在一个实施方案中,提供了一种包含核酸试剂的核酸构建体。所述核酸构建体可具有经构建以促进可操作地连接至在宿主细胞中起作用的一个或多个启动子序列的本发明的rna分子的转录的多核苷酸序列。可以将多核苷酸序列置于通常存在于宿主基因组中的内源启动子的控制之下。在可操作地连接的启动子序列的控制下,本发明的多核苷酸序列可以进一步侧接有利地影响其转录和/或所得转录本的稳定性的其他序列。这样的序列通常位于表达构建体的启动子的上游和/或3'端的下游。如涉及调节序列和结构核苷酸序列的术语“可操作地连接”是指该调节序列引起所连接的结构核苷酸序列的调节表达。遗传“控制元件”是指位于结构核苷酸序列上游、内部或下游的核苷酸序列,其影响转录的时间和水平或量、rna加工或稳定性或相关结构核苷酸序列的翻译。调节序列可以包括启动子、翻译前导序列、内含子、增强子、茎环结构、阻遏物结合序列、终止序列、暂停序列、聚腺苷酸化识别序列等。应当理解,可以以多种方式将核酸试剂递送至动物,例如虾。根据一个实施方案,一些实施方案的组合物包含细胞,所述细胞包含核酸试剂。如本文所用,相对于宿主,术语“细胞”可以指处于其生命周期的任何阶段的动物细胞。在某些实施方案中,副转基因系统可以建立在宿主的整个生命周期中都是内源的基因修饰的细菌。根据一个具体的实施方案,该细胞是细菌细胞。在另一个实施方案中,可以将包含被配置为表达dsrna的基因修饰的细菌的组合物配制成饲料和/或可以被进一步配置为分散到环境中的水分散性颗粒或粉末。在又一个实施方案中,本发明的组合物还可包含可湿性粉剂、喷雾剂、乳剂、胶体、水溶液或有机溶液、粉剂、丸剂或胶体浓缩物。可以将组合物的干燥形式配制成在润湿时立即溶解,或者以控制释放,持续释放或其他时间依赖性方式溶解。替代地或另外地,该组合物可以包含水溶液。这样的水溶液或悬浮液可以以浓缩的储备溶液的形式提供,该浓缩的储备溶液在施用前被稀释,或者作为一种可供立即使用的稀释溶液。这样的组合物可以以多种方式配制。通过与各种惰性材料(例如无机矿物(有机硅或硅衍生物、层状硅酸盐、碳酸盐、硫酸盐、磷酸盐等)或植物性材料(粉状玉米芯、稻壳、核桃壳等))混合,它们可以用作可湿性粉剂、颗粒剂或粉剂。含有副转基因细菌的制剂或组合物可以包括铺展剂-佐剂、稳定剂、其他杀虫添加剂或表面活性剂。液体制剂可以用作泡沫、悬浮液、乳油等。成分可以包括神学剂、表面活性剂、乳化剂、分散剂或聚合物。根据一个实施方案,通过喂养将组合物施用于宿主。用组合物喂养宿主可以在数小时、数天、数周、数月甚至数年的时间内一次、定期或半定期地进行。如上所述,本发明的dsrna可以作为裸dsrna施用。备选地,本发明的dsrna可以与本领域技术人员已知的载体偶联,例如转染剂,例如pei或壳聚糖或蛋白质/脂质载体或偶联至纳米颗粒。组合物可以在给药前以合适的方式配制,例如冻干、冷冻干燥、微囊化、干燥、或在水性载体、介质或合适的稀释剂如盐水或其他缓冲液中配制。合适的农业载体可以是固体、半固体或液体,并且是本领域众所周知的。这样的组合物可以被认为是“农业上可接受的载体”,其可以覆盖通常在农药配制技术中使用的所有佐剂,例如惰性组分、分散剂、表面活性剂、增粘剂、粘合剂等。根据一些实施方案,以有效减少或抑制至少一种宿主病原体抗性基因产物的表达的量提供核酸剂。如本文所用,“抑制量”或“有效量”或“治疗有效量”是指足以下调靶基因(减少表达)至少20%、30%、40%、50%或更多(例如60%、70%、80%、90%或什至100%)的dsrna的量,或将动物或动物种群(例如水产养殖中的虾)的死亡率降低至少可测量的百分比,最好在1%-100%。可以使用本领域已知的任何方法来测试基因沉默的功效。例如,使用定量rt-pcr测量基因敲低。因此,例如,可以收集每个治疗组的宿主动物并将其汇集在一起。可以从中提取rna,并可以进行cdna合成。然后可以使用qrt-pcr通过测量基因表达水平来使用cdna评估rnai的程度。可以将本发明的试剂包装在包括核酸试剂(例如dsrna),将核酸试剂,构建体或组合物施用于特定宿主的说明书的试剂盒中。如本文所用,术语“基因”或“多核苷酸”是指任意长度的单核苷酸或核酸残基的聚合物。多核苷酸可以包含脱氧核糖核苷酸,核糖核苷酸和/或其类似物,并且可以是双链或单链的。多核苷酸可以包含修饰的核酸(例如,甲基化的)、核酸类似物或非天然存在的核酸,并且可以被非核酸残基中断。例如,多核苷酸包括基因、基因片段、cdna、分离的dna、mrna、trna、rrna、任何序列的分离的rna、重组多核苷酸、引物、探针、质粒和载体。定义中包括已被修饰的核酸聚合物,无论是天然修饰还是通过干预修饰。可以通过本领域已知的用于合成dna和rna分子的任何方法来制备本发明的构建体,包括化学合成本领域众所周知的寡脱氧核糖核苷酸和寡核糖核苷酸的技术,例如固相亚磷酰胺化学合成。或者,可通过在体外和体内转录编码反义rna分子的dna序列来产生rna分子。可以将此类dna序列掺入多种载体中,这些载体掺入合适的rna聚合酶启动子,例如t7或sp6聚合酶启动子。或者,取决于所使用的启动子,组成性或诱导性合成反义rna的反义cdna构建体可以稳定地引入细胞系。而且,可以引入对核酸分子的各种众所周知的修饰作为增加细胞内稳定性和半衰期的手段。可能的修饰包括但不限于在分子的5'和/或3'末端添加核糖核苷酸或脱氧核糖核苷酸的侧翼序列,或在寡脱氧核糖核苷酸主链内使用硫代磷酸酯或2'o-甲基而不是磷酸二酯酶键。如本文所用,术语“大约”或“大概”是指±10%。每当在本文中指示数值范围时,其意图包括在指示范围内的任何引用数字(分数或整数)。短语“在第一指示数字和第二指示数字之间的范围/范围”和从第一指示数字“到第二指示数字”的“范围/范围”在本文中可互换使用,并且意在包括第一个和第二个指示数字以及它们之间的所有小数和整数。术语“包括”、“含有”、“包含”、“具有”及其缀合物表示“包括但不限于”。术语“由...