F-box蛋白编码基因OsFBX250在水稻选育中的应用
f-box蛋白编码基因osfbx250在水稻选育中的应用
技术领域
1.本发明涉及基因工程和植物遗传育种领域,具体涉及f-box蛋白编码基因osfbx250在水稻选育中的应用。
背景技术:
2.f-box蛋白在真核生物中广泛存在,其n端有一个高度保守的f-box基序,该基序能介导蛋白质与蛋白质之间的互作,在泛素-蛋白酶体途径中因能够特异识别相关底物蛋白,从而参与多种生命活动,如细胞周期调控、细胞信号转导、细胞凋亡、转录调控、免疫反应、雄性不育及植物叶片发育等。多数f-box蛋白的c端常含有相关功能域,根据其c端的不同,将f-box蛋白分为3类:富含亮氨酸重复序列(fbl)、富含wd-40重得序列(fbw)及含有一些未确定或无明显结构特征的序列(fbx)。
3.在拟南芥和水稻中分别有779和687个f-box蛋白,这些f-box蛋白在许多信号通路中发挥作用。在拟南芥中,f-box蛋白与ask蛋白结合,形成scf复合体特异识别底物蛋白,通过泛素蛋白酶体途径参与蛋白质的降解,从而发挥相应的生物学功能。ufo(unusual floral organs)是植物中鉴定出的第一个f-box基因,该基因突变后可导致拟南芥一系列花发育异常,且ufo和atcul1有直接的相互作用,scf-ufo复合体的形式参与了对植物花器官发育的多方面调控。sleepy1(sly1)是一个151个氨基酸的f-box蛋白质,在赤霉素(ga)信号途径中也起重要作用:在ga的存在下,sly1可识别rga和gai并介导这两个della蛋白质类泛素化降解。拟南芥f-box蛋白tir1作为生长素受体通过与生长素直接相互作用,介导一系列生长素反应。也有研究发现f-box蛋白参与aba、干旱和高盐等胁迫反应。
4.水稻中大部分的f-box蛋白属于fbx亚家族,在水稻生长发育过程中发挥着重要的作用。osfbk12通过与osk1互作参与26s蛋白酶体降解途径,进一步识别底物ossams1使其降解,从而改变乙烯水平调控叶片的衰老。osfbx352的表达水平受脱落酸(aba)的诱导,通过影响aba的合成与代谢维持体内的aba水平,从而调控水稻种子的萌发。apo1为一个编码429个氨基酸与拟南芥ufo同源的f-box蛋白,提高该基因的表达量导致花序分枝和小穗增加,从而导水稻籽粒数和产量的增加。ddf1(dwarf and deformed flower 1),一个f-box蛋白编码基因,对水稻的营养和花发育至关重要。osfbk12编码含有kelch重复基序的f-box蛋白,与sams1相互作用以调节水稻的叶片衰老、种子大小以及粒数。
5.f-box蛋白编码基因在水稻生长发育过程中具有重要的作用,但在水稻籽粒发育方面报道还非常少,而水稻osfbx250基因对水稻的生长发育的影响目前暂未有任何研究。本发明发现osfbx250基因对水稻籽粒的发育有极其重要的调控作用,可应用于植物籽粒性状的遗传改良。
技术实现要素:
6.本发明的目的在于提供f-box蛋白编码基因osfbx250在水稻选育中的应用。
7.本发明的目的通过下述技术方案实现:
8.本发明发现一个t-dna插入突变体材料lg7的粒长、粒宽、粒厚与千粒重和对照品种中花11(zh11)相比均显著增加。经鉴定t-dna的插入位点,成功克隆了一个新的调控粒型的基因osfbx250,且该基因在t-dna插入突变体的幼穗中表达水平显著高于对照zh11,因此推测osfbx250基因正向调控水稻的粒型。进一步通过遗传转化实验,发现该基因经crispr技术敲除后籽粒粒长变短、粒宽变窄及千粒重下降;而过表达osfbx250可使籽粒的粒长变长、粒宽变宽、粒厚变厚且粒重增加。这些结果表明osfbx250基因具有重要的育种应用潜力,同时可用于深入研究f-box蛋白调控水稻粒型的分子机制。
9.基于本发明发现的osfbx250基因的功能,其可用于水稻选育中。所述的水稻选育为使水稻粒型变大,包括提高水稻籽粒的粒长、粒宽、粒厚和粒重,通过使水稻粒型变大提高水稻产量。所述的使水稻粒型变大通过提高osfbx250基因的表达实现,具体可通过转基因或改变osfbx250基因的启动子序列提高osfbx250基因的表达。
