一组B7H3单克隆抗体及其医药用途的制作方法
一组b7h3单克隆抗体及其医药用途
技术领域
1.本发明属于肿瘤抗体治疗以及分子免疫学治疗领域,涉及b7h3抗体及其用途。具体地,本发明涉及到多种b7h3单克隆抗体。
技术背景
2.b7h3也被称为cd276,是一种i型跨膜蛋白(chapoval a.i.,et al.,(2001),nat. immunol.2:269)。在小鼠中,b7h3由9号染色体编码;而在人类中,由15号染色体编码。b7h3与其它b7家族配体具有20
‑
27%的氨基酸同源性(sun m.,et al.,(2002),j. immunol.168:6294;loos m.,(2010),clin.dev.immunol.2010:683875)。在蛋白质组成、结构方面,鼠类的b7h3仅具2个igb7h3类型(由细胞外的一对igv和igc组成);而人类的b7h3具4个igb7h3类型,几乎是完全相同的igv
‑
igc串联性重复(sun m.,et al., (2002),j.immuno1.168:6294;steinberger p.,et al.,(2004),j.immunol.172:2352)。虽然b7h3 的mrna存在于一些正常组织中,如肝脏、小肠、胰腺、睾丸、心脏和结肠等组织,b7h3 蛋白在正常组织中相对少见(greenwald r.j.,et al.,(2005),annu.rev.immunol.23:515
‑ꢀ
48)。b7h3 mrna和蛋白质之间的这种差异性也许反映了转录后紧密调控机制的存在 (hofmeyer k.a.,et al.,(2008),proc.natl.acad.sci.105:10277;calabro l.,et al.,(2011),j. cell physiol.226:2595)。总体上,b7h3仅存在于一些非免疫性成纤维细胞、内皮细胞和成骨细胞,以及一些免疫细胞,如b细胞,t细胞,单核细胞,树突状细胞或nk细胞; b7h3的表达可由粒细胞
‑
巨噬细胞的csf或脂多糖刺激诱导产生(suh w.k.,et al.,(2003), nat.immunol.4∶899;chapoval a.i.,et al.,(2001),nat.immunol.2:269;greenwald r.j.et al., (2005),annu.rev.immunol.23:515)。
3.b7h3属于重要的免疫检查点蛋白。虽然初步研究曾显示b7h3对t细胞有刺激作用,可在一定程度上促进cd4+和cd8+t细胞的增殖、增强t淋巴细胞的细胞毒性及刺激干扰素
‑
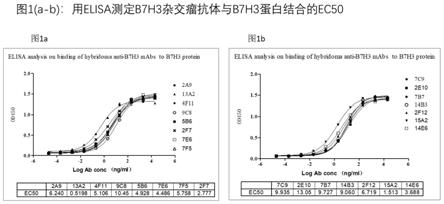
γ(ifn
‑
γ)的产生(chapoval a.i.,et al.,(2001),nat.immunol.2:269),并且b7h3 的缺失可减轻同种异体心脏移植的慢性排斥作用(wang l.et al.,(2005),eur.j.immunol. 35:428)。然而,更多的研究表明,b7h3有着显著的免疫抑制作用。例如,b7h3可显著抑制cd3抗体或同种异体dc细胞对t细胞的激活作用,而b7h3阻断抗体可有效逆转这种抑制作用(prasad d.v.r.,et al.,(2004),j.immunol.173:2500)。b7h3蛋白的结构信息揭示了b7h3 igv域中的fg环可能对t细胞抑制起着关键作用(vigdorovich v.,et al., (2013),structure 21:707)。另外除了t细胞,b7h3可能还对天然杀伤细胞(nk)有着抑制作用(castriconi r.,et al.,(2004),proc.natl.acad.sci.101:12640)。可能的机理涉及调节性t细胞(tregs)诱发树突状细胞(dc)b7h3的表达,以致于抑制dc细胞对t淋巴细胞的刺激作用(mahnke k.,et al.,(2007),eur.j.immunol.37:2117)。在小鼠模型中,缺乏 b7h3的小鼠会出现较严重的气道炎症,并且也会使某些自体免疫性疾病如实验性自身免疫应变性脑脊髓炎等更早地发生(suh w.k.,et al.,(2003),nat.immunol.4:899)。
4.在许多恶性肿瘤中,b7h3的表达水平相当高,例如黑色素瘤(wang j.,et al.,
(2013),j. invest.dermatol.133:2050)、白血病(hu y.,et al.,(2015),hematology 20:187;sun j.,et al., (2014),oncotargets ther.7:1979)、前列腺癌(zang x.,et al.,(2007),proc.natl.acad.sci. 104:19458)、卵巢癌(zang x.,et al.,(2010),mod.pathol.23:1104)和胰腺癌(chen y.,etal.,(2014),onco.targets ther.7:1465