组成”是指“包括并限于”。术语“基本上由……组成”是指所述组合物,方法或结构可以包括另外的成分、步骤和/或部分,但仅当附加成分、步骤和/或部分不会实质性改变所要求保护的组合物、方法或结构的基本和新颖特征时。如本文所使用的,单数形式“一个”、“一种”和“该”包括复数引用,除非上下文另外明确指出。例如,术语“一种化合物”或“至少一种化合物”可以包括多种化合物,包括其混合物。在整个申请中,本发明的各种实施例可以以范围格式呈现。应当理解,范围格式的描述仅是为了方便和简洁,而不应被解释为对本发明范围的不灵活的限制。因此,应该认为范围的描述已经具体公开了所有可能的子范围以及该范围内的各个数值。例如,对范围(例如1到6)的描述应视为已明确公开了子范围,例如1到3、1到4、1到5、2到4、2到6、3至6等,以及该范围内的单个数字,例如1、2、3、4、5和6。这与范围的广度无关。如本文所使用的,术语“方法”是指用于完成给定任务的方式、手段、技术和过程,包括但不限于化学、药理、生物学、生化和医学领域的从业者已知或容易从已知方式、手段、技术和程序发展起来的那些方式、手段、技术和程序。如本文所用,术语“治疗”包括消除、基本上抑制、减慢或逆转病情的进展、基本上改善病情的临床或美学症状或基本上防止病情的临床或美学症状的出现。现在参考以下实施例将更容易理解一般描述的本发明,这些实施例仅出于说明本发明实施方案的某些方面的目的而包括在内。所述实例无意于限制本发明,如本领域技术人员将从以上教导和以下实例中认识到的,其他技术和方法可以满足权利要求,并且可以在不脱离所要求保护的本发明的范围的情况下被采用。确实,尽管已经参考本发明的优选实施例具体示出和描述了本发明,但是本领域技术人员将理解,在不脱离所附权利要求书所涵盖的本发明的范围的情况下,可以在形式和细节上进行各种改变。实施例实施例1:表达载体和基因修饰的细菌菌株的产生通过在pad-43-25质粒中的两个收敛的pupp启动子之间克隆dsrna序列来制备用于dsrna表达的质粒。如图2-3所示,本发明人生成了pad-dsrna质粒图和pad-43-25(表达pgfp)质粒图。rnaseiii缺陷型枯草芽孢杆菌菌株bg322用pad-dsrna质粒转化(3个用于wssv特异性dsrna表达的质粒–pad-dswsv477(seqidno.1)、pad-dsvp28(seqidno.2)、pad-dsvp19(seqidno.3),以及用于表达非特异性dslucpad-dsluc的质粒(seqidno.4)。实施例2:制备含有基因修饰的细菌的虾饲料细菌在含有5mg/ml氯霉素的lb中生长过夜,然后离心并混合入浓度为1e+08cfu/gm的普通商品虾饲料(zeiglerpl40)中并冷藏。将准备好的饲料以体重的10%饲喂给一克虾,每天分成三批。实施例3:基因修饰的细菌在虾肠中的定殖和持久性本发明人已经证明示例性的基因修饰的枯草芽孢杆菌菌株bg322能够在虾肠中定植并持续存在。在该实施方案中,本发明人用编码荧光gfp蛋白的质粒pad-43-25转化了bg322。向虾提供含bg322pad-43-25的饲料10天,并在第5天和第10天通过荧光显微镜下对虾肠的分析(在肠中检测到gfp荧光)和分离虾的肠板计数法(bg322菌落通过gfp荧光鉴定)检测到bg322的存在。bg322喂食5天后,bg322肠中的细菌滴度为~1.1e+06cfu/g,在第10天时保持在大致相同的水平(表1)。由于喂食5天后虾肠中细菌的浓度恒定,所以本发明人选择喂食细菌后的第5天作为开始病毒攻击的适当日期。实施例4:将dsrna表达载体导入虾在7加仑水族箱(n=12)中保持1g体重的spf虾(shrimpimprovementsystem,islamorada,fl)。虾被随机分为五个处理组:1)用商业食品喂虾,不注射病毒;2)注射商业食品的虾,注射bg322pad-dsluc/病毒;2)用bg322pad-dsvp19/病毒注射液喂虾商业饲料;3)用商业食品喂食bg322pad-dsvp28/病毒注射液的虾;4)用商业食品喂食bg322pad-dswsv477/病毒注射液的虾,以及5)用商业食品喂食bg322pad-dsluc/病毒注射液的虾。在病毒注射前和攻击过程中通过饲料为虾提供细菌5天。这种虾通常被称为“处理虾”。实施例5:在表达dsrna的(处理过的)虾中的wssv攻击本发明人通过注射源自虾的感染组织(厄瓜多尔2001株)的接种物,将处理过的虾暴露于wssv中,并储存在-80℃下。将组织在混合器中以组织与经紫外线处理的冷冻盐井水的比例为1:10匀化30秒钟。将制剂倒入50ml管中,并在4℃以2500rpm离心10分钟。将所得的上清液用0.45μm(pes)过滤器(whatman)过滤。制备后立即将滤液以20μl/g虾的重量肌肉注射到第二和第三尾段之间。实施例6:在表达dsrna(处理的)的虾中wssv诱导的死亡率计数本发明人在病毒注射后第7-10天进行死亡率计数。在该实施方案中,针对阴性对照组(无病毒)分析了6个生物学重复(单独的罐),对通过dsluc(非特异性dsrna)喂养的对虾进行4个重复,对通过bg322-dsvpl9、bg322-dsvp28或bg322-dswsv477(抗wssv特异性dsrna)喂养的每个对虾组进行了5个重复。从时间过程的最后几天开始的死亡率计数显示,与非特异性dsrna喂养的虾相比,以wssv为目标的dsrna喂养的虾死亡率降低了约50%。本发明人进一步证明,在分别由dsvp19、dsvp28和dswsv477饲喂的虾中观察到30%、33%或38%的平均死亡率,而dsluc饲喂组具有64%的死亡率。对于p<0.05,此差异具有统计学意义(图1)。这样,本发明人证明了表达病毒特异性dsrna的bg322提供了虾对病毒的保护。实施例7:对表达wssv-靶向dna的bg322饲喂的虾中wssv复制的序列特异性抑制本发明人使用qpcr来评估wssv攻击10天后虾中病毒复制的水平。