一种直链麦芽五糖生成酶在枯草芽孢杆菌中分泌表达的方法与流程
本发明涉及一种直链麦芽五糖生成酶在枯草芽孢杆菌中分泌表达的方法,属于酶工程技术领域。
背景技术:
麦芽五糖(g5)是以α-d-吡喃葡萄糖为单元通过α-1,4-糖苷键连接而成的小分子,是直链麦芽低聚糖的一种。g5具有良好的食品加工适应性:可作为口感柔和的甜味剂;在烘焙、膨化食品中可作为水分调节剂;在冷饮中可改善冷饮的抗融性能、提高冰淇淋的膨胀率;在巧克力、果酱等食品中可有效抑制晶体结晶;在流质食品中可作为增稠剂;此外还能抑制淀粉老化和速冻食品中的蛋白质变性,从而延长保质期。
g5还具有以下独特的生理功效:其在小肠被消化吸收,引起的血糖反应比葡萄糖平稳,能为人体缓慢而持续地供能,可用于运动员的能量补充或胰脏切除病人、肾病患者的膳食治疗;其不易被细菌发酵利用,有利于预防龋齿;其能选择性抑制肠道腐败菌的生长而促进益生菌增殖,从而维护肠道健康;此外还可以促使人体对钙的吸收,有效预防中老年人骨质疏松。因此,g5在运动员食品、婴幼儿食品和保健食品中均有很好的应用前景。
g5的工业化生产与其他直链麦芽低聚糖的生产方式相似,是通过直链麦芽低聚糖生成酶水解淀粉α-1,4-糖苷键切下多个葡萄糖单元而成。现有的生产直链麦芽低聚糖的方法被日本垄断,通常需要α-淀粉酶先进行淀粉液化再由直链麦芽低聚生成酶进行糖化,并添加钙离子以改善酶的稳定性,而国内尚未实现直链麦芽低聚糖的工业化生产。
在目前的国内外生产直链麦芽低聚糖的研究报道中,所用的底物浓度均较低(1~5%),转化率水平一般在50%~75%左右;在对于g5-淀粉酶的已有研究中,产物中的g5含量只有定性分析而未做定量说明,不可直接扩大并应用于工业化生产。随着直链麦芽低聚糖在各个领域中的应用前景越来越广泛,亟需在国内开发具有工业应用价值的高纯度单一直链麦芽低聚糖。
g5-淀粉酶作为生产g5的关键酶,开发g5-淀粉酶在枯草芽孢杆菌中高效分泌表达的方法可以提高我国g5的生产水平,降低应用成本,而且对于其他直链麦芽低聚糖的生产具有指导及借鉴意义,并对于玉米淀粉的的综合高效利用具有重要意义。
技术实现要素:
本发明所要解决的技术问题在于g5-淀粉酶在大肠杆菌(escherichiacoli,e.coli)中的表达效率低,而且e.coli表达系统存在致病性,不适用于生产食品级g5-淀粉酶。针对这一问题提出一种g5-淀粉酶在枯草芽孢杆菌中分泌表达的方法,通过来源于bacillusstearothermophilus(b.stearothermophilus)的可生产g5的g5-淀粉酶编码基因在枯草芽孢杆菌中进行克隆表达,来建立一种g5-淀粉酶在枯草芽孢杆菌中高效分泌表达的方法。
本发明的第一个目的是提供一种重组枯草芽孢杆菌,是以枯草芽孢杆菌中为宿主,表达直链麦芽五糖生成酶;所述直链麦芽五糖生成酶含seqidno.1所示的氨基酸序列。
在本发明的一种实施方式中,编码直链麦芽五糖生成酶的基因的核苷酸序列如seqidno.2所示。
在本发明的一种实施方式中,以枯草芽孢杆菌wb600为表达宿主。
在本发明的一种实施方式中,以pst为表达载体。
本发明的第二个目的是提供一种构建上述重组枯草芽孢杆菌的方法,其特征在于,所述方法包括:
(1)以bacillusstearothermophilus基因组dna为模板,通过pcr扩增获得两端分别含ncoⅰ和xhoⅰ限制性酶切位点的g5目的基因片段;
(2)将(1)中获得的g5基因片段插入pmd18-tsimple质粒中,获得克隆载体pmd18-tsimple/g51;
(3)利用定点突变去除pmd18-tsimple/g51中目的基因内部的ncoⅰ位点,获得载体pmd18-tsimple/g52;
(4)载体pmd18-tsimple/g52经ncoⅰ和xhoⅰ双酶切后,回收含有粘性末端的目的g5基因片段并插入pst质粒中,获得pst/g5质粒,将表达载体pst/g5转化宿主枯草芽孢杆菌wb600,获得基因工程菌。
