CsLYK基因及其编码蛋白在提高柑橘溃疡病抗性的应用的制作方法
本发明涉及分子生物学领域,具体涉及一种cslyk基因及其编码蛋白在提高柑橘溃疡病抗性的应用。
背景技术:
柑橘是我国南方第一大水果,也是全球重要的经济作物。然而,柑橘细菌性溃疡病(citrusbacterialcanker,cbc)却严重阻碍了柑橘产业的健康发展(胡军华等,2015)。柑橘溃疡病是由黄单胞杆菌柑橘致病变种(xanthomonascitrisubsp.citri,xcc)引起,它起源于印度、爪哇等地(胡军华等,2015)。我国福建、湖南、广东等柑橘主产区受柑橘溃疡病侵害较为严重。溃疡病菌主要侵染柑橘叶片、枝梢、果实等,其中苗木、幼树受害较为严重(何秀玲等,2007)。病树会出现落叶、枯梢、树势衰弱以及落果等现象,严重的影响了柑橘的产量和品质。受到柑橘溃疡病危害的芸香科植物有几十种,其中绝大部分为经济栽培品种。有研究发现甜橙最易感病,其次是酸橙和柚类(袁承东等,1997;李敏等,2013)。目前全世界已经有30多个国家和地区将柑橘细菌性溃疡病列为重要的植物检疫对象。
目前为控制柑橘溃疡病危害通常采取一种综合防治策略:其中以化学防治为主,生物防治为辅。但是由于化学防治措施对环境不友好,极易对环境造成污染,且需要投入大量的人力、物力,生物防治效果差,成本高,所以亟待通过培育抗病新品种来减少溃疡病带来的损失(陈力等,2008;朱雪梅等,2017)。在抗病育种方面一代代柑橘人进行了长期杂交育种,但由于杂交育种周期长使得育种效率低。随着分子生物学的兴起,人们开始从抗病基因工程方向对病原菌本身和植物的抗病防卫反应进行研究。基因工程手段也在抗溃疡病研究中进行了尝试(段敏杰等,2016;贾瑞瑞等,2017),获得了一些对溃疡病有抗性的转基因材料。例如陈善春等获得了对柑橘溃疡病有抗性的转柞蚕抗菌肽d基因的锦橙、新会橙、脐橙株系(陈善春等,1996)。外源基因nls,chit42,xa21和ptha在转移至冰糖橙,椪柑和甜橙后对柑橘溃疡病具有抗性(mendesetal,2010;yangetal,2011)。li的研究发现csbzip40是响应柑橘溃疡病侵染的一个重要的转录因子,并且猜测是通过sa途径影响柑橘品种的抗性(lietal.,2017)。cslob1基因作为溃疡病病原基因ptha的靶蛋白,被认为是是柑橘溃疡菌的感病因子,使得植物对溃疡病更为敏感(lietal,2014),peng等对柑橘溃疡病感病基因cslob1启动子进行crispr/cas9靶向敲除从而获得了对柑橘溃疡病抗性提高的植株。
细胞溶素基序(lysm)结构域存在于大多数动植物体中,其长度约为40个氨基酸(buistetal.,2008;zhangetal.,2009)。lysm类受体激酶(lysmreceptor-likekinase,lyk)是在植物中发现的一类重要的类受体激酶,在拟南芥中有4个,在水稻中有10个成员(shiuetal.,2004;lehti-shiuetal.,2009)。大多数植物lysm蛋白都与病原菌几丁质和肽聚糖识别有关,lyks在多种植物病害发生、与微生物共生过程中的重要作用已被证实:拟南芥中的lyks能够与几丁质和肽聚糖直接结合,促进atlyk1形成二聚体并将激活位于胞内的激酶结构域,从而将胞外信号传递到胞内,突变植株感病性明显增强(liuetal.,2012);水稻中的两个lyp(lyp4与lyp6)与pgn、几丁质均能结合,沉默二者中的任何一个都会削弱植株抗性(liuetal.,2012)。大麦、苜蓿、番茄等作物中的一些含有lysm结构域的蛋白的抗病功能也陆续被鉴定(tanakaetal.,2010;fliegmannetal.,2011;zengetal.,2011)由此可见,lyks可能是病原菌相关基因,参与了植物抵御病原侵害的过程。利用lyks基因进行抗病分子育种具有重大的潜力。目前,尚没有利用lyks提高柑橘对柑橘细菌性溃疡病的抗性的研究和应用。
技术实现要素:
本发明为提高柑橘对细菌性溃疡病的抗性提供一种新的选择,公开一种cslyk基因及其编码蛋白在提高柑橘溃疡病抗性的应用。该应用是将一个柑橘lysm类受体蛋白磷酸激酶编码基因通过表达载体整合到柑橘中,有效提高柑橘对细菌性溃疡病的抗性,这对于柑橘的抗细菌性溃疡病育种具有重大的应用价值。
发明人在柑橘抗感柑橘细菌性溃疡病后转录组中获得了一个在抗病品种中受柑橘溃疡病菌诱导上调的lyks基因,而感病品种则表达变化不大。因此,构建超量表达载体通过农杆菌介导转化柑橘研究转化柑橘对柑橘细菌性溃疡病的抗性。
本发明通过下述技术方案实现:
cslyk基因及其编码蛋白在提高柑橘溃疡病抗性的应用,所述cslyk基因的编码蛋白为柑橘lysm类受体蛋白磷酸激酶,命名为cslyk蛋白,cslyk基因的核苷酸序列为(a1)或(a2):
(a1)seqidno.1所示的核苷酸序列;
(a2)来源于柑橘且与(a1)具有95%以上同一性且编码所述cslyk蛋白的核苷酸序列。
进一步的,所述cslyk蛋白为(b1)或(b2)或(b3)或(b4):
(b1)氨基酸序列如seqidno.2所示的蛋白质;
(b2)在(b1)所述蛋白质的n端和/或c端连接标签得到的融合蛋白;
(b3)将(b1)经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与柑橘溃疡病抗病性相关的蛋白质;
(b4)来源于柑橘且与(b1)具有98%以上同一性且与柑橘溃疡病抗病性相关的蛋白质。
cslyk基因及其编码蛋白在提高柑橘溃疡病抗性的应用,包括以下步骤:
(1)克隆柑橘cslyk编码序列,然后进行超量表达载体构建;
(2)超量表达载体转化柑橘,得到溃疡病抗性提高的转基因植株。
其中,步骤(1)中,克隆柑橘cslyk基因编码序列所采用的pcr引物为oe-f和oe-r,分别具有如seqidno.3和seqidno.4所示核苷酸序列。
进一步的,步骤(1)中,柑橘cslyk基因编码序列的克隆方法为:提取柑橘总rna,然后反转录为cdna,最后采用高保真酶pcr扩增cslyk基因编码序列dna片段。
进一步的,步骤(1)中,超量表达载体构建方法为:以plgne为载体,plgne载体带有camv35s启动子,camv35s启动子为花菜花叶病毒启动子,具有seqidno.5所示的核苷酸序列,利用kpnⅰ和ecorⅰ酶切目的片段后连接到kpnⅰ和ecorⅰ酶切回收的载体上,构建超量表达载体plgne-lyk。
进一步的,步骤(2)中,超量表达载体转化柑橘的方法为:超量表达载体通过电击法转化根癌农杆菌,再用根癌农杆菌介导转化柑橘外植体,遗传转化后的外植体细胞经离体培养、染色鉴定、嫁接后得到转基因植株。
进一步的,步骤(2)得到转基因植株后,对转基因植株进行抗性评价,判定cslyk基因超量表达可提高柑橘对细菌性溃疡病的抗性。
进一步的,对转基因植株进行抗性评价前,通过pcr验证转基因植株,采用的引物为id-f和id-r,id-f是取自plgne载体上camv35s的一段序列,id-r是根据cslyk基因末端序列设计,分别具有如seqidno.6和seqidno.7所示核苷酸序列,阳性植株可以得到2574bp的扩增片段。
进一步的,pcr验证后,利用实时荧光定量pcr进行cslyk基因表达量检测,采用的引物为rt-f和rt-r,分别具有如seqidno.8和seqidno.9所示核苷酸序列,实时荧光定量pcr内参为柑橘actin基因,采用引物为ac-f和ac-r,分别具有如seqidno.10和seqidno.11所示核苷酸序列。
本发明与现有技术相比,具有如下的优点和有益效果:
本发明cslyk基因及其编码蛋白在提高柑橘溃疡病抗性的应用,通过克隆柑橘cslyk基因编码序列,进行超量表达载体构建,然后转化柑橘,得到的转基因植株溃疡病发病程度可最大降低至现有柑橘的40%,能够显著的减轻溃疡病的发病程度,减小病斑面积,通过超量表达cslyk基因可大大提高植株对细菌性溃疡病的抗性,这对于柑橘的抗细菌性溃疡病育种具有重大的应用价值。