10.所述的osfbx250基因编码的蛋白氨基酸序列如seq id no.1所示;所述的osfbx250基因的cdna序列优选如seq id no.2所示;所述的osfbx250基因的启动子序列如seq id no.3所示。
11.所述的osfbx250基因具有植物品种改良或制备转基因植物的应用。
12.本发明的优点和效果:
13.(1)本发明通过实验证明,将osfbx250基因转入野生型水稻品种zh11,可获得粒型变大的转基因水稻;在zh11中敲除该基因可以获得粒型减小的水稻材料。与对照zh11相比,osfbx250的过表达转基因水稻株系中,粒宽显著增加、粒长显著变长及粒重显著增加。因此,osfbx250基因与水稻粒型相关,为培育粒型改变的转基因植物奠定基础。
14.(2)osfbx250基因的成功克隆,进一步证实了f-box基因家族在水稻生长发育过程中的重要作用,对阐明f-box基因家族的生物学功能有重要的意义。
15.(3)尽管目前已克隆到了一些调控水稻粒型的基因,但f-box基因家族是如何通过关键基因影响植物生长发育的分子机制仍不清楚。而本发明克隆的osfbx250基因能够提高水稻籽粒大小,对进一步阐明植物籽粒发育分子机理并通过基因工程手段培育优质、高产的作物新品种具有重要的理论意义和现实意义。
附图说明
16.图1是对照zh11和lg7突变体的籽粒表型图。标尺为1cm。
17.图2是对照zh11和lg7突变体的籽粒的性状统计柱状图。数据采用graphpad软件进行显著性分析(t-test),a、b、c、d分别指粒长、粒宽、粒厚与千粒重统计图,其中“**”与“***”分别代表p《0.01与p《0.001。
18.图3是lg7突变体经三代测序后的序列比对图。a为测序产生的序列数据,b为序列数据经blast比对后的结果。其中蓝色标记序列为水稻日本睛参考序列,绿色标记序列为pcr1301载体序列。
19.图4是对lg7突变体中t-dna的插入位点的鉴定图。a为t-dna插入位点模式图;b与c为pcr对插入位点的鉴定图;d与e分别为rt-pcr与rt-qpcr鉴定osfbx250的表达水平差异图,“***”代表p《0.001。
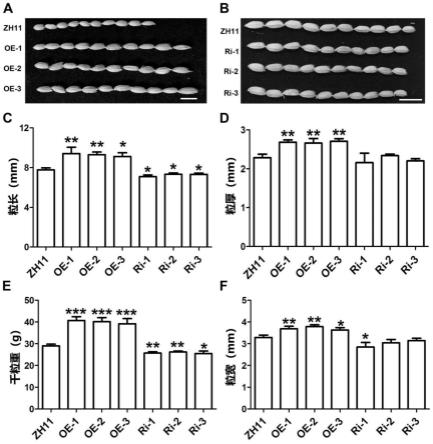
20.图5是对照zh11和osfbx250基因过表达、osfbx250基因干扰株系籽粒性状统计图。
a与b分别为osfbx250基因过表达与干扰株系的籽粒图;c、d、e、f分别为转基因株系籽粒的粒长、粒厚、千粒重与粒宽统计图。其中,oe-1、oe-2与oe-3指osfbx250基因过表达的三个株系,ri-1、ri-2与ri-3指osfbx250基因干扰的三个株系。“*”、“**”与“***”分别代表p《0.05、p《0.01与p《0.001。
21.图6是在zh11背景下采用crispr技术敲除osfbx250基因后表型图。a是osfbx250基因的敲除株系测序验证图;b是敲除株系的籽粒表型图;c、d、e、f与g分别是敲除株系籽粒的粒长、粒宽、粒厚、千粒重和结实率的统计图。c10、c12、c14与c15指osfbx250基因不同的敲除株系。“*”、“**”与“***”分别代表p《0.05、p《0.01与p《0.001。
具体实施方式
22.下面结合实施例对本发明做进一步详细的说明,但本发明的实施方式不限于此。若未特别指明,下述实施例所用的技术手段为本领域技术人员所熟知的常规手段;所用的实验方法均为常规方法,并可按照已描述的重组技术(参见分子克隆,实验室手册,第2版,冷泉港实验室出版社,冷泉港,纽约)完成;所用的材料、试剂等,均可从商业途径得到。
23.实施例1lg7基因的克隆
24.本发明鉴定到一个水稻籽粒增大的t-dna突变体lg7(图1),相对于对照zh11,lg7突变体籽粒的粒长、粒宽、粒厚及粒重均显著增加(图2)。通过提取lg7突变体的基因组dna,并于北京百迈客生物科技有限公司进行重测序。利用t-dna部分序列和重测序产生的dna序列进行序列比对,分析该突变体中t-dna的插入位点,初步确定t-dna插入到水稻基因组的第7号染色体的22066392bp位置(图3)。