‑
72)等。虽然正常和肿瘤组织中都有b7h3 mrna的存在,但b7h3蛋白仅在肿瘤细胞中有较高水平;这种差异可能与mirna
‑
29调控机制有关,如在正常或肿瘤组织以及癌细胞系中,有研究表明mirna
‑
29与b7h3蛋白水平呈现了反比关系(xu h.,et al.,(2009),cancer res.69:6275)。在肿瘤病人中,b7h3的异常表达与不良预后、肿瘤分级水平和转移程度增加、治疗抗性、肿瘤病人总体存活率下降有着比较密切的关系(picarda e.,et al.,(2016),clin.cancer res.22:3425)。
5.b7h3对t细胞、nk和dc细胞的抑制作用会显著促进肿瘤细胞的免疫逃逸;另外, b7h3对肿瘤细胞的增殖、迁移、侵袭、血管生成、上皮到间质转化(emt)、癌干性和 warburg效应、以及肿瘤细胞耐药性等也有着重要作用。b7h3的增加可促进bcl
‑
2和bcl
‑ꢀ
x1的表达,通过jak2
‑
stat3信号途径的激活从而增强肿瘤细胞抗细胞凋亡的活性(zhangt.,et al.,(2015),world j.gastroenterol.21:1804)。同时,akt、erk、以及jak2/stat3 途径的激活可诱导与肿瘤细胞转移有关的蛋白因子表达,包括mmp2、mmp9和cxcr4 等,从而增强肿瘤细胞的迁移力和侵袭力(tekle c.,et al.,(2013),int.j cancer 130:2282;liy.,et al.,(2017),oncotarget 8∶71725;wang l.,et al.,(2013),plos one 8:e70689;liu f.,et al., (2015),mol.med.rep.12:5455)。多项研究表明,肿瘤内皮细胞的b7h3也可基于tlr4依赖性机制诱导nf
‑
κb的活化,使得vegf和il
‑
8的表达水平显著增加(tekle c.,et al., (2012),int.j.cancer 130:2282),从而促进肿瘤的侵袭作用和血管生成(ferrara n.,et al., (2002),nat.rev.cancer 2:795)。另一方面,b7h3也可通过降低e
‑
钙粘蛋白以及增加n
‑ꢀ
钙粘蛋白、波形蛋白、cd133以及cd44的表达,起到调节肿瘤细胞emt和肿瘤干细胞活性的作用(jiang b.,et al.,(2016),oncotarget 731755)。由于b7h3可激活stat3信号传导,从而增加己糖激酶2的表达,所以b7h3也有促进肿瘤细胞有氧糖酵解的作用(shi t., et al.,(2019),cell death dis.10∶308)。同时,b7h3通过抑制转录因子nrf2的活性,致使活性氧(ros)和hif1a水平升高,从而进一步增强肿瘤细胞的有氧糖酵解作用,促进肿瘤细胞的生长(lim s.,et al.,(2016),cancerres.76∶2231)。越来越多的研究表明,抑制或降低b7h3的表达可增加肿瘤细胞对一些药物的反应,如抑制dna复制的药物(包括烷基化药物)以及抑制pi3k/akt/mtor和ras/raf/mek信号蛋白的药物(flem
‑
karlsen k., et al.,(2017),pigment cell melanoma res.30∶467;kasten b.b.,et al.,(2017),nucl.med.biol. 47:23;liu h.,et al.,(2011),mol.cancer ther.10:960)。
6.把肿瘤细胞移植在缺失b7h3的小鼠中或对肿瘤小鼠进行b7h3抗体处理,肿瘤的生长会受到明显的抑制(cai d.,et al.,(2020),cell.mol.immunol.17:227;lee y.h.,et al.,(2017), cell res.27:1034),说明阻断b7h3的信号传导可用于肿瘤治疗。此外,通过与抗pd
‑
1抗体的联用,产生对b7h3和pd
‑
1的双重阻断,产生协同性的抗肿瘤效果(lee y.h.,etal.,(2017),cell res.27:1034)。由于正常组织和肿瘤组织在b7h3表达水平上的显著差异,可以通过b7h3抗体的adcc效应或毒素偶联来有效地杀死肿瘤细胞,而对正常组织不导致太大的影响(koenig s.,et al.,(2014),medicographia 36:285)。总之,根据当
前的研究信息,b7h3是一个比较有希望的癌症治疗靶点。
技术实现要素:
7.本发明通过杂交瘤技术获得了一组抗b7h3单克隆抗体,这些抗体具较高的b7h3亲和力、显著的细胞内吞作用(可通过与毒素偶联选择性地毒杀或抑制肿瘤)、显著的抗体依赖性细胞毒性(adcc)、或能有效地阻断b7h3引起的免疫抑制作用;本发明成功地对其中6个候选抗体进行了人源化改造。所述抗体在制备应用于抑制癌细胞及调节b7h3的作用、水平以及增强机体免疫力的相关药物,尤其是治疗癌症相关药物方面具有广阔的应用前景。
