基于线性片段同源重组进行须糖多孢菌基因簇阻断的方法与流程
本发明涉及一种须糖多孢菌进行基因簇阻断的方法,具体涉及一种基于线性片段同源重组进行须糖多孢菌基因簇阻断的方法。
背景技术:
丁烯基多杀菌素是由须糖多孢菌产生的一类具有杀虫活性的大环内酯类化合物,且具有比多杀菌素更强的杀虫活性和杀虫谱,如对多杀菌素难以控制的世界性检疫性害虫和重要农业害虫烟青虫具有很好的生物防治作用,已成为当前最具发展前景的新型生物农药之一。但是,野生型须糖多孢菌合成丁烯基多杀菌素的能力较弱且发酵周期长,制约着丁烯基多杀菌素的大规模工业化生产和应用。因此,如何缩短发酵周期以及提高丁烯基多杀菌素的产量成为当前急需解决的热点问题。
丁烯基多杀菌素的生物合成是采用典型的i型pks(polyketidesynthase,聚酮合酶)途径来完成,通过催化前体物质进行反复的缩合反应,形成聚酮体,随后进行糖基化和甲基化修饰形成最终产物。然而,该过程的进行可能受到其它次生代谢产物基因簇的竞争性抑制,如竞争性利用前体物质。须糖多孢菌全基因组测序结果显示,该菌株基因组存在除丁烯基多杀菌素基因簇外的其它27个次生代谢产物基因簇。这些基因簇的表达很可能使得丁烯基多杀菌素生物合成前体供应减少,最终影响丁烯基多杀菌素生物合成。因此,通过阻断非靶标基因簇来减少对合成前体的利用,有望成为提高丁烯基多杀菌素产量的有效方法。如meng等人在阿维链霉菌中通过对ⅲ型基因簇rpp进行阻断,促使阿维菌素产量由1,024mg/l提高到1,262mg/l(mengl,xiongz,chuj,wangy.enhancedproductionofavermectinbydeletionoftypeiiipkssbiosyntheticclusterrppinstreptomycesavermitilis.lettapplmicrobiol.2016nov;63(5):384-390.doi:10.1111/lam.12635.)。
目前,已建立起多种遗传修饰系统来进行微生物基因组编辑,如crispr/cas9基因组编辑技术(aamirmir,alirezaedraki,jooyounglee,anderikj.sontheimer.typeii-ccrispr-cas9biology,mechanism,andapplication.acschembiol.2017,doi:10.1021/acschembio.7b00855.)、red/et同源重组技术(hailongwang,zhenli,ruonanjia,yuhou,jiayin,xiaoyingbian,aiyingli,rolfmüller,afrancisstewart,junfu,youmingzhang.recetdirectcloningandredαβrecombineeringofbiosyntheticgeneclusters,largeoperonsorsinglegenesforheterologousexpression.natprotoc.2016,11(7):1175-1190.)、cre/loxp位点特异性重组技术(zhengqiangliu,yalixie,xuzhang,xiaofenghu,yushengli,xuezhiding,liqiuxia,shengbiaohu.efficientconstructionoflargegenomicdeletioninagrobacteriumtumefaciensbycombinationofcre/loxpsystemandtriplerecombineering.currmicrobiol.2016,72(4):465-472.)和i-scei内切酶系统(zhongqiuchen,wenling,guangdongshang.recombineeringandi-scei-mediatedpseudomonasputidakt2440scarlessgenedeletion.femsmicrobiologyletters.2016,363(21):fnw231.)等,都适用于微生物基因组大片段敲除。但是,这些遗传操作系统都需要通过质粒介导或者引入外源重组酶,有些还需要进行两次质粒转化才能完成基因组编辑工作,由于须糖多孢菌外源质粒转化极为困难,从而限制着上述遗传修饰系统在须糖多孢菌中的快速应用。
已有研究表明,线性片段介导的同源重组技术同样可用于微生物基因组编辑,如mayumi等人通过线性片段重组成功敲除酵母基因组657.3kb序列(mayumisasaki,hiromichikumagai,kaorutakegawa,hidekitohda.characterizationofgenome-reducedfissionyeaststrains.nucleicacidsresearch.2013,41(10):5382-5399.)。除酵母外,该重组技术在其它微生物如枯草芽胞杆菌也成功运用。
