阿尔茨海默病新突变、其稳转细胞模型及医药用途的制作方法
本发明属于生物医药领域,涉及一种阿尔茨海默病新突变、其稳转细胞模型及医药用途。具体地,本发明涉及家族性阿尔茨海默病致病基因psen1编码的早老素蛋白1(presenilin1)2个新突变位点。
背景技术:
阿尔茨海默病(alzheimer’sdisease,ad)是最常见的痴呆类型,其三大致病基因psen11、app2和psen23,4的突变及其临床病例以及生化和动物模型的研究共同指向aβ对ad的致病作用,对其家系的报道和突变的探究有助于进一步揭示ad发病机制。
aβ由app加工生成。app加工有两条竞争性途径5:一条途径是不生成aβ,即先被α-分泌酶剪切生成sappα和αapp-ctf(c83),c83再被γ-分泌酶剪切生成与发生ad无关的p3和aicd;另一条途径是生成aβ,即先被β-分泌酶(bace1)剪切生成sappβ和βapp-ctf(c99)6,c99再在膜内被γ-分泌酶剪切:γ-分泌酶先对app的跨膜/胞浆内侧面发挥内肽酶活性,剪切生成aβ48/49或aβ49/50(ε-剪切),随后发挥羧肽酶活性多次剪切,每次按顺序从c端移除一圈helix包含3或4个氨基酸,产生两种产物链(aβ49/46/43/40和aβ48/45/42/38)7-11。其中,aβ40和aβ42是最为常见的与发生ad相关的产物,而且一般情况下主要生成aβ40,aβ42仅占总分泌型aβ的约10%12。
psen1基因突变导致的ad是ad的少见类型,但在三大致病基因突变中最为常见。携带该基因突变的大多数家系符合常染色体显性遗传,发病早、进展快、预后差、无特效治疗方法,在身心、经济、社会功能等方面对患者及其家庭的生活质量构成巨大威胁。正常情况下,psen1基因表达生成ps1-fl蛋白(后面亦简称为ps1),ps1-fl蛋白本身不具有γ-分泌酶催化活性,它通过自身的内切水解作用形成具有γ-分泌酶催化活性的ps1-ntf和ps1-ctf。ps1-ctf和ps1-ntf形成异二聚体(heterodimers),与另外的其它三种蛋白共同构成γ-分泌酶复合体。ps1-ctf和ps1-ntf均具有天冬氨酸催化位点(d257和d385),作为γ-分泌酶复合体的催化亚单位共同发挥对app的剪切作用。ps1-ntf和ps1-ctf上面各有一个发挥内切水解作用和γ-分泌酶活性所必需的位点,这两个位点的突变(例如d385a突变)都可抑制ps1的内切水解作用并废除ps1的催化活性13。当psen1基因错义突变导致ps1蛋白氨基酸改变时(例如d257和d385发生突变),可能直接影响γ-分泌酶催化活性或通过引起ps1蛋白构象改变从而影响γ-分泌酶催化活性,使底物app剪切发生异常。psen1突变使γ-分泌酶的羧肽酶活性减弱,导致更长aβ肽的产量相对增加,更长的aβ肽更为疏水,更易自我聚集,更具神经毒性,继而引起aβ沉积等ad的标志性病理改变,导致ad发病。
apoe基因是目前ad最主要的风险基因,有ε2/ε3/ε4这三种等位碱基形式,其中ε3是人群中最常见的类型,ε4被认为增加ad发病风险,ε2则被认为对ad具有保护作用(cordereh,etal.protectiveeffectofapolipoproteinetype2alleleforlateonsetalzheimerdisease.natgenet.1994jun;7(2):180-4.)。与其它类型的apoe相比,apoeε4对aβ的分解作用较弱,这导致apoeε4携带者更易罹患ad(jiangqetal.apoepromotestheproteolyticdegradationofabeta.neuron.2008jun12;58(5):681-93.)。
目前,尚需要发现新的阿尔茨海默病致病基因突变。
技术实现要素:
本发明人对与γ-分泌酶催化活性密切相关的2个psen1新突变报道其家系临床表型,并通过慢病毒表达载体体系构建其神经细胞模型探究对ad的致病机制。这2个fad突变的临床和基础模型研究将对揭示psen1基因在ad发病机制中的作用具有重要意义。
具体地,本发明人在两个中国ad家系中发现了与γ-分泌酶催化活性密切相关的2个psen1新突变,其临床特点分别为精神症状、缺血和白血病风险升高,以及超早发。通过慢病毒表达载体体系成功建立了这两种突变的稳转细胞模型,发现p433s突变显著抑制ps1-fl蛋白的内切水解作用,而i249l突变对ps1-fl蛋白的内切水解无明显抑制作用。i249l和p433s都能引起aβ42和或aβ42/40比值的升高,支持这2种fad新突变的致病作用。对ps1内切水解作用的分析和对aβ40、42、43的分析表明,这2种新突变与d385a突变的致病机制存在不同。该模型的应用将有助进一步揭示psen1基因在ad发病机制中的作用。
由此提供了下述发明:
本发明的一个方面一种突变的蛋白,其为野生型人psen1蛋白发生选自如下的任意1个或者2个突变:
i249l和p433s;
优选地,所述野生型人psen1蛋白的氨基酸序列如seqidno:5所示;
优选地,所述蛋白的氨基酸序列如seqidnos:1-2中任一序列所示。
表述形式i249l或p433s具有本领域技术人员知悉的含义,例如,分别是指野生型人psen1蛋白的第249个氨基酸由异亮氨酸(i)突变为亮氨酸(l),第433位的脯氨酸(p)突变为丝氨酸(s)。
psen1i249l的氨基酸序列如下:467aa,突变位点用边框标出。
psen1p433s的氨基酸序列如下:467aa,突变位点用边框标出。
在本发明的一些实施方式中,所述蛋白为分离的蛋白。
在本发明的一些实施方式中,两个突变是i249l和p433s。
本发明的另一方面涉及一种核酸分子,其编码本发明所述的蛋白;优选地,所述核酸分子如seqidnos:3-4中任一序列所示。
编码psen1i249l的核苷酸序列如下:1404bp,突变位点用边框标出。
编码psen1p433s的核苷酸序列如下:1404bp,突变位点用边框标出。
在本发明的一些实施方式中,所述核酸分子为分离的核酸分子。
本发明还涉及一种核酸构建体,其含有本发明所述的核酸分子;优选地,所述核酸构建体为重组载体;优选地,所述重组载体为重组表达载体。
本发明还涉及一种重组宿主细胞,其表达本发明所述的蛋白,或者含有本发明所述的核酸分子或者本发明所述的核酸构建体;
优选地,所述重组宿主细胞为重组的人神经元细胞;
优选地,所述重组宿主细胞为重组的人神经母细胞瘤sh-sy5y细胞。
本发明的再一方面涉及一种组合物,其包含本发明的蛋白、本发明的核酸分子、本发明的核酸构建体或者本发明的重组宿主细胞。可选地,所述组合物还包含一种或多种药学上可接受的辅料。
本发明的再一方面涉及一种单克隆抗体或其抗原结合片段,其能够特异性地结合本发明的蛋白。
本发明的再一方面涉及一种偶联物,其包含本发明的单克隆抗体或其抗原结合片段以及偶联部分,其中,所述偶联部分为可检测的标记;优选地,所述偶联部分为放射性同位素、荧光物质、发光物质、有色物质或酶。
本发明的再一方面涉及一种试剂盒,其包含本发明的单克隆抗体或其抗原结合片段,或者包括本发明的偶联物;
优选地,所述试剂盒还包括第二抗体,其特异性识别所述单克隆抗体或其抗原结合片段;任选地,所述第二抗体还包括可检测的标记,例如放射性同位素、荧光物质、发光物质、有色物质或酶;
可选地,所述试剂盒还包含说明书。
本发明的再一方面涉及一种引物或探针,其能够特异性地结合本发明的核酸分子;
优选地,所述探针的5’端标记有荧光报告基团,3’端标记有荧光淬灭基团;
优选地,所述荧光报告基团选自fam、hex、vic、rox和cy5;
优选地,所述荧光淬灭基团选自bhq1、tamra、joe、bhq2和bhq3。
本发明的再一方面涉及一种试剂盒,其包含本发明的引物或探针;
可选地,所述试剂盒还包含说明书。
本发明的再一方面涉及选自如下的(1)-(8)项中的任意一项在制备治疗和/或预防或者诊断阿尔茨海默病的药物中的用途;或者在制备降低aβ42水平和/或降低aβ42/40比值的药物中的用途;或者在制备筛选药物的模型例如细胞模型或者动物模型中的用途,所述药物用于治疗和/或预防和/或诊断阿尔茨海默病:
(1)本发明所述的蛋白;
(2)本发明所述的核酸分子;
(3)抑制或阻断本发明所述的蛋白的药物,例如本发明所述的单克隆抗体或其抗原结合片段;
(4)抑制或降低本发明所述的核酸分子的表达水平的药物;
(5)将本发明的蛋白修复为野生型人psen1蛋白(例如seqidno:5所示)的药物;
(6)将本发明的核酸分子修复为野生型人psen1基因(例如seqidno:6所示)的药物;
(7)检测本发明所述的蛋白的药物,例如本发明所述的单克隆抗体或其抗原结合片段或者本发明所述的偶联物;
(8)检测本发明所述的核酸分子的表达水平的药物,例如本发明所述的引物或探针;
优选地,所述阿尔茨海默病为家族性阿尔茨海默病;优选地,所述阿尔茨海默病为早发性家族性阿尔茨海默病。
