热稳定性提高的双果糖酐水解酶突变体E160F
热稳定性提高的双果糖酐水解酶突变体e160f
技术领域
1.本发明涉及热稳定性提高的双果糖酐水解酶突变体e160f,属于酶工程技术领域。
背景技术:
2.菊糖是一种膳食纤维,广泛用于食品领域。菊糖是一种由果糖基通过β-(2,1)键聚合而成的果聚糖,其末端含有一分子葡萄糖。菊糖可以被多种酶代谢掉,包括被菊粉酶降解为膳食纤维低聚果糖,广泛用于酸奶、饮料等制品中。或者可以被菊糖果糖转移酶(inulinfructotransferase,iftase)转化成为益生元双果糖酐(difructose anhydride iii,dfa-iii)。这种代谢途径是一种新型途径,所生成的双果糖酐是一种新型功能性甜味剂,具有益生元功效,已经在日本应用于食品体系中了。双果糖酐不被人体消化吸收,但是在大肠中被肠道微生物中的双果糖酐水解酶水解为菊二糖,菊二糖进一步被微生物果糖苷酶水解成为果糖而被人体吸收利用。
3.目前对于菊糖代谢中菊粉酶途径已经研究的非常清晰,但是对于双果糖酐水解酶合成菊二糖的研究较少。菊二糖可能是一种新型的功能甜味剂,但是其生理功能未有相关报道,甚至理化性质研究也鲜有报道。菊糖代谢途径中其它糖都具有重要生理功能,因此,菊二糖值得探索研究。然而市场上并无菊二糖纯样品可售,目前也极少有关菊二糖合成报道。菊二糖的有限生产可能限制了其性质研究,这主要是由于双果糖酐水解酶活力较低以及热稳定性较差(55℃条件下半衰期为2小时)的原因。因此,本发明的目的是探索解决其限制稳定性差这一限制因素,通过酶工程手段筛选获得热稳定性提高的双果糖酐水解酶。
技术实现要素:
4.为了解决上述问题,本发明通过对微生物arthrobacter chlorophenolicus a6来源的双果糖酐水解酶进行分子改造,获得酶活与野生型酶无明显变化且热稳定性显著提升的双果糖酐水解酶突变体。
5.本发明提供了一种双果糖酐水解酶突变体e160f,所述突变体e160f的氨基酸序列如seq id no.3所示。
6.本发明提供了编码上述双果糖酐水解酶突变体e160f的基因。
7.在本发明的一种实施方式中,所述基因的核苷酸序列如seq id no.4所示。
8.本发明还提供了携带上述基因的重组载体。
9.在本发明的一种实施方式中,所述重组载体以pet系列为表达载体。
10.在本发明的一种实施方式中,所述重组载体以pet-22a(+)为表达载体。
11.本发明提供了携带上述基因,或上述重组载体的微生物细胞。
12.在本发明的一种实施方式中,所述微生物细胞以细菌或真菌为表达宿主。
13.在本发明的一种实施方式中,所述微生物细胞以大肠杆菌为表达宿主。
14.本发明还提供了一种基因工程菌,所述基因工程菌以大肠杆菌为表达宿主,以pet-22b(+)为载体,表达上述双果糖酐水解酶突变体e160f。
15.在本发明的一种实施方式中,所述大肠杆菌包括e.coli bl21(de3)。
16.本发明还提供了一种提高双果糖酐水解酶热稳定性的方法,所述方法为在氨基酸序列如seq id no.1所示的双果糖酐水解酶的第160位谷氨酸突变成苯丙氨酸。
17.本发明还提供了上述突变体,或上述基因,或上述重组载体,或上述微生物细胞,或上述基因工程菌或上述方法在食品领域中的应用。
18.本发明还提供了上述突变体,或上述基因,或上述重组载体,或上述微生物细胞,或上述基因工程菌或上述方法在制备含有菊二糖的产品中的应用。
19.有益效果:
20.本发明微生物arthrobacter chlorophenolicus a6来源的双果糖酐水解酶进行分子改造,对氨基酸序列如seq id no.1所示的双果糖酐水解酶的160位谷氨酸进行定点突变,获得了热稳定性提高的突变酶e160f。野生酶在55℃条件下半衰期为2小时,65℃下半衰期为30分钟,而本发明提供的突变体e160f在55℃条件酶活基本不受影响,65℃条件下半衰期为8小时。这有利于在酶最适温度55℃下固定化长时间催化生产,解决了双果糖酐水解酶热稳定性差的问题,为拓宽双果糖酐水解酶工业应用奠定了基础。
附图说明
21.图1:双果糖酐水解酶及突变体e160f纯化后的sds-page;
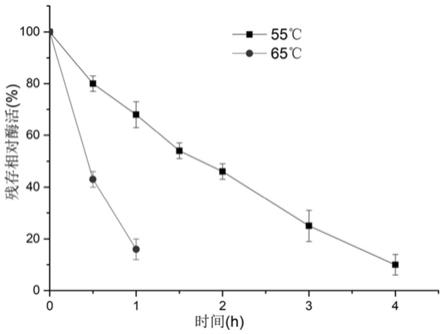