8.本发明提供的鼠源或人源化b7h3抗体或功能性片段,其包括了重链序列和轻链序列。抗人b7h3鼠源抗体,重链可变区、轻链可变区的氨基酸序列信息如下:7f5重链可变区、轻链可变区的氨基酸序列分别为seqidno:1、2;9c8重链可变区、轻链可变区的氨基酸序列分别为seqidno:3、4;5b6重链可变区、轻链可变区的氨基酸序列分别为seqidno:5、6;7c9重链可变区、轻链可变区的氨基酸序列分别为seqidno:7、8;2a9重链可变区、轻链可变区的氨基酸序列分别为seqidno:9、10;4f11重链可变区、轻链可变区的氨基酸序列分别为seqidno:11、12;15a2重链可变区、轻链可变区的氨基酸序列分别为seqidno:13、14;7b7重链可变区、轻链可变区的氨基酸序列分别为seqidno:15、16;7e6重链可变区、轻链可变区的氨基酸序列分别为seqidno:17、18;2e10重链可变区、轻链可变区的氨基酸序列分别为seqidno:19、20;2f12重链可变区、轻链可变区的氨基酸序列分别为seqidno:21、22;2f7重链可变区、轻链可变区的氨基酸序列分别为seqidno:23、24;13a2重链可变区、轻链可变区的氨基酸序列分别为seqidno:25、26;14b3重链可变区、轻链可变区的氨基酸序列分别为seqidno:27、28。
9.以上抗体重链、轻链的cdr1、cdr2、cdr3氨基酸序列信息如下:7f5重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:29、30、31,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:32、33、34;9c8重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:35、36、37,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:38、39、40;5b6重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:41、42、43,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:44、45、46;7c9重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:47、48、49,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:50、51、52;2a9重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:53、54、55,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:56、57、58;4f11重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:59、60、61,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:62、63、64;15a2重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:65、66、67,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:68、69、70;7b7重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:71、72、73,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:74、75、76;7e6重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:77、78、79,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:80、81、82;2e10重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:83、84、85,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:86、87、88;2f12重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:89、90、91,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:92、93、94;2f7重链cdr1、cdr2、cdr3氨基酸序列分别为seqidno:95、96、97,其轻链