上述第(5)项或(6)项中,所述药物可以包括设计的引物,将突变的基因修复为野生型人psen1基因,通过本领域技术人员知悉的基因转染或者转导手段,转入到细胞中表达野生型蛋白。
野生型人psen1蛋白的氨基酸序列如下:467aa
mtelpaplsyfqnaqmsednhlsntvrsqndnrerqehndrrslghpeplsngrpqgnsrqvveqdeeedeeltlkygakhvimlfvpvtlcmvvvvatiksvsfytrkdgqliytpftedtetvgqralhsilnaaimisvivvmtillvvlykyrcykvihawliissllllfffsfiylgevfktynvavdyitvalliwnfgvvgmisihwkgplrlqqaylimisalmalvfikylpewtawlilavisvydlvavlcpkgplrmlvetaqernetlfpaliysstmvwlvnmaegdpeaqrrvsknskynaesteresqdtvaenddggfseeweaqrdshlgphrstpesraavqelsssilagedpeergvklglgdfifysvlvgkasatasgdwnttiacfvailiglcltllllaifkkalpalpisitfglvfyfatdylvqpfmdqlafhqfyi(seqidno:5)
野生型人psen1基因的读码框序列如下:1404bp
atgacagagttacctgcaccgttgtcctacttccagaatgcacagatgtctgaggacaaccacctgagcaatactgtacgtagccagaatgacaatagagaacggcaggagcacaacgacagacggagccttggccaccctgagccattatctaatggacgaccccagggtaactcccggcaggtggtggagcaagatgaggaagaagatgaggagctgacattgaaatatggcgccaagcatgtgatcatgctctttgtccctgtgactctctgcatggtggtggtcgtggctaccattaagtcagtcagcttttatacccggaaggatgggcagctaatctataccccattcacagaagataccgagactgtgggccagagagccctgcactcaattctgaatgctgccatcatgatcagtgtcattgttgtcatgactatcctcctggtggttctgtataaatacaggtgctataaggtcatccatgcctggcttattatatcatctctattgttgctgttctttttttcattcatttacttgggggaagtgtttaaaacctataacgttgctgtggactacattactgttgcactcctgatctggaattttggtgtggtgggaatgatttccattcactggaaaggtccacttcgactccagcaggcatatctcattatgattagtgccctcatggccctggtgtttatcaagtacctccctgaatggactgcgtggctcatcttggctgtgatttcagtatatgatttagtggctgttttgtgtccgaaaggtccacttcgtatgctggttgaaacagctcaggagagaaatgaaacgctttttccagctctcatttactcctcaacaatggtgtggttggtgaatatggcagaaggagacccggaagctcaaaggagagtatccaaaaattccaagtataatgcagaaagcacagaaagggagtcacaagacactgttgcagagaatgatgatggcgggttcagtgaggaatgggaagcccagagggacagtcatctagggcctcatcgctctacacctgagtcacgagctgctgtccaggaactttccagcagtatcctcgctggtgaagacccagaggaaaggggagtaaaacttggattgggagatttcattttctacagtgttctggttggtaaagcctcagcaacagccagtggagactggaacacaaccatagcctgtttcgtagccatattaattggtttgtgccttacattattactccttgccattttcaagaaagcattgccagctcttccaatctccatcacctttgggcttgttttctactttgccacagattatcttgtacagccttttatggaccaattagcattccatcaattttatatctag(seqidno:6)
本发明的再一方面涉及一种治疗和/或预防阿尔茨海默病的方法或者一种降低aβ42水平和/或降低aβ42/40比值的方法,包括减少或者降低受试者中的seqidnos:1-2中任一序列所示蛋白的水平的步骤,或者包括减少或者降低受试者中的seqidnos:3-4中任一序列所示核酸分子的水平的步骤。在本发明的一个实施方案中,所述方法包括将seqidnos:1-2中任一序列所示的蛋白修正为相应的野生型蛋白的步骤。在本发明的一个实施方案中,所述方法包括将seqidnos:3-4中任一序列所示的核酸分子修正为相应的野生型核酸分子的步骤。
本发明的再一方面涉及一种诊断阿尔茨海默病的方法,包括检测受试者是否存在seqidnos:1-2中任一序列所示的蛋白的步骤,或者包括检测受试者是否存在seqidnos:3-4中任一序列所示的核酸分子的步骤;如果存在所述蛋白或核酸分子,则诊断为阳性。
本发明的再一方面涉及一种筛选治疗和/或预防阿尔茨海默病的药物的方法,包括检测候选药物是否减少或者降低受试者中或者细胞中的seqidnos:1-2中任一序列所示的蛋白的水平的步骤,或者包括检测候选药物是否减少或者降低受试者中或者细胞中的seqidnos:3-4中任一序列所示的核酸分子的水平的步骤。如果受试者中或者细胞中的所述蛋白或核酸的水平降低或者减少,则作为阳性药物。可选地,以不加入候选药物的细胞作为对照。
受试者中或者细胞中的所述蛋白或核酸的水平降低或者减少,可以是相对于向受试者给药之前的该蛋白或核酸的水平,或者相对于向细胞中加入候选药物之前的该蛋白或核酸的水平。
在本发明中,除非另有说明,否则本文中使用的科学和技术名词具有本领域技术人员所通常理解的含义。并且,本文中所用的细胞培养、分子遗传学、核酸化学、免疫学实验室操作步骤均为相应领域内广泛使用的常规步骤。同时,为了更好地理解本发明,下面提供相关术语的定义和解释。
本发明中,术语“分离的”或“被分离的”指的是,从天然状态下经人工手段获得的。如果自然界中出现某一种“分离”的物质或成分,那么可能是其所处的天然环境发生了改变,或从天然环境下分离出该物质,或二者情况均有发生。例如,某一活体动物体内天然存在某种未被分离的多聚核苷酸或多肽,而从这种天然状态下分离出来的高纯度的相同的多聚核苷酸或多肽即称之为分离的。术语“分离的”或“被分离的”不排除混有人工或合成的物质,也不排除存在不影响物质活性的其它不纯物质。
本发明中,术语“核酸构建体”,在文中定义为单链或双链核酸分子,优选是指人工构建的核酸分子。可选地,所述核酸构建体还包含有可操作地连接的1个或多个调控序列。
本发明中,术语“可操作地连接”是指两个或多个核苷酸区域或核酸序列的功能性的空间排列。所述“可操作地连接”可以通过基因重组的手段实现。
本发明中,术语“载体”指的是,可将抑制某蛋白的多核苷酸插入其中的一种核酸运载工具。举例来说,载体包括:质粒;噬菌粒;柯斯质粒;人工染色体如酵母人工染色体(yac)、细菌人工染色体(bac)或p1来源的人工染色体(pac);噬菌体如λ噬菌体或m13噬菌体及动物病毒等。用作载体的动物病毒种类有逆转录酶病毒(包括慢病毒)、腺病毒、腺相关病毒、疱疹病毒(如单纯疱疹病毒)、痘病毒、杆状病毒、乳头瘤病毒、乳头多瘤空泡病毒(如sv40)。一种载体可能含有多种控制表达的元件。
本发明中,术语“宿主细胞”指的是导入载体的细胞,包括如下许多细胞类型,如大肠杆菌或枯草菌等原核细胞,如酵母细胞或曲霉菌等真菌细胞,如s2果蝇细胞或sf9等昆虫细胞,或者如纤维原细胞,cho细胞,cos细胞,nso细胞,hela细胞,bhk细胞,hek293细胞或人细胞的动物细胞。
本发明中,术语“单克隆抗体”或“单抗”是指,来自一群高度同源的抗体分子中的一个抗体或抗体的一个片断,也即除可能自发出现的自然突变外,一群完全相同的抗体分子。单抗对抗原上的单一表位具有高特异性。多克隆抗体是相对于单克隆抗体而言的,其通常包含至少2种或更多种的不同抗体,这些不同的抗体通常识别抗原上的不同表位。单克隆抗体通常可采用kohler等首次报道的杂交瘤技术获得(nature,256:495,1975),但也可采用重组dna技术获得(如参见u.s.p4,816,567)。