目前,有关须糖多孢菌基因组编辑工作主要停留在单基因水平上,对基因簇大片段阻断的研究还未见有报道,而且外源质粒转化也较为困难。
技术实现要素:
本发明所要解决的技术问题是,克服现有技术存在的上述缺陷,提供一种能对基因簇进行大片段阻断,且无需构建载体、引入外源重组酶的基于线性片段同源重组进行须糖多孢菌基因簇阻断的方法。
本发明解决其技术问题所采用的技术方案如下:一种基于线性片段同源重组进行须糖多孢菌基因簇阻断的方法,包括以下步骤:
(1)在须糖多孢菌基因组中找到ura4作为负筛选标记基因;
(2)在ura4基因的两端设计分别设计一段大小为1kb的序列用于进行ura4基因替换的同源臂;
(3)采用融合pcr将ura4基因上、下游同源臂与阿泊拉抗性基因进行融合,并与质粒poj260进行连接,获得重组质粒poj260-ura4;
(4)将含有重组质粒poj260-ura4的原生质体转入须糖多孢菌中,进行ura4基因的敲除,获得ura4缺失菌株s.pogona-δura4,通过线性片段同源重组技术对所述ura4缺失菌株s.pogona-δura4进行须糖多孢菌基因簇的阻断。
进一步,步骤(1)中所述ura4基因序列如序列表seqid№1所示。
进一步,步骤(2)中所述用于替换ura4基因的上游同源臂序列如序列表seqid№2所示;所述用于替换ura4基因的下游同源臂序列如序列表seqid№3所示。
进一步,步骤(3)中所述利用阿泊拉霉素抗性基因取代ura4基因,使须糖多孢菌具有阿泊拉霉素抗性,在后续对须糖多孢菌进行转化子筛选时,添加阿泊拉霉素。
进一步,步骤(4)获得ura4缺失菌株s.pogona-δura4作为须糖多孢菌基因簇进行大片段阻断的模式菌株。
进一步,步骤(4)所述通过线性片段同源重组技术对所述ura4缺失菌株s.pogona-δura4进行须糖多孢菌14号基因簇的阻断,包括以下步骤:
(a)在14号基因簇待阻断区域下游,基因组上位置为2,627,531~2,631,482,设计两对引物fc14up/rc14up和fc14down/rc14down进行上、下游同源臂扩增,用于将线性片段插入基因组中;
(b)在14号基因簇待敲除区域上游,基因组上位置为2,609,020~2,609,519,设计一对引物fds/rds进行directsequence扩增,用于将引入的外源ura4基因环外敲除;
(c)在须糖多孢菌基因组中设计一对引物fura4/rura4进行ura4基因扩增,作为负筛选标记基因进行转化子筛选;
(d)以质粒poj260-cm-perm*为模板,设计一对引物fperm/rperm用于红霉素强启动子(perm*)扩增,用于启动ura4基因表达;
(e)采用融合pcr将步骤(a)到步骤(d)获得的基因片段进行融合,获得可用于14号基因簇阻断的线性重组片段uac14-perm*-ura4-ds-dac14;
(f)原生质体转化线性重组片段进入s.pogona-δura4中进行重组、筛选,获得14号基因簇阻断的工程菌株s.pogona-δura4-δc14。
进一步,步骤(f)所得的须糖多孢菌14号基因簇阻断的工程菌株s.pogona-δura4-δc14的阻断区域为20kb序列。
本发明在确定好负筛选标记基因ura4的基础上,利用双交换同源重组将须糖多孢菌基因组中ura4基因敲除,获得不含ura4基因的突变菌株s.pogona-δura4,即完成进行基因簇大片段阻断的所需“底盘”细胞的构建;生理、生化分析结果表明,ura4基因敲除并不影响出发菌株的生长、菌丝形态改变以及丁烯基多杀菌素生物合成;对待阻断的14号基因簇进行线性片段的设计(包含两个同源臂,一个负筛选标记基因ure4和一个定向重复序列),组装(融合pcr),并转入突变菌株s.pogona-δura4原生质体中,使用5-foa(5-氟乳清酸)进行负筛选,获得基因簇cluster14阻断的工程菌株s.pogona-δura4-δ14,且pcr鉴定结果表明,已成功将cluster14的核心区域(约20kb)进行阻断。
发明有益效果:在须糖多孢菌基因组编辑中不仅避免了质粒的使用,还可以快速得到对特定基因簇阻断的突变株,是进行须糖多孢菌基因组编辑的有力工具。而且,本发明通过工程菌株s.pogona-δura4-δ14的获得,证实通过阻断竞争性基因簇对共有前体物质的利用可有效提高丁烯基多杀菌素产量。
附图说明
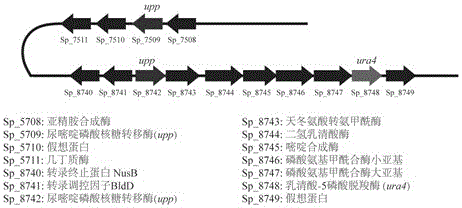
图1为本发明之可作为负筛选标记基因的ura4与upp在基因组上的分布;
图2为本发明之重组质粒poj260-ura4构建流程图;
图3为本发明之突变菌株s.pogona-δura4构建流程图;
图4为本发明之突变株菌s.pogona-δura4pcr验证图;
图5为本发明之突变株菌s.pogona-δura4生长曲线测定;
图6为本发明之突变株菌s.pogona-δura4第3天、第5天和第7天菌丝形态扫描电镜观察;
图7为本发明之突变株菌s.pogona-δura4(a)与原始菌株s.pogona(b)丁烯基多杀菌素产量分析;