附图说明
此处所说明的附图用来提供对本发明实施例的进一步理解,构成本申请的一部分,并不构成对本发明实施例的限定。在附图中:
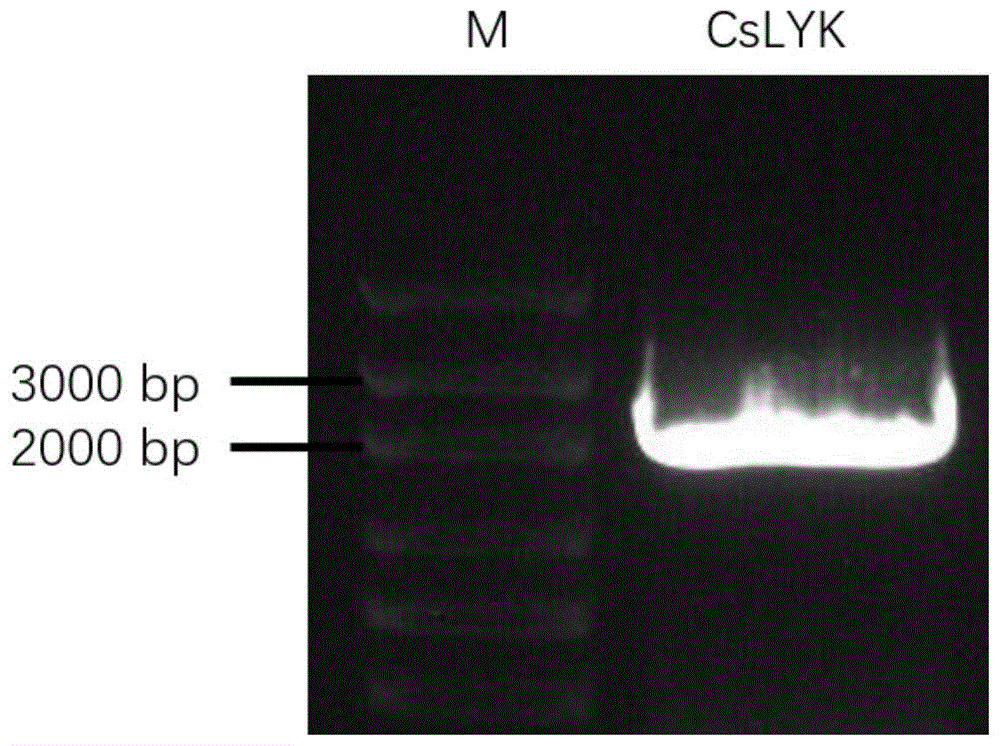
图1为本发明cslyk基因克隆的pcr扩增图谱:m为dna分子量标准。
图2为本发明cslyk蛋白的结构域示意图:sp,信号肽;lysm:细胞溶素基序结构域;tm,跨膜结构域;pkinase,磷酸激酶结构域。
图3为本发明cslyk植物超量表达载体构建流程图:gus,β-葡萄糖酸苷酶基因;camv35s,来源于花椰菜花叶病毒的植物组成性启动子;nos,冠瘿碱合成酶基因终止子;载体plgne具有camv35s启动子调控下的gus基因,便于在植物遗传转化过程中对转化子进行gus染色筛选。
图4为本发明cslyk基因在柑橘基因组中pcr重组验证:m为dna分子量标准;oe1、oe2、oe3分别代表3个转基因植株,wt为野生型晚锦橙植株,(下同)。
图5为本发明转基因植株gus染色图。
图6为本发明转基因植株中cslyk基因相对表达量分析图:**表示同野生型比较差异极显著(p=0.01),(下同)。
图7为本发明转基因植株表型图。
图8为本发明超量表达植株叶片接种溃疡病菌的10天后症状图。
图9为本发明转基因柑橘叶片病斑大小统计图。
图10为本发明转基因柑橘叶片发病程度(病情指数)统计图。
图11为本发明工艺流程图。
具体实施方式
为使本发明的目的、技术方案和优点更加清楚明白,下面结合实施例和附图,对本发明作进一步的详细说明,本发明的示意性实施方式及其说明仅用于解释本发明,并不作为对本发明的限定。
本发明实施例以晚锦橙为试验对象,在实际应用中,该方法也能够用于提高其他柑橘品种对溃疡病的抗性改良。
实施例1
柑橘cslyk基因编码序列的克隆
1.rna提取及cdna合成
用rna提取试剂盒(艾德莱,cat:rn09)提取柑橘(晚锦橙)叶片总rna,琼脂糖凝胶电泳验证rna质量,浓度计测定其浓度。使用recombinantdnasei合成cdna(takara),cdna于-20℃保存备用。
2.cslyk基因编码序列的pcr扩增
使用引物oe-f(seqidno.3)和oe-r(seqidno.4)从柑橘cdna中扩增获得cslyk片段(图1),片段长度为2054bp(包含酶切位点),扩增的dna片段经测序分析为柑橘cslyk基因编码序列,软件分析对应的氨基酸序列及结构域,验证蛋白质的正确性,其具有两个lysm结构域、1个激酶结构域、1段信号肽、1段跨膜结构域(图2)。
扩增体系:10xpcrmix:2.5μl;引物oe-f(5μmol/l):1μl;引物oe-r(5μmol/l):1μl;cdna约60ng;加ddh2o至25μl。
扩增程序:94℃,5min;94℃,30s,56℃,30s,72℃,1.5min,35个循环;72℃延伸10min。
3.dna片段回收
紫外灯下,用洁净的刀片切下含有目的片段的琼脂糖凝胶块。使用试剂盒(艾德莱)回收片段。
实施例2
超量表达载体的构建并转化农杆菌
载体构建流程图如图3,所有限制性内切酶购自(thermo)公司,按照使用说明操作。
具体操作如下:将cslyk基因片段和超表达载体plgne用限制性内切酶ecorⅰ和kpnⅰ双酶切后回收进行连接过夜,连接采用t4dnaligasekit(takara)。连接产物转化大肠杆菌dh5α,阳性克隆提取质粒即得到cslyk的超量表达载体plgne-lyk。质粒提取采用试剂盒(艾德莱)。