本发明中,术语“抗原结合片段”是指包含全长抗体的片段的多肽,其保持特异性结合全长抗体所结合的相同抗原的能力,和/或与全长抗体竞争对抗原的特异性结合,其也被称为“抗原结合部分”。通常参见,fundamentalimmunology,ch.7(paul,w.,ed.,第2版,ravenpress,n.y.(1989),其以其全文通过引用合并入本文,用于所有目的。可通过重组dna技术或通过完整抗体的酶促或化学断裂产生抗体的抗原结合片段。在一些情况下,抗原结合片段包括fab、fab'、f(ab')2、fd、fv、dab和互补决定区(cdr)片段、单链抗体(例如,scfv)、嵌合抗体、双抗体(diabody)和这样的多肽,其包含足以赋予多肽特异性抗原结合能力的抗体的至少一部分。
在一些情况下,抗原结合片段是单链抗体(例如,scfv),其中vl和vh结构域通过使其能够产生为单个多肽链的连接体配对形成单价分子(参见,例如,bird等人,science242:423426(1988)和huston等人,proc.natl.acad.sci.usa85:58795883(1988))。此类scfv分子可具有一般结构:nh2-vl-接头-vh-cooh或nh2-vh-接头-vl-cooh。合适的现有技术接头由重复的ggggs氨基酸序列或其变体组成。例如,可使用具有氨基酸序列(ggggs)4的接头,但也可使用其变体(holliger等人(1993),proc.natl.acad.sci.usa90:6444-6448)。可用于本发明的其他接头由alfthan等人(1995),proteineng.8:725-731,choi等人(2001),eur.j.immunol.31:94-106,hu等人(1996),cancerres.56:3055-3061,kipriyanov等人(1999),j.mol.biol.293:41-56和roovers等人(2001),cancerimmunol.描述。
本发明中,术语“app(beta淀粉样蛋白前体,amyloidprecursorprotein)”编码aβ前体蛋白,是aβ的直接来源,app经β-分泌酶剪切后产生c99,c99继而被γ-分泌酶剪切产生aβ。psen1和psen2基因分别编码早老素1和早老素2,是γ-分泌酶的催化亚单位。由此可见,app、psen1、psen2基因编码的蛋白都位于app酶切通路上,突变可能是通过影响app的酶切位点和早老素蛋白的结构,从而影响app的酶切过程,导致aβ生成异常。
本发明中,术语“aβ”是app依次经β-分泌酶和γ-分泌酶酶切的代谢产物,是包含约40个氨基酸的多肽。aβ40(含40个氨基酸)和aβ42(含42个氨基酸)是aβ的主要类型,aβ40约占80%-90%,aβ42约占5%-10%,其他更长的aβ肽则更少。aβ异常聚集形成淀粉样斑块的核心,是ad的主要病理特征之一。aβ40抑制淀粉样斑块沉积,而aβ42、aβ43和更长的aβ肽疏水性更强,更易聚集形成斑块和具有神经毒性,可导致淀粉样斑块(amyloidplaque)沉积。aβ42/aβ40比值的升高与ad的突触功能障碍、神经元丢失和认知损害直接相关,这强烈提示ad致病性。含有aβ43的斑块见于psen1l435f突变患者的脑中。aβ43在一些psen1突变中升高,可能具有ad致病性。
aβ40的氨基酸序列如下:(40aa)
daefrhdsgyevhhqklvffaedvgsnkgaiiglmvggvv(seqidno:7)
aβ42的氨基酸序列如下:(42aa)
daefrhdsgyevhhqklvffaedvgsnkgaiiglmvggvvia(seqidno:8)
aβ43的氨基酸序列如下:(43aa)
daefrhdsgyevhhqklvffaedvgsnkgaiiglmvggvviat
(seqidno:9)
本发明中,术语“有效量”是指可在受试者中实现治疗、预防、减轻和/或缓解本发明所述疾病或病症的剂量。
术语“疾病和/或病症”是指所述受试者的一种身体状态,该身体状态与本发明所述疾病和/或病症有关。
术语“受试者”可以指患者或者其它接受本发明药物组合物以治疗、预防、减轻和/或缓解本发明所述疾病或病症的动物,特别是哺乳动物,例如人、狗、猴、牛、马等。
发明的有益效果
本发明人在中国eofad中发现psen12个新突变,它们与各家系ad的发病密切相关,具有应用于制备治疗和/或预防或者诊断阿尔茨海默病的药物或者试剂的潜力。
附图说明
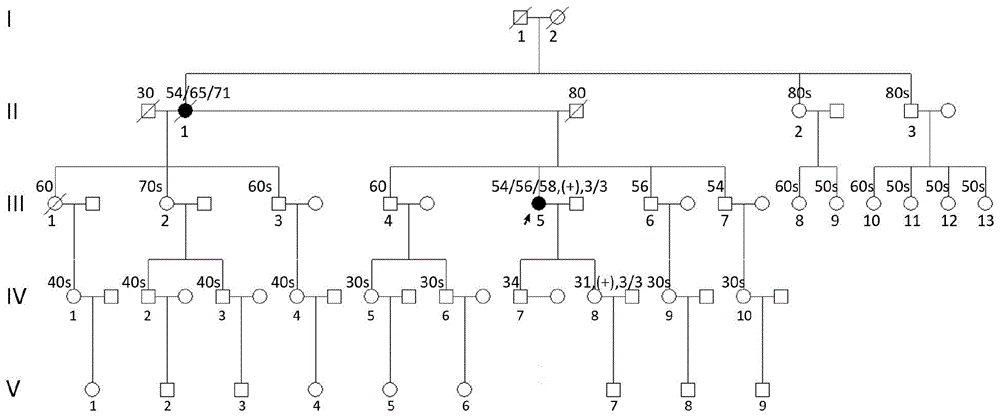
图1a:psen1i249l突变家系图。数字表示发病年龄、痴呆年龄以及就诊时年龄或死亡年龄。“(+)”或“(-)”表示是否携带psen1该新突变,“3/3”表示apoe基因型为ε3/ε3。
图1b:psen1p433s突变家系图。
图1c:测序发现图1a家系的先证者携带psen1i249l突变。
图1d:测序发现图1b家系的先证者携带psen1p433s突变。
图2a:核磁共振成像图(mri)。psen1i249l先证者头mri示全脑萎缩和海马萎缩,海马萎缩评分(mta)=1。
图2b:氟代脱氧葡萄糖单光子发射计算机断层成像图(fdgspect)。共成像24张,从左到右、从上至下依次是psen1p433s先证者脑部从上到下的不同水平截面,脑中代谢减低的部位需通过双侧对比、与其他水平的截面对比综合得出。代谢减低的各部位不在同一个水平截面上。psen1p433s先证者头fdgspect示双侧顶叶、颞叶、枕叶葡萄糖代谢普遍减低,左侧额叶葡萄糖代谢较对侧减低;左侧基底节区及丘脑葡萄糖代谢较对侧减低。其中第一行第4个图像箭头示双侧顶叶葡萄糖代谢减低,第一行第7个图像箭头示左侧额叶葡萄糖代谢较对侧减低,第二行第1个图像箭头示双侧颞叶葡萄糖代谢减低,第二行第5个图像箭头示双侧枕叶葡萄糖代谢减低,第二行第6个图像上方箭头和下方箭头分别示左侧基底节区葡萄糖代谢较对侧减低,左侧丘脑葡萄糖代谢较对侧减低。
图3:3种突变型质粒pcr扩增产物的电泳图(1.5%tae琼脂糖凝胶)。其中:
泳道m,trans2kplusdnamarker;
泳道1-5,plvx-psen1p433s-ires-zsgreen1;
泳道6-10,plvx-psen1d385a-ires-zsgreen1;
泳道11-15,plvx-psen1i249l-ires-zsgreen1;
其中泳道1-5的5个样品之间,泳道6-10的5个样品之间,泳道11-15的5个样品之间,除了退火温度不同(分别为45℃/50℃/55℃/60℃/65℃),其它条件均相同。
图4a:i249l突变反向测序,图中第707位t>g。
图4b:p433s突变反向测序,图中第146位g>a。
图4c:d385a突变反向测序,图中第291位t>g。
图5a至图5e:各组慢病毒表达载体转染293t细胞的荧光表达情况。各列分别表示10x和20x物镜下白光、荧光以及白光荧光融合(merge)时的镜下表现,各行分别表示转染后0h、6h、30h、54h的镜下表现。其中:
图5a是空载组(vehicle);
图5b是野生组(wt);
图5c是i249l突变组;图5d是d385a突变组;图5e是p433s突变组。
图6:各组sh-sy5y感染后6d荧光表达情况(20x物镜)。各列分别表示20x物镜下白光、荧光以及白光荧光融合(merge)时的镜下表现,各行分别表示m0-m5各组的镜下表现。m0,空白组(blank);m1,空载组(vehicle);m2,野生组(wt);m3,i249l突变组;m4,d385a突变组;m5,p433s突变组。
图7a至图7e:westernblot检测ps1全长(ps1-fl)和ps1-c端片段(ps1-ctf)。其中:
图7a,westernblot示ps1-fl、ps1-ctf、ps1-ctflongexposure(longexposure是指延长了曝光时间,目的是尽可能显示较淡的ps1-ctf条带,避免因原曝光时间过短导致ps1-ctf条带未显现)、β-actin蛋白表达情况。m0,空白组(blank);m1,空载组(vehicle);m2,野生组(wt);m3,i249l突变组;m4,d385a突变组;m5,p433s突变组。右侧标明了被检测的蛋白的大小(kda)。(b-e)ps1-fl的3次独立重复实验结果(统计方法均为anovabonferroni校正);
图7b,ps1-fl(wt标化为1):p=0.000;m0/1vsm2/3/4/5***p=0.000;
图7c,ps1-ctf(blank标化为1):p=0.000;m0/1/4/5vsm2/3**p<0.01;
图7d,ps1-ctflongexposure(blank标化为1):p=0.000;m0/1/4/5vsm2/3***p=0.000;
图7e,ps1-ctf/(ps1-fl+ps1-ctf)(%)(blank标化为100%):p=0.000;m4/5***vsm0/1/2/3p=0.000。
图8a至图8b,westernblot检测sappβ。其中:
图8a是westernblot示sappβ和β-actin蛋白表达情况。m0,空白组(blank);m1,空载组(vehicle);m2,野生组(wt);m3,i249l突变组;m4,d385a突变组;m5,p433s突变组。右侧标明了被检测的蛋白的大小(kda);
图8b是sappβ的3次独立重复实验结果(统计方法为anovabonferroni校正):p=0.895。
图9a至图9f:elisa法检测aβ40、42、43的3次独立重复实验结果。aβ40、42、43已按冻干浓缩比例修正至原培养液浓度值。统计方法均为anovabonferroni。m0,空白组(blank);m1,空载组(vehicle);m2,野生组(wt);m3,i249l突变组;m4,d385a突变组;m5,p433s突变组。其中,为了便于直观地比较,*、**或***表示不同的组别。例如图9a中,m4用*表示,m0/1/5用***表示。图9b中,**表示m3组,***表示m5组。图9c中,***表示m5组。图9d中,*表示m3组,***表示m4组,**表示m5组。图9e中,***表示m5组。图9f中,*表示m4组,***表示m5组。其中:
图9a,aβ40:p=0.000;m0/1/5***vsm2/3p=0.000;m0/1/5***vsm4*p=0.000;m2/3vsm4*p=0.000;
图9b,aβ42:p=0.000;m0/1vsm3p<0.05;m2/4vsm3**/5***p=0.000;m0/1vsm5***p=0.000;m3**vsm5***p=0.000;
图9c,aβ43:p=0.000;m5***vsm0/1/2/3/4p=0.000;
图9d,aβ42/40:p=0.000;m0/1/2vsm3*p<0.05;m0/1/2vsm4***p=0.000;m0/1/2vsm5**p<0.01;m4***vsm3*p<0.01;m4***vsm5**p<0.05;
图9e,aβ43/40:p=0.000;m5***vsm0/1/2/3/4p=0.000;
图9f,(aβ42+43)/40:p=0.000;m4*vsm0/1/2/3p<0.05;m5***vsm0/1/2/3p=0.000;m4*vsm5***p=0.000。
具体实施方式
下面将结合实施例对本发明的实施方案进行详细描述,但是本领域技术人员将会理解,下列实施例仅用于说明本发明,而不应视为限定本发明的范围。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规产品。
实施例1:阿尔茨海默病致病基因的筛查和测序
1.实验方法
家系采集和遗传学检测:对中国家族性阿尔茨海默病注册登记网(cfan,www.chinacfan.org)两个ad家系采集临床资料和血液标本,筛查psen1、psen2、app基因突变并检测apoe基因型。已获取书面知情同意。
2.实验结果
发现了psen1突变,即两个家系分别携带psen1i249l和p433s新突变,其家系图分别如图1a、和图1b所示,测序结果分别如图1c和图1d所示。
i249l突变家系的先证者是一名58岁家庭主妇,进行性记忆减退4年,发病1年内出现人格改变和社会退缩,病情逐渐加重,近1年出现定向力障碍和重复动作。头mri示进行性全脑萎缩和海马萎缩(图2a)。既往史包括冠心病和发作性眩晕。其母发病年龄(aao)、症状与其相似,晚期出现视听幻觉。先证者的女儿也携带该突变,31岁时诊断白血病。
p433s突变家系累及3代7名患者,其aao和病程相似。先证者39岁男性,进行性记忆力减退5年,诊断mci(轻度认知功能障碍),就诊时发现患者伴有抑郁和易激惹。头fdgspect示双侧顶颞枕叶葡萄糖代谢普遍减低(图2b)。
没有发现psen2、app基因突变。
另外,上述所有个体apoe基因型均为ε3/ε3。
由于apoe基因是ad最主要的风险基因,检测apoe基因型是ad患者的常规检测项目之一,目的是识别apoeε4是否参与该ad家系的发病。上述所有个体apoe基因型均为ε3/ε3,说明从遗传因素角度考虑,apoeε4并未参与这些个体ad的发生,进一步证明这些个体发生ad很可能是由于致病基因psen1突变所致。
实施例2:细胞学实验
1.实验方法
(1)构建慢病毒表达载体、定点突变和鉴定
pcdna3.1-wtpsen1-egfp质粒来自于本实验室前期工作(方伯言.中国人家族性阿尔茨海默病早老素1基因突变病理功能研究[博士]:首都医科大学;2006.)。慢病毒表达载体plvx-ires-zsgreen1(clontech,catalogno.632187)、慢病毒包装质粒pspax2(addgene,catalogno.12260)和包膜质粒pmd2g(addgene,catalogno.12259)由温州医科大学李红智教授馈赠14。
使用pcdna3.1-wtpsen1-egfp质粒(也可以使用插入有psen1其它质粒),通过pcr扩增、纯化pcr产物,回收得到目的片段psen1。也可以用本领域技术人员知悉的其它手段制备目的片段psen1(例如人工合成)。
双酶切环状质粒plvx-ires-zsgreen1,回收线性载体plvx-ires-zsgreen1,双酶切体系:plvx-ires-zsgreen11μg,flycutbamhi1μl,flycutecori1μl,10×flycutbuffer5μl,ddh2otofinalvolume50μl。
使用easypurequickgelextractionkit(code#eg101)纯化回收目的片段psen1和双酶切后的质粒plvx-ires-zsgreen1,详细操作步骤参照说明书进行。
利用peasy-uniseamlesscloningandassemblykit(购于北京全式金生物技术有限公司,货号cu101-01)将psen1和plvx-ires-zsgreen1进行连接,经过stbl3菌落pcr和双酶切鉴定获得阳性克隆后,进行测序;以测序正确的质粒plvx-psen1-ires-zsgreen1为模板分别进行定点突变并测序鉴定。定点突变包括i249l突变、p433s突变和ps1/γ-分泌酶天冬氨酸活性催化位点之一d385a突变这3种突变。得到3种突变型质粒:plvx-psen1i249l-ires-zsgreen1、plvx-psen1p433s-ires-zsgreen1和plvx-psen1d385a-ires-zsgreen1。stbl3感受态菌扩增这3种突变型质粒后对所提质粒进行测序鉴定。
其中,所述定点突变的具体操作如下:
定点突变的引物如下表所示,其中下划线部分表示碱基突变位点:
pcr体系:模板8ng,正向引物(10μm)1μl,反向引物(10μm)1μl,5×transstartfastpfuflybuffer10μl,2.5mmdntps4μl,transstartfastpfuflydnapolymerase1μl,加ddh2o至50μl。
点突变pcr反应条件如下:
各退火温度下条带均清晰,均适合进行下游实验,这里选取退火温度60℃的产物进行后面的实验。
由此构建如下的5种重组慢病毒表达载体:
空载体(vehicle,即plvx-ires-zsgreen1);
野生型(即含有野生型psen1的plvx-psen1-ires-zsgreen1,简称为wt);
3种突变型质粒(即含有突变型psen1的plvx-psen1-ires-zsgreen1):plvx-psen1i249l-ires-zsgreen1、plvx-psen1p433s-ires-zsgreen1和plvx-psen1d385a-ires-zsgreen1。
上面制得的3种突变型质粒分别通过琼脂糖凝胶电泳和测序进行鉴定。
(2)重组慢病毒包装
将vehicle、wt和3种mt的重组慢病毒表达载体分别使用pspax2和pmd2.g质粒通过lipo3000转染293t细胞(购自北京全式金生物技术有限公司),转染后6h换液,换液后24h和48h荧光显微镜观察荧光表达率。换液后24h和48h收集病毒液(转染的293t细胞的上清液),3000rpm离心5min,0.45μm滤膜过滤于4℃,分装后-80℃储存。
293t细胞培养于含10%胎牛血清(fetalbovineserum,fbs)(gibco,grandisland,ny,usa)的dmem(dulbecco’smodifiedeaglemedium)(gibco,grandisland,ny,usa)中,培养液含100iu/ml青霉素和100μg/ml链霉素(gibco)。细胞放于37℃5%co2培养箱中培养。
(3)慢病毒感染sh-sy5y细胞
以人神经母细胞瘤sh-sy5y细胞(购自国家实验细胞资源共享服务平台(北京总部),网址http://www.crcpumc.com/)汇合度为40%-60%时,每1cm2底面积铺30μl前面步骤(2)制备的病毒液,分别感染sh-sy5y细胞,观察荧光表达率。