本发明的第三个目的是提供一种生产直链麦芽五糖生成酶的方法,应用上述重组枯草芽孢杆菌进行发酵。
在本发明的一种实施方式中,所用发酵培养基为:酵母粉30-36g/l,玉米淀粉5-7g/l,kh2po42.0-2.5g/l,k2hpo412-13g/l,ph为7.0~8.0。
在本发明的一种实施方式中,所用发酵培养基为:酵母粉30g/l,玉米淀粉6g/l,kh2po42.3g/l,k2hpo412.5g/l,ph7.5。
在本发明的一种实施方式中,发酵前需要经过种子活化,种子活化包括:将上述重组枯草芽孢杆菌接种到lb培养基中,于35-38℃,200-220r/min培养10-12h。
在本发明的一种实施方式中,将活化后的种子液以1%-5%的接种量转接至发酵培养基中,于35-38℃,200-220r/min培养48-96h。
本发明的第四个目的是提供上述重组枯草芽孢杆菌在制备麦芽五糖或含麦芽五糖的产品中的应用。
本发明的有益效果:
本发明通过构建g5-淀粉酶的枯草芽孢杆菌分泌表达系统,并对基因工程菌的摇瓶发酵条件进行优化,最终确定较佳培养基组成为:酵母粉30g/l,玉米淀粉6g/l,kh2po42.3g/l,k2hpo412.5g/l,ph7.5。在发酵温度37℃,200r/min下振荡培养48h后酶活达451.1u/ml,比初始培养条件下的酶活水平提高了127.5%,比文献报道水平提高了10.9倍。本发明实现了来源于b.stearothermophilus的直链麦芽五糖生成酶在bacillussubtiliswb600,即b.subtiliswb600中的高效分泌表达,有利于打破国外垄断,提高我国g5生产水平,降低应用成本,而且对于其他直链麦芽低聚糖的生产具有指导及借鉴意义,并对于玉米淀粉的的综合高效利用具有重要意义。
附图说明
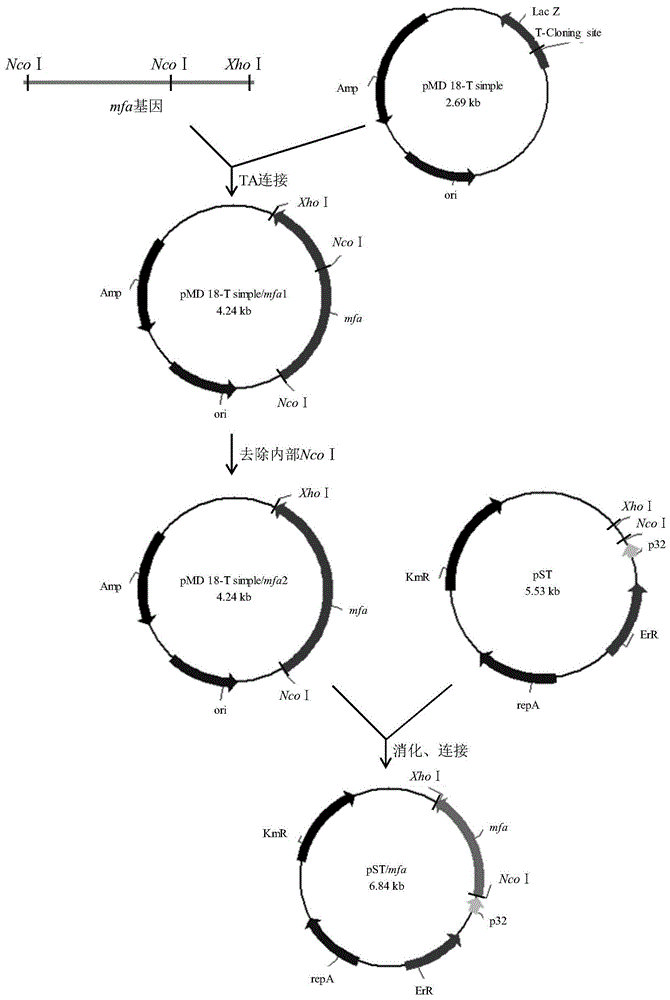
图1为表达载体pst/g5的构建流程。
图2为lb培养基摇瓶发酵b.subtiliswb600(pst/g5)的生长曲线。
图3为发酵出发培养基类型对g5-淀粉酶摇瓶发酵的影响。其中,1:lb;2:tsb;3:tb;4:初始发酵培养基;5:m9。每个值为3次独立实验的平均值,柱状图上不同字母表示存在显著性差异(p<0.05)。
图4为发酵温度和时间对g5-淀粉酶摇瓶发酵的影响。
图5为发酵初始ph对g5-淀粉酶摇瓶发酵的影响。
图6为不同碳源对基因工程菌发酵产酶的影响。其中,0:未添加;1:甘油;2:马铃薯淀粉;3:玉米淀粉;4:木薯淀粉;5:麦芽糊精;6:葡萄糖;7:麦芽糖;8:蔗糖。每个值为三次独立实验的平均值,柱状图上不同字母表示存在显著性差异(p<0.05)。
图7为玉米淀粉浓度对g5-淀粉酶酶摇瓶发酵的影响。其中,每个值为三次独立实验的平均值,柱状图上不同字母表示存在显著性差异(p<0.05)。
图8为无机氮源对g5-淀粉酶摇瓶发酵的影响。每个值为三次独立实验的平均值,柱状图上不同字母表示存在显著性差异(p<0.05)。
图9为有机氮源对g5-淀粉酶摇瓶发酵的影响。其中,1:未添加;2:胰蛋白胨;3:大豆蛋白胨;4:酵母粉;5:酵母膏;6:鱼粉蛋白胨;7:牛肉膏。每个值为三次独立实验的平均值,柱状图上不同字母表示存在显著性差异(p<0.05)。
图10为酵母粉浓度对g5-淀粉酶摇瓶发酵的影响。其中,每个值为三次独立实验的平均值,柱状图上不同字母表示存在显著性差异(p<0.05)。
具体实施方式
(1)dna的提取、验证和回收
b.stearothermophilus基因组dna的提取方法、采用琼脂糖凝胶电泳对提取的质粒dna进行验证的方法参见江南大学李兆丰博士学位论文(李兆丰.软化类芽孢杆菌α-环糊精葡萄糖基转移酶在大肠杆菌中的表达及其产物特异性分析[d].江南大学,2009)。