用电激法将构建的超量表达载体导入根癌农杆菌eha105。预先取冻存的eha105农杆菌感受态细胞(50μl),于冰上融化,加入2μl所构建的超量表达载体的质粒于感受态细胞中,吹打混匀后,冰上放置5min。将混合液转入事先吹干的电击杯底,将电击杯放入卡槽调整到正确位点。将电击装置调节为“agr”档,按下电击按钮,检查电击数据确保电击成功。加入1mllb液体培养基至电击杯中,移液枪吹打混匀,移至无菌离心管中,260r/min,28℃摇床振荡培养60min。10000r/min将菌液离心1min,弃上清液(剩约100μl重悬菌体),重悬后涂布,28℃倒置暗培养2天。待菌斑长出后,挑取单菌落进行pcr验证。
实施例3
遗传转化柑橘(晚锦橙)
1.柑橘实生苗上胚轴的获得
新鲜柑橘(晚锦橙)果实洗净,70%酒精表面消毒,无菌的条件下取出种子,剥掉种皮,种子萌发培养基上萌发,28℃下暗培养2周,然后在16h光照/8h黑暗条件下培养1周。无菌条件取萌发幼苗上胚轴切成1cm茎段,用于根癌农杆菌遗传转化。
2.根癌农杆菌菌液制备
用于转染的农杆菌菌液(含cslyk超量表达载体)加入30%的无菌甘油保存于-80℃的超低温培养箱中。转染前,在含50mg/l卡那霉素的lb固体培养基上划线培养。挑单菌落,接种于25ml含有相同抗生素的lb液体培养基中,28℃震荡培养过夜。测浓度后将菌液稀释成od=0.1的菌液进行二摇,约3h后,菌液od=0.5时,5000r/min离心10min,弃上清,用ph5.4的ms液体培养基重悬用于转染。
3.柑橘上胚轴茎段转化
将柑橘上胚轴茎段在农杆菌菌液中浸泡13min后擦干,将茎段转移到共培养培养基中,26℃暗培养2天。
4.转化子的筛选
共培养完成后,将上胚轴转移到筛选培养基中,28℃暗培养7天,外殖体在28℃,16h光照/8h黑暗培养,每两周继代一次,然后gus染色初步鉴定。
5.转化子的成苗培养
待幼苗长到1cm以上时,将其切下后嫁接到无菌试管中的晚锦橙苗,在成苗培养基中进行培养;待幼苗长到5cm左右时将其嫁接到枳实生苗上,在温室中进行培养。
实施例4
转基因植株验证
1.pcr检测外源基因整合
初筛得到的植株叶片100mg,使用dna提取试剂盒(艾德莱,cat:dn15)提取基因组dna,pcr检测cslyk基因在柑橘基因组的整合。pcr反应条件:94℃3min;94℃30s,58℃30s,72℃30s,30次循环;72℃10min。检测引物为id-f(seqidno.6)和id-r(seqidno.7)。pcr结果见图4,阳性植株可以得到2574bp的扩增片段wt植株无扩增。
2.转基因植株gus染色鉴定
转基因植株叶片切成叶圆片(直径7mm),进行gus组织化学染色(24h),如图5所示,阳性植株叶圆片边缘显蓝色,wt植株叶圆片未显色。
3.cslyk基因表达量分析
提取柑橘叶片总rna(艾德莱,catno:rn09),使用recombinantdnasei合成cdna。利用qrt-pcr检测目的基因的表达量,检测引物为rt-f(seqidno.8)和rt-r(seqidno.9);内参基因actin的检测引物为rt-f(seqidno.10)和rt-r(seqidno.11)。
反应体积20μl,反应条件:95℃3min,94℃10s;56℃10s,72℃10s,40次循环;72℃10min。实验重复三次。
采用2-△△ct法计算转基因植株中cslyk基因的相对表达量:定义水处理的样本为参照因子,即其cslyk的表达水平为1,然后计算转基因柑橘中相对参照因子基因表达的倍数2-△△ct,为其相对表达量。检测结果见图6。结果显示,cslyk基因在转基因植株中相比野生型植株有高水平表达(>50倍)。
4.转基因植株表型观察
观察分析3株转基因植株表型,发现外观和长势上并无明显异常(图7)。说明超量表达cslyk基因并未对植株的表型和发育产生直接的明显的变化。
实施例5
转基因植株的抗性评价