cdr1、cdr2、cdr3氨基酸序列分别为seq idno:98、99、100;13a2重链cdr1、cdr2、cdr3氨基酸序列分别为seq id no:101、 102、103,其轻链cdr1、cdr2、cdr3氨基酸序列分别为seq id no:104、105、106。 14b3重链cdr1、cdr2、cdr3氨基酸序列分别为seq id no:107、108、109,其轻链 cdr1、cdr2、cdr3氨基酸序列分别为seq id no:110、111、112。
10.进一步,抗人b7h3抗体或片段经过改造后,成为人源化抗体。
11.抗人b7h3人源化抗体2a9重链可变区氨基酸序列为seq id no:113,其轻链可变区氨基酸序列为seq id no:114;人源化抗体4f11重链可变区氨基酸序列为seq id no: 115,其轻链可变区氨基酸序列为seq id no:116;人源化抗体9c8重链可变区氨基酸序列为seq id no:117,其轻链可变区氨基酸序列为seq id no:118;人源化抗体15a2重链可变区氨基酸序列为seq id no:119,其轻链可变区氨基酸序列为seq id no:120;人源化抗体5b6重链可变区氨基酸序列为seq id no:121,其轻链可变区氨基酸序列为 seq id no:122;人源化抗体7b7重链可变区氨基酸序列为seq id no:123,其轻链可变区氨基酸序列为seq id no:124。
12.表达载体,包含上述抗体的核酸分子。
13.药物组合物,其包含上述的抗体或其功能片段,以及药用载体。
14.上述的抗体或其功能性片段,核酸分子、表达载体、宿主细胞、药物组合物在制备 b7h3免疫功能药物的用途。
15.本专利采用哺乳动物细胞表达系统制备重组的b7h3受体蛋白作为抗原,免疫小鼠后,将小鼠脾脏细胞与骨髓瘤细胞融合获得杂交瘤细胞。通过对大量杂交瘤细胞进行多次克隆及筛选,得到多个单克隆杂交瘤细胞株。这些杂交瘤细胞株可以分泌产生与b7h3受体特异性结合的单克隆抗体(图1、图2),这些单克隆抗体中,部分具有明显的被细胞内吞的作用(图3、图4)、部分具有促进人免疫细胞分泌细胞因子如ifn
‑
γ的作用(图5)、以及部分具有明显的介导adcc的作用(图6、图7、图8),其中部分单克隆抗体可被用于将来的adc,adcc以及b7h3拮抗剂的开发。进一步地,通过rt
‑
pcr(reverse transcription
‑ꢀ
polymerase chain reaction,逆转录聚合酶链式反应)克隆编码抗体轻链和重链可变区的基因,采用互补决定簇嫁接方法(complementarity
‑
determining regions graft,cdr
‑
graft)构建人源化抗体。体外功能实验表明,人源化b7h3抗体能特异性地结合b7h3受体蛋白(图9、图 10),同时保持了明显的被细胞内吞的作用(图11)及偶联mmae毒素后,对calu
‑
6肿瘤细胞有着明显的毒杀作用(图12)、介导adcc的作用(图13)、介导nk92mi
‑
hcd16细胞对jurkat
‑
b7h3和pc9细胞的adcc毒杀作用(图14)、以及促进人免疫细胞产生和分泌细胞因子如ifn
‑
γ的作用(图15)。本专利中采用的阳性对照抗体来自专利us8802091和 wo2017180813al。
附图说明
16.图1(a
‑
b):用elisa方法测定b7h3杂交瘤单克隆抗体与b7h3
‑
hfc蛋白的结合特性
17.图2(a
‑
b):用facs方法测定b7h3杂交瘤单克隆抗体与pc9细胞的结合特性
18.图3(a
‑
b):用facs测定b7h3杂交瘤抗体被jurkat
‑
b7h3细胞内吞的活性
19.图4(a
‑
b):用facs测定b7h3杂交瘤抗体被pc9细胞内吞的活性
20.图5(a
‑
b):b7h3杂交瘤抗体对人的pbmc细胞ifn
‑
γ分泌的促进作用
21.图6(a
‑
b):用报告基因方法测试b7h3杂交瘤或嵌合抗体所介导的对jurkat
‑
b7h3
细胞的 adcc作用
22.图7(a
‑
b):用报告基因方法测试一些b7h3嵌合抗体所介导的对pc9肿瘤细胞的adcc 作用
23.图8:用报告基因方法测试一些b7h3嵌合抗体所介导的对dld1肿瘤细胞的adcc作用
24.图9:用elisa方法测定人源化b7h3单克隆抗体与b7h3
‑
his蛋白的结合特性
25.图10:用facs方法测定人源化b7h3单克隆抗体与pc9细胞中b7h3蛋白的结合特性
26.图11:用facs测定人源化b7h3单克隆抗体被pc9细胞内吞的作用
27.图12:人源化b7h3抗体adc对calu
‑
6肿瘤细胞的毒杀作用
28.图13(a
‑
b):用报告基因方法测试人源化b7h3抗体对jurkat
‑