采用上海生工生物工程公司的质粒小量提取试剂盒(货号:b518191)提取质粒dna,具体提取方法参见说明书。
采用上海碧云天生物技术研究所的胶回收试剂盒(货号:d0056)从琼脂糖凝胶中回收目的基因片段,具体操作方法参见说明书。
(2)感受态细胞的制备和转化
采用上海生工生物工程公司的超级感受态制备试剂盒(货号:b529303)制备e.coli感受态细胞,方法参见说明书,感受态细胞的转化方法参见江南大学李兆丰博士学位论文。
b.subtilis感受态细胞的制备及转化方法参见江南大学张佳瑜硕士学位论文(张佳瑜.软化芽孢杆菌α-环糊精葡萄糖基转移酶在毕赤酵母和枯草杆菌中的表达[d].2010)。
(3)g5-淀粉酶基因的克隆
以b.stearothermophilus基因组dna作为模板,根据g5-淀粉酶的序列分别设计含ncoⅰ和xhoⅰ限制性酶切位点的两端引物(由苏州金唯智生物科技有限公司合成),经primestarhsdna聚合酶扩增后得到g5基因。
pcr操作方法参见江南大学李兆丰博士学位论文。
正反引物设计如下:
正向:5’-catgccatggccgcaccgtttaac-3’(下划线表示ncoⅰ酶切位点)
反向:5’-ccgctcgagaggccatgccaccaa-3’(下划线表示xhoⅰ酶切位点)
将g5基因克隆到pmd18-tsimple质粒中后得到克隆载体,转化e.colijm109,筛选得到阳性菌株后进一步培养,提取质粒pmd18-tsimple/g51,并进行酶切电泳和测序验证,其中测序由苏州金唯智生物科技有限公司完成。
(4)ncoⅰ位点的突变
以质粒pmd18-tsimple/g51为模板,设计互补引物(由苏州金唯智生物科技有限公司合成),在保证氨基酸种类不变的前提下对g5基因中的ncoⅰ位点进行一步突变。
pcr操作方法参见江南大学李兆丰博士学位论文。
引物设计如下:
正向:5’-atgggtcgacccgtggttcaaacc-3’(下划线碱基表示突变碱基)
反向:5’-ggtttgaaccacgggtcgacccat-3’(下划线碱基表示突变碱基)
突变产物经dpnⅰ处理后转化e.colijm109,筛选并培养阳性菌株后提取质粒pmd18-tsimple/g52并验证。
(5)表达载体pst/g5的构建和转化
质粒pmd18-tsimple/g52经ncoⅰ和xhoⅰ双酶切后,回收含有粘性末端的目的基因片段,并用t4dna连接酶将其与经相同限制性核酸内切酶和碱性磷酸酶处理过的pst质粒片段连接,转化e.colijm109后涂布到含有5μg/ml卡那霉素的lb琼脂平板上,于37℃培养10~12h,挑取单菌落在含相同抗性的lb培养基中培养10~12h。提取质粒并经测序验证,获得含正确g5基因序列的表达载体pst/g5,再转化宿主枯草芽孢杆菌b.subtiliswb600,获得可表达g5基因的基因工程菌b.subtiliswb600(pst/g5)。
(6)种子活化
在装有50mllb培养基的250ml锥形瓶中加入终浓度为5μg/ml的卡那霉素,然后移取100μl保藏于-80℃的甘油管菌株b.subtiliswb600(pst/g5),接至瓶中,于37℃摇床中培养10~12h(转速为200r/min)。
(7)发酵培养
在装有50ml发酵培养基的250ml锥形瓶中加入终浓度为5μg/ml的卡那霉素,然后移取2ml活化后的种子液(od600为1.2左右)转接至瓶中,于37℃摇床中培养48-96h(转速为200r/min)。停止发酵后,在4℃、10000×g条件下离心发酵液15min,收集上清液即得到g5-淀粉酶的粗酶液。
(8)酶活的定义及检测方法
酶活的定义:以每分钟生成1μmol还原糖(以葡萄糖计)所需的酶量定义为1个酶活单位(u)。