图8为本发明之线性片段介导须糖多孢菌基因组大片段敲除的方法的流程图;
图9为本发明之须糖多孢菌14号基因簇基因结构分析;
图10为本发明之线性重组片段(a)与工程菌株s.pogona-δura4-δ14(b)pcr验证图;
图11为本发明之工程菌株s.pogona-δura4-δ14(a)与原始菌株s.pogona(b)丁烯基多杀菌素产量分析。
具体实施方式
下面结合实施例和附图对本发明作进一步说明。
本发明实施例所使用的化学试剂,如无特殊说明,均通过常规商业途径获得。
实施例1
(一)具体培养基配方和培养条件
(1)s.pogona种子活化培养基csm(completesyntheticmedium):trypticsoybroth4.5g/l,glucose0.9g/l,yeastextract0.3g/l,mgso4.7h2o0.22g/l。接种单克隆或2%菌种保藏液加入装有20mlcsm的150ml摇瓶中,30°c,280r/min培养48h;
(2)s.pogona合成发酵培养基(syntheticfermentationmedium,sfm):glucose20.0g/l,yeastextract4.0g/l,tryptone4.0g/l,kno31.0g/l,k2hpo4•3h2o0.5g/l,mgso4•7h2o0.5g/l,feso4•7h2o0.01g/l。接种2%的菌种活化液加入装有50mlsfm的300ml摇瓶中,30°c,280r/min培养12d。
(3)原生质体转化采用r5培养基:蔗糖103g/l,葡萄糖10g/l,酪蛋白氨基酸0.1g/l,yeastextract5g/l,k2so40.25g/l,mgcl2•6h2o10.12g/l,tes5.73g/l,微量元素溶液2ml(zncl240mg/l,fecl3•6h2o200mg/l,cucl2•2h2o10mg/l,mncl2•4h2o10mg/l,na2b4o7•10h2o10mg/l,(nh4)6mo7o4•4h2o10mg/l),琼脂粉15-20g/l,使用之前在培养基中分别加入已过滤除菌的kh2po3(0.5%)2.5ml/l,l-脯氨酸(20%)3.75ml/l和cacl2•2h2o(5m)1ml/l。
(4)e.colidh5α液体培养基lb(luria-bertani):tryptone1g/l,yeastextract0.5g/l和nacl1g/l。在上述液体中加入15-20g/l的琼脂粉可配制成lb固体培养基。
(二)须糖多孢菌基因组中负筛选标记基因的分析
在已报道的文献中,大多选择ura4(乳清苷-5-磷酸脱羧酶基因)和upp(尿嘧啶转磷酸核糖基酶)这两类基因作为负筛选标记基因。其中乳清苷-5-磷酸脱羧酶在5-foa(5-氟乳清酸)的存在下可将其转化为5-fu(5-氟尿嘧啶),而尿嘧啶转磷酸核糖基酶可将5-fu转化为5-fluoro-ump(5-氟尿苷酸),最终抑制胸苷酸合成酶的活性,引起细胞死亡。这两个负筛选标记基因的筛选条件分别是5-foa和5-fu。须糖多孢菌全基因组测序结果表明,该菌株存在1个ura4基因和2个upp基因,其在基因组位置(参见图1)。因此,为简化操作,我们最终选择ura4基因作为本次发明的负筛选基因。
(三)ura4基因敲除质粒构建
以s.pogona基因组为模板,设计两对引物fura4up/rura4up和fura4down/rura4down分别在ura4基因的上、下游扩增各1kb左右序列作为双交换的同源臂,引物序列见表1所示,其中引物fura4up和rura4down分别带有hindⅲ和ecori酶切位点,将用于与载体poj260进行连接。两段同源臂扩增的pcr反应体系为:h2o16.5µl,2xprimestargcbuffer25µl,dntps4µl,引物各2µl,模板s.pogona基因组dna2µl,primestar0.5µl。pcr反应程序设置为:95℃预变性5min,随后按95℃变性30s、56℃退火30s和72℃延伸90s,循环30次,最后再72℃维持10min,20℃终止反应。
以质粒poj260为模板,设计一对引物fapr/rapr用于扩增阿泊拉霉素抗性基因,引物序列见表1所示。上述引物对fapr/rapr的5’端各带有35bp可与ura4基因的上、下游同源臂重叠的序列,可用于进行三片段融合。pcr反应体系和反应程序与ura4基因上、下游同源臂设置一致。
将上述3种pcr产物进行纯化、回收,并以获得的回收产物作为模板,fura4up和rura4down为引物,采用fusionpcr方法,将ura4基因上、下游同源臂与阿泊拉霉素抗性基因apr进行无缝拼接,获得三片段融合产物uaura4-apr-daura4(fusionfragment)。随后对fusionfragment和poj26分别进行hindⅲ和ecori双酶切(双酶切体系:ddh2o10μl,10×mbuffer5μl,fusionfragment/poj26030μl,hindⅲ5μl,ecori5μl),产物回收纯化后,在t4dna连接酶的作用下进行融合片段与载体poj260连接(酶连体系:t4dnaligasebuffer1:t4dnaligase:linearpoj260:fusionfragment=1:1:1:7)。最后将连接产物热转入e.colidh5α感受态细胞中,并在lb抗性平板(阿泊拉霉素)上进行转化子筛选和验证,获得ura4敲除质粒poj260-ura4,重组质粒构建流程(参见图2)。
