玉米基因ZmLIP2及其应用的制作方法
本发明涉及玉米基因zmlip2及其应用,属于植物基因工程领域。
背景技术:
玉米是世界范围内重要的粮食作物之一。然而随着生态环境的不断恶化,土壤中重金属的含量日益增加,其生长发育和产量品质受到包括重金属在内的非生物胁迫的严重影响,这严重制约了作物产量。挖掘抗逆相关基因,研究其逆境应答机制,培育抗逆性较强的玉米新品种是应对逆境胁迫,提高玉米产量的最有效途径。目前,我国玉米抗逆种质资源十分匮乏,常规育种得到抗逆种质资源不仅使一个非常艰难的过程,而且育种周期也比较长,很难培育出理想的抗性品种。随着现代分子生物学的飞速发展,为解决这个问题提供了一个很好的契机。
根据玉米对铅的积累特性和遗传规律,通过分子标记技术挖掘铅吸收和转运的相关基因是针对土壤铅污染的对策育种的基础研究和重要組成部分。目前,虽然己有关于不同玉米材料及不同组织对铅吸附特性和可能机制的研究报道,但对控制玉米重金属铅相关基因的定位研究尚未见报道
技术实现要素:
本发明克服了上述现有技术的不足,提供玉米基因zmlip2及其应用。本发明提供一个玉米基因zmlip2,明确了玉米zmlip2基因与植物耐铅胁迫的关系,验证了该基因在玉米耐铅应用中的重要性。
玉米基因zmlip2,其核苷酸序列如seqidno.1所示。
进一步的,上述玉米基因zmlip2的氨基酸序列为seqidno.2所示。
进一步的,上述玉米基因zmlip2定位于玉米第6号染色体chr06.156,292,425~chr06.156,294,916之间。
上述玉米基因zmlip2在提高植物耐铅能力方面的应用。
一种提高植物耐铅能力的方法,所述方法包括:下调玉米基因zmlip2在植物株系的表达。
进一步的,上述方法包括如下步骤:构建重组载体rnai-zmlip2,将rnai-zmlip2通过农杆菌介导转入玉米幼胚中,降低玉米基因zmlip2在植物株系中的表达量,提高植物耐铅能力。
上述玉米基因zmlip2在培育耐铅性增强的植物方面的应用。
一种培育耐铅性增强的植物的方法,包括如下步骤:构建重组载体rnai-zmlip2,将rnai-zmlip2通过农杆菌介导转入玉米幼胚中或拟南芥中,得到耐铅性增强的转基因植物,获得t0代植株,自交获得t1代植株,即可获得耐铅性增强的植物。
本申请还保护含有上述玉米基因zmlip2的重组载体、表达盒、转基因细胞系或重组菌。
有益效果:
(1)本发明研究了玉米基因zmlip2的功能,首次明确了玉米基因zmlip2与植物耐铅的关系,为利用该基因在提高植物耐铅性的应用奠定了理论依据。
(2)本发明利用农杆菌介导的转基因技术对玉米进行zmlip2基因的转化,并获得玉米zmlip2基因的rnai干涉转化事件。对rnai干扰的植株进行表达量分析发现,与受体材料相比,其表达量显著降低。对其进行铅胁迫发现,zmlip2基因的干涉株系与野生型相比在根系水平和叶片水平均增强了玉米在铅胁迫下的生长状况,增强了植株的耐铅性。
(3)本发明研究表明,玉米zmlip2基因能够负向调控植物对铅的吸收以及转运,首次明确了玉米zmlip2基因与植物耐铅胁迫的关系,验证了该基因在玉米耐铅应用中的重要性。为该基因在玉米及其他植物上的应用以及提高抗逆性提供了理论依据和实践价值。玉米zmlip2基因在植物抗重金属污染领域,特别是玉米抗土壤铅毒领域具有广阔的应用前景,其经济效益潜力巨大。
(4)本发明提出可利用转基因zmlip2来增强植物耐铅性,培育耐铅性植物,减少植株体内重金属铅的积累,降低植株受到的毒害作用。
附图说明
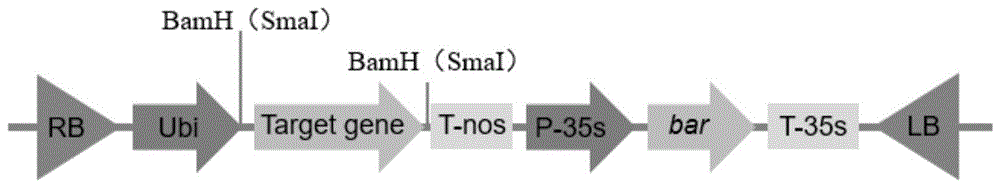
图1为rnai干涉株系所用的表达元件图谱。
图2为利用qrt-pcr技术对zmlip2基因的rnai株系的基因相对表达量。
图3为非处理对照和铅处理(3mmol/l)一周过后rnai干涉株系和非转基因阴性对照的表型照片。
图4为rnai干涉株系以及野生型对照在铅处理(3mmol/l)和非处理之后的植株表型数据。
图5为rnai干涉株系以及野生型对照在铅处理(3mmol/l)之后的植株铅含量。
具体实施方式
为了使本技术领域人员更好地理解本申请中的技术方案,下面结合实施例对本发明作进一步说明,所描述的实施例仅是本申请一部分实施例,而不是全部,本发明不受下述实施例的限制。
以下实施例中进一步定义本发明,根据以上描述和实施例,本领域技术人员可以确定本发明的基本特征,并且在不偏离本发明精神和范围的情况下,可以对本发明作出各种改变和修改,以使其适用各种用途和条件;其中本发明将上述表达载体导入植物细胞中,导入方法都是本领域人员熟知的,这些方法包括但不仅限于:农杆菌介导的转化法、基因枪法、电激法、子房注射法等。本发明所用选择标记基因为双丙氨酰磷抗性基因(bar),可进一步包括其它选择标记基因和报告基因。本发明选用的筛选抗生素为卡那霉素,除此之外,本发明所采用的均为本领域现有技术。