sh-sy5y细胞培养于含10%胎牛血清(fetalbovineserum,fbs)(gibco,grandisland,ny,usa)的dmem/f12(gibco,grandisland,ny,usa)中,培养液含100iu/ml青霉素和100μg/ml链霉素(gibco)。细胞放于37℃5%co2培养箱中培养。
(4)westernblot方法检测ps1蛋白
感染后第7天收集细胞提取蛋白样品,用westernblot检测ps1蛋白(一抗:ps1抗体,1:8000,abcam公司,货号ab76083;二抗:山羊抗兔hrp,1:8000,abcam公司,货号ab6721)、sappβ蛋白(一抗:sappβ抗体,1:2000,biolegend公司,货号813401;二抗:山羊抗兔hrp,1:8000,abcam公司,货号ab6721)和β-actin蛋白表达水平(一抗β-actin抗体,1:1000,santacruz公司,货号sc-47778;二抗山羊抗小鼠hrp,1:8000,abcam公司,货号ab6789)。
(5)elisa方法检测aβ40、aβ42和aβ43浓度
收集感染后第7-9天之间的细胞培养液离心冻干浓缩后用aβelisa试剂盒检测aβ40、aβ42和aβ43浓度(aβ40elisa试剂盒、aβ42elisa试剂盒和aβ43elisa试剂盒均购自ibl公司,货号分别为jp27718、jp27719、jp27710)。
2.实验结果
(1)质粒的pcr鉴定和测序鉴定
电泳结果显示3种突变质粒的扩增条带均清晰(图3)。
stbl3感受态菌扩增质粒后所提质粒经测序鉴定提示各组载体构建成功(图4a-图4c)。
(2)293t细胞转染质粒后观察荧光表达
各组荧光表达率在转染后30h约为70%,54h约为80%(图5a-图5e),提示慢病毒包装成功。
(3)sh-sy5y细胞感染慢病毒后观察荧光表达
感染后6d各组荧光表达率一致,约为70%(图6),提示慢病毒感染后绿色荧光蛋白在sh-sy5y细胞中成功表达,且组间慢病毒感染效率一致。
共有6组细胞样品:
m0(blank,即只有sh-sy5y细胞,不进行病毒感染)、m1(vehicle)、m2-m5(wt、i249l、d385a、p433s)。
(4)westernblot检测ps1蛋白表达水平
m0(blank)和m1(vehicle)几乎未见ps1-fl,m2-m5(wt、i249l、d385a、p433s)的ps1-fl蛋白表达水平显著高于m0(blank)和m1(vehicle)(图7a、图7b)。
m2(wt)和m3(i249l)的ps1-ctf显著高于m0(blank)、m1(vehicle)、m4(d385a)和m5(p433s);m4(d385a)和m5(p433s)较m0(blank)、m1(vehicle)有降低趋势但无显著统计学差异(图7a、7c、7d)。m4(d385a)和m5(p433s)的ps1-ctf/(ps1-fl+ps1-ctf)(%)显著低于m0(blank)、m1(vehicle)、m2(wt)和m3(i249l)(图7e)。
上述结果说明:
①目的基因突变型psen1已成功导入sh-sy5y细胞并成功表达和经历内切水解作用等内源性加工过程,psen1突变稳转细胞模型成功建立;
②不同突变对ps1-fl的内切水解作用的影响不同:m3(i249l)几乎不影响ps1-fl的内切水解作用。m4/5(d385a、p433s)的存在会部分抑制ps1-fl的内切水解作用,同时还会部分抑制细胞原有内源性正常ps1-fl的内切水解作用,使得具有γ-分泌酶催化活性的ps1-ctf减少。
(5)westernblot检测sappβ蛋白表达水平
各组间sappβ蛋白水平未见显著差异(图8a、8b)。上述结果说明ps1突变不影响sappβ蛋白水平,且sappβ组间未见显著差异,便于后续sappβ剪切产物aβ的组间比较。
(6)elisa检测aβ40、aβ42、aβ43
检测结果如下(图9):
①aβ40(图9a):m0(blank)、m1(vehicle)、m5(p433s)的aβ40显著高于m2(wt),m4(d385a)的aβ40显著低于m2(wt),但m3(i249l)与m2(wt)的aβ40无显著统计学差异。
②aβ42(图9b):m3(i249l)、m5(p433s)的aβ42显著高于m2(wt),且m5(p433s)的aβ42显著高于m3(i249l),但m4(d385a)与m2(wt)无显著统计学差异。
③aβ43(图9c):m0(blank)和m1(vehicle)的aβ43很少,m2(wt)、m3(i249l)、m4(d385a)的aβ43未测出。m5(p433s)的aβ43显著升高。
④aβ42/40(图9d):m3(i249l)、m4(d385a)、m5(p433s)的aβ42/40比值显著高于m2(wt),m4(d385a)的aβ42/40显著高于m3(i249l)和m5(p433s),m5(p433s)较m3(i249l)的aβ42/40有升高趋势但无显著统计学差异。
⑤aβ43/40(图9e):同aβ43,m5(p433s)的aβ43/40显著升高。
⑥(aβ42+43)/aβ40(图9f):m4(d385a)和m5(p433s)的(aβ42+43)/aβ40较m2(wt)显著升高,且m5(p433s)的(aβ42+43)/aβ40显著高于m4(d385a),m3(i249l)的(aβ42+43)/aβ40较m2(wt)有升高趋势但无统计学差异。
根据上述结果,小结各突变对aβ的影响如下:
①i249l突变对aβ40水平无明显影响,使aβ42水平显著升高,aβ42/40比值显著升高,与wt一样几乎不生成aβ43,(aβ42+43)/aβ40有升高趋势。
②d385a使aβ40水平显著降低,对aβ42水平无明显影响,aβ42/40比值显著升高,与wt一样几乎不生成aβ43,(aβ42+43)/aβ40显著升高。
③p433s使aβ40水平显著升高,aβ42水平显著升高,aβ42/40比值显著升高,aβ43水平显著升高,aβ43/40比值升高,(aβ42+43)/aβ40显著升高。
④3种突变相比较,p433s的aβ42显著高于i249l,p433s较i249l的aβ42/40有升高趋势,而d385a突变对aβ42/40比值的升高显著高于i249l和p433s;p433s的(aβ42+43)/aβ40显著高于d385a。
3.结果分析
本研究首次在2个中国fad家系中报道了psen1i249l和p433s新突变,通过慢病毒表达载体体系成功建立了这2种突变的稳转神经细胞模型,对其ps1蛋白表达水平的检测和aβ的检测提示这些突变的致病机制存在不同。
i249l和p433s突变家系的早发和超早发现象可能与这些突变在ps1中处于影响γ-分泌酶正常催化活性的关键位置有关。i249l突变邻近d257天冬氨酸催化位点,其携带者aao及记忆和精神症状与天冬氨酸催化位点附近已报道位点特征基本一致,且该家系中该突变携带者还具有易发生缺血和白血病的特征。p433s突变位于ps1蛋白高度保守的palp区(palp区是指ps1蛋白第433-436个氨基酸,其氨基酸缩写分别为palp,该区域在物种进化过程中高度保守),其超早发和精神症状与该位点附近已报道的a434c和a434t突变一致15,16。palp区对γ-分泌酶催化位点的正常构象至关重要,可能参与底物识别17-19。
aβ42水平和/或aβ42/40比值两者任一都可作为psen1突变致病性的主要衡量标志,本研究结果支持两种新突变(i249l和p433s)以及ps1天冬氨酸活性催化位点之一d385a突变对ad的致病作用。此外,p433s的aβ42绝对值和相对比值都高于i249l,这与临床p433s突变家系发病年龄明显早于i249l突变家系相一致。
由于ps1/γ-分泌酶剪切aβ的两条主要途径是aβ49-aβ46-aβ43-aβ40和aβ48-aβ45-aβ42-aβ38,本研究发现i249l突变aβ42和aβ42/40比值升高,几乎不生成aβ43而aβ40水平正常,且i249l突变不影响具有γ-分泌酶催化活性的ps1-ctf的产量,故提示i249l突变可能对aβ49-aβ46-aβ43-aβ40途径无明显影响,而是可能通过干扰aβ48-aβ45-aβ42-aβ38途径发挥ad致病作用,且这种致ad作用可能是由于突变后的ps1-ctf的γ-分泌酶催化活性异常,而非ps1-ctf总量的减少所致。
本研究发现p433s突变的aβ40、aβ42、aβ43、aβ42/40、aβ43/40和(aβ42+43)/aβ40比值均升高,且p433s突变通过抑制ps1-fl的内切水解作用使具有γ-分泌酶催化活性的ps1-ctf减少,故提示p433s突变可能干扰γ-分泌酶对aβ49-aβ46-aβ43-aβ40途径和aβ48-aβ45-aβ42-aβ38途径的分配、抑制aβ43-aβ40途径并干扰aβ48-aβ45-aβ42-aβ38途径,使aβ42和aβ43的相对产量增加从而发挥ad致病作用,且p433s突变的这种致ad作用的部分原因可能是通过使ps1-ctf总量的减少,使得较长的aβ肽产量增加,从而导致ad。