b7h3和pc9细胞的adcc 作用
29.图14(a
‑
b):人源化b7h3抗体介导nk92mi
‑
hcd16细胞对jurkat
‑
b7h3和pc9细胞的 adcc毒杀作用
30.图15:人源化b7h3单克隆抗体对人的pbmc细胞ifn
‑
γ产生和分泌的促进作用。
具体实施方式
31.实施例1
32.产生抗b7h3抗体的小鼠免疫和细胞融合
33.用人源b7h3(ncbi reference sequence:nm_001024736.2)胞外区与mfc融合蛋白 (b7h3
‑
ecd
‑
mfc)作为抗原,与等体积完全弗氏佐剂(sigma,cat.no.:f5581)充分乳化后,经皮下免疫6
‑
8周龄balb/c小鼠(购自昭衍(苏州)新药研究中心有限公司),抗原免疫量为50μg/只。随后每隔2周,用相同剂量的抗原与不完全弗氏佐剂(sigma,cat.no.: f5506)充分乳化后,经皮下免疫小鼠三次。三次免疫后测定小鼠血清效价,融合前3天通过腹腔注射进行一次加强免疫。以peg hybri
‑
max(sigma,cat.no.:7181)作为融合剂,将小鼠脾脏细胞与sp2/0细胞按照4:1比例混合,将融合的细胞加入到96孔板中(1x105个/孔),每孔含有0.1ml 1x hat(invitrogen,cat.no.:21060
‑
017)培养基。在第3天加入0.1mlht(invitrogen,cat.no.:11067
‑
030)培养基,在第7天吸掉96孔板中的培养基,补加0.2 ml新鲜ht培养基。第9天收取上清液进行各种筛选和测试。
34.实施例2
35.杂交瘤抗体的抗原结合力以及亚克隆
36.杂交瘤抗体的结合力测试包括elisa和facs,以及采用有限稀释法的亚克隆:
37.1)用elisa筛选b7h3结合阳性克隆:用50μl b7h3
‑
hfc(终浓度:2μg/ml)包被 96孔elisa板(corning,cat.no.:9018),室温过夜;用洗涤缓冲液(pbs+0.05%tween20) 洗涤3次后,加入封闭缓冲液(pbs+2%bsa(sigma,cat.no.:v90093))室温孵育1小时,用洗涤缓冲液洗涤elisa板3次;加入杂交瘤上清液,室温孵育1小时,洗涤3次;每孔加入100μl的10000倍稀释的hrp偶联羊抗鼠igg二抗(thermo,cat.no.:31432),室温避光孵育1小时,洗涤3次;每孔加入100μl tmb(北京百奥赛博,cat.no.:es
‑
002),室温孵育显色2分钟,加入100μl/孔的终止液(2n h2so4)终止显色反应,用酶标仪(tecanspark)读取各孔od450数值。
38.2)用facs筛选b7h3结合阳性克隆:取50μl在上述检测中呈阳性的杂交瘤上清或纯
化的杂交瘤抗体与50μl 293t
‑
b7h3细胞混合(2x105个/孔),加至96孔u底细胞板中4℃孵育1小时,用facs缓冲液(pbs+3%fcs)洗涤并离心两次,加入400倍稀释的pe标记的羊抗鼠二抗(biolegend,cat.no.:405307),4℃避光孵育30分钟,用facs缓冲液洗涤并离心两次,用bd accuri c6流式细胞仪检测细胞在pe通道信号值。
39.3)亚克隆:采用有限稀释法进行亚克隆
‑
对在上述elisa和facs检测中呈阳性的多克隆杂交瘤细胞进行亚克隆,随后再重复采用elisa和facs方法进行检测筛选,获得阳性杂交瘤单克隆。
40.抗体纯化、浓度测定及内毒素测定:
41.将阳性单克隆杂交瘤细胞置于50ml无血清培养基中(invitrogen,cat.no.:12045
‑
076) 培养8
‑
9天后,离心收取上清液。用protein a亲和层析纯化单克隆抗体,纯化后的抗体样品经超滤离心管(millipore,cat.no.:acs500024)换液浓缩后,用bca方法测定蛋白浓度,用鲎试剂(厦门鲎试剂生物科技股份有限公司)检测纯化的抗体样品的内毒素含量。
42.用elisa和facs检测纯化的抗体样品与b7h3的结合能力,结果如图1(a
‑
b)、图2(a
‑ꢀ
b)和表1。所选杂交瘤抗体都具较高的亲和力。
43.表1,b7h3杂交瘤单克隆抗体与b7h3抗原结合力ec50值
[0044][0045]
实施例3
[0046]
杂交瘤单克隆抗体的细胞内吞作用和b7h3阻断作用的测试
[0047]
对抗体的细胞内吞噬作用分析:在96孔板中,每孔加入50μl 2
×
106/ml pc9细胞,以及50μl标记的b7h3抗体,将96孔细胞培养板置于4℃孵育1小时,用facs buffer洗涤 2次后加入ph依赖性荧光染料cypher5e(ge,cat.no.:pa15401)标记的goat anti
‑
mfc (jackson,cat.no.:115
‑
005
‑
071)二抗,4℃孵育0.5小时,洗涤2次后加入培养基 (1640+10%fbs),放置培养箱中孵育3小时,离心去培养基,于ph9.0的pbs中重悬,用bd c6流式细胞仪检测pc9细胞的cypher5e信号,计算b7h3抗体进入细胞内部的效率。如图3(a