(四)突变菌株s.pogona-δura4的构建
使用原生质体转化的方式将外源重组质粒poj260-ura4转入s.pogona中,以上原生质体制备与转化方法可参考罗玉双,丁学知,夏立秋,王海龙,黄璠,唐滢在《生物工程学报》(2009年第25卷第3期第360-367页)上发表的“多杀菌素生产菌原生质体的制备条件及其再生菌株生理特性”文章记载的进行。获得转化子后,将其置于tsb液体培养基中,30℃摇床培养48h。收集1ml菌液并提取基因组作为模板,使用引物对fapr/rapr进行阿泊拉霉素抗性基因扩增,并以野生型s.pogona基因组为阴性对照,重组质粒poj260-ura4为阳性对照(由于poj260属于自杀型质粒,不能在s.pogona进行复制,导致传代n次后,该质粒会丢失,从而不会对转化子鉴定产生干扰)。将pcr鉴定正确的转化子(突变菌株),命名为s.pogona-δura4。结果(参见图3和图4),s.pogona-δura4在以引物对fapr/rapr进行pcr扩增时有约为1.5kb的条带出现,而野生型s.pogona无对应条带出现,说明apr基因成功替换掉ura4基因,至此,用于进行无痕敲除所需的宿主菌构建完成。
(五)突变菌株s.pogona-δura4表型分析
(1)ura4基因敲除对菌株生长的影响
分别将活化好的突变菌株s.pogona-δura4和原始菌株等量转入合成发酵培养基中,每12h取样进行菌体od600值测定(测定值在0.2~08之间,以保证测定结果的准确性,否则对样品进行稀释),取样周期为12d。实验结果(参见图5),突变菌株s.pogona-δura4与原始菌株s.pogona相比,生长曲线无明显变化。
(2)ura4基因敲除对菌株菌丝形态的影响
分别将活化好的突变菌株s.pogona-δura4和原始菌株等量转入合成发酵培养基中,并取第3天、第5天和第7天的菌液进行扫描电镜观察。样品经2.5%的戊二醛固定以及乙醇脱水处理后,使用冷场发射扫描电子显微镜su8010进行突变菌株s.pogona-δura4与原始菌株菌丝体形态观察。扫描电镜观察结果显示(参见图6),突变菌株s.pogona-δura4菌丝体形态在第3天和第5天呈现分支丝状,培养至第7天后,菌丝体形态逐渐变为菌丝断裂状态且少分支,其菌丝体变化过程与原始菌株s.pogona一致。
(3)ura4基因敲除对丁烯基多杀菌素生物合成的影响
分别将活化好的突变s.pogona-δura4和原始菌株等量转入合成发酵培养基中,取培养至第10天的发酵液进行丁烯基多杀菌素产量检测。样品处理过程可参与文献(罗林根,杨燕,魏慧,穰杰,唐琼,胡胜标,孙运军,余子全,丁学知,夏立秋在《生物工程学报》(2016年第32卷第2期第259-263页)上发表的“须糖多孢菌saccharopolysporapogona的核糖体工程改造对丁烯基多杀菌素合成的影响”文章记载的进行。hplc1290检测结果(参见图7),突变菌株s.pogona-δura4与原始菌株s.pogona丁烯基多杀菌素峰面积分别为,统计学上无明显差异。
上述表型分析结果显示,ura4基因敲除后并未对菌株生长、发育和丁烯基多杀菌素生物合成产生不利影响,说明将ura4基因作为须糖多孢菌负筛选标记基因选择正确。
(六)线性片段同源重组技术用于目标基因簇cluster14的阻断
须糖多孢菌基因组中存在28个次级代谢产物基因簇,我们随机选择14号基因簇,非核糖体多肽基因簇(参见图9),作为靶标进行上述大片段敲除方法的验证。
(1)线性重组片段uac14-perm*-ura4-ds-dac14体外构建
以s.pogona基因组为模板,设计四对引物fura4/rura4,fc14up/rc14up、fc14down/rc14down和fds/rds进行ura4基因、上、下游同源臂和定向重复序列扩增,分别用于进行重组菌株负筛选、线性重组片段整合入基因组以及负筛选标记基因(ura4)删除,引物序列如表1所示,其中引物fc14down,fsd和rsd的5’各添加有35bp用于进行片段融合的碱基序列。pcr反应体系和反应程序与上述设置一致。
表1本发明所用菌株、质粒和引物
以质粒poj260-cm-perm*为模板,设计一对引物fperm/rperm用于扩增红霉素强启动子(perm*),引物序列如表1所示,其中引物rprem的5’添加有35bp用于进行片段融合的碱基序列。pcr反应体系和反应程序与上述设置一致。