d385作为ps1/γ-分泌酶天冬氨酸催化位点之一,本研究中d385a突变的aβ40降低,aβ42水平无明显变化,aβ42/40比值升高,几乎不生成aβ43,且d385a突变通过抑制ps1-fl的内切水解作用使具有γ-分泌酶催化活性的ps1-ctf减少,这与既往研究(kretnerb,etal.generationanddepositionofaβ43bythevirtuallyinactivepresenilin-1l435fmutantcontradictsthepresenilinloss-of-functionhypothesisofalzheimer'sdisease.embomolmed.2016may2;8(5):458-65.)发现稳定转染d385a突变的hek293/sw细胞其ps1内切水解作用显著受抑制且几乎不生成aβ的结果相一致。故本研究支持d385a突变可能通过显著抑制γ-分泌酶催化活性和抑制具有γ-分泌酶催化活性的ps1-ctf的生成这两方面抑制aβ49-aβ46-aβ43-aβ40途径,从而引起aβ42/40等与ad相关的aβ指标的升高。
综上,i249l和p433s都能引起aβ42和/或aβ42/40比值的升高,支持这2种fad新突变的致病作用。对ps1内切水解作用的分析和对aβ40、42、43的分析表明,这2种新突变与d385a突变的致病机制存在不同。
参考文献
1.sherringtonr,rogaevei,liangy,etal.cloningofagenebearingmissensemutationsinearly-onsetfamilialalzheimer'sdisease.nature.jun291995;375(6534):754-760.
2.goatea,chartier-harlinmc,mullanm,etal.segregationofamissensemutationintheamyloidprecursorproteingenewithfamilialalzheimer'sdisease.nature.feb211991;349(6311):704-706.
3.rogaevei,sherringtonr,rogaevaea,etal.familialalzheimer'sdiseaseinkindredswithmissensemutationsinageneonchromosome1relatedtothealzheimer'sdiseasetype3gene.nature.aug311995;376(6543):775-778.
4.levy-lahade,wijsmanem,nemense,etal.afamilialalzheimer'sdiseaselocusonchromosome1.science.aug181995;269(5226):970-973.
5.haassc,kaetherc,thinakarang,sisodias.traffickingandproteolyticprocessingofapp.coldspringharborperspectivesinmedicine.may2012;2(5):a006270.
6.haassc.takefive--baceandthegamma-secretasequartetconductalzheimer'samyloidbeta-peptidegeneration.theembojournal.feb112004;23(3):483-488.
7.qi-takaharay,morishima-kawashimam,tanimuray,etal.longerformsofamyloidbetaprotein:implicationsforthemechanismofintramembranecleavagebygamma-secretase.thejournalofneuroscience:theofficialjournalofthesocietyforneuroscience.jan122005;25(2):436-445.
8.takamim,nagashimay,sanoy,etal.gamma-secretase:successivetripeptideandtetrapeptidereleasefromthetransmembranedomainofbeta-carboxylterminalfragment.thejournalofneuroscience:theofficialjournalofthesocietyforneuroscience.oct142009;29(41):13042-13052.
9.chavez-gutierrezl,bammensl,benilovai,etal.themechanismofgamma-secretasedysfunctioninfamilialalzheimerdisease.theembojournal.may162012;31(10):2261-2274.
10.okochim,tagamis,yanagidak,etal.gamma-secretasemodulatorsandpresenilin1mutantsactdifferentlyonpresenilin/gamma-secretasefunctiontocleaveabeta42andabeta43.cellreports.jan312013;3(1):42-51.
11.fernandezma,klutkowskija,frerett,wolfems.alzheimerpresenilin-1mutationsdramaticallyreducetrimmingoflongamyloidbeta-peptides(abeta)bygamma-secretasetoincrease42-to-40-residueabeta.thejournalofbiologicalchemistry.nov072014;289(45):31043-31052.
12.tomitat,watabikit,takikawar,etal.thefirstprolineofpalpmotifatthecterminusofpresenilinsisobligatoryforstabilization,complexformation,andgamma-secretaseactivitiesofpresenilins.thejournalofbiologicalchemistry.aug312001;276(35):33273-33281.
13.wolfems,xiaw,ostaszewskibl,diehlts,kimberlywt,selkoedj.twotransmembraneaspartatesinpresenilin-1requiredforpresenilinendoproteolysisandgamma-secretaseactivity.nature.apr081999;398(6727):513-517.
14.yuanw,chenj,caoy,etal.15.comparativeanalysisandoptimizationofprotocolsforproducingrecombinantlentiviruscarryingtheanti-her2chimericantigenreceptorgene.jgenemed.2018jul;20(7-8):e3027.
15.devig,fotioua,jyrinjid,tyckob,dearmands,rogaevae,songyq,medierosh,liangy,orlacchioa,williamsonj,stgeorge-hyslopp,mayeuxr.novelpresenilin1mutationsassociatedwithearlyonsetofdementiainafamilywithbothearly-onsetandlate-onsetalzheimerdisease.archneurol.2000.
16.jiaob,tangb,liux,xuj,wangy,zhoul,zhangf,yanx,zhouy,shenl.mutationalanalysisinearly-onsetfamilialalzheimer'sdiseaseinmainlandchina.neurobiolaging.2014.
17.wangj,beherd,nyborgac,shearmanms,goldete,goatea.c-terminalpalmotifofpresenilinandpresenilinhomologuesrequiredfornormalactivesiteconformation.journalofneurochemistry.2006.