酶活的检测方法:重组酶活力的测定采用3,5-二硝基水杨酸(dns)法。以c6h8o7-na2hpo4缓冲液(10mm,ph5.5)配制1%(w/v)可溶性淀粉溶液作为底物,在1.98ml底物中加入20μl稀释后的酶液,60℃下反应15min,加入2.0mldns溶液终止反应,沸水浴中显色5min后立即冰浴冷却,于540nm下测定吸光值,根据葡萄糖标准曲线计算出体系中还原糖含量。
实施例1
表达载体pst/g5的构建流程如图1所示。以b.stearothermophilus基因组dna为模板,通过pcr扩增获得两端分别含ncoⅰ和xhoⅰ限制性酶切位点的目的g5基因片段,并将其插入pmd18-tsimple质粒中,获得克隆载体pmd18-tsimple/g51,以便改善亚克隆效果。经分析发现,在目的基因内部大约1029bp处也存在一个相同的ncoⅰ位点,为了更精准地进行双酶切,利用定点突变去除该处酶切位点,获得载体pmd18-tsimple/g52,并经酶切和测序验证。
载体pmd18-tsimple/g52经ncoⅰ和xhoⅰ双酶切后,回收含有粘性末端的目的g5基因片段并插入pst质粒中,获得pst/g5质粒,将经测序验证的分泌型表达载体pst/g5转化宿主b.subtiliswb600,获得基因工程菌b.subtiliswb600(pst/g5)。
实施例2
在装有50mllb培养基的250ml摇瓶中接入适量基因工程菌b.subtiliswb600(pst/g5),在37℃、200r/min条件下培养不同时间取样,测定菌体浓度,结果如图2所示。可见0~6h为延滞期,6~14h为对数生长期,14h以后进入平稳期。对数生长期的菌体适应性和繁殖能力较强,拥有较高的活性,可作为种子液用于发酵产酶。
实施例3
将经lb培养基活化的b.subtiliswb600(pst/g5)种子液分别接入五种较典型的b.subtilis培养基(lb、tsb、m9、tb、初始发酵培养基)中培养24h,结果如图3所示。从酶活水平来看较佳发酵培养基为tb培养基,37℃下发酵24h酶活可达198.3u/ml,显著优于其它培养基;而用m9培养基培养基因工程菌则效果较差,相同条件下酶活仅为16.9u/ml。
实施例4
将活化后的种子液转接至tb培养基中,分别于25℃、30℃、37℃摇床中振荡培养,按发酵时间定点取样测定酶活力,结果如图4所示。由图可见,37℃条件发酵粗酶液的酶活明显高于30℃和25℃条件下的酶活,发酵48h后酶活达338.0u/ml,是相同时间下30℃摇床中发酵酶活的2.6倍,25℃摇床中发酵酶活的4.5倍;发酵48~96h,酶活增长速度放缓,渐趋稳定。
实施例5
在发酵过程中,细菌的生命活动受ph的影响,而其代谢产物会又导致培养基ph的变化,因此不易控制摇瓶发酵过程中的ph,只能优化发酵初始时培养基的ph。将活化后的种子液分别接至不同ph的tb培养基中,37℃下发酵48h后测定酶活力,结果如图5所示。结果表明该基因工程菌在中性偏碱(ph7.0~8.0)的条件下发酵产酶水平较高,较佳发酵初始ph为7.5,相应酶活可达353.6u/ml。当发酵初始ph呈酸性或碱性较大时,则酶活显著降低。
实施例6
在tb培养基的基础上,参照甘油的含碳量,分别以甘油、马铃薯淀粉、玉米淀粉、木薯淀粉、麦芽糊精(de4)、葡萄糖、麦芽糖、蔗糖作为碳源,探究其对基因工程菌发酵产酶水平的影响,结果如图6所示。结果表明以马铃薯淀粉、玉米淀粉、木薯淀粉和麦芽糊精作为碳源时,发酵产酶水平比以甘油为碳源时更高,其中以马铃薯淀粉为碳源时,酶活力可达392.8u/ml,比甘油为碳源提高了23.6%,以玉米淀粉、木薯淀粉和麦芽糊精为碳源则分别提高了16.6%、19.6%和10.6%。由于三种淀粉为碳源时的酶活水平无显著性差异,实际生产中可选择成本相对较低的玉米淀粉为碳源。以蔗糖和麦芽糖为碳源时,酶活力均不如以甘油为碳源时的酶活水平。而葡萄糖则明显抑制了基因工程菌的发酵产酶水平。