22.图2:双果糖酐水解酶温度稳定性;
23.图3:双果糖酐水解酶的突变体e160f温度稳定性。
具体实施方式
24.以下结合说明书附图和具体实施例来进一步说明本发明,但实施例并不对本发明做任何形式的限定。除非特别说明,本发明采用的试剂、方法和设备为本技术领域常规试剂、方法和设备。
25.除非特别说明,以下实施例所用试剂和材料均为市售商品或者可以通过已知方法制备。
26.下述实施例中涉及的培养基如下:
27.培养基均使用ddh2o配制,配制完成后121℃灭菌15~20min。
28.lb液体培养基:酵母粉5.0g/l、胰蛋白胨10.0g/l、nacl 10.0g/l。
29.lb固体培养基:酵母粉5.0g/l、胰蛋白胨10.0g/l、nacl 10.0g/l、琼脂粉15g/l。
30.实施例1:野生型双果糖酐水解酶的制备和酶活测定
31.微生物arthrobacter chlorophenolicus a6菌种在genbank中基因组编号是cp001341,包含双果糖酐水解酶基因(编号achl_2895),基因全长为1338个核苷酸(seq id no.2)。该基因编码的蛋白质酶编号是acl40859.1,共445个氨基酸(seq id no.1)。
32.1)重组菌株的构建:
33.将编码双果糖酐水解酶的基因在大肠杆菌工程菌中进行分子克隆表达,将核苷酸序列如seq id no.2双果糖酐水解酶基因构建至质粒载体pet-22b(+)的xho i和nde i酶切位点之间,构建得到重组质粒,并命名为pet-22b(+)-acdfa-iiiase。
34.将质粒pet-22b(+)-acdfa-iiiase转化入e.coli bl21(de3)感受态细胞中,在含
有100μg ml-1
氨苄青霉素的lb固体培养基37℃培养过夜后,挑阳性单克隆转化子于含有50μg ml-1
的氨苄青霉素的lb液体培养基中,37℃、200rpm培养至菌体吸光度值od为0.6~0.8,添加终浓度为0.5-1mm异丙基硫代半乳糖苷(iptg)进行诱导表达6小时获得发酵液。
35.2)蛋白纯化:
36.发酵液经过离心20min(4℃,l0000 rpm)后取菌体沉淀部分。加入20ml缓冲液(50mm pbs,200mm nacl,ph 6.5)重悬菌体并使用超声波细胞破碎仪进行细胞破碎处理,超声条件为:超声ls,停止2s,共计15min。破碎液进行低温高速离心30min(4℃,10000rpm)得粗酶液。用0.45μm微孔滤膜过滤备用。
37.使用镍离子亲和层析纯化和透析后得到双果糖酐水解酶acdfa-iiiase纯酶液,见图1,sds-page显示acdfa-iiiase达到电泳纯级别。
38.3)acdfa-iiiase的的酶活测定
39.将acdfa-iiiase与双果糖酐反应,在1ml酶催化反应体系中包括10g l-1
双果糖酐,50mm ph 6.5磷酸盐缓冲液和100noml l-1
acdfa-iiiase纯酶液。在55℃条件下反应10min后沸水浴10min以终止反应。1u酶活定义为在ph 6.5,55℃条件下反应,每分钟生成1μmol菊二糖所需要的酶量。
40.反应液使用0.22μm滤膜过滤后的溶液上样至高效液相中,配备糖柱sugar-pak i(4.6mm
×
250mm,waters,ma,usa)和示差折光显示器检测菊二糖产量,双果糖酐水解酶acdfa-iiiase酶活为101.25u mg-1
。
41.实施例2:双果糖酐水解酶突变体e160f的制备及表达纯化
42.(1)突变体e160f的制备
43.核苷酸序列如seq id no.2双果糖酐水解酶基因为模板进行引物设计,以实施例1中构建的携带双果糖酐水解酶acdfa-iiiase基因的载体pet-22b(+)-acdfa-iiiase为模板进行定点突变构建突变质粒pet-22b(+)-e160f。
44.突变正向引物:5
’‑
gcggtcgccatccgttcaataccta-3’45.突变反向引物:5
’‑
attggcataggtattgaacggatggcga-3’46.pcr反应体系为:10
×