最后,将上述5种pcr产物进行回收纯化,并以获得的回收产物作为模板,fc14up和rc14down为引物,采用fusionpcr方法,将上述5种片段进行“缝合”,获得五片融合产物(uac14-perm*-ura4-ds-dac14),构建方法(参见图8)。
(2)工程菌株s.pogona-δure4-δ14的构建
使用原生质体转化的方式将线性重组片段uac14-perm*-ura4-ds-dac14转入s.pogona-δura4中并发生重组;取100μl菌液涂布于含有5-foa的mm(基础培养基)平板上进行负筛选,30℃恒温培养箱倒置培养直至白色转化子出现;挑取单克隆划线于含有5-foa的mm平板上进行分离纯化,重复2-3次后,获得最终的转化子。
在mm平板上挑取部分菌苔于tsb液体培养基中,30℃摇床培养48h,收集1ml菌液并提取基因组作为模板,使用引物对fsd/rc14down进行pcr验证,并以野生型s.pogona基因组为阴性对照。pcr鉴定正确的转化子(工程菌株),命名为s.pogona-δura4-δc14。结果显示,s.pogona-δura4-δc14在以引物对fsd/rc14down进行验证时会出现约为1.8kb的条带,而原始菌株s.pogona无条带出现(参见图9),说明已成功对该基因簇目标区域进行阻断。
(七)工程菌株s.pogona-δura4-δc14丁烯基多杀菌素产量分析
分别将活化好的工程菌株s.pogona-δura4-δc14和原始菌株s.pogona等od600值转入合成发酵培养基中,取培养至第10天的发酵液进行丁烯基多杀菌素产量检测。hplc1290检测结果如图10所示,工程菌株s.pogona-δura4-δc14与原始菌株s.pogona相比,除5.3min的色谱峰外,在8min出现明显的色谱峰,该峰经质谱鉴定为丁烯基多杀菌素衍生物spinosynαa,因此,工程菌株s.pogona-δura4-δc14丁烯基多杀菌素总峰面积分别为1904.7,而原始菌株总峰面积为342.1mau*min,最终基因簇cluster14阻断后,使得丁烯基多杀菌素产量提高了5.57倍。
序列表
<110>湖南师范大学
<120>基于线性片段同源重组进行须糖多孢菌基因簇阻断的方法
<160>3
<170>siposequencelisting1.0
<210>1
<211>849
<212>dna
<213>须糖多孢菌(saccharopolysporapogona)
<400>1
atgactgatccgctgcgggcgggcttcggccagcggctggcggagtcggtgcgggcgcgc60
ggtgcgctgtgcgtcggcatcgacccgcacccgtcgctgctgcacgcgtgggatctcgac120
gagaccccgcagtcgctggagcggttcgcgatgaccgcggtggaggcgctggccggcgag180
gtcgcggtgctcaagccgcagtcggcgttcttcgaggtgtacggctcggccggaatcgcc240
gtgctggagaagacgatccgggaggcgcggcaggccggtgcgctggtgctgctggacatc300
aagcgcggcgacatcggctccacgatggcggcgtacgcgatggcctacctggacgagcac360
gcgccgctggcggcggatgcgatcacggtgtcgccgtacctcggctacggctcgctcgcc420
ccggcgatcggcatcgccgagcagacgggccgcggggtgttcgtgctggcgcgcacctcc480
aacccggagggcgcgggcctccagcggtcgatccacggcagcggcgactcggtcgcccag540
tacatcgtggactcggccgcggagaccaatgcggacgcggagccgatggggcacgtcgga600
gtggtggcgggagccaccatcggcgccgacgagctggacttctcccgcctcaacggcccg660
atcctggcacccggcctgggtgcgcagggtgcgacggtgcagagcctgcgcgcgctgttc720
ggcgcggcgctgccgaacgtcctaccggcgacggcccgcgacgtcctccggcacggcccg780
acggtcaacggcctccgcgccgcggcccacaaggtccgcgacgaggtgagcgacctcctc840
cgagactga849
<210>2
<211>1274
<212>dna
<213>须糖多孢菌(saccharopolysporapogona)
<400>2
tggtggacaacttcctcgacgacgcgatcgagatcgacgtggacgcgctctgcgacggca60
ccgacatctacctcggtggcgtgatggaacacatcgaggaggccggtatccactccggcg120