成熟叶片清洗后用75%的酒精消毒并用无菌水冲洗,置于超净台;以叶脉为中心进行针刺,六针为一组,每侧两组,用移液器点样溃疡病菌液,每针孔点样1μl(1x105cfu/ml)。于28℃恒温光照培养箱中培养(16h光照/8h黑暗)。叶片点菌后培养10天拍照,用imagejv1.47软件统计病斑面积。根据病情指数公式计算发病程度。按照病斑面积将病情分为0-7级,以字母r表示病斑面积,0级(r≤0.25mm2),1级(0.25mm2<r≤0.5mm2),2级(0.5mm2<r≤0.75mm2),3级(0.75mm2<r≤1mm2),4级(1.0mm2<r≤1.25mm2),5级(1.25mm2<r≤1.5mm2),6级(1.5mm2<r≤1.75mm2),7级(r>1.75mm2);根据公式计算发病程度:di=100xσ【各级病斑数x相应级数值】/(病斑总数x最大级数)。
超量表达植株和同时期嫁接的wt植株,选取发育一致的叶片针刺接种溃疡病菌。离体接种溃疡病菌10天后,接种溃疡病菌的植株均不同程度发病,病斑大小存在一定的差异(图8)。上述实验重复三次以确保结果的准确性。
对离体接种柑橘溃疡病菌的叶片的病斑大小进行统计。分析发现转基因植株病斑面积均显著小于转空载体对照组,其中oe1最小(图9)。转基因植株的发病程度统计发现转基因植株发病程度显著小于转空载体对照组,尤其是oe1,约为对照组的40%(图10)。
由此可见,cslyk的超量表达能够显著减小柑橘细菌性溃疡病的病斑面积,减轻柑橘细菌性溃疡病的发病程度。
本发明中,根癌农杆菌转化所用培养基:
种子萌发培养基:
ms+30g/l蔗糖+2.5g/lgelrite,ph5.8。
共培养培养基:
ms+2mg/lba+0.5mg/liaa+1mg/l2,4–d+100μmolas+30g/l蔗糖+2.5g/lgelrite,ph5.8。
筛选培养基:
ms+2mg/lba+0.5mg/liaa+500mg/lcef+50mg/lkan+30g/l蔗糖+2.5g/lgelrite,ph5.8。
成苗培养基:
ms+30g/l蔗糖,ph5.8。
以上所述的具体实施方式,对本发明的目的、技术方案和有益效果进行了进一步详细说明,所应理解的是,以上所述仅为本发明的具体实施方式而已,并不用于限定本发明的保护范围,凡在本发明的精神和原则之内,所做的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
序列表
<110>西南大学
<120>cslyk基因及其编码蛋白在提高柑橘溃疡病抗性的应用
<160>11
<170>siposequencelisting1.0
<210>1
<211>2037
<212>dna
<213>人工序列(1)
<400>1
atggagccaccatattcattgtgtacctacctttcaatctggttgatgattctgtgtaat60
tttcttcatgatttgcagcaatcaaactgccaacaaacttaccttggtaatgttagtcta120
gaatgcaacaacacccctgccatatcaaaaggctatctctgtaatggtcctcaaaagtta180
tgccagtccttcataaccttccgatctcaaccaccttacgacacacccgtcagtattgcg240
tatcttttaggttcagaagcctccagcataaccttgatcaataaaatttcatctactgat300
gagaaattgcctactgacaaattagtaatagttcctatttcgtgttcctgtgctgccagt360
atataccaacacagcactccttacaccatcaaggccaacgacacgtattttaagttagca420
cgctatacttaccagggcttgacaacttgccaggctctcctcggacagaactacttcgac480
gcaccaaatattactattggtgcgcaggtgatgatcccgctgagatgtgcttgtccgacc540
gcaaagcagatagacaatggagtgagctatttgttggcttatatggcaacaaagggtgat600
actatttcatcaattggacataaatttggagttgatcaacagagcattttggaggcaaac660
atgctgtcaaaagctgactcaattttcccttttgcacctcttttgattccactcaaaaac720
ggaagttgctctgcaaatcctgagaatttcttttgccattgtaagaatggtttccttgta780
gatggcaagctagagggtctccattgcaaacctgatggcaaaaagtttccggtcaaattg840
gttgctctcttaggtttggggattggtttgggatttttgagtgtggtggttgtggggtgc900
tatttgtacagattttttaaggataaaagaaatagaatgcttaaagagaagctgttcaag960
caaaatggaggctacttgctgcaacaacaactatcatcctgtggaagtagcgaaagagct1020
aaagtatttacagcagacgagcttcaaagagcaacagacaattacaaccagagccggttt1080
cttggccaaggaggatttggcactgtctacaaagggatgttacctgatggtagcatagtt1140
gctgtcaaaaggtcaaaagagattgataaaactcaaattcatcaattcattaatgaagtt1200
gtcattctctctcagatcaatcatcgacatattgtcaagcttctgggttgttgtttagag1260
actgaagtaccggtactagtatacgagtacatctccagtggaacactttcccaccacatt1320
catgatcaccagcaacagcaagaacagaaacaaaaacaagaattgtcttcactttcatgg1380
gagaatcgagttagggttgcgtgtgaagttgcaggagcagtagcatatatgcattcctca1440
gcctctatccctatctttcatcgcgacatcaaatcttccaacatactcttggatgataaa1500
tttagcgcaaaagtttctgattttggtatttcaaggtctataccgaacgataagactcat1560
ttaacaacaacaattcaaggaaccttcggatacttggatccagagtattttcaatcaagt1620
caattcacagataaaagcgatgtgtatagctttggggttgttctattagagcttctaacc1680
ggtaagaaaccaatctgttttgctagagtagaagaagaaagaaatttagttgcatgcttc1740
atttcattagcaaaggagaaccaactgcttgagattttagatgctagagtagctaaagaa1800
gcaagagaagaagatattggagctatggcagagcttgcaatgagatgtttgagattgaat1860
agcaaaaaaagaccgacgatgaaacaagtttcaatggagcttgaagggttgagaagatca1920
cagagatgcttagaaatgtgccaagtgaatcagttgttggcagatgagatttcattagca1980
gacaacttagtaatttccatgcaaatggattcaaaatcattttattcatcagcttga2037
<210>2
<211>678
<212>prt
<213>人工序列(2)
<400>2
metgluproprotyrserleucysthrtyrleuseriletrpleumet
151015
ileleucysasnpheleuhisaspleuglnglnserasncysglngln
202530