来自喂食bg322-dsluc、bg322-dsvp19、bg322-dsvp28和bg322-dswsv477组的幸存者用于总dna提取(n=4-6)。用invitrogen组织dna试剂盒提取总dna,并使用nanodropnd-100分光光度计(nanodroptechnologies)定量提取的dna浓度。50-100ng的总dna用作qpcr的模板。用sybrgreenpowerup预混液进行sybrgreen测定。产生69bp复制子的引物wssv1011(5’)和wssv1079(3’)用于反应(表2)(durand等人,2002)。循环条件如下:第一步是在95℃下进行10分钟的初始步骤,然后是在95℃下进行30s、58℃下进行30s和72℃下进行30s的40个循环。检查每个样品的解链温度以验证pcr产物的纯度。pcr反应在stratagenemx3005实时pcr系统上进行,并使用mx3005软件进行分析。为了进行比较,将得出的数字根据bg322-dsluc(阳性对照组)饲喂的虾的读数进行归一化。本发明人观察到与表达非特异性dslucdsrna的虾饲喂细菌的虾的细菌相比,在所有以dssv基因为靶标的wssv基因(vp19,vp28,wsv477)的虾饲喂细菌的虾中检测到的病毒数量减少了3-4倍(负控制)。如图4所示,非特异性dsrna组与其他组之间的差异在p<0.01上具有统计学意义。实施例8:在wssv感染的虾中检测抗病毒vp19sirna本发明人通过对从经wssv感染而幸存的bgg322vp19喂养的虾中纯化的小rna样品的northern印迹分析证实了细菌特异性小rna的积累。在该实施方案中,本发明人制备了针对小rna的dig3’末端标记的探针。表3列出了与vp19dsrna互补的dna寡核苷酸序列。通过idtdna订购探针,并用dig寡核苷酸3'末端标记试剂盒进行标记。加入200mmedta(ph8.0)终止反应。标记的dna探针存储在-20℃。根据制造商的说明,使用e.z.n.amirna分离试剂盒(omega)从虾肌肉样品中分离总rna。通过在15%聚丙烯酰胺-8m尿素凝胶上进行电泳来评估总rna的质量和完整性。使用nanodropnd-100分光光度计(nanodroptechnologies)对提取的rna的浓度进行定量。在含有8m尿素的变性15%聚丙烯酰胺凝胶中,将至少10μg总rna进行电泳,然后转移至带正电的尼龙膜(roche)。与紫外线交联后,将膜在perfecthyb(sigma)中预杂交1h,然后在44℃下与洋地黄毒苷(dig)标记的与dsrna序列互补的dna探针杂交20h。膜用补充有0.1%sds(分别为2×、0.5×和0.1×)的ssc洗涤缓冲液洗涤四次。按照dighighprimedna标记和检测入门工具包ii(roche)的说明进行信号检测。14nt、18nt、22nt、26nt和30nt的ssrna标记用作小rna(takara)的标记。最后,杂交信号通过bio-radchemidocxrs可视化。如图5中总体所示,本发明人在从被感染的虾制备的样品中显示了强病毒特异的小rna,证实了病毒感染通过小干扰rna抑制了vp19基因表达。在未感染wssv的天然虾中未检测到显着的小病毒rna(泳道4)。在三分之二的虾中发现了大小为26nt至18nt的vp19特异性sirna的积累(1-2条泳道)。该结果证明bgg322vp19抗病毒保护是通过sirna途径介导的。参考文献以下参考文献通过引用整体并入本文:[1]sanjuktham,stalinrajv,aravindank,alavandisv,poornimam,santiagotc.comparativeefficacyofdouble-strandedrnastargetingwssvstructuralandnonstructuralgenesincontrollingviralmultiplicationinpenaeusmonodon.archvirol.2012may;157(5):993-8.doi:10.1007/s00705-012-1258-2.epub2012feb21.pubmedpmid:22350694.[2]kumar,anil&allnutt,fcthomas&sayre,richard.(2015).doublestrandedrnasimultaneouslytargetingfourwhitespotsyndromevirus(wssv)genesprovidesprotectionagainstwssvinlitopenaeusvannamei.internationaljournalofmarinescienceandoceantechnology.2.5-10.[3]lezzerinim,vandevenk,veermanm,bruls,budovskayayv.specificrnainterferenceincaenorhabditiselegansbyingesteddsrnaexpressedinbacillussubtilis.plosone.2015apr30;10(4):e0124508.doi:10.1371/journal.pone.0124508.ecollection2015.pubmedpmid:25928543;pubmedcentralpmcid:pmc4416053.[4]durand,s.v.andlightner,d.v.(2002),quantitativerealtimepcrforthemeasurementofwhitespotsyndromevirusinshrimp.journaloffishdiseases,25:381–389.doi:10.1046/j.1365-2761.2002.00367.x.