‑
d)和图4(a
‑
d)所示,jurkat
‑
b7h3和pc9细胞对b7h3抗体,包括15a2,5b6,7c9, 2f7,7b7,2e10,4f11等,具有显著的内吞活性。
[0048]
测试b7h3杂交瘤抗体对人pbmc细胞分泌细胞因子的影响:在96孔(corning,cat.no.: 3799)加入用完全培养基(rpmi1640+10%fcs)重悬的pbmc细胞(tpcs,cat.no.: pb025c),再加入40ng/ml okt3(ebioscience,cat.no.:16
‑
0037
‑
85),置37℃孵育72小时。活化的pbmc细胞计数后,用完全培养基重悬(2.5
×
105细胞/ml)。在96孔板中,每孔加入 100μl pbmc细胞,50μl不同浓度的b7h3抗体(起始浓度为20μg/ml,10倍系列稀释),将96孔细胞培养板(corning,cat.no.:3599)置于37℃,5%co2培养箱中孵育48小时,收集上清液。用ifn
‑
γelisa试剂盒(r&d systems,cat.no.:dy285)检测细胞因子的浓度。如图 5所示,b7h3抗体包括5b6,2f12,2f7,15a2,13a2,14b3,4f11,2a9等能够显著促进 pbmc细胞分泌
ifn
‑
γ。
[0049]
实施例4
[0050]
b7h3抗体可变区基因克隆和测序
[0051]
用trizon(cwbiotech,cat.no.:cw0580)裂解b7h3单克隆杂交瘤细胞株,提取杂交瘤细胞总rna。用hifi script cdna合成试剂盒(cwbiotech,cat.no.:cw2569)将杂交瘤细胞的rna反转录为cdna。以cdna为模板,用简并引物通过pcr方法(kettleborough et al., (1993),eur j immunology 23:206
‑
211;strebe,et al.,(2010),antibody engineering 1:3
‑
14)扩增抗体的重链和轻链的可变区基因。将pcr扩增产物连接到t/a载体后,转化dh5a感受态细胞,涂板并置37℃过夜培养。从培养板上挑取单克隆,扩大培养后抽提质粒,测定抗体的基因序列。根据抗体的基因序列,分析其互补决定簇(cdr)和骨架区。b7h3抗体序列编号如表2所示,具体序列信息见序列表。
[0052]
表2.b7h3抗体序列编号说明
[0053]
[0054]
[0055]
[0056][0057]
实施例5
[0058]
杂交瘤单克隆抗体的adcc活性测试
[0059]
用报告基因(reporter)法测试抗体的adcc活性:以293t-b7h3、pc9或dld1肿瘤细胞为靶向细胞,jurkat
‑
mcd16.2
‑
nf
‑
kb或jurkat
‑
hcd16
‑
nf
‑
kb细胞为效应细胞,利用 mcd16.2或hcd16介导的转录因子nf
‑
kb的活化信号筛选adcc阳性克隆。取50μl在上述elisa和facs检测中呈阳性的杂交瘤抗体与25μl的293t
‑
b7h3、pc9或dld1肿瘤细胞(7.5x104个/
孔)混合,加至96孔板,再加入25μl jurkat
‑
mcd6.2
‑
nf
‑
kb细胞 (2.5x104个/孔),混匀,置于37℃培养箱中孵育4小时。每孔加入25μl预热的bright
‑ꢀ
glo溶液(promega,cat.no.:e2620),室温避光静置3分钟,用酶标仪(tecan spark)测定各样品的冷光(luminescence)信号值。
[0060]
由于杂交瘤抗体中,小鼠的igg1抗体没有adcc活性。经基因测序后,我们确认了14 个杂交瘤抗体中,9个是以小鼠igg1为基础的,5个是以igg2b或igg2a为基础的(见表 3)。图6a显示了杂交瘤抗体基于jurkat
‑
b7h3为靶向细胞的adcc报告基因测试结果。跟预期的一样,5个以igg2b或igg2a为基础的杂交瘤抗体具adcc活性,而9个以小鼠igg1 为基础的,没有活性。因此对于9个以小鼠igg1为基础的杂交瘤瘤抗体,我们作fc的替换,把这些抗体变成了以人源igg1为基础的嵌合抗体,如图6b所示。图7和图8反映的是基于pc9或dld1肿瘤细胞为靶向细胞的adcc报告基因测试结果,抗体的形式是以人源 igg1为基础的嵌合抗体。综合adcc报告基因的测试结果,adcc活性相对较强的包括9c8, 5b6,7c9,f5,7e6,2a9,4f11等。
[0061]
表3,b7h3杂交瘤抗体的fc类型
[0062]
abs2a913a24f119c85b67e67f52f77c92e107b714b32f1215a2fcigg2bigg2bigg1igg1igg1igg1igg1igg2bigg1igg2aigg2bigg1igg1igg1
