粉棒束孢Cyanovirin-N基因、重组蛋白及应用的制作方法
本发明属于分子生物学领域,具体涉及粉棒束孢cyanovirin-n基因、基因和重组蛋白的制备方法、重组蛋白及应用。
背景技术:
粉棒束孢(isariafarinosa),是丝状真菌棒束孢属(isariafries)模式种,是一种主要的昆虫病原真菌,同时也是一种极具开发价值和商业利用价值的真菌,具体表现在:1)现有研究表明,粉棒束孢对多种类型的害虫具有良好的杀虫效果;2)粉棒束孢作为蝙蝠蛾幼虫的定殖真菌,其药用活性也越来越多地被研究和发现;3)已从粉棒束孢培养菌丝体中分离得到包括多糖、虫草酸、生物碱、吡啶酮、天然苯腙、喹唑啉酮、环状五肽、蒽醌类化合物等多种新化合物;4)研究证明粉棒束孢菌的代谢产物中具有类似生长素和细胞分裂素的物质等。然而目前针对粉棒束孢的研究主流还停留在宏观层面上,主要以粉棒束孢的代谢产物或孢子悬液或菌丝体作为研究对象,其微观分子层面的研究才刚刚起步,且现有的利用分子生物学手段对粉棒束孢进行的深入和系统研究,主要都集中在真菌分类及作为生防菌的角度进行。
申请号为201610476131.6的发明专利公开了一株粉棒束孢的发酵培养物在制备预防林业害虫的生防制剂中的应用,该发明主要是利用了菌株的发酵培养液。杨俊媛等学者从粉棒束孢培养菌丝体中分离得到多种有价值的化合物,但其研究依然停留在宏观层面。因此,有必要在全面、系统研究活性代谢产物的基础上深入开展粉棒束孢药理活性与基因功能之间的关系,以便构建出能够产生特定活性成分的超级生产菌株。
鉴于此,本发明从一株具有较强杀虫活力的粉棒束孢菌株ftrsy-2出发,在筛选分析基因信息时,发现了一个与蓝藻抗病毒蛋白-n(cyanovirin-n)同源的基因,并首次从粉棒束孢基因组中克隆出粉棒束孢cvn基因,命名为ifcvn。本发明以该基因为出发点,通过分子生物学的手段构建重组载体、工程菌、重组cvn蛋白等,并通过构建ifcvn基因敲除与回复株验证该基因的功能,为进一步研究粉棒束孢功能基因及其潜在的药用价值奠定了基础,具有重要的研究意义和潜在的商业价值。
技术实现要素:
有鉴于此,本发明的目的之一,在于提供粉棒束孢的cyanovirin-n基因。
粉棒束孢的cyanovirin-n基因,所述的cyanovirin-n基因的核苷酸序列包括seqidno.1所示和/或其中的片段。该基因由本发明的课题组首次从粉棒束孢基因组中克隆得到,命名为ifcvn。
本发明的目的之二,在于提供粉棒束孢的cyanovirin-n基因的制备方法。
上述cyanovirin-n基因的制备方法,根据粉棒束孢全基因组序列预测得到seqidno.1所示序列,根据所述seqidno.1序列设计若干引物对;提取粉棒束孢总rna,反转cdna,进行pcr扩增,获得如seqidno.1所示的cyanovirin-n基因。cyanovirin-n基因的cdnaorf全长为369bp,编码122个氨基酸,分子量约为12.9kda,与现有报道的天然成熟的cvn氨基酸残基数目及相对分子质量有一定差异。
进一步,所述引物对包括如序列seqidno.2和seqidno.3所示的一组引物对,所述序列seqidno.2的引物命名为ifcvnf;所述序列seqidno.3的引物命名为ifcvnr。
本发明的目的之三,在于提供一种重组质粒载体。
重组质粒载体,所述重组质粒载体包含权利要求1所述的seqidno.1序列和原核表达质粒;所述seqidno.1序列与原核表达质粒连接。
作为一种优选,所述原核表达质粒采用sumo-pet28a质粒。
本发明的目的之四,在于提供一种基因工程菌。
重组质粒载体转化得到的产重组cyanovirin-n蛋白的基因工程菌。
进一步,所述重组cyanovirin-n蛋白在所述基因工程菌的包涵体和培养上清液中表达。本发明的基因工程菌用大肠杆菌bl21(de3)制备。
本发明的目的之五,在于提供一种重组cvn蛋白。
粉棒束孢的重组cyanovirin-n蛋白,所述重组cyanovirin-n蛋白的氨基酸序列包括seqidno.4所示和/或其中的片段。