pcr buffer 5μl,dntp(2mmol/l)4μl,突变正向引物(10μm)1μl,突变反向引物(10μm)1μl,模板pet-22b(+)-acdfa-iiiase 1μl,taq plus dna polymerase(5u/μl)0.5μl,使用双蒸水将反应体系体积补足至50μl。
47.pcr扩增条件为:94℃预变性4min;94℃变性1min,56℃退火lmin,72℃延伸1min,共35个循环后72℃保温10min。
48.扩增产物利用琼脂糖电泳检测,并经测序为正确的e160f突变体后备用,将鉴定正确的突变质粒命名为pet-22b(+)-e160f。
49.(2)突变体e160f的表达纯化
50.将突变质粒pet-22b(+)-e160f转化至大肠杆菌bl21(de3)感受态细胞中涂布于含有100μg/ml氨苄青霉素的lb固体平板上,37℃培养过夜后,挑取阳性转化子于lb液体培养基中37℃、200rpm过夜培养后获得种子液;将种子液按照体积比1.5%接种量接入到500ml液体lb培养基进行扩大培养,于37℃条件下培养3~4h至od值为0.6~0.8后降温到28℃,加入终浓度为1mm iptg诱导8h,得到发酵液。
51.发酵液经过离心20min(4℃,l0000 rpm)后取菌体沉淀部分。加入20ml缓冲液
(50mm pbs,200mm nacl,ph 6.5)重悬菌体并使用超声波细胞破碎仪进行细胞破碎处理,超声条件为:超声ls,停止2s,共计15min。破碎液进行低温高速离心30min(4℃,10000rpm)得粗酶液。用0.45μm微孔滤膜过滤备用。
52.对e160f突变体粗酶液使用镍离子亲和层析柱在4℃环境下进行纯化,得到e160f突变体的纯酶液,sds-page显示纯酶液达到电泳纯,见图1。
53.(3)e160f突变体的酶活测定
54.将e160f突变体与双果糖酐反应,在1ml酶催化反应体系中包括10g l-1
双果糖酐,50mm ph 6.5磷酸盐缓冲液和100noml l-1
e160f突变体纯酶液。在55℃条件下反应10min后沸水浴10min以终止反应。1u酶活定义为在ph 6.5,55℃条件下反应,每分钟生成1μmol菊二糖所需要的酶量。
55.反应液使用0.22μm滤膜过滤后的溶液上样至高效液相中,配备糖柱sugar-pak i(4.6mm
×
250mm,waters,ma,usa)和示差折光显示器检测菊二糖产量,双果糖酐水解酶e160f突变体酶活为98.3u mg-1
,突变体酶活相较于原始酶活变化不大,即突变并不会显著影响酶活。
56.实施例3:酶热稳定性测定
57.(1)最适温度的检测:
58.在1ml的反应体系中,包括终浓度10g l-1
底物双果糖酐、终浓度50mm的ph 6.5磷酸盐缓冲液和100nmol l-1
纯酶液,分别在30℃、40℃、45℃、50℃、55℃、60℃、65℃、70℃、80℃条件下反应10min,沸水浴10min终止反应。18000
×
g 4℃离心20min后,反应液经过0.22μm滤膜过滤到液相小瓶中,高效液相配备糖柱sugar-pak i(4.6mm
×
250mm,waters,ma,usa)和示差折光显示器用于检测菊二糖。1u酶活定义为在ph 6.5,55℃条件下反应,每分钟生成1μmol菊二糖所需要的酶量。
59.结果显示,双果糖酐水解酶acdfa-iiiase和e160f突变体的最适催化温度均为55℃。
60.(2)热稳定性检测:在1ml的反应体系中,包括10g l-1
双果糖酐、50mm ph 6.5磷酸盐缓冲液和100nmol l-1
纯酶,分别在55℃、65℃条件下反应,期间取样进行残存酶活测定。
61.结果如果2和图3所示,在55℃条件下双果糖酐水解酶acdfa-iiiase酶活半衰期为2小时,65℃下半衰期为30分钟,这不利于工业化生产。而突变体e160f在55℃条件下酶活随着反应时间的延长并没有发生明显变化,65℃条件下半衰期达到了8小时,大幅度提高了热稳定性。这使得酶可应用到固定化生产中,即在最适温度55℃下长时间固定化生产菊二糖,解决了双果糖酐水解酶热稳定性差问题,为拓宽双果糖酐水解酶工业应用奠定了基础。
62.虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1