[0063]
实施例6
[0064]
人源化b7h3抗体2a9、4f11、9c8、15a2、5b6和7b7
[0065]
综合杂交瘤抗体的各方面特性,我们挑选了6个b7h3抗体,包括2a9、4f11、9c8、 15a2、5b6和7b7,进行了人源化改造。
[0066]
采用互补决定簇嫁接法进行b7h3抗体的人源化改造。首先,在imgt数据库中分别搜寻与鼠源2a9、4f11、9c8、15a2、5b6和7b7抗体的轻、重链可变区序列同源性最高的人胚系抗体(germline antibody)序列。2a9抗体轻链可变区人源化选取的胚系为igkv3
‑
11*01,重链可变区人源化选取ighv1
‑
2*02。4f11抗体轻链可变区人源化选取的胚系为igkv3
‑
11*01,重链可变区人源化选取ighv1
‑
2*02。9c8 抗体轻链可变区人源化选取的胚系为igkv3
‑
11*01,重链可变区人源化选取 ighv1
‑
46*01。15a2抗体轻链可变区人源化选取的胚系为igkv3
‑
11*01,重链可变区人源化选取ighv1
‑
8*01。5b6抗体轻链可变区人源化选取的胚系为igkv1
‑ꢀ
39*01,重链可变区人源化选取ighv1
‑
69*02。7b7抗体轻链可变区人源化选取的胚系为igkv1
‑
39*01,重链可变区人源化选取ighv3
‑
21*01。保留鼠源抗体的cdr 区,将鼠源抗体的框架区(framework)序列用人胚系抗体的框架区序列置换。建立鼠源抗体的结构模型,逐个对比人源抗体与相应鼠源抗体框架区中每个位点的氨基酸,如果框架区的某个位点采用人的氨基酸序列没有导致cdr区域空间结构的破坏或改变,则该位点使用人的氨基酸序列,否则在该位点使用对应的鼠源序列(即回复突变为鼠源序列)。
[0067]
根据结构模拟,将2a9抗体人源化重链的第24位ala回复突变为thr,第48 位met回复突变为ile,第67位val回复突变为ala,第69位met回复突变为 leu,第71位arg回复突变为val,第73位thr回复突变为lys。将2a9抗体人源化轻链的第46位leu回复突变为arg,第47位leu回复突变为trp,第48位ile回复突变为val,第71位phe回复突变为tyr。将4f11抗体人源化重链的第24位ala 回复突变为thr,第48位met回复突变为ile,第67位val回复突变为ala,第69 位met回复突变为leu,第71位arg回复突变为val,第73位thr回复突变为 lys。将
4f11抗体人源化轻链的第46位leu回复突变为arg,第47位leu回复突变为trp,第48位ile回复突变为val,第71位phe回复突变为tyr。将9c8抗体人源化重链的第48位met回复突变为ile,第67位val回复突变为ala,第69位met 回复突变为leu,第71位arg回复突变为val,第73位thr回复突变为lys。将 9c8抗体人源化轻链的第2位ile回复突变为thr,第46位leu回复突变为arg,第 47位leu回复突变为trp,第71位phe回复突变为tyr。将15a2抗体人源化重链的第48位met回复突变为ile,第67位val回复突变为ala,第69位met回复突变为leu,第71位arg回复突变为ala,第73位thr回复突变为lys。将15a2抗体人源化轻链的第36位tyr回复突变为phe,第47位leu回复突变为trp,第49位 tyr回复突变为his,第58位ile回复突变为phe,第71位phe回复突变为tyr。将 5b6抗体人源化重链的第27位gly回复突变为tyr,第30位ser回复突变为ile,第 48位met回复突变为ile,第67位val回复突变为ala,第69位ile回复突变为 leu。将5b6抗体人源化轻链的第46位leu回复突变为arg,第47位leu回复突变为pro,第66位gly回复突变为ala,第71位phe回复突变为tyr,第98位phe回复突变为ile。将7b7抗体人源化重链的第49位ser回复突变为ala。将7b7抗体人源化轻链的第2位ile回复突变为ser,第48位ile回复突变为val。
[0068]
人源化抗体重链和轻链的可变区氨基酸序列号分别为seq id no:113和seqid no:114。4f11人源化抗体重链和轻链的可变区氨基酸序列号分别为seq id no: 115和seq id no:116。9c8人源化抗体重链和轻链的可变区氨基酸序列号分别为 seq id no:117和seq id no:118。15a2人源化抗体重链和轻链的可变区氨基酸序列号分别为seq id no:119和seq id no:120。5b6人源化抗体重链和轻链的可变区氨基酸序列号分别为seq id no:121和seq id no:122。7b7人源化抗体重链和轻链的可变区氨基酸序列号分别为seq id no:123和seq id no:124。将以上人源化抗体构建为igg1亚型。人源化抗体的重链和轻链的可变区氨基酸序列信息见表4。