粉棒束孢的重组cyanovirin-n蛋白,所述重组cyanovirin-n蛋白由上述的seqidno.1核苷酸序列翻译得到。
天然的cvn蛋白最初是从蓝藻椭孢念株藻(nostocelliposporum)中分离得到的一种水溶性糖蛋白。cvn具有较广谱的抗病毒活性,同时还具有稳定的理化特性,能抵抗变性剂、去污剂和有机溶剂的处理。cvn的基因工程研究在cvn被发现后就开始了,但椭孢念株藻养殖困难,从中分离纯化cvn不仅成本高、工艺复杂,而且原料浪费严重。因此本发明首次从粉棒束孢基因组中克隆出粉棒束孢cvn基因,并借助分子生物学手段得到稳定高产的重组cvn蛋白,具有突破性的意义。
本发明的目的之六,在于提供一种多克隆抗体。
一种多克隆抗体,所述多克隆抗体包括上述的重组cyanovirin-n蛋白。
本发明的目的之七,在于提供上述重组cvn蛋白的一系列应用。
上述重组cyanovirin-n蛋白在制备用于防治林业害虫的生防制剂中的应用。所述应用还可以由上述seqidno.1的核苷酸序列和上述重组质粒载体得到。
进一步,所述林业害虫包括油松毛虫、松梢螟、苹果蠹蛾、美国白蛾、落叶松卷蛾、黄刺蛾幼虫、板栗象甲、松墨天牛、柑桔粉蚧中的一种或多种。传统的生防制剂主要是以菌株或菌株代谢产物为基础制备,而以本发明的重组cvn蛋白为基础来制备,效力更强,可控性更高。
上述重组cyanovirin-n蛋白在制备广谱抗菌药物中的应用。cvn蛋白具有广谱的抗病毒活性和稳定的生化特性,是一种新型的抗菌活性物。所述应用还可以由上述seqidno.1的核苷酸序列和上述重组质粒载体得到。
进一步,所述细菌包括水生拉恩氏菌(rahnellaaquatilis)、e.persicina、普城沙雷菌(serratiaplymuthica)、平凡假单胞菌(pseudomonastrivialis)、菠萝泛菌(pantoeaananatis)、土生柔特勒菌(raoultellaterrigena)、河生肠杆菌(lelliottiaamnigena)中的一种或多种。
进一步,所述重组cyanovirin-n蛋白可以为任意药学可接受剂型,所述广谱抗菌药物还包括药学上可接受的载体和/或助剂。
上述重组cyanovirin-n蛋白在制备多克隆抗体中的应用。所述应用还可以由上述seqidno.1的核苷酸序列和上述重组质粒载体得到。
上述重组cyanovirin-n蛋白在制备抗艾滋病疫苗中的应用。所述应用还可以由上述seqidno.1的核苷酸序列和上述重组质粒载体得到。天然的cvn蛋白从椭孢念株藻(nostocelliposporum)的培养基提取物中分离得到,前期临床研究证实其能够不可逆地阻止hiv进入宿主细胞,因此有望成为第1种多肽类的天然抗hiv药物,但目前对其他藻类及真菌是否能分离出cvn少见报道。本发明成功构建出一种稳定性较强、产量和纯度较高的重组cvn蛋白,为后续进一步制备抗艾滋病疫苗提供了高纯度、高效、廉价的核心原材料。
上述重组cyanovirin-n蛋白在制备提高人体免疫功能的保健品中的应用。所述应用还可以由上述seqidno.1的核苷酸序列和上述重组质粒载体得到。通过对cvn蛋白的系统进化分析发现粉棒束孢与冬虫夏草菌进化关系最近,进化树聚为一枝,表明在虫生真菌尤其是虫草真菌中具有同一起源,而冬虫夏草被公认为具有调节免疫、抗疲劳等多种功效。因此本发明的重组cyanovirin-n蛋白及相应菌株具备了与冬虫夏草相当的开发成保健品的潜力。
上述重组cyanovirin-n蛋白在增强虫草抗逆性能中的应用。所述应用还可以由上述seqidno.1的核苷酸序列和上述重组质粒载体得到。
本发明的有益效果在于:
1)本发明从一株具有较强杀虫活力的粉棒束孢ftrsy-2菌株出发,首次从粉棒束孢基因组中克隆出粉棒束孢cvn基因,命名为ifcvn。以该基因为出发点,通过分子生物学的手段构建重组载体、工程菌、重组cvn蛋白等,并通过构建ifcvn基因敲除与回复株验证该基因的功能。从而为粉棒束孢功能基因的研究及其对寄主、环境互作分子机制的阐明、药用价值的深入研究奠定理论基础,同时为构建产生特定功能的活性成分的超级菌株奠定了基础。