实施例7
选择成本相对较低的玉米淀粉作为碳源,进一步对碳源浓度进行优化,结果如图7所示。可见,以玉米淀粉为g5-淀粉酶摇瓶发酵的碳源时,添加量为6g/l对应的酶活较高(402.4u/ml)。
实施例8
用含无机氮源的培养基对基因工程菌进行发酵,从图8可见,在发酵上清液中测得的酶活非常低,当以8g/l尿素为氮源时,酶活相对较高,但也仅达40.7u/ml。而从图9可见有机氮源明显更适合用于基因工程菌的发酵产酶(添加量均为24g/l)。基因工程菌对胰蛋白胨的利用效果相对较差,酶活为210.6u/ml;而酵母粉为较佳氮源,基因工程菌胞外酶活可达405.8u/ml,与相同条件下tb培养基(以玉米淀粉为碳源)的产酶水平(407.6u/ml)相差不大,这也间接说明了tb培养基中胰蛋白胨对基因工程菌的发酵产酶作用不大。
实施例9
氮源浓度也是发酵需要考虑的因素。从图10可见,当以酵母粉为氮源时,较佳氮源浓度为30g/l,相应的酶活可达451.1u/ml,当进一步提高氮源浓度到36g/l时,基因工程菌发酵产酶水平与30g/l时无显著差异。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
sequencelisting
<110>江南大学
<120>一种直链麦芽五糖生成酶在枯草芽孢杆菌中分泌表达的方法
<160>4
<170>patentinversion3.3
<210>1
<211>542
<212>prt
<213>人工序列
<400>1
metlyslyspheleulysserthralaalaleualaleuglyleuser
151015
leuthrpheglyleupheserproalaglnalaalaalapropheasn
202530
glythrmetmetglntyrpheglutrptyrleuproaspaspglythr
354045
leutrpthrlysvalalaasnglualaasnasnleuserserleugly
505560
ilethralaleutrpleuproproalatyrlysglythrserargser
65707580
aspvalglytyrglyvaltyraspleutyraspleuglyglupheasn
859095
glnlysglythrvalargthrlystyrglythrlysalaglntyrleu
100105110
glnalaileglnalaalahisalaalaglymetglnvaltyralaasp
115120125
valvalpheasphislysglyglyalaaspglythrglutrpvalasp
130135140
alavalgluvalasnproseraspargasnglngluileserglythr
145150155160
tyrglnileglnalatrpthrlyspheasppheproglyargglyasn
165170175
thrtyrserserphelystrpargtrptyrhispheaspglyvalasp
180185190
trpaspgluserarglysleuserargiletyrlyspheargglyile
195200205
glylysalatrpasptrpgluvalaspthrgluasnglyasntyrasp
210215220
tyrleumettyralaaspleuaspmetasphisprogluvalvalthr
225230235240
gluleulysasntrpglylystrptyrvalasnthrthrasnileasp
245250255
glypheargleuaspalavallyshisilelyspheserphephepro