实施例1玉米自交系178根部的转录组分析
1-1、玉米自交系材料的铅胁迫处理
选择饱满的玉米自交系178种子用75%乙醇消毒1min,10℅的h2o2溶液浸泡15min并且消毒期间不断摇晃,然后用去离子水漂洗3~5次直到将残留的h2o2漂洗干净,再用去离子水浸泡4个小时,最后在28℃黑暗条件下用滤纸发芽。约2-3天后,将发芽的玉米转移到漂浮板上,在28℃光照(16h)/黑暗(8h)条件下用hoagland营养液培养进行水培,一周换两次培养液,在三叶期选取6株长势一致的幼苗进行胁迫处理。将这6株幼苗去除胚乳后移入装有营养液的塑料容器中。本实验设置2个处理:正常水分供给(即全营养液,ck)和铅胁迫处理(即全营养液+3mmol/lpb(no3)2,t)。营养液为现用现配,调其ph为6.0左右。分别取胁迫0h,24h,48h和72h之后的根系提取rna,进行转录组测序。
1-2、玉米总rna的提取
rna提取参照trizol试剂盒中的方法,具体步骤如下:(1)用液氮冷却研磨器:将已称量好的材料迅速放入研钵中并快速研磨,直至材料研磨成极细粉末;(2)按照每0.1克材料1ml的用量,将trizol加入研钵中;(3)20分钟后,拆开研钵继续研磨,直至研钵内trizol呈透明状,将其分装到2ml离心管中;(4)在上述离心管中再加入氯仿300μl,颠倒1分钟充分混匀,并静置5分钟后离心15min(4℃,12000rpm),小心吸取上清液至另一2ml离心管中;(5)重复第4步加氯仿及之后步骤,再次吸取上清液转至另一1.5ml离心管中;(6)再次吸取上清后,使用北京天恩泽基因科技有限公司的trizol伴侣提取总rna。(7)待rna沉淀晾干后,加入适量depc处理水溶解。
1-3、cdna的获得(利用takara公司的
(1)基因组dna的除去反应(totalrna用量:最大1μl)
(2)反转录反应,反应体系如下所示
1-4、zmlip2基因的时空表达模式分析
以1-3处理样品的cdna为模板,gapdh作内参基因,通过荧光定量pcr技术对zmlip2基因在铅胁迫条件下的表达进行相对定量分析。zmlip2基因其核苷酸序列如序列3所示,其编码蛋白氨基酸序列如序列4所示。
其中反应体系如下:
表3qrt-pcr扩增体系
反应程序为:95℃30s;95℃5s,tm(引物最适退火温度)30s,40cycles;95℃10s;65℃5s,65℃到95℃绘制融解曲线。所用基因定量pcr引物序列如下表所示:
表4qrt-pcr引物
对胁迫下的玉米根系进行转录组测序后发现,与对照条件相比,zmlip2基因是玉米根系响应铅胁迫最显著上调表达的基因。
实施例2.zmlip2干涉株系的获得及分析
本实施例以ubi作启动子,nos做终止子,bar作为选择标记基因,以卡那霉素作为选择标记基因,根据zmlip2序列设计目的基因片段引物,以cub载体为骨架,采用同源重组法,利用bamhⅰ和smalⅰ双酶切构建zmlip2基因的干涉载体。将构建好的干涉载体转化农杆菌感受态细胞。农杆菌浸染野生型玉米c01幼胚培养得到t0代zmlip2基因干涉玉米。
以下为具体步骤:
2-1、玉米rna的提取和纯化
利用trizol法提取玉米自交系齐c01(以下也称为野生型玉米)的总rna,具体方法参考1-2。cdna第一链的合成参考1-3。
2-2、zmlip2基因的克隆
以反转录的cdna为模板,采用以下引物对进行pcr扩增:
上游引物:5'-acggatcccactagtctggtcacgctcccc-3',如seqidno.5所示;
下游引物:5'-atcccggggtataaattgtaatgaccatac-3',如seqidno.6所示。
其中上游引物划横线部分为bamhⅰ酶切位点,下游引物划横线部分为smalⅰ酶切位点;pcr扩增体系为2μl上游引物(5μmol/μl),2μl下游引物(5μmol/μl),5μl5×phusionhfbuffer(gc),5μldntp混合液(2.5mmol/l),0.25μlphusiondnapolymerase,1μlcdna模板,加ddh2o将总体积补充至11.75μl;扩增条件为:98℃预变性30秒;98℃变性10秒,59℃退火25秒,72℃延伸1分40秒,循环30次;72℃延伸10分钟。取4μlpcr产物与pmd19-tsimple载体连接,操作步骤按照takara公司产品pmd19-tsimplevector说明书进行。然后连接产物转化大肠杆菌dh5α感受态细胞,在含有氨苄青霉素(100mg/l)的lb固体培养基上培养过夜。挑取白色菌落,在含有氨苄青霉素(100mg/l)的lb液体培养基中培养过夜。碱法提取质粒dna,酶切鉴定后进行序列测定。