2)本发明得到的重组cvn蛋白性质稳定,产量和纯度较高,并具有广谱杀菌效力和药理活性,有潜力开发成为生防制剂、抗菌药品、多肽疫苗、保健产品。
附图说明
图1:ifcvn基因的dna序列及所对应的氨基酸序列(框中为信号肽)。
图2:重组cvn蛋白二级结构预测。
图3:重组cvn蛋白三维结构图。
图4:粉棒束孢重组cvn与其他真菌cvn的氨基酸序列相似性分析(*标注为活性位点)。
图5:粉棒束孢重组cvn蛋白与其他真菌cvn蛋白的系统进化树。
图6:粉棒束孢ifcvn基因扩增。
图7:ifcvn基因同源敲除及回复载体构建过程图。
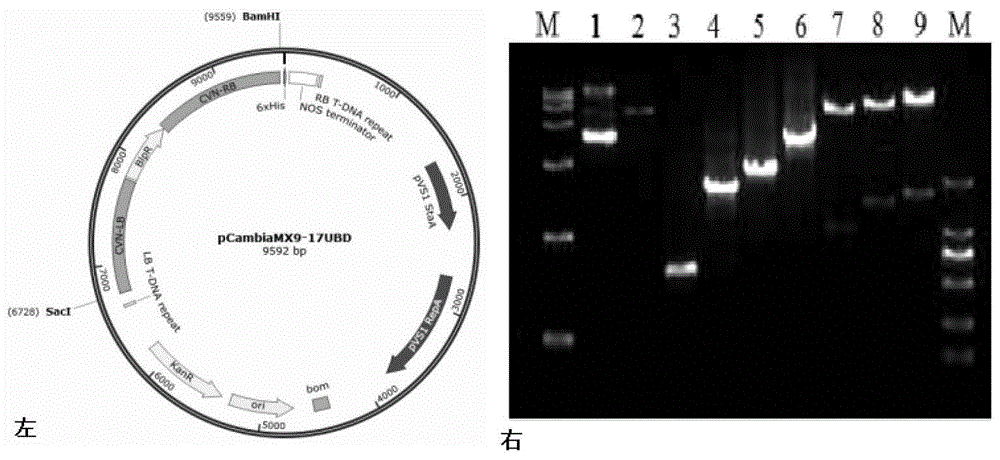
图8:ifcvn基因同源敲除载体图谱(左)及pcr验证(右)。
图9:ifcvn基因回复载体图谱(左)及pcr验证(右)。
图10:ifcvn基因同源敲除转化子bar基因pcr验证(m:dl2000marker;1-12:ifcvn基因敲除转化子;p:pcambiamx9-ifcvnubd;w:ftrsy-2)。
图11:ifcvn基因同源敲除、回复转化子pcr验证(左)及回复转化子荧光观察(右)(m:dl2000marker;1-3:ifcvn基因敲除转化子;4-6:ifcvn基因回复转化子)。
图12:ifcvn基因同源敲除、回复转化子菌落形态及逆境胁迫下的生长情况(wt:野生菌株;△ifcvn:ifcvn基因敲除突变株;rc:回复突变株;1-7:pda正常培养基;刚果红;nacl;山梨醇;ph11;uv;35℃)。
图13:ifcvn基因在感病大蜡螟不同时间及不同组织中的表达量。
图14:ifcvn基因扩增、表达载体酶切验证及重组蛋白表达(a、b:m:dl2000marker;1:ifcvn扩增产物;2:bamhi/sali酶切验证;c:1:0.4mg/mlbsa;2:上清;3marker;4:sumo-pet28a-sumo空载诱导表达;5:上清2(2m尿素溶解包涵体);6:包涵体2倍稀释(2m尿素溶解包涵体);7:包涵体2倍稀释(2m尿素溶解包涵体))。
图15:cvn重组蛋白特异性检测及感病大蜡螟血液中ifcvn基因表达量测定(a:多克隆抗体特异性及灵敏度检测;b:感病大蜡螟血液中cvn蛋白表达量检测;(m:marker;1-2:ftrsy-2野生株总蛋白;3:ck;4-8:0h、12h、24h、48h、72h感病大蜡螟血液总蛋白;9:阴性对照)。
图16:不同浓度重组cvn的wst测试及(左)对sf9细胞影响的增殖的影响(右)(a:cell+cck8;b:cell+sumo-cvn+cck8;c:sumo-cvn+cck8;a-h:sumo-cvn使用浓度梯度)。
具体实施方式
以下将参照附图,对本发明的优选实施例进行详细描述。按照常规条件,所举实施例是为了更好地对本发明的内容进行说明,但并不是本发明的内容仅限于所举实施例。所以熟悉本领域的技术人员根据上述发明内容对实施方案进行非本质的改进和调整,仍属于本发明的保护范围。
本发明的实验材料
1.实验材料
1.1供试菌株及昆虫
粉棒束孢ftrsy-2菌株;大肠杆菌bl21(de3);大蜡螟幼虫
1.2质粒载体
pcambiamx9质粒(genbankaccessionnumber:kx755248,具kana抗性);
pgapneor12质粒(genbankaccessionnumber:ky363244,具g418抗性)。
实施例1
1.基因组dna提取
基因组dna提取采用天根植物基因组提取试剂盒,按说明书步骤提取。
2.基因组rna提取
基因组rna提取采用天根植物基因组提取试剂盒,按说明书步骤提取。
3.微量快速pcr验证
采取微量菌丝pcr扩增:无菌牙签挑取少量粉棒束孢菌丝至ep管中,液氮速冻研磨后,加入50μllysisbuffer,混合均匀,95℃,10min;取菌丝裂解液2μl进行常规pcr扩增。
4.生物信息学分析
运用常规的生物信息分析软件进行。
5.生物信息学分析结果