thrtyrleuglyasnvalserleuglucysasnasnthrproalaile
354045
serlysglytyrleucysasnglyproglnlysleucysglnserphe
505560
ilethrpheargserglnproprotyraspthrprovalserileala
65707580
tyrleuleuglyserglualaserserilethrleuileasnlysile
859095
serserthraspglulysleuprothrasplysleuvalilevalpro
100105110
ilesercyssercysalaalaseriletyrglnhisserthrprotyr
115120125
thrilelysalaasnaspthrtyrphelysleualaargtyrthrtyr
130135140
glnglyleuthrthrcysglnalaleuleuglyglnasntyrpheasp
145150155160
alaproasnilethrileglyalaglnvalmetileproleuargcys
165170175
alacysprothralalysglnileaspasnglyvalsertyrleuleu
180185190
alatyrmetalathrlysglyaspthrileserserileglyhislys
195200205
pheglyvalaspglnglnserileleuglualaasnmetleuserlys
210215220
alaaspserilepheprophealaproleuleuileproleulysasn
225230235240
glysercysseralaasnprogluasnphephecyshiscyslysasn
245250255
glypheleuvalaspglylysleugluglyleuhiscyslysproasp
260265270
glylyslyspheprovallysleuvalalaleuleuglyleuglyile
275280285
glyleuglypheleuservalvalvalvalglycystyrleutyrarg
290295300
phephelysasplysargasnargmetleulysglulysleuphelys
305310315320
glnasnglyglytyrleuleuglnglnglnleusersercysglyser
325330335
sergluargalalysvalphethralaaspgluleuglnargalathr
340345350
aspasntyrasnglnserargpheleuglyglnglyglypheglythr
355360365
valtyrlysglymetleuproaspglyserilevalalavallysarg
370375380
serlysgluileasplysthrglnilehisglnpheileasngluval
385390395400
valileleuserglnileasnhisarghisilevallysleuleugly
405410415
cyscysleugluthrgluvalprovalleuvaltyrglutyrileser
420425430
serglythrleuserhishisilehisasphisglnglnglnglnglu
435440445
glnlysglnlysglngluleuserserleusertrpgluasnargval
450455460
argvalalacysgluvalalaglyalavalalatyrmethisserser
465470475480
alaserileproilephehisargaspilelysserserasnileleu