actcggcgtgcgcgctgccgccgatcacgttggggcgtcaggacatcgagcaggtgcgaa180
agtccaccgaggccatcgcgcgcggcatcggcgtgcacggcctgctgaacgtgcagtacg240
cgctcaaggacgacgtgctgtacgtgctggaggccaacccgcgcgcctcgcggaccgtgc300
cgttcgtgtccaaggcgaccgccgtgccgctggccaaggcggcggcccggatcatgctcg360
gcgccaagatcgcggacctgcgcgcggagggcctgctgccgccggagggcgacggcgcgg420
aactgccgatcgacgcgccggtcgcggtgaaggaagccgtgctgccgttccaccggttcc480
gcacccgcgagggcgtcggcgtggactcgctgctcgggccggagatgaagtccaccggcg540
aggtgatgggcatcgacacctccttcgggcaggccttcgccaagtcgcaggccggcgcct600
acggttcgctgccgaccggcggccgggtgttcgtgtcggtggccaacaaggacaagcggt660
ccctggtgttcccggtcaagcggctggcggacctcggcttcgagatcctggccaccagcg720
gcaccgcggaggtgctgcgccgcaacggaatcccgtccacggtggtgcgcaagcacatcg780
agaagaacggcgcggagcgcgacatcgtcgagctgatcaaggccggcgaggtcgacatgg840
tgatcaacaccccgtacggcaacccgggcccgcgcgtggacggctacgagatccgcaccg900
ccgcggtatcccgcgacatcccgtgcatcaccacggtgcagggcgcggcggcggccgtgc960
agggcatcgaggccgccatccaaggcaacatcggggtccggcccctgcaggcgttgcagg1020
cggcccttcggcagggcggtgagcagcgatgactgatccgctgcgggcgggcttcggcca1080
gcggctggcggagtcggtgcgggcgcgcggtgcgctgtgcgtcggcatcgacccgcaccc1140
gtcgctgctgcacgcgtgggatctcgacgagaccccgcagtcgctggagcggttcgcgat1200
gaccgcggtggaggcgctggccggcgaggtcgcggtgctcaagccgcagtcggcgttctt1260
cgaggtgtacggct1274
<210>3
<211>1172
<212>dna
<213>须糖多孢菌(saccharopolysporapogona)
<400>3
ggaaacggctatctggtacgaagttcgcatgcatcccgcaacgaacggcgccctcgcgca60
gaagaaaccgccgcgcgtgctgcgctggatcatcgttctcgtgctgctctggctggtcac120
cggtggctatttcctggtttccaccgaggtcaccatctggtcccggaccttcggctcggg180
cgaggccacggccgacccagtcgcacacgtggagctggagcgggggcagccgttcggcct240
ggccgcgcaacgggatgagtacgtgaagtgccaggtgatcccgcaggccggtgagccgcg300
tgagttcattcccgaccgcccggccaacagcctgcgcagctcgcggctgcccgggccgga360
acccgcgtggttctccggcccggccgaggcccgctgcaccgggcccaccacggtgctcct420
gccggagcgctacgacacgaccgggctgaacgtcctgctggggctgattgctgcggccag480
cctcatgatcgtggtcgtttttgtacaattcgcgcgtcgtgcccgtttcaacaataggtt540
gtacgcccgcgactgatttgctcctgggaacggatctccgaacggcatgaggcgccgtgg600
gcggtcagatagtgtcgatccaacggttcggaagctaaggggacagacggtgagcagcgc660
gtctgagcgcatccaggagatgatgcgcaagttcgaggagcaggcgcagaaggcgtcgga720
actgcagtccgcgatgcagggcatgcaggccacggcgagcagcccggaccggtcggtgac780
