一种新型IL-15超级激动剂融合蛋白的制作方法
一种新型il-15超级激动剂融合蛋白
技术领域
1.本发明涉及分子生物学技术领域,尤其涉及一种新型il-15超级激动剂融合蛋白。
背景技术:
2.在过去的几十年中,癌症一般是通过化学疗法、放射疗法、靶向疗法和手术进行治疗,但免疫疗法一直是基础和临床研究中一个快速发展的领域。通过阻断t淋巴细胞上的pd-1和ctla-4等共抑制受体的抗体对淋巴细胞的去抑制可有效治疗晚期癌症。新出现的证据表明,基于细胞因子的治疗也有望用于治疗癌症和病毒性疾病。白细胞介素(il)-15已成为免疫疗法中极具前景的候选药物之一,目前正在进行临床试验,以测试其治疗晚期癌症患者的疗效。il-15对于自然杀伤(nk)、nkt和记忆(m)cd8
+
t细胞的发育和功能至关重要,目前正在研究作为治疗癌症的免疫治疗剂。
3.然而,游离il-15具有半衰期短和体内生物活性有限等缺点,这需要进行频繁的团注。另外,异源三聚体il-15受体复合物由独特的il-15rα亚基、il-2/il-15rβ和常见的γ链/il-2rγ亚基组成。与il-2不同,il-15以高亲和力结合il-15rα,il-15rα内含一个sushi结构域,能与il-15结合,并且是使结合后的il-15发挥生物学功能所必需的。然后与由il-2/il-15rβ和常见γ链/il-2rγ亚基组成的复合物结合,在同一细胞(顺式呈递)或不同细胞(反式呈递)上表达。最近,受il-15的自然反式呈递的启发,通过在溶液中复合il-15及其高亲和力受体α(il-15rα)来产生il-15超激动剂,提高了基于il-15的潜力肿瘤免疫治疗。与单体il-15相比,il-15超激动剂显示出令人鼓舞的优势,例如由于延长半衰期而维持高循环浓度和更有效地刺激nk和cd8
+
t效应淋巴细胞。到目前为止,基于构型修饰的重组il-15超激动剂融合蛋白有三种不同形式。第一种,rli(il-15-linker-il-15rαsushi domain),是由人il-15与人il-15rα的高亲和力il-15结合域(sushi domain)通过接头组成的重组il-15超激动剂;第二种,il-15/il-15rα-igg1-fc复合物,是通过高亲和力非共价结合在溶液中偶联il-15和il-15rα-igg1-fc嵌合体来产生。第三种,alt-803(il-15n72d/il-15rα-igg1-fc嵌合体),是非共价结合了人il-15突变体(il-15n72d)和融合了igg1 fc结构域的人il-15rα的sushi结构域的组成的。
4.迄今为止,il-15超级激动剂,主要是alt-803,正在进行10多项临床试验,以测试其在晚期实体或血液恶性肿瘤患者中的安全性和潜在疗效。2022年5月份immunitybio公司将il-15超激动剂alt-803(n-803)的生物制品许可申请(bla),用于对卡介苗芽孢杆菌(bcg)无反应的高危非肌肉浸润型的膀胱癌(cis、乳头状病灶)患者治疗。
5.另外,已有不同的国内外研究团队基于上述三种超激动剂,创新改进了不同融合蛋白,激活记忆样nk(ciml-nk)的融合蛋白hcw9201(结合了il12、il15/il15 rasu和il18)、hcw9207(在hcw9201基础上添加了抗cd16抗体)和18/12/txm(结合了il12、il15/il15rasu和il18);目前正在i期临床研究中产品是由il-15/il-15rα异二聚体fc融合组成。靶向纤连蛋白