[5]chakrabortya.,ottas.k.,kumarb.j.s.,hossainm.s.,karunasagari.,venugopalm.n.,karunasagari.,prevalenceofwhitespotsyndromevirusinwildcrustaceansalongthecoastofindia,curr.sci.(2002)82:1392–1397.).[6]nguyendv,christiaenso,bossierpandsmaggheg(2016)rnainterferenceinshrimpandpotentialapplicationsinaquaculture.reviewsinaquaculture(2016)0:1-12.doi:10.1111/raq.l2187.[7]sanchez-martinezj.g.,aguirre-guzmang.,mejia-ruizh.,whitespotsyndromevirusinculturedshrimp:areview,aquac.res.(2007)38:1339–1354.[8]flegelt.w.,nielsenl.,thamavitv.,kongtims.,pasharawipast.,presenceofmultiplevirusesinnon-diseased,cultivatedshrimpatharvest,aquaculture(2004)240:55-68.[9]hoat.,hodgsonr.,oanhd.,phuongn.,prestonn.,walkerp.,genotypicvariationsintandemrepeatdnasegmentsbetweenribonucleotidereductasesubunitgenesofwhitespotsyndromevirus(wssv)isolatesfromvietnam,in:walkerp.,lesterr.,reantasom.(eds.),diseasesinasianaquaculture,asianfisheriessociety,manila,2005,pp.395–403.[10]ma,d.;hu,y.;wang,j.;ye,s.&li,a.(2006).effectsofantibacterialsuseinaquacultureonbiogeochemicalprocessesinmarinesediment.thescienceofthetotalenvironment,vol.367,no.l,pp.273-277.[11]martinezcruz,patriciaetal."useofprobioticsinaquaculture."isrnmicrobiology2012(2012).[12]kobayashi,m.&brummett,r.2014.diseasemanagementinaquaculture.in:forumforagriculturalriskmanagementindevelopment.[13]walker,peterj.,andjamesr.winton."emergingviraldiseasesoffishandshrimp."veterinaryresearch41.6(2010).表表1.用表达gfp的bg322喂养的虾肠中的荧光负荷表2.qpcr反应中使用的引物wssv1011tggtcccgtcctcatctcagwssv1079gctgccttgccggaaatta表3.用于micrornanorthern印迹分析且与dsrnavp19互补的寡核苷酸表4:家禽种群中的目标病原体表5:蜜蜂种群中的目标病原体表6:哺乳动物种群中的目标病原体序列表如上所述,本申请包含完整的序列表,该序列表已以ascii格式电子提交,并在此全文引入作为参考。随此进一步提供以下序列,并在此将其完整引入本说明书中:seqidno.1dnadswsv477人工atgtatatcttcgtcgaaggttcccccctcacagggaagagttcatggatgtccaagttgatagatacaggatcatgtggaatgtctttcctcaattttcttcgtatgaacacttctgactactacaactggcctgccgaaatcgggacagaacatctccagttaggtttcagagaaaccagagtggtggatggaatgtttgaacctgtcctaaagacctttgtcgactcgtggaagaaagagcaaggaaaagagagtttgaaggaatatctggactacaacggccaagtcatggagatctacatcgcagaatggttgagacaaaggccactagccttccacgtgtttacctatacagatgaagctgtcaagagtggattcttgaacgaggaggatctagatatggatactgcaaccaagtggatggctgaaattattagagagaagaggggcaatattcaagaaataaaagtgacccctagagtagtcttcaatggcaatggttgtagtgcatgtttctctaacactaagagaaacttgtataactttggaacaaactataacaatgttgtacattgtgatttgttgtgcccttttgcaaggcataggattgtacatttcttataaseqidno.2dnadsvp28人工tcactctttcggtcgtgtcggccatcctcgccatcactgctgtgattgctgtatttattgtgatttttaggtatcacaacactgtgaccaagaccatcgaaacccacacaggcaatatcgagacaaacatggatgaaaacctccgcattcctgtgactgctgaggttggatcaggctacttcaagatgactgatgtgtcctttgacagcgacaccttgggcaaaatcaagatccgcaatggaaagtctgatgcacagatgaaggaagaagatgcggatcttgtatcactcccgtggagggccgagcactcgaagtgactgtggggcggaatctcacctttgaggggacattcaaggtgtggaacaacacatcaagaaagatcaacatcactggtatgcagatggtgccaaagattaacccatcaaaggcctttgtcggtagctccaacacctcctccttcacccccgtctctattgatgaggatgaagttggcacctttgtgtgtggtaccacctttggcgcaccaattgcagctaccgccggtggaaatcttttcgacatgtacgtgcacgtcacctactctggcactgagaccgagtaaseqidno.3dnadsvp19人工gccaccacgactaacactcttcctttcggcaggaccggagcccaggccgctggcccttcttacaccatggaagatcttgaaggctccatgtctatggctcgcatgggtctctttttgatcgttgctatctcaattggtatcctcgtcctggccgtcatgaatgtatggatgggaccaaagaaggacagcgattctgacactgataaggacaccgatgatgatgacgacactgccaacgataacgatgatgaggacaaatataagaacaggaccagggatatgatgcttctggctgggtccgctcttctgttcctcgtttccgccgccaccgtttttatgtcttaccccaaseqidno.4dnadsluc人工acagcctgggcatcagcaagcccaccatcgtgttcagcagcaagaagggcctggacaaagtcatcaccgtgcagaaaaccgtgaccaccatcaagaccatcgtgatcctggacagcaaggtggactaccggggctaccagtgcctggacaccttcatcaagcggaacaccccccctggcttccaggccagcagcttcaagaccgtggaggtggaccggaaagaacaggtggccctgatcatgaacagcagcggcagcaccggcctgcccaagggcgtgcagctgacccacgagaacaccgtgacccggttcagccacgccagggaccccatctacggcaaccaggtgtcccccggcaccgccgtgctgaccgtggtgcccttccaccacggcttcggcatgttcaccaccctgggctacctgatctgcggcttccgggtggtgatgctgaccaagttcgacgaggaaaccttcctgaaaaccctgcaggactacaagtgcacctacgtgattctggtgcccaccctgttcgccatcctgaacaagagcgagctgctgaacaseqidno.5氨基酸vp19白斑综合症病毒matttntlpfgrtgaqaagpsytmedlegsmsmarmglflivaisigilvlavmnvwmgpkkdsdsdtdkdtdddddtandnddedkyknrtrdmmllagsallflvsaatvfmsypkrrqseqidno.6dnavp19白斑综合症病毒atggccaccacgactaacactcttcctttcggcaggaccggagcccaggccgctggcccttcttacaccatggaagatcttgaaggctccatgtctatggctcgcatgggtctctttttgatcgttgctatctcaattggtatcctcgtcctggccgtcatgaatgtatggatgggaccaaagaaggacagcgattctgacactgataaggacaccgatgatgatgacgacactgccaacgataacgatgatgaggacaaatataagaacaggaccagggatatgatgcttctggctgggtccgctcttctgttcctcgtttccgccgccaccgtttttatgtcttaccccaagaggaggcagtaaseqidno.7氨基酸vp28白斑综合症病毒vtktiethtdnietnmdenlripvtaevgsgyfkmtdvsfdsdtlgkikirngksdaqmkeedadlvitpvegralevtvgqnltfegtfkvwnntsrkinitgmqmvpkinpskafvgssntssftpvsidedevgtfvcgttfgapiaataggnlfdmyvhvtysgteteseqidno.8dnavp28白斑综合症病毒gtgaccaagaccatcgaaacccacacagacaatatcgagacaaacatggatgaaaacctccgcattcctgtgactgctgaggttggatcaggctacttcaagatgactgatgtgtcctttgacagcgacaccttgggcaaaatcaagatccgcaatggaaagtctgatgcacagatgaaggaagaagatgcggatcttgtcatcactcccgtggagggccgagcactcgaagtgactgtggggcagaatctcacctttgagggaacattcaaggtgtggaacaacacatcaagaaagatcaacatcactggtatgcagatggtgccaaagattaacccatcaaaggcctttgtcggtagctccaacacctcctccttcacccccgtctctattgatgaggatgaagttggcacctttgtgtgtggtaccacctttggcgcaccaattgcagctaccgccggtggaaatcttttcgacatgtacgtgcacgtcacctactctggcactgagaccgagseqidno.9氨基酸wsv477白斑综合症病毒myifvegspltgksswmsklidtgscgmsflnflrmntsdyynwpaeigtehlqlgfretrvvdgmfepvlktfvdswkkeqgkeslkeyldyngqvmeiyiaewlrqrplafhvftytdeavksgflneedldmdtatkwmaeiirekrgniqeikvtprvvfngngcsacfsntkrnlynfgtnynnvvhcdllcpfarhrivhflseqidno.10dnawsv477白斑综合症病毒atgtatatcttcgtcgaaggttcccccctcacagggaagagttcatggatgtccaagttgatagatacaggatcatgtggaatgtctttcctcaattttcttcgtatgaacacttctgactactacaactggcctgccgaaatcgggacagaacatctccagttaggtttcagagaaaccagagtggtggatggaatgtttgaacctgtcctaaagacctttgtcgactcgtggaagaaagagcaaggaaaagagagtttgaaggaatatctggactacaacggccaagtcatggagatctacatcgcagaatggttgagacaaaggccactagccttccacgtgtttacctatacagatgaagctgtcaagagtggattcttgaacgaggaggatctagatatggatactgcaaccaagtggatggctgaaattattagagagaagaggggcaatattcaagaaataaaagtgacccctagagtagtcttcaatggcaatggttgtagtgcatgtttctctaacactaagagaaacttgtataactttggaacaaactataacaatgttgtacattgtgatttgttgtgcccttttgcaaggcataggattgtacatttcttataa序列表<110>卵石实验室公司<120>水产养殖和其他动物系统中病原菌生物控制的新系统<130>90115.00200<150>us62/480,138<151>2017-03-31<160>10<170>patentinversion3.5<210>1<211>627<212>dna<213>人工(artificial)<220><221><222><223>dswsv477<400>1atgtatatcttcgtcgaaggttcccccctcacagggaagagttcatggatgtccaagttg60atagatacaggatcatgtggaatgtctttcctcaattttcttcgtatgaacacttctgac120tactacaactggcctgccgaaatcgggacagaacatctccagttaggtttcagagaaacc180agagtggtggatggaatgtttgaacctgtcctaaagacctttgtcgactcgtggaagaaa240gagcaaggaaaagagagtttgaaggaatatctggactacaacggccaagtcatggagatc300tacatcgcagaatggttgagacaaaggccactagccttccacgtgtttacctatacagat360gaagctgtcaagagtggattcttgaacgaggaggatctagatatggatactgcaaccaag420tggatggctgaaattattagagagaagaggggcaatattcaagaaataaaagtgacccct480agagtagtcttcaatggcaatggttgtagtgcatgtttctctaacactaagagaaacttg540tataactttggaacaaactataacaatgttgtacattgtgatttgttgtgcccttttgca600aggcataggattgtacatttcttataa627<210>2<211>601<212>dna<213>人工(artificial)<220><221><222><223>dsvp28<400>2tcactctttcggtcgtgtcggccatcctcgccatcactgctgtgattgctgtatttattg60tgatttttaggtatcacaacactgtgaccaagaccatcgaaacccacacaggcaatatcg120agacaaacatggatgaaaacctccgcattcctgtgactgctgaggttggatcaggctact180tcaagatgactgatgtgtcctttgacagcgacaccttgggcaaaatcaagatccgcaatg240gaaagtctgatgcacagatgaaggaagaagatgcggatcttgtatcactcccgtggaggg300ccgagcactcgaagtgactgtggggcggaatctcacctttgaggggacattcaaggtgtg360gaacaacacatcaagaaagatcaacatcactggtatgcagatggtgccaaagattaaccc420atcaaaggcctttgtcggtagctccaacacctcctccttcacccccgtctctattgatga480ggatgaagttggcacctttgtgtgtggtaccacctttggcgcaccaattgcagctaccgc540cggtggaaatcttttcgacatgtacgtgcacgtcacctactctggcactgagaccgagta600a601<210>3<211>350<212>dna<213>人工(artificial)<220><221><222><223>dsvp19<400>3gccaccacgactaacactcttcctttcggcaggaccggagcccaggccgctggcccttct60tacaccatggaagatcttgaaggctccatgtctatggctcgcatgggtctctttttgatc120gttgctatctcaattggtatcctcgtcctggccgtcatgaatgtatggatgggaccaaag180aaggacagcgattctgacactgataaggacaccgatgatgatgacgacactgccaacgat240aacgatgatgaggacaaatataagaacaggaccagggatatgatgcttctggctgggtcc300gctcttctgttcctcgtttccgccgccaccgtttttatgtcttaccccaa350<210>4<211>561<212>dna<213>人工(artificial)<220><221><222><223>dsluc<400>4acagcctgggcatcagcaagcccaccatcgtgttcagcagcaagaagggcctggacaaag60tcatcaccgtgcagaaaaccgtgaccaccatcaagaccatcgtgatcctggacagcaagg120tggactaccggggctaccagtgcctggacaccttcatcaagcggaacaccccccctggct180tccaggccagcagcttcaagaccgtggaggtggaccggaaagaacaggtggccctgatca240tgaacagcagcggcagcaccggcctgcccaagggcgtgcagctgacccacgagaacaccg300tgacccggttcagccacgccagggaccccatctacggcaaccaggtgtcccccggcaccg360ccgtgctgaccgtggtgcccttccaccacggcttcggcatgttcaccaccctgggctacc420tgatctgcggcttccgggtggtgatgctgaccaagttcgacgaggaaaccttcctgaaaa480ccctgcaggactacaagtgcacctacgtgattctggtgcccaccctgttcgccatcctga540acaagagcgagctgctgaaca561<210>5<211>121<212>prt<213>vp19白斑综合症病毒(vp19whitespotsyndromevirus)<400>5metalathrthrthrasnthrleupropheglyargthrglyalagln151015alaalaglyprosertyrthrmetgluaspleugluglysermetser202530metalaargmetglyleupheleuilevalalaileserileglyile354045leuvalleualavalmetasnvaltrpmetglyprolyslysaspser505560aspseraspthrasplysaspthraspaspaspaspaspthralaasn65707580aspasnaspaspgluasplystyrlysasnargthrargaspmetmet859095leuleualaglyseralaleuleupheleuvalseralaalathrval100105110phemetsertyrprolysargarggln115120<210>6<211>366<212>dna<213>vp19白斑综合症病毒(vp19whitespotsyndromevirus)<400>6atggccaccacgactaacactcttcctttcggcaggaccggagcccaggccgctggccct60tcttacaccatggaagatcttgaaggctccatgtctatggctcgcatgggtctctttttg120atcgttgctatctcaattggtatcctcgtcctggccgtcatgaatgtatggatgggacca180aagaaggacagcgattctgacactgataaggacaccgatgatgatgacgacactgccaac240gataacgatgatgaggacaaatataagaacaggaccagggatatgatgcttctggctggg300tccgctcttctgttcctcgtttccgccgccaccgtttttatgtcttaccccaagaggagg360cagtaa366<210>7<211>172<212>prt<213>vp28白斑综合症病毒(vp28whitespotsyndromevirus)<400>7valthrlysthrilegluthrhisthraspasnilegluthrasnmet151015aspgluasnleuargileprovalthralagluvalglyserglytyr202530phelysmetthraspvalserpheaspseraspthrleuglylysile354045lysileargasnglylysseraspalaglnmetlysglugluaspala505560aspleuvalilethrprovalgluglyargalaleugluvalthrval65707580glyglnasnleuthrphegluglythrphelysvaltrpasnasnthr859095serarglysileasnilethrglymetglnmetvalprolysileasn100105110proserlysalaphevalglyserserasnthrserserphethrpro115120125valserileaspgluaspgluvalglythrphevalcysglythrthr130135140pheglyalaproilealaalathralaglyglyasnleupheaspmet145150155160tyrvalhisvalthrtyrserglythrgluthrglu165170<210>8<211>516<212>dna<213>vp28白斑综合症病毒(vp28whitespotsyndromevirus)<400>8gtgaccaagaccatcgaaacccacacagacaatatcgagacaaacatggatgaaaacctc60cgcattcctgtgactgctgaggttggatcaggctacttcaagatgactgatgtgtccttt120gacagcgacaccttgggcaaaatcaagatccgcaatggaaagtctgatgcacagatgaag180gaagaagatgcggatcttgtcatcactcccgtggagggccgagcactcgaagtgactgtg240gggcagaatctcacctttgagggaacattcaaggtgtggaacaacacatcaagaaagatc300aacatcactggtatgcagatggtgccaaagattaacccatcaaaggcctttgtcggtagc360tccaacacctcctccttcacccccgtctctattgatgaggatgaagttggcacctttgtg420tgtggtaccacctttggcgcaccaattgcagctaccgccggtggaaatcttttcgacatg480tacgtgcacgtcacctactctggcactgagaccgag516<210>9<211>208<212>prt<213>wsv477白斑综合症病毒(wsv477whitespotsyndromevirus)<400>9mettyrilephevalgluglyserproleuthrglylyssersertrp151015metserlysleuileaspthrglysercysglymetserpheleuasn202530pheleuargmetasnthrserasptyrtyrasntrpproalagluile354045glythrgluhisleuglnleuglyphearggluthrargvalvalasp505560glymetphegluprovalleulysthrphevalaspsertrplyslys65707580gluglnglylysgluserleulysglutyrleuasptyrasnglygln859095valmetgluiletyrilealaglutrpleuargglnargproleuala100105110phehisvalphethrtyrthraspglualavallysserglypheleu115120125asnglugluaspleuaspmetaspthralathrlystrpmetalaglu130135140ileileargglulysargglyasnileglngluilelysvalthrpro145150155160argvalvalpheasnglyasnglycysseralacyspheserasnthr165170175lysargasnleutyrasnpheglythrasntyrasnasnvalvalhis180185190cysaspleuleucysprophealaarghisargilevalhispheleu195200205<210>10<211>627<212>dna<213>wsv477白斑综合症病毒(wsv477whitespotsyndromevirus)<400>10atgtatatcttcgtcgaaggttcccccctcacagggaagagttcatggatgtccaagttg60atagatacaggatcatgtggaatgtctttcctcaattttcttcgtatgaacacttctgac120tactacaactggcctgccgaaatcgggacagaacatctccagttaggtttcagagaaacc180agagtggtggatggaatgtttgaacctgtcctaaagacctttgtcgactcgtggaagaaa240gagcaaggaaaagagagtttgaaggaatatctggactacaacggccaagtcatggagatc300tacatcgcagaatggttgagacaaaggccactagccttccacgtgtttacctatacagat360gaagctgtcaagagtggattcttgaacgaggaggatctagatatggatactgcaaccaag420tggatggctgaaattattagagagaagaggggcaatattcaagaaataaaagtgacccct480agagtagtcttcaatggcaatggttgtagtgcatgtttctctaacactaagagaaacttg540tataactttggaacaaactataacaatgttgtacattgtgatttgttgtgcccttttgca600aggcataggattgtacatttcttataa627当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1