ggtgacggtcgcgccgtccggggcggtgctggacctgcgtctggcgccgaacgcggtgcg840
ccagtcggccaacgacctgcagcagcagatcatggcgaccatccgccaagcaacggcaag900
cgcggcggagcagctcaacaacacggtggcgccgatcctgggcgaccagttcgacaagtt960
ccaggaagcgttcaacgtcgaaggcatggcgatcaagccgaccgccggagacgaaacgcc1020
cgcaccggagagccccgcaccgccgcccgccgcgcaccgcgcgccgccggagagtccggc1080
aacgccgcggcggccgcagcaggcgagcgccgattacgacggtgacgacttctcttcggg1140
ctcgttcctgcgctgagggaagttgtcttgac1172
序列表
<110>湖南师范大学
<120>基于线性片段同源重组进行须糖多孢菌基因簇阻断的方法
<160>3
<170>siposequencelisting1.0
<210>1
<211>849
<212>dna
<213>须糖多孢菌(saccharopolysporapogona)
<400>1
atgactgatccgctgcgggcgggcttcggccagcggctggcggagtcggtgcgggcgcgc60
ggtgcgctgtgcgtcggcatcgacccgcacccgtcgctgctgcacgcgtgggatctcgac120
gagaccccgcagtcgctggagcggttcgcgatgaccgcggtggaggcgctggccggcgag180
gtcgcggtgctcaagccgcagtcggcgttcttcgaggtgtacggctcggccggaatcgcc240
gtgctggagaagacgatccgggaggcgcggcaggccggtgcgctggtgctgctggacatc300
aagcgcggcgacatcggctccacgatggcggcgtacgcgatggcctacctggacgagcac360
gcgccgctggcggcggatgcgatcacggtgtcgccgtacctcggctacggctcgctcgcc420
ccggcgatcggcatcgccgagcagacgggccgcggggtgttcgtgctggcgcgcacctcc480
aacccggagggcgcgggcctccagcggtcgatccacggcagcggcgactcggtcgcccag540
tacatcgtggactcggccgcggagaccaatgcggacgcggagccgatggggcacgtcgga600
gtggtggcgggagccaccatcggcgccgacgagctggacttctcccgcctcaacggcccg660
atcctggcacccggcctgggtgcgcagggtgcgacggtgcagagcctgcgcgcgctgttc720
ggcgcggcgctgccgaacgtcctaccggcgacggcccgcgacgtcctccggcacggcccg780
acggtcaacggcctccgcgccgcggcccacaaggtccgcgacgaggtgagcgacctcctc840
cgagactga849
<210>2
<211>1274
<212>dna
<213>须糖多孢菌(saccharopolysporapogona)
<400>2
tggtggacaacttcctcgacgacgcgatcgagatcgacgtggacgcgctctgcgacggca60
ccgacatctacctcggtggcgtgatggaacacatcgaggaggccggtatccactccggcg120
actcggcgtgcgcgctgccgccgatcacgttggggcgtcaggacatcgagcaggtgcgaa180
agtccaccgaggccatcgcgcgcggcatcggcgtgcacggcctgctgaacgtgcagtacg240
cgctcaaggacgacgtgctgtacgtgctggaggccaacccgcgcgcctcgcggaccgtgc300
cgttcgtgtccaaggcgaccgccgtgccgctggccaaggcggcggcccggatcatgctcg360
gcgccaagatcgcggacctgcgcgcggagggcctgctgccgccggagggcgacggcgcgg420
aactgccgatcgacgcgccggtcgcggtgaaggaagccgtgctgccgttccaccggttcc480
gcacccgcgagggcgtcggcgtggactcgctgctcgggccggagatgaagtccaccggcg540