2-3、植物重组载体rnai-zmlip2的构建
提取玉米自交系齐c01的rna,反转录得到cdna作为模板,用上游引物和下游引物进行pcr扩增,得到1775bp的pcr扩增产物(zmlip2基因)。将其反向互补后的片段用限制性内切酶bamhi和smalⅰ酶双酶切,再与用bamhi和smalⅰ双酶切pcub::ubi表达载体大片段连接,操作步骤按照fermentas公司产品t4dnaligase说明书进行。然后连接产物转化大肠杆菌dh5α感受态细胞,在含有壮观霉素(50mg/l)的lb固体培养基上培养过夜。挑取白色菌落,在含有壮观霉素(50mg/l)的lb液体培养基中培养过夜。碱法提取质粒dna并进行bamhi和smalⅰ酶切鉴定,得到大小为1775bp的基因片段为阳性质粒。将该阳性质粒命名为rnai-zmlip2,载体部分结构示意图如图1所示,图中targetgene区为转基因插入目标片段的位置。将rnai-zmlip2转化农杆菌lba4404感受态细胞,并获得可供转化使用的农杆菌菌株,命名lba4404/rnai-zmlip2。
2-4、农杆菌介导的玉米幼胚转化及抗性植株的获得
浸染前一天,挑取活化的农杆菌菌落lba4404/rnai-zmlip2(携带重组质粒的农杆菌单菌落)接种于含有50mg/l利福平和50mg/l壮观霉素的液体yep培养基中,28℃220rpm过夜振荡培养;将上述农杆菌溶液装入80ml离心管中,6000rpm离心5分钟,弃去上清,收集沉淀并用ab诱导培养液重悬,28℃摇床上培养至少5小时后方可进行沉淀、浸染。挑取授粉后10-12天的野生型玉米(玉米自交系c01)幼胚放入预浸染培养液中,2700rpm离心5秒,弃去培养基;加预浸染培养基2ml重新悬浮,2700rpm离心5秒;46℃水浴3分钟,冰浴1分钟,2700rpm离心5秒,弃去培养基,加入2ml预浸染培养液,4℃14000rpm离心10分钟,弃去培养液;将经过ab诱导培养基培养的菌液移入80ml离心管中,6000rpm离心5分钟收集菌体,用浸染培养液将菌体重新悬浮;将1ml菌液加入装有幼胚的离心管中,轻柔混匀数次,室温静置5分钟,弃去菌液;将幼胚移至共培养培养基上,28℃暗培养一周后,转移至筛选培养基,每两周继代一次;筛选两次后转移至分化培养基,将分化得到的幼苗转入生根培养基,当幼苗生长出3根以上粗壮的根后,进行炼苗;炼苗7天后,移至温室种植,得到t0代干涉zmlip2玉米。本实施例中的预侵染培养基为添加了1.5mg/l2,4-d的ms培养基(murashigeandskoog,1962),筛选培养基为添加了2.0mg/l2,4-d的n6培养基,其中的2,4-d为2,4-二氯苯氧乙酸。
2-5、干涉zmlip2基因玉米的分子鉴定
采用ctab法(sambrookandrussell,分子克隆实验指南,2001)提取t0代干涉zmlip2玉米植株基因组dna,分别设计引物检测标记基因bar和ubi-lip2融合片段,引物对序列如下:
标记基因bar基因引物对如下:
上游引物:5'-gtggagaggctattcggctatgactg-3',如seqidno.7所示;
下游引物:5'-agctcttcagcaatatcacgggtagc-3',如seqidno.8所示。
标记基因bar扩增序列长度为650bp,扩增条件为:94℃预变性5分钟;94℃变性30秒,59℃退火30秒,72℃延伸40秒,循环35次;72℃延伸10分钟。ubi-lip2融合片段引物对(上游引物为表达载体中启动子区序列,下游引物为zmlip2基因序列)如下:
上游引物:5'-gccttcatacgctatttatttgct-3',如seqidno.9所示;
下游引物:5'-ggctttacactttatgcttccg-3',如seqidno.10所示。
目标片段扩增序列长度为550bp,扩增条件:扩增条件为:94℃预变性5分钟;94℃变性30秒,61℃退火30秒,72℃延伸40秒,循环35次;72℃延伸10分钟。
采用同样的方法将空载体pcub::ubi转入野生型玉米中,得到t0代转pcub::ubi玉米。采用同样的方法检测,其结果与野生型玉米无显著差异。将t0代植株播种,自交得到t1代植株。