485490495
leuaspasplyspheseralalysvalserasppheglyileserarg
500505510
serileproasnasplysthrhisleuthrthrthrileglnglythr
515520525
pheglytyrleuaspproglutyrpheglnserserglnphethrasp
530535540
lysseraspvaltyrserpheglyvalvalleuleugluleuleuthr
545550555560
glylyslysproilecysphealaargvalgluglugluargasnleu
565570575
valalacyspheileserleualalysgluasnglnleuleugluile
580585590
leuaspalaargvalalalysglualaargglugluaspileglyala
595600605
metalagluleualametargcysleuargleuasnserlyslysarg
610615620
prothrmetlysglnvalsermetgluleugluglyleuargargser
625630635640
glnargcysleuglumetcysglnvalasnglnleuleualaaspglu
645650655
ileserleualaaspasnleuvalilesermetglnmetaspserlys
660665670
serphetyrserserala
675
<210>3
<211>33
<212>dna
<213>人工序列(3)
<400>3
cggggtaccatggagccaccatattcattgtgt33
<210>4
<211>31
<212>dna
<213>人工序列(4)
<400>4
cggaattctcaagctgatgaataaaatgatt31
<210>5
<211>345
<212>dna
<213>人工序列(5)
<400>5
gagacttttcaacaaagggtaatatccggaaacctcctcggattccattgcccagctatc60
tgtcactttattgtgaagatagtggaaaaggaaggtggctcctacaaatgccatcattgc120
gataaaggaaaggccatcgttgaagatgcctctgccgacagtggtcccaaagatggaccc180
ccacccacgaggagcatcgtggaaaaagaagacgttccaaccacgtcttcaaagcaagtg240
gattgatgtgatatctccactgacgtaagggatgacgcacaatcccactatccttcgcaa300
gacccttcctctatataaggaagttcatttcatttggagagaaca345
<210>6
<211>20
<212>dna
<213>人工序列(6)
<400>6
cgacacgcttgtctactcca20
<210>7
<211>31
<212>dna
<213>人工序列(7)
<400>7
cggaattctcaagctgatgaataaaatgatt31
<210>8
<211>20
<212>dna
<213>人工序列(8)
<400>8
ttgacaacttgccaggctct20
<210>9
<211>20
<212>dna
<213>人工序列(9)
<400>9
ggtcggacaagcacatctca20
<210>10
<211>18
<212>dna
<213>人工序列(10)
<400>10
catccctcagcaccttcc18
<210>11
<211>19
<212>dna
<213>人工序列(11)
<400>11
ccaaccttagcacttctcc19
- 还没有人留言评论。精彩留言会获得点赞!