aggtgatgggcatcgacacctccttcgggcaggccttcgccaagtcgcaggccggcgcct600
acggttcgctgccgaccggcggccgggtgttcgtgtcggtggccaacaaggacaagcggt660
ccctggtgttcccggtcaagcggctggcggacctcggcttcgagatcctggccaccagcg720
gcaccgcggaggtgctgcgccgcaacggaatcccgtccacggtggtgcgcaagcacatcg780
agaagaacggcgcggagcgcgacatcgtcgagctgatcaaggccggcgaggtcgacatgg840
tgatcaacaccccgtacggcaacccgggcccgcgcgtggacggctacgagatccgcaccg900
ccgcggtatcccgcgacatcccgtgcatcaccacggtgcagggcgcggcggcggccgtgc960
agggcatcgaggccgccatccaaggcaacatcggggtccggcccctgcaggcgttgcagg1020
cggcccttcggcagggcggtgagcagcgatgactgatccgctgcgggcgggcttcggcca1080
gcggctggcggagtcggtgcgggcgcgcggtgcgctgtgcgtcggcatcgacccgcaccc1140
gtcgctgctgcacgcgtgggatctcgacgagaccccgcagtcgctggagcggttcgcgat1200
gaccgcggtggaggcgctggccggcgaggtcgcggtgctcaagccgcagtcggcgttctt1260
cgaggtgtacggct1274
<210>3
<211>1172
<212>dna
<213>须糖多孢菌(saccharopolysporapogona)
<400>3
ggaaacggctatctggtacgaagttcgcatgcatcccgcaacgaacggcgccctcgcgca60
gaagaaaccgccgcgcgtgctgcgctggatcatcgttctcgtgctgctctggctggtcac120
cggtggctatttcctggtttccaccgaggtcaccatctggtcccggaccttcggctcggg180
cgaggccacggccgacccagtcgcacacgtggagctggagcgggggcagccgttcggcct240
ggccgcgcaacgggatgagtacgtgaagtgccaggtgatcccgcaggccggtgagccgcg300
tgagttcattcccgaccgcccggccaacagcctgcgcagctcgcggctgcccgggccgga360
acccgcgtggttctccggcccggccgaggcccgctgcaccgggcccaccacggtgctcct420
gccggagcgctacgacacgaccgggctgaacgtcctgctggggctgattgctgcggccag480
cctcatgatcgtggtcgtttttgtacaattcgcgcgtcgtgcccgtttcaacaataggtt540
gtacgcccgcgactgatttgctcctgggaacggatctccgaacggcatgaggcgccgtgg600
gcggtcagatagtgtcgatccaacggttcggaagctaaggggacagacggtgagcagcgc660
gtctgagcgcatccaggagatgatgcgcaagttcgaggagcaggcgcagaaggcgtcgga720
actgcagtccgcgatgcagggcatgcaggccacggcgagcagcccggaccggtcggtgac780
ggtgacggtcgcgccgtccggggcggtgctggacctgcgtctggcgccgaacgcggtgcg840
ccagtcggccaacgacctgcagcagcagatcatggcgaccatccgccaagcaacggcaag900
cgcggcggagcagctcaacaacacggtggcgccgatcctgggcgaccagttcgacaagtt960
ccaggaagcgttcaacgtcgaaggcatggcgatcaagccgaccgccggagacgaaacgcc1020
cgcaccggagagccccgcaccgccgcccgccgcgcaccgcgcgccgccggagagtccggc1080
aacgccgcggcggccgcagcaggcgagcgccgattacgacggtgacgacttctcttcggg1140
ctcgttcctgcgctgagggaagttgtcttgac1172
- 还没有人留言评论。精彩留言会获得点赞!