ifcvncdnaorf全长为369bp,编码122个氨基酸(图1),分子量约为12.9kda,等电点为5.99,平均疏水性为-0.236,且稳定性比较好(instabilityindex(ii))为27.64。信号肽预测结果表明该序列具有信号肽,其切割位点在n端第26和27个氨基酸之间。二硫键预测表明该序列2个二硫键分别位于第8位和第22位及第53位和第79位的半胱氨酸残基上;二级结构预测如图2所示,表明该蛋白序列由α螺旋、β折叠、β转角及无规则卷曲构成;三级结构预测结果如图3所示,其主要由β折叠片形成的链状蛋白,折叠以后的构象呈现假二重对称;多序列比对结果显示粉棒束孢cvn氨基酸序列和稻曲霉、巴西青霉、层生镰刀菌、底栖蓝藻、亚麻色单歧藻、霍氏伪枝蓝细菌的cvn氨基酸序列相似度不高,序列较特异(图4),保守性最突出的区域为cyanovirin-n的活性中心(亮氨酸leul、异亮氨酸ilei、天冬氨酸aspd、甘氨酸glyg),这些保守结构域和氨基酸残基对cvn功能起着非常重要的作用;对cvn蛋白系统进化分析发现,粉棒束孢与冬虫夏草菌(cordycepsconfragosa)聚为一枝(图5)。
实施例2
基因cdnaorf全长克隆:
查找粉棒束孢全基因组序列得到预测的ifcvn基因全长序列,根据该序列设计引物ifcvnf/r。提取粉棒束孢总rna,反转cdna,进行pcr扩增,获得ifcvn基因cdnaorf全长。取5μlpcr扩增产物进行电泳验证。
实验结果:
扩增得到全长为369bp大小的片段,pcr扩增结果如图6所示。
实施例3
1.ifcvn基因敲除盒的构建
(1)ifcvn基因上、下游及草丁膦抗性基因片段克隆
以野生型菌株ftrsy-2基因组为模板,使用引物ifcvnqcupf和ifcvnqcupr扩增ifcvn基因上游序列,产物命名为ifcvnu;使用引物ifcvnqcdownf和ifcvnqcdownr扩增ifcvn基因下游序列,产物命名为ifcvnd;以pcambaregfp质粒为模板,使用引物bar5/6扩增草丁膦抗性基因,产物命名为b。电泳检测后切胶回收备用。
(2)质粒pcambiamx9-ifcvnu构建
将提取的质粒pcambiamx9和回收的ifcvn基因上游回收片段ifcvnu分别进行saci和hindiii双酶切,电泳检测后切胶回收,将回收的线性化质粒pcambiamx9和ifcvn基因上游片段ifcvnu进行连接,并将连接产物转化大肠杆菌dh5α,用引物mx9f/r对进行菌液pcr验证;提取pcr检测结果为阳性菌落的质粒,用saci和bamhi对质粒进行双酶切验证。
(3)质粒pcambiamx9-ifcvnud构建
将提取的质粒pcambiamx9-ifcvnu和回收的ifcvn基因下游回收片段ifcvnd分别进行hindiii和bamhi双酶切,电泳检测后切胶回收,将回收的线性化质粒pcambiamx9-ifcvnu和ifcvn基因下游片段ifcvnd进行连接,并转化大肠杆菌dh5α,用引物mx9f/r对进行菌液pcr验证;提取pcr检测结果为阳性菌落的质粒,用saci和bamhi对质粒进行双酶切验证。
(4)质粒pcambiamx9-ifcvnubd构建
将提取的质粒pcambiamx9-ifcvnud和回收的草丁膦抗性基因片段b分别进行hindiii酶切。电泳检测后切胶回收,将回收的线性化质粒pcambiamx9-ifcvnud和草丁膦抗性基因片段b连接,将连接产物转化大肠杆菌dh5α,用引物mx9f/r对进行菌液pcr验证;提取pcr检测结果为阳性的质粒,用saci和bamhi对质粒进行双酶切验证。
实验结果:
敲除载体构建及验证过程如图7所示,成功扩增获得ifcvn基因左右臂侧翼序列和bar基因元件,测序正确后按顺序依次连接后形成ifcvn基因敲除盒,成功连接到敲除骨架载体pcambiamx9上,图谱及验证结果见图8。
2.ifcvn基因回补盒的构建
采用同源重组方法构建基因回补盒。
(1)ifcvn基因回补片段扩增回收
以野生型菌株ftrsy-2基因组为模板,用引物hb1/2扩增ifcvn及其假定的启动子序列,产物命名为ifcvnh。pcr反应电泳检测后,切胶回收。