2-6、对阳性转基因干涉植株进行铅胁迫,并分析其表型
选择饱满的t1代zmlip2干涉株系种子采用1-1的方法对其进行处理,在铅处理14天之后,分别对各个材料取样,进行rna提取、表型鉴定和铅含量测定。其中rna提取方法参照1-2,样品的消煮用mas6,铅含量的测定用电感耦合等离子质谱仪。
对铅处理后野生型、干涉株系的阳性转基因植株进行表达量分析,结果如图2所示,其中wt表示野生型对照,rnai-1、rnai-2表示干涉株系的两个转化事件,可以看出干涉株系的表达量显著下降。铅处理前后的野生型对照、干涉株系的表型图片如图3所示,图3中从左往右分别是非铅处理组的非转基因阴性对照和干涉株系以及铅处理组的非转基因阴性对照和干涉株系。从图中可以看出,在正常生长条件下,干涉株系与非转基因阴性对照长势没有明显的区别,其根长、叶片大小以及株高都没有明显差异。而在铅处理下,干涉株系植株地上部分叶片比野生型长,根系分支比野生型多,其株高与根长较野生型长。说明干涉株系植株能降低玉米植株受铅胁迫毒害,使植株能够正常生长。由此可以看出,zmlip2基因具有负向调控的作用。其干涉植株能够显著增强玉米苗期对铅胁迫的抗性,且受到铅胁迫的诱导。如图4,图4中的a、b、c分别为株高、叶鲜重、根鲜重的生长情况。根据图4也可以看出,在非铅处理时,野生型、干涉株系的株高、叶鲜重、根鲜重均没有显著差异。而在铅处理时,干涉株系受到的铅胁迫伤害要显著低于野生型。这也说明了该基因的负向调控作用。对苗期地上部分和根部的铅含量进行测定,结果如图5所示,从图中可以看出,不论地上部分还是根部,rnai干涉株系的铅含量普遍低于野生型植株(wt)。联合以上表达量、表型照片和铅含量结果可以得出,zmlip2基因干涉株系可以减少植株体内的重金属铅的积累,从而降低植株受到的毒害。这进一步说明了zmlip2基因在玉米对铅胁迫中的负向调控的重要作用。
虽然,上文已经用一般性说明、具体实施方式及试验,对本发明作了详尽的描述,但在本发明基础上,可以对之做出一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
sequencelisting
<110>四川农业大学
<120>玉米基因zmlip2及其应用
<130>2020
<160>10
<170>patentinversion3.3
<210>1
<211>2492
<212>dna
<213>序列
<400>1
ccatgtataaattgtaatgaccatacgaaaaacaaattaaagtgcaaaatgtgtaaccca60
tcagtatgtcatagctcacatttttacatgcttggcgtcatcacaaaaaaaaagattact120
cacttgtgaacaccccaatttaactgccacaaccaccaagtccaagtggctccaaggtcc180
cacatattgagctccgtacacgccaagtgcacgaagccacttcatctgcgtggcccttcc240
acgtttgaagccttcgaagctgaacacacccgcccttatcattgctcctcggtttcctcc300
tcctcgttatcttcgtcgtcgtctgaagaagaagatgacgaggagtctgaatggtccgat360
tctttgtcttccttcggctccggggcggcggcaggcggtggagcggcaggggccggtggg420
tcaggcggcatcgccgtgaggaagcctagcgtcatgacgacgtcggccatcagcggtcgc480
acggagtcctcttcctgcaagcacatcgcagccatggcgaccacctggttcaacgctttg540
gcagggtattccttctttatgagtggatcaaccagctcatggtatctcttctggtccttg600
aacatcggcatggcctgaaaatgtgtcgtgcagatgcatgccatgccatgccatgccaat660
ataatgcagctcattagcaaaagcaacgttgaagaagcagcaaaaacgcgtagcttataa720
tacttacccaggtgacaacgttctgctcgtccaccggcttgctggtatcgacggccctcc780
tcccggagatgagctgcaccagcaccaccccgaagctgtacgcgtcggacttcatggtgg840
cctgccctgtgcgatcgtactccggcgcgcagcagccgaaggagcccatcatgggcgacg900