18.satoc,takagis,tomitat,iwatsubot.thec-terminalpalmotifandtransmembranedomain9ofpresenilin1areinvolvedintheformationofthecatalyticporeofthegamma-secretase.thejournalofneuroscience:theofficialjournalofthesocietyforneuroscience.2008.
19.baixc,yanc,yangg,etal.anatomicstructureofhumangamma-secretase.nature.2015.
尽管本发明的具体实施方式已经得到详细的描述,本领域技术人员将会理解。根据已经公开的所有教导,可以对那些细节进行各种修改和替换,这些改变均在本发明的保护范围之内。本发明的全部范围由所附权利要求及其任何等同物给出。
sequencelisting
<110>首都医科大学宣武医院
<120>阿尔茨海默病新突变、其稳转细胞模型及医药用途
<130>idc180343
<160>17
<170>patentinversion3.2
<210>1
<211>467
<212>prt
<213>artificial
<220>
<223>psen1i249l的氨基酸序列
<400>1
metthrgluleuproalaproleusertyrpheglnasnalaglnmet
151015
sergluaspasnhisleuserasnthrvalargserglnasnaspasn
202530
arggluargglngluhisasnaspargargserleuglyhisproglu
354045
proleuserasnglyargproglnglyasnserargglnvalvalglu
505560
glnaspgluglugluaspglugluleuthrleulystyrglyalalys
65707580
hisvalilemetleuphevalprovalthrleucysmetvalvalval
859095
valalathrilelysservalserphetyrthrarglysaspglygln
100105110
leuiletyrthrprophethrgluaspthrgluthrvalglyglnarg
115120125
alaleuhisserileleuasnalaalailemetileservalileval
130135140
valmetthrileleuleuvalvalleutyrlystyrargcystyrlys
145150155160
valilehisalatrpleuileileserserleuleuleuleuphephe
165170175
pheserpheiletyrleuglygluvalphelysthrtyrasnvalala
180185190
valasptyrilethrvalalaleuleuiletrpasnpheglyvalval
195200205
glymetileserilehistrplysglyproleuargleuglnglnala
210215220
tyrleuilemetileseralaleumetalaleuvalpheilelystyr
225230235240
leuproglutrpthralatrpleuleuleualavalileservaltyr
245250255
aspleuvalalavalleucysprolysglyproleuargmetleuval
260265270
gluthralaglngluargasngluthrleupheproalaleuiletyr
275280285
serserthrmetvaltrpleuvalasnmetalagluglyaspproglu
290295300
alaglnargargvalserlysasnserlystyrasnalagluserthr
305310315320
gluarggluserglnaspthrvalalagluasnaspaspglyglyphe
325330335
sergluglutrpglualaglnargaspserhisleuglyprohisarg
340345350
serthrprogluserargalaalavalglngluleuserserserile
355360365
leualaglygluaspproglugluargglyvallysleuglyleugly
370375380
asppheilephetyrservalleuvalglylysalaseralathrala
385390395400
serglyasptrpasnthrthrilealacysphevalalaileleuile
405410415
glyleucysleuthrleuleuleuleualailephelyslysalaleu
420425430
proalaleuproileserilethrpheglyleuvalphetyrpheala
435440445
thrasptyrleuvalglnprophemetaspglnleualaphehisgln
450455460
phetyrile
465
<210>2
<211>467
<212>prt
<213>artificial
<220>
<223>psen1p433s的氨基酸序列
<400>2
metthrgluleuproalaproleusertyrpheglnasnalaglnmet
151015
sergluaspasnhisleuserasnthrvalargserglnasnaspasn
202530
arggluargglngluhisasnaspargargserleuglyhisproglu
354045
proleuserasnglyargproglnglyasnserargglnvalvalglu
505560
glnaspgluglugluaspglugluleuthrleulystyrglyalalys
65707580
hisvalilemetleuphevalprovalthrleucysmetvalvalval
859095
valalathrilelysservalserphetyrthrarglysaspglygln
100105110
leuiletyrthrprophethrgluaspthrgluthrvalglyglnarg
115120125
alaleuhisserileleuasnalaalailemetileservalileval
130135140
valmetthrileleuleuvalvalleutyrlystyrargcystyrlys
145150155160
valilehisalatrpleuileileserserleuleuleuleuphephe
165170175
pheserpheiletyrleuglygluvalphelysthrtyrasnvalala
180185190
valasptyrilethrvalalaleuleuiletrpasnpheglyvalval
195200205
glymetileserilehistrplysglyproleuargleuglnglnala
210215220
tyrleuilemetileseralaleumetalaleuvalpheilelystyr
225230235240
leuproglutrpthralatrpleuileleualavalileservaltyr
245250255
aspleuvalalavalleucysprolysglyproleuargmetleuval
260265270
gluthralaglngluargasngluthrleupheproalaleuiletyr
275280285
serserthrmetvaltrpleuvalasnmetalagluglyaspproglu
290295300
alaglnargargvalserlysasnserlystyrasnalagluserthr
305310315320
gluarggluserglnaspthrvalalagluasnaspaspglyglyphe
325330335
sergluglutrpglualaglnargaspserhisleuglyprohisarg
340345350
serthrprogluserargalaalavalglngluleuserserserile
355360365
leualaglygluaspproglugluargglyvallysleuglyleugly
370375380
asppheilephetyrservalleuvalglylysalaseralathrala
385390395400
serglyasptrpasnthrthrilealacysphevalalaileleuile
405410415
glyleucysleuthrleuleuleuleualailephelyslysalaleu
420425430
seralaleuproileserilethrpheglyleuvalphetyrpheala
435440445
thrasptyrleuvalglnprophemetaspglnleualaphehisgln
450455460
phetyrile
465
<210>3
<211>1404
<212>dna
<213>artificial
<220>
<223>编码psen1i249l的核苷酸序列
<400>3
atgacagagttacctgcaccgttgtcctacttccagaatgcacagatgtctgaggacaac60
cacctgagcaatactgtacgtagccagaatgacaatagagaacggcaggagcacaacgac120
agacggagccttggccaccctgagccattatctaatggacgaccccagggtaactcccgg180
caggtggtggagcaagatgaggaagaagatgaggagctgacattgaaatatggcgccaag240
catgtgatcatgctctttgtccctgtgactctctgcatggtggtggtcgtggctaccatt300
aagtcagtcagcttttatacccggaaggatgggcagctaatctataccccattcacagaa360
gataccgagactgtgggccagagagccctgcactcaattctgaatgctgccatcatgatc420
agtgtcattgttgtcatgactatcctcctggtggttctgtataaatacaggtgctataag480
gtcatccatgcctggcttattatatcatctctattgttgctgttctttttttcattcatt540
tacttgggggaagtgtttaaaacctataacgttgctgtggactacattactgttgcactc600
ctgatctggaattttggtgtggtgggaatgatttccattcactggaaaggtccacttcga660
ctccagcaggcatatctcattatgattagtgccctcatggccctggtgtttatcaagtac720
ctccctgaatggactgcgtggctcctcttggctgtgatttcagtatatgatttagtggct780
gttttgtgtccgaaaggtccacttcgtatgctggttgaaacagctcaggagagaaatgaa840
acgctttttccagctctcatttactcctcaacaatggtgtggttggtgaatatggcagaa900
ggagacccggaagctcaaaggagagtatccaaaaattccaagtataatgcagaaagcaca960
gaaagggagtcacaagacactgttgcagagaatgatgatggcgggttcagtgaggaatgg1020
gaagcccagagggacagtcatctagggcctcatcgctctacacctgagtcacgagctgct1080
gtccaggaactttccagcagtatcctcgctggtgaagacccagaggaaaggggagtaaaa1140
cttggattgggagatttcattttctacagtgttctggttggtaaagcctcagcaacagcc1200
agtggagactggaacacaaccatagcctgtttcgtagccatattaattggtttgtgcctt1260
acattattactccttgccattttcaagaaagcattgccagctcttccaatctccatcacc1320
tttgggcttgttttctactttgccacagattatcttgtacagccttttatggaccaatta1380
gcattccatcaattttatatctag1404
<210>4
<211>1404
<212>dna
<213>artificial
<220>
<223>编码psen1p433s的核苷酸序列
<400>4
atgacagagttacctgcaccgttgtcctacttccagaatgcacagatgtctgaggacaac60
cacctgagcaatactgtacgtagccagaatgacaatagagaacggcaggagcacaacgac120
agacggagccttggccaccctgagccattatctaatggacgaccccagggtaactcccgg180
caggtggtggagcaagatgaggaagaagatgaggagctgacattgaaatatggcgccaag240