(2)回补质粒pgapneocvn构建
将提取的质粒pgapneor12进行bamhi酶切线性化。电泳检测后切胶回收,将回收的线性化质粒pgapneor12和ifcvn基因回补片段ifcvnh进行同源重组连接,并产物转化大肠杆菌dh5α,用引物r12f/r对转化子进行菌液pcr验证;pcr检测结果为阳性的菌落,提取质粒,用bamhi对质粒进行酶切验证。
实验结果:
回复载体构建及验证过程如图9所示。
实施例4
1.粉棒束孢ifcvn基因敲除及回复突变株筛选
(1)转化子筛选用g418浓度的确定
确定突变株筛选时的抗生素浓度,分别测试野生菌株ftrsy-2菌株对草丁膦的敏感度、对头孢霉素的敏感度及头孢霉素对农杆菌lba4404的毒性,从而确定筛选转化子的草丁膦和头孢霉素的最适使用浓度。由于回复采用的pgapneor12质粒带有遗传霉素(g418)抗性标签,因此需要对野生株进行g418敏感度测定,确定筛选转化子所需g418的最适浓度。
(2)农杆菌介导转化ftrsy-2菌株
(a)将已转入载体的农杆菌在加有利福平(100μg/ml)、链霉素(100μg/ml)、卡那霉素(100μg/ml)的yeb固体平板上活化,28℃,黑暗培养2-3d;
(b)挑取单菌落到加有利福平(100μg/ml)、链霉素(100μg/ml)、卡那霉素(100μg/ml)的1mlyeb液体的1.5ml离心管中,28℃,200rpm培养过夜;
(c)第二天待菌液浑浊后,吸取100ul菌液至10ml加有利福平(100μg/ml)、链霉素(100μg/ml)、卡那霉素(100μg/ml)的yeb液体培养基中,28℃,200rpm避光培养至od600约为0.15。
(d)取5ml的菌液于10ml的离心管中,6000rpm离心5min,弃上清。再往离心管中加入im液体,调整菌液od600为0.15;
(e)用tween-20制备粉棒束孢孢子悬液,并稀释至1×106分生孢子/ml;
(f)取400μl农杆菌菌液、400μl孢子悬液混匀,加入200μlaim液体培养基,200rpm28℃黑暗诱导培养2d;
(g)吸取100μl农杆菌与孢子悬浮液混合液,涂布于铺有灭菌玻璃纸的aim固体平板上,28℃倒置黑暗培养3-4d;
(h)将玻璃纸正向转移到含有草丁膦与头孢霉素的czm平板培养基上,28℃倒置黑暗培养。
(3)粉棒束孢突变株的筛选及验证
当czm平板的玻璃纸上长出粉棒束孢菌落时,用牙签挑取,并接种到ppda平板培养基上,28℃培养;待菌落长大后,挑取少量菌丝,压片法在荧光显微镜下观察;选取带有绿色荧光的转化子,单孢分离并多次传代培养,确保egfp是稳定遗传的荧光标记;随机挑选10株突变株接种到无筛选药物的pda平板上,重复转接10代后,再接种ppda平板上,观察是否能够正常生长,以确定抗性基因能否稳定遗传。随机选取15个突变株,提取基因组dna,采用基于草丁膦抗性基因设计的特异性引物barf/r对突变株进行检测;随机挑取转化子在荧光显微镜下观察是否有绿色荧光。
(4)表型突变的转化子筛选及抗逆性测定
以野生ftrsy-2菌株为对照,根据菌落形态及颜色、孢子产量等特征,从突变体库中筛选表型差异较大的突变株。将表型差异较大的突变株,接种于含有筛选药物的ppda培养基上,28℃条件下培养7d,测定其菌落直径和产孢量,计算日平均生长速率及单位面积产孢量,每个处理3次重复,以野生菌株为对照,确定由于t-dna的插入对突变株生长和产孢的影响。
将ftrsy-2的转化子培养7d后,刮取菌丝孢子,用终浓度为0.05%吐温-20的ddh20制作孢子悬浮液,使其浓度为1×105分生孢子/ml,分别取1μl点接于ph11及分别含有刚果红(500μg/mg)、nacl(1mol/l)、山梨醇(1mol/l)的pda制成的十二孔板上,设立空白对照,28℃培养7d,观察其生长状态。设置紫外线照射1h处理及亚致死温度35℃培养7d,以观察转化子对紫外线及亚致死温度的敏感性。
实验结果:
利用bar基因引物barf/r微量快速pcr验证初筛敲除转化子,以pcambiamx9为阳性对照,野生菌株为阴性对照,结果表明野生菌株无扩增片段,而阳性质粒及随机挑选的阳性同源敲除突变子能够扩增出500bp左右的bar基因片段,证明bar基因已经成功插入,为进一步验证敲除转化子的准确性,同时采用原目的基因引物ifcvnf/r进行pcr扩增,结果表明野生菌株能扩到约360bp左右的目的基因片段,而突变株由于目的基因破坏的缘故,没有扩增出条带,因此表明ifcvn基因已经成功被敲除(图10)。采用原目的基因引物ifcvnf/r进行pcr扩增,结果表明野生菌株及回复菌株均能扩到约360bp左右的目的基因片段,表明ifcvn基因已经重新整合到基因组上,由于回复载体带有egfp基因盒,可使用倒置荧光显微镜于400-500nm波长激发光下进行观察,突变株菌丝及孢子形态如图11所示。
ifcvn基因对粉棒束孢生长和产孢的影响:结果如表1所示,粉棒束孢野生型、ifcvn基因同源敲除及回复转化子的生长速率差异显著,其中敲除株生长较慢,但三者之间产孢量无差异;逆境胁迫实验结果如图12所示,与野生对照相比,敲除株对刚果红及碱性环境表现敏感,菌丝生长速率减慢,对nacl、山梨醇等高渗条件胁迫不敏感,能够正常生长并产孢,回复株与野生株表现一致,而紫外线照射1h和35℃处理时野生株和敲除株及回复株的生长速度均受到抑制。
表1转化子的生长速率和产孢量
2.ifcvn基因敲除株毒力测定
(1)体表侵染
以0.02%吐温-80为溶剂,使用新鲜活化的粉棒束孢孢子配制浓度为1×107分生孢子/ml的wt、敲除菌株分生孢子悬液。实验对象为4龄大蜡螟幼虫,接种方式为体表浸渍,每个处理3次重复,每个重复20头,浸于事先准备好的分生孢子悬浮液25s后取出,滤纸吸干体表水分并放入150mm培养皿中,置于28℃恒温培养箱保湿培养,逐日观察统计幼虫死亡数,及时清理病死幼虫,避免二次感染,将死亡个体挑出保湿培养,连续观察7d。
(2)体内注射
以pbs为溶剂,使用新鲜活化的粉棒束孢孢子配制浓度为1×107分生孢子/ml的wt、敲除菌株分生孢子悬液。实验对象为4龄大蜡螟幼虫,接种方式为血腔注射,使用微量注射器从幼虫第三腹节气孔处注射孢悬液至幼虫血腔,接种量为10μl孢悬液/头虫(对照组为10μlpbs),每处理30头虫,每12小时观察取样。直至幼虫全部死亡(或停止死亡)时停止取样。
实验结果:
结果如表2所示。在采用浸渍法处理时,由于ifcvn基因的敲除,导致粉棒束孢对大蜡螟侵染能力增强,其校正死亡率由24.44%增加到33.33%,lt50值由26.53d降低到12.64d;采用注射法处理时与野生株无差异。由此表明ifcvn基因影响粉棒束孢的致病力,主要通过影响粉棒束孢对昆虫体壁吸附、穿透的过程来调控其毒力。
表2野生株与δifcvn菌株处理大蜡螟幼虫的校正死亡率及致死中时间(lt50)
3.ifcvn基因在感病大蜡螟组织中表达分析
配制浓度为1×107分生孢子/ml的wt菌株孢悬液,注射接种,28℃恒温培养箱保湿培养,接种0h、12h、24h、48h、72h之后取血淋巴、表皮、脂肪体及中肠组织,提rna后,反转cdna,进行rt-qpcr。
实验结果:
结果如图13所示:随侵染时间延长,ifcvn基因在大蜡螟幼虫各个组织中的表达量不同,在侵染后12h各组织中的表达量均达到最高,其中在血液中的表达量最高,在中肠中的表达量次之,在脂肪体中表达量较少,在表皮的表达量最低;随侵染时间延长,所有组织中ifcvn基因表达量均开始降低,其中在血液及脂肪体中的表达量迅速降低,而中肠中表达量的下降幅度较小,侵染后72h在表皮检测到少量ifcvn基因表达,而血液、中肠及脂肪体中ifcvn基因表达量均降到最低。
4.ifcvn基因原核表达及多克隆抗体制备
(1)原核表达载体sumo-pet28a-cvn构建
按步骤提取sumo-pet28a质粒,并以引物ifcvnf/r扩增ifcvn基因cdnaorf全长,命名为cvn。电泳验证后回收目的条带。