ccaccaccggcatgctgcccccggcctggccgagctgcgcgagcccgaagtcggagagct960
tgggcgtgagcttctcgtcgagaagtatgtgggaggccttgaattccccgtacaccaccg1020
gcgggttcgctgtctcgtgcaggtactccagcccctgcgcggcgccgtgcgccaccttca1080
tcctcgtgcaccagtccatcggcttcttgtccgccggcagatctggcgcgcgccggcagc1140
ttgttcagcagagacatagacgacgaaactgacaaattctgcaggagcttgccagttgcc1200
agataccggcgtgcacgtacgtacgtaccgaacaggtggtcttctaaggtgccggcgggc1260
acggactcgtacaccagcagccgctggtctccgtcagcgcagtagccaatgatgtcgacg1320
aggttctcgtggtggagctggctgagcttggcgaccccggtcaagaacgccttgttgtcc1380
tggaacccatgcttgtccagctgcttgatggccacggtctggagcaaaagtagcagaggc1440
acgccatgcatcacacatttcattcagagagacgcatggacgacgacacgtgaggcgtgg1500
cagttcacgtacgtacctgcccgctcttctctaaccggcccttgtagacccggaagaagc1560
cgccttcaccgacgaggttgtacggcgtgaagtggtctgtggcggcggcgagctcgcgga1620
acgcgaacgcctgccccgtgatggcgagccgcagcgccgagtcatcggcggaagcctcgt1680
gcgcctggttccctgcatcgcgagatcaaatcaaattcgttgggcgttgcaggcgcgacg1740
agatttggacgtgctaagcgagatgcatgaagaaaccgatcgaagcagagagggccatct1800
caccgccaccgggcttgggcggcgccgcggccgttggagcggcggcgggagctgggaagt1860
acgcggcgggcgcctgcaccatcgcgggtggcgtcatgttagacgccggggccgcgtcgc1920
ccgggctctccgcgtcggcgtcgtcgttgttgccctttccgccgccgaagcacggaaagc1980
agctcatcttccttttccctctgcgcctccccgtggcggcgaccctgtggacgcggggcg2040
agcggcgtgggcgcgagcgagcgacgagccgatgactcgacaggggcaagaatgctgggc2100
acgcacgcacgcacggacggaccgagccgccggccgccgtcactgcgcgccgaccatgcc2160
ccggcgtctcagcatcgacgcggcgtctccacttcgtggaacgccatgcccgagctcgcc2220
gcacgcgcaccaaatgaaacccgcccgtccggtgcggagcaggggagcgtgaccagacta2280
gtggtggcttgctcaggagaccggagaggggcacaagaccaccacgagacgagaacgacg2340
gtcgatggctatggcaatggaaggcaggccttggagatcgagagatccaagggccttccc2400
agagggaggggagaccggaggaggacaccgataccggtggaggggaggcgggaggagagg2460
gatgctaagtttggcgcgtggagtgaatcgag2492
<210>2
<211>426
<212>prt
<213>序列
<400>2
metsercyspheprocyspheglyglyglylysglyasnasnaspasp
151015
alaaspalagluserproglyaspalaalaproalaserasnmetthr
202530
proproalametvalglnalaproalaalatyrpheproalaproala
354045