catgtgatcatgctctttgtccctgtgactctctgcatggtggtggtcgtggctaccatt300
aagtcagtcagcttttatacccggaaggatgggcagctaatctataccccattcacagaa360
gataccgagactgtgggccagagagccctgcactcaattctgaatgctgccatcatgatc420
agtgtcattgttgtcatgactatcctcctggtggttctgtataaatacaggtgctataag480
gtcatccatgcctggcttattatatcatctctattgttgctgttctttttttcattcatt540
tacttgggggaagtgtttaaaacctataacgttgctgtggactacattactgttgcactc600
ctgatctggaattttggtgtggtgggaatgatttccattcactggaaaggtccacttcga660
ctccagcaggcatatctcattatgattagtgccctcatggccctggtgtttatcaagtac720
ctccctgaatggactgcgtggctcatcttggctgtgatttcagtatatgatttagtggct780
gttttgtgtccgaaaggtccacttcgtatgctggttgaaacagctcaggagagaaatgaa840
acgctttttccagctctcatttactcctcaacaatggtgtggttggtgaatatggcagaa900
ggagacccggaagctcaaaggagagtatccaaaaattccaagtataatgcagaaagcaca960
gaaagggagtcacaagacactgttgcagagaatgatgatggcgggttcagtgaggaatgg1020
gaagcccagagggacagtcatctagggcctcatcgctctacacctgagtcacgagctgct1080
gtccaggaactttccagcagtatcctcgctggtgaagacccagaggaaaggggagtaaaa1140
cttggattgggagatttcattttctacagtgttctggttggtaaagcctcagcaacagcc1200
agtggagactggaacacaaccatagcctgtttcgtagccatattaattggtttgtgcctt1260
acattattactccttgccattttcaagaaagcattgtcagctcttccaatctccatcacc1320
tttgggcttgttttctactttgccacagattatcttgtacagccttttatggaccaatta1380
gcattccatcaattttatatctag1404
<210>5
<211>467
<212>prt
<213>artificial
<220>
<223>野生型人psen1蛋白的氨基酸序列
<400>5
metthrgluleuproalaproleusertyrpheglnasnalaglnmet
151015
sergluaspasnhisleuserasnthrvalargserglnasnaspasn
202530
arggluargglngluhisasnaspargargserleuglyhisproglu
354045
proleuserasnglyargproglnglyasnserargglnvalvalglu
505560
glnaspgluglugluaspglugluleuthrleulystyrglyalalys
65707580
hisvalilemetleuphevalprovalthrleucysmetvalvalval
859095
valalathrilelysservalserphetyrthrarglysaspglygln
100105110
leuiletyrthrprophethrgluaspthrgluthrvalglyglnarg
115120125
alaleuhisserileleuasnalaalailemetileservalileval
130135140
valmetthrileleuleuvalvalleutyrlystyrargcystyrlys
145150155160
valilehisalatrpleuileileserserleuleuleuleuphephe
165170175
pheserpheiletyrleuglygluvalphelysthrtyrasnvalala
180185190
valasptyrilethrvalalaleuleuiletrpasnpheglyvalval
195200205
glymetileserilehistrplysglyproleuargleuglnglnala
210215220
tyrleuilemetileseralaleumetalaleuvalpheilelystyr
225230235240
leuproglutrpthralatrpleuileleualavalileservaltyr
245250255
aspleuvalalavalleucysprolysglyproleuargmetleuval
260265270
gluthralaglngluargasngluthrleupheproalaleuiletyr
275280285
serserthrmetvaltrpleuvalasnmetalagluglyaspproglu
290295300
alaglnargargvalserlysasnserlystyrasnalagluserthr
305310315320
gluarggluserglnaspthrvalalagluasnaspaspglyglyphe
325330335
sergluglutrpglualaglnargaspserhisleuglyprohisarg
340345350
serthrprogluserargalaalavalglngluleuserserserile
355360365
leualaglygluaspproglugluargglyvallysleuglyleugly
370375380
asppheilephetyrservalleuvalglylysalaseralathrala
385390395400
serglyasptrpasnthrthrilealacysphevalalaileleuile
405410415
glyleucysleuthrleuleuleuleualailephelyslysalaleu
420425430
proalaleuproileserilethrpheglyleuvalphetyrpheala
435440445
thrasptyrleuvalglnprophemetaspglnleualaphehisgln
450455460
phetyrile
465
<210>6
<211>1404
<212>dna
<213>artificial
<220>
<223>野生型人psen1基因的读码框序列
<400>6
atgacagagttacctgcaccgttgtcctacttccagaatgcacagatgtctgaggacaac60
cacctgagcaatactgtacgtagccagaatgacaatagagaacggcaggagcacaacgac120
agacggagccttggccaccctgagccattatctaatggacgaccccagggtaactcccgg180
caggtggtggagcaagatgaggaagaagatgaggagctgacattgaaatatggcgccaag240
catgtgatcatgctctttgtccctgtgactctctgcatggtggtggtcgtggctaccatt300
aagtcagtcagcttttatacccggaaggatgggcagctaatctataccccattcacagaa360
gataccgagactgtgggccagagagccctgcactcaattctgaatgctgccatcatgatc420
agtgtcattgttgtcatgactatcctcctggtggttctgtataaatacaggtgctataag480
gtcatccatgcctggcttattatatcatctctattgttgctgttctttttttcattcatt540
tacttgggggaagtgtttaaaacctataacgttgctgtggactacattactgttgcactc600
ctgatctggaattttggtgtggtgggaatgatttccattcactggaaaggtccacttcga660
ctccagcaggcatatctcattatgattagtgccctcatggccctggtgtttatcaagtac720
ctccctgaatggactgcgtggctcatcttggctgtgatttcagtatatgatttagtggct780
gttttgtgtccgaaaggtccacttcgtatgctggttgaaacagctcaggagagaaatgaa840
acgctttttccagctctcatttactcctcaacaatggtgtggttggtgaatatggcagaa900
ggagacccggaagctcaaaggagagtatccaaaaattccaagtataatgcagaaagcaca960
gaaagggagtcacaagacactgttgcagagaatgatgatggcgggttcagtgaggaatgg1020
gaagcccagagggacagtcatctagggcctcatcgctctacacctgagtcacgagctgct1080
gtccaggaactttccagcagtatcctcgctggtgaagacccagaggaaaggggagtaaaa1140
cttggattgggagatttcattttctacagtgttctggttggtaaagcctcagcaacagcc1200
agtggagactggaacacaaccatagcctgtttcgtagccatattaattggtttgtgcctt1260
acattattactccttgccattttcaagaaagcattgccagctcttccaatctccatcacc1320
tttgggcttgttttctactttgccacagattatcttgtacagccttttatggaccaatta1380
gcattccatcaattttatatctag1404
<210>7
<211>40
<212>prt
<213>artificial
<220>
<223>aβ40的氨基酸序列
<400>7
aspalagluphearghisaspserglytyrgluvalhishisglnlys
151015
leuvalphephealagluaspvalglyserasnlysglyalaileile
202530
glyleumetvalglyglyvalval
3540
<210>8
<211>42
<212>prt
<213>artificial
<220>
<223>aβ42的氨基酸序列
<400>8
aspalagluphearghisaspserglytyrgluvalhishisglnlys
151015
leuvalphephealagluaspvalglyserasnlysglyalaileile
202530
glyleumetvalglyglyvalvalileala
3540
<210>9
<211>43
<212>prt
<213>artificial
<220>
<223>aβ43的氨基酸序列
<400>9
aspalagluphearghisaspserglytyrgluvalhishisglnlys
151015
leuvalphephealagluaspvalglyserasnlysglyalaileile
202530
glyleumetvalglyglyvalvalilealathr
3540
<210>10
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>10
tggactgcgtggctcctcttggctgtgattt31
<210>11
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>11
ggagccacgcagtccattcagggaggtactt31
<210>12
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>12
ttcaagaaagcattgtcagctcttccaatct31
<210>13
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>13
acaatgctttcttgaaaatggcaaggagtaa31
<210>14
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>14
ttcaagaaagcattggcagctcttccaatct31
<210>15
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>15
ccaatgctttcttgaaaatggcaaggagtaa31
<210>16
<211>33
<212>dna
<213>artificial
<220>
<223>引物
<400>16
aacttggattgggagctttcattttctacagtg33
<210>17
<211>31
<212>dna
<213>artificial
<220>
<223>引物
<400>17
gctcccaatccaagttttactcccctttcct31
- 还没有人留言评论。精彩留言会获得点赞!