将提取的质粒sumo-pet28a和回收的ifcvn基因片段cvn进行bamhi/sali双酶切。电泳验证后回收目的条带。将回收的线性化质粒sumo-pet28a和ifcvn基因cdna回收片段cvn进行连接,转化大肠杆菌dh5α,用引物t7f/r对转化子进行菌液pcr验证;阳性菌落扩繁后提取质粒,用bamhi/sali对质粒进行双酶切验证后,转化大肠杆菌bl21(de3)备用。
(2)cvn蛋白诱导表达及表达形式鉴定
将含ifcvn基因的重组大肠杆菌bl21(de3)菌液在液体lb培养基中按1%接种量接种,37℃摇床振荡过夜培养至对数生长期,od600值为0.5-0.6范围时,加入终浓度为1mm/l的itpg诱导剂,再培养5-8h后离心收集菌体,ddh2o重悬后破碎,8000rpm离心10min,沉淀用5mlddh2o重悬。取10μl上清液及沉淀重悬液进行sds-page分析,考马斯亮蓝r-250染色4h后进行脱色,鉴定表达产物的存在形式。纯化蛋白并制备相应多克隆抗体。
(3)抗原及内源cvn蛋白westernblot检测
以制备的多克隆抗体(1:1000)为一抗、羊抗兔igg-hrp(1:5000)为二抗,采用western-blot方法分析多克隆抗体的特异性;分别提取粉棒束孢侵染大蜡螟不同时期各组织的总蛋白,检测感病大蜡螟组织中粉棒束孢cvn蛋白的表达量。
实验结果:
扩增获得ifcvn基因cdnaorf全长共369bp,pcr产物电泳鉴定大小正确(图14a),测序成功后克隆到sumo-pet28a载体上,酶切鉴定(图14b)后诱导表达。重组大肠杆菌bl21(de3)菌液在浓度达到od600值为0.5-0.6范围时,加入终浓度为1mm/l的itpg诱导剂,以空载体为对照,37℃诱导培养8h后,经sds-page分析,结果如图14c所示。由可以看出,目标蛋白大小大约为32kda左右,在上清液中检测到少量目标蛋白的表达,但大多数均以聚集的形式存在,即包涵体,说明在大肠杆菌bl21(de3)中cvn蛋白表达在包涵体和上清中。
western-blot方法分析抗ifcvn基因重组蛋白多克隆抗体的特异性,结果显示,重组蛋白能与多克隆抗体发生特异性血清学反应,在32kda处可见一明显的免疫反应条带,其大小与预测ifcvn基因重组蛋白分子量相近,同时抗体灵敏度高,1:1000稀释后可检测到500pg抗原(图15a)。以多克隆抗体检测粉棒束孢侵染后大蜡螟幼虫血液中的总蛋白,来表征ifcvn基因在感病大蜡螟组织中表达量(图15b),结果显示侵染后12h和24h可检测到cvn蛋白。
实施例5
cvn重组蛋白活性检测
(1)抑菌活性检测
采用生长抑制法进行cvn重组蛋白的体外抑菌活性检测(马驰,2014)。用双蒸水作空白对照,以氨苄青霉素(ampicillin)、链霉素(streptomycin)、头孢霉素(cefalothin)、硫酸卡那霉素(kanamycin)为阳性对照,对照药剂和重组蛋白均采用1×pbs稀释,使其终浓度为100μg/ml。在37℃的条件下,将水生拉恩氏菌(rahnellaaquatilis)、e.persicina、普城沙雷菌(serratiaplymuthica)、平凡假单胞菌(pseudomonastrivialis)、菠萝泛菌(pantoeaananatis)、土生柔特勒菌(raoultellaterrigena)、河生肠杆菌(lelliottiaamnigena)等从粉棒束孢生境及宿主虫尸上分离的供试菌株培养12h,再用lb培养基将其稀释至od600值为0.001;取稀释菌液加入96孔板中,加入供试药物,每个测试菌株三次重复;37℃培养12h后测量od600值,观察重组蛋白的抑菌活性。
实验结果:
抑菌活性实验结果见表3,与阳性对照相比,重组sumo-cvn在100μg/ml的使用浓度下,对p.ananatis的抑制作用最为明显,抑制率为11.05%;对e.persicina的抑制率为10.41%,稍强于氨苄青霉素阳性对照的抑制作用,表明其对e.persicina的效价与氨苄青霉素相当。
表3cvn重组蛋白对不同细菌的抑制率
(2)细胞毒性检测