260265270
asptrpleusertyrvalargserglnthrglylysproleuphethr
275280285
valglyglutyrtrpsertyraspileasnlysleuhisasntyrile
290295300
thrlysthrasnglythrmetserleupheaspalaproleuhisasn
305310315320
lysphetyrthralaserlysserglyglyalapheaspmetargthr
325330335
leumetthrasnthrleumetlysaspglnprothrleualavalthr
340345350
phevalaspasnhisaspthrgluproglyglnalaleuglnsertrp
355360365
valaspprotrpphelysproleualatyralapheileleuthrarg
370375380
glngluglytyrproglyvalphetyrglyasptyrtyrglyilepro
385390395400
glntyrasnileproserleulysserlysileaspproleuleuile
405410415
alaargargasptyralatyrglythrglnhisasptyrleuasphis
420425430
seraspileileglytrpthrarggluglyvalthrglulysprogly
435440445
serglyleualaalaleuilethraspglyproglyglyserlystrp
450455460
mettyrvalglylysglnhisalaglylysvalphetyraspleuthr
465470475480
glyasnargseraspthrvalthrilethrseraspglytrpglyglu
485490495
phelysvalasnglyglyservalservaltrpvalproarglysthr
500505510
thrvalserthrilethrargproilethrthrargprotrpthrgly
515520525
gluphevalargtrpthrgluproargleuvalalatrppro
530535540
<210>2
<211>1626
<212>dna
<213>人工序列
<400>2
atgaagaaatttctgaaatcgacagctgcgcttgccctgggattatcgctgacgttcggg60
cttttcagccctgcccaggccgccgcaccgtttaacggcaccatgatgcagtattttgaa120
tggtacttgccggatgatggcacgttatggaccaaagtggccaatgaagccaacaactta180
tccagccttggcatcaccgctctttggctgccgcccgcttacaaaggaacaagccgcagc240
gacgtagggtacggagtatacgacttgtatgacctcggcgaattcaatcaaaaagggacc300
gtccgcacaaaatacggaacaaaagctcaatatcttcaagccattcaagccgcccacgcc360
gctggaatgcaagtgtacgccgatgtcgtgttcgaccataaaggcggcgccgacggcacg420
gaatgggtggacgccgtcgaagtcaatccgtccgaccgcaaccaagaaatctcgggcacc480
tatcaaatccaagcatggacgaaatttgattttcccgggcggggcaacacctactccagc540