alaalaprothralaalaalaproprolysproglyglyglyasngln
505560
alahisglualaseralaaspaspseralaleuargleualailethr
65707580
glyglnalaphealaphearggluleualaalaalathrasphisphe
859095
thrprotyrasnleuvalglygluglyglyphepheargvaltyrlys
100105110
glyargleuglulysserglyglnthrvalalailelysglnleuasp
115120125
lyshisglypheglnaspasnlysalapheleuthrglyvalalalys
130135140
leuserglnleuhishisgluasnleuvalaspileileglytyrcys
145150155160
alaaspglyaspglnargleuleuvaltyrgluservalproalagly
165170175
thrleugluasphisleupheaspleuproalaasplyslyspromet
180185190
asptrpcysthrargmetlysvalalahisglyalaalaglnglyleu
195200205
glutyrleuhisgluthralaasnproprovalvaltyrglygluphe
210215220
lysalaserhisileleuleuaspglulysleuthrprolysleuser
225230235240
asppheglyleualaglnleuglyglnalaglyglysermetproval
245250255
valalaserprometmetglyserpheglycyscysalaproglutyr
260265270
aspargthrglyglnalathrmetlysseraspalatyrserphegly
275280285
valvalleuvalglnleuileserglyargargalavalaspthrser
290295300
lysprovalaspgluglnasnvalvalthrtrpalametprometphe
305310315320
lysaspglnlysargtyrhisgluleuvalaspproleuilelyslys
325330335
glutyrproalalysalaleuasnglnvalvalalametalaalamet
340345350
cysleuglnglugluaspservalargproleumetalaaspvalval
355360365
metthrleuglypheleuthralametproproaspproproalapro
370375380
alaalaproproproalaalaalaprogluprolysgluasplysglu
385390395400
serasphisseraspserserserserserserseraspaspaspglu
405410415
aspasnglugluglugluthrgluglugln
420425
<210>3
<211>20
<212>dna
<213>人工序列
<400>3
gacaagaagccgatggactg20
<210>4
<211>20
<212>dna
<213>人工序列
<400>4
gccttgaattccccgtacac20
<210>5
<211>30
<212>dna
<213>人工序列
<400>5
acggatcccactagtctggtcacgctcccc30
<210>6
<211>30
<212>dna
<213>人工序列
<400>6
atcccggggtataaattgtaatgaccatac30
<210>7
<211>26
<212>dna
<213>人工序列
<400>7
gtggagaggctattcggctatgactg26
<210>8
<211>26
<212>dna
<213>人工序列
<400>8
agctcttcagcaatatcacgggtagc26
<210>9
<211>24
<212>dna
<213>人工序列
<400>9
gccttcatacgctatttatttgct24
<210>10
<211>22
<212>dna
<213>人工序列
<400>10
ggctttacactttatgcttccg22
- 还没有人留言评论。精彩留言会获得点赞!