采用wst-8试剂盒法测试重组cvn蛋白的细胞增值毒性检测,试验采用草地贪夜蛾细胞sf9细胞系,向96孔板各孔中加入100μl细胞悬液,28℃下预孵育培养板24h,待细胞贴壁后,加入10μl不同浓度的重组蛋白稀释液,使其终浓度依次为100μg/ml、10μg/ml、1μg/ml、100ng/ml、10ng/ml、1ng/ml、100pg/ml;28℃下孵育不同时间后,向各孔中加入10μl的cck-8溶液,继续28℃下孵育1-4h,用酶标仪测定450nm处的od值,分析重组蛋白的细胞毒性。
实验结果:
wst-8实验表明,在所测试的使用浓度及作用时间下,除在最高使用浓度100μg/ml下作用72h时,重组cvn蛋白影响草地贪夜蛾细胞sf9细胞正常增值外,其余各试验条件下,重组cvn蛋白对sf9细胞生长无影响,同时短时间内一定浓度的重组cvn蛋白能够促进sf9的增值,因此表明重组cvn蛋白对正常sf9细胞具有一定的保护效应(图16)。
最后说明的是,以上实施例仅用以说明本发明的技术方案而非限制,尽管参照较佳实施例对本发明进行了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的宗旨和范围,其均应涵盖在本发明的权利要求范围当中。
序列表
<110>太极集团有限公司;西南大学
<120>粉棒束孢cyanovirin-n基因、重组蛋白及应用
<160>4
<170>patentinversion3.5
<210>1
<211>369
<212>dna
<213>人工序列(artificialsequence)
<400>1
atgcgctggacaacaaacaccgtcgcggccgctctggcttttgctactctcggtagctgc60
gtccactacacagacagctgcgacgacaccaacctgtctggcaccacgctgtctggccac120
tgcggtgacaacaagggcaacagtccctacagcagcgttgacctggcccaaaaggttggc180
aacaactggggtgttttggcttggggcggtgtcaacttccagcagagttgctccgagatt240
gtatacaactcttggaacggtgttttgtccgccaagtgcggcaacggcggtggccgtgat300
gtccgcacagttctgaatctgaacaactacatttccaacaactttggcaaactagcattt360
gactcgtag369
<210>2
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>2
atatggatccatgcgctggacaacaaacac30
<210>3
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>3
atatgtcgacctacgagtcaaatgctagtt30
<210>4
<211>122
<212>prt
<213>人工序列(artificialsequence)
<400>4
metargtrpthrthrasnthrvalalaalaalaleualaphealathr
151015
leuglysercysvalhistyrthraspsercysaspaspthrasnleu
202530
serglythrthrleuserglyhiscysglyaspasnlysglyasnser
354045
protyrserservalaspleualaglnlysvalglyasnasntrpgly
505560
valleualatrpglyglyvalasnpheglnglnsercyssergluile
65707580
valtyrasnsertrpasnglyvalleuseralalyscysglyasngly
859095
glyglyargaspvalargthrvalleuasnleuasnasntyrileser
100105110
asnasnpheglylysleualapheaspser
115120
- 还没有人留言评论。精彩留言会获得点赞!