tttaagtggcgctggtaccattttgacggcgttgattgggacgaaagccgaaaattgagc600
cgcatttacaaattccgcggcatcggcaaagcgtgggattgggaagtagacacggaaaac660
ggaaactatgactacttaatgtatgccgaccttgatatggatcatcccgaagtcgtgacc720
gagctgaaaaactgggggaaatggtatgtcaacacaacgaacattgatgggttccggctt780
gatgccgtcaagcatattaagttcagtttttttcctgattggttgtcgtatgtgcgttct840
cagactggcaagccgctatttaccgtcggggaatattggagctatgacatcaacaagttg900
cacaattacattacgaaaacaaacggaacgatgtctttgtttgatgccccgttacacaac960
aaattttataccgcttccaaatcagggggcgcatttgatatgcgcacgttaatgaccaat1020
actctcatgaaagatcaaccgacattggccgtcaccttcgttgataatcatgacaccgaa1080
cccggccaagcgctgcagtcatgggtcgacccgtggttcaaaccgttggcttacgccttt1140
attctaactcggcaggaaggatacccgggcgtcttttatggtgactattatggcattcca1200
caatataacattccttcgctgaaaagcaaaatcgatccgctcctcatcgcgcgcagggat1260
tatgcttacggaacgcaacatgattatcttgatcactccgacatcatcgggtggacaagg1320
gaaggggtcactgaaaaaccaggatccggactggccgcactgatcaccgatgggccggga1380
ggaagcaaatggatgtacgttggcaaacaacacgccggaaaagtgttctatgaccttacc1440
ggcaaccggagtgacaccgtcaccatcaccagtgatggatggggggaattcaaagtcaat1500
ggcggttcggtttcggtttgggttcctagaaaaacgaccgtttctaccatcactcggccg1560
atcacaacccgaccgtggactggtgaattcgtccgttggaccgaaccacggttggtggca1620
tggcct1626
<210>3
<211>24
<212>dna
<213>人工序列
<400>3
atgggtcgacccgtggttcaaacc24
<210>4
<211>24
<212>dna
<213>人工序列
<400>4
ggtttgaaccacgggtcgacccat24
- 还没有人留言评论。精彩留言会获得点赞!
- 一种高效表达β-甘露聚糖酶的枯草芽孢杆菌及其酶产品和生产方法
- 转芽孢杆菌源α-淀粉酶基因重组乳酸杆菌及制品和应用的制作方法
- 一种以豆饼粉为主要原料生产枯草芽孢杆菌菌剂的方法
- 产酶芽孢杆菌菌株的制作方法
- 枯草芽孢杆菌编码PRPP转酰胺酶突变基因purF及应用的制作方法
- 一株类芽孢杆菌及其用于κ-卡拉胶酶的制备方法
- 一种含枯草芽孢杆菌菌株xp的菌酶联合制剂及其在加速烟草薄片中淀粉降解方面的应用的制作方法
- 枯草芽孢杆菌菌株xp、菌酶联合制剂及其在降解烟草制品中淀粉类化合物方面的应用的制作方法
- 一种含枯草芽孢杆菌菌株xp的菌酶联合制剂及其在加速烟梗中淀粉降解方面的应用的制作方法
- 含抗氧化酶的黄孢原毛平革菌提取液和制备方法