[0069]
合成编码2a9、4f11、9c8、15a2、5b6和7b7人源化抗体轻链和重链的核酸序列,并插入到表达载体pcdna3.1。用0.1mg抗体轻链和0.1mg抗体重链表达质粒共转染200ml 的293细胞(细胞密度为1
×
106/ml),在37℃摇瓶振摇培养6天,离心收集上清液,用 protein a纯化人源化抗体,纯化后的人源化抗体进行活性检测。
[0070]
表4.b7h3人源化抗体的序列表。
[0071][0072]
实施例7
[0073]
人源化b7h3抗体与b7h3的结合作用
[0074]
用elisa、facs检测人源化b7h3抗体样品与b7h3蛋白的结合活性,具体方法参考实施例2。人源化抗体hu2a9、hu4f11、hu5b6、hu9c8、hu15a2和hu7b7的测定结果见表 5
‑
6和图9
‑
10。总体上,本发明的抗体经人源化后,保持了人源化前的高抗原亲和力的作用,尤其是hu15a2、hu7b7和hu5b6。
[0075]
表5.用elsia检测人源化b7h3抗体与b7h3
‑
mfc蛋白的结合作用
[0076][0077]
表6.用facs测定人源化b7h3抗体和细胞表面b7h3的结合作用
[0078][0079]
实施例8
[0080]
人源化b7h3抗体细胞内吞作用的检测
[0081]
具体测定方法参考实施例3。结果如表7和图11所示;pc9细胞对人源化b7h3抗体具有显著的内吞作用。与阳性对照(macrogenics的mga018和mskcc的8h9)比较,本发明抗体的细胞内吞作用比8h9强不少;另外,hu15a2和hu7b7也胜过mgc018。
[0082]
表7.pc9肿瘤细胞对人源化b7h3抗体的内吞作用
[0083][0084]
实施例9
[0085]
与毒素偶联的人源化b7h3抗体(b7h3
‑
adc)对calu
‑
6肿瘤细胞的毒杀作用
[0086]
通过化学偶联方法,将vc
‑
mmae偶联到b7h3抗体上。按3:1摩尔比用tcep还原b7h3 抗体,在37℃孵育45分钟,通过超滤去除tcep。用0.5ml pbs重悬抗体,加入b7h3抗体 6倍摩尔数的vc
‑
mmae,于4℃孵育120分钟,用超滤去除未偶联的vc
‑
mmae,检测抗体浓度备用。将100μl pc9(每孔5000细胞)和100μl梯度稀释的b7h3
‑
adc加入到96孔板。在37℃孵育5天后,每孔加入xμl celltiter
‑
glo,检测荧光强度,计算抗体抑制肿瘤细胞增殖的能力。结果如图12和表8所示,人源化b7h3抗体偶联mmae毒素后对calu
‑
6肿瘤细胞具有显著的毒杀作用;比macrogenics的阳性对照(mgc018
‑
mmae)具更强的活性。
[0087]
表8.人源化b7h3抗体与mmae的偶联物对calu
‑
6肿瘤细胞的毒杀作用
[0088][0089]
实施例10
[0090]
人源化b7h3抗体adcc作用的检测
[0091]
具体测定方法参考实施例5。结果如表9和图13,展示了人源化b7h3抗体具显著的 adcc活性。mga271(mg
‑
ab)是macrogenics的抗体;与之相比较,人源化9c8、5b6、 2a9和4f11抗体具更小的ec50值相,反映了更好的adcc活性。
[0092]
表9.人源化b7h3抗体的adcc作用
[0093][0094]
实施例11
[0095]
人源化b7h3抗体介导nk92mi
‑
hcd16细胞对jurkat
‑
b7h3和pc9细胞的adcc毒杀作用
[0096]
采用nk92mi
‑
hcd16模拟nk细胞来评估b7h3抗体的杀伤肿瘤活性。将25μl用cfse 染色的pc9(每孔0.5x105细胞)和25μl nk92mi
‑
hcd16(每孔1x105细胞)混合,加入50 μl梯度稀释的b7h3抗体。在37℃下孵育4小时后,每孔加入5μl 7
‑
aad,室温孵育10 分钟,将96孔板离心去上清液重悬后,使用流式细胞仪检测cfse和7
‑
aad双阳性的细胞在cfse阳性细胞的比例,得出b7h3抗体介导的nk细胞杀伤活性。结果如图14所示,人源化 b7h3抗体9c8、5b6、2a9和4f11可明显引起nk92mi
‑
hcd16细胞对pc9肿瘤细胞的adcc 杀伤作用。
[0097]
实施例12
[0098]
人源化b7h3抗体对人pbmc细胞分泌细胞因子的促进作用
[0099]
具体测定方法参考实施例3。结果如图15所示,人源化b7h3抗体5b6、2a9和4f11 有显著促进人pbmc细胞分泌ifn
‑
γ的作用。
[0100]
以上仅为本发明的较佳实施例而已,不能以此限定本发明的保护范围,即大凡依本发明权利要求书及发明内容所做的简单的等效变化与修改,皆仍属于本发明专利申请的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1