25.根据重测序的结果,于水稻7号染色体的22066392bp左右两侧及t-dna分别设计引物,进一步验证t-dna的插入位点。相关引物序列如下:
26.gsp-f:5'-ggtctcggccttggtat-3';
27.gsp-r:5'-gcgacagggttcgttct-3';
28.tsp-f:5'-ggcggtaacaagaaaggga-3'。
29.分别用gsp-f/r与tsp-f/gsp-r引物对扩增zh11与lg7突变体的dna,对扩增片段进行测序鉴定,确定t-dna的插入位置,从图4中可以看出t-dna插入于osfbx250基因起始密码子前的-21bp处,且osfbx250基因在突变体lg7的幼穗中表达水平显著高于对照zh11。
30.实施例2lg7基因的功能分析
31.提取水稻zh11的rna,并将其反转录成cdna,利用引物对:
32.f1:5'-agaggatccggcagcgagaagccg-3'(bamh i),
33.r1:5'-gagggtaccccagcagaagcagcc-3'(kpn i);
34.f2:5'-gatactagtccagcagaagcagcc-3'(spei),
35.r2:5'-ttcgagctcggcagcgagaagccg-3'(sac i);
36.分别扩增出osfbx250基因的cdna片段,通过上述相应的限制性内切酶酶切后连入ptck303载体,构建出osfbx250基因的干扰表达载体osfbx250-ptck303。采用农杆菌eha105介导的遗传转化方法,将干扰表达载体导入正常粳稻品种zh11中。
37.采用kod高保真dna聚合酶以zh11的cdna为模板,利用引物对:
38.5'-acgggggacgagctcggtaccatggccggcgacgaaatggcctcgc-3',
39.5'-tcgtcgactctagaggatcctcatggccgcgtcagcttcccgccc-3';
40.扩增出osfbx250基因的cdna序列,通过同源重组技术将osfbx250基因的cdna插入pcambia1301-35s-nos载体的多克隆位点5'kpni/bamhi 3'之间,构建出osfbx250基因的超表达载体pcambia1301-35s-osfbx250-nos。采用农杆菌eha105介导的遗传转化方法,将干扰表达载体导入正常粳稻品种zh11中。
41.将经潮霉素溶液筛选48小时得到的所有转基因幼苗移栽于温室中种植,阳性植株单株收种,直至t2代鉴定出纯合的转基因植株,即分别得到osfbx250基因的干扰株系与超表达株系。
42.osfbx250基因超表达植株籽粒的粒长、粒宽、粒厚和粒重均显著高于对照zh11,且osfbx250基因干扰植株籽粒的粒长、粒宽和粒重均显著低于对照zh11(图5),表明osfbx250基因正调控水稻籽粒的大小。
43.实施例3lg7基因的敲除分析
44.根据osfbx250基因的cdna序列,选择如下2个靶标进行crispr敲除:
45.target 1:gataccaaggccgagaccggagg,
46.target 2:ggaggacgatacctcattcaagg。
47.并于武汉伯远生物科技有限公司进行crispr敲除载体的构建及遗传转化,获得osfbx250基因在zh11背景的敲除株系。取敲除株系叶片,提取dna,采用如下引物对靶序列进行pcr扩增:
48.5'-gcccttcgtcggcttcctcg-3',
49.5'-ccggtgacgaaaacactaggctc-3'。
50.将扩增产物切胶回收后,进行测序分析,结果显示有4个株系敲除成功,分别是c10、c12、c14与c15,且敲除株系籽粒的粒长、粒宽、粒重均显著低于对照zh11(图6),进一步证明了osfbx250基因对水稻籽粒发育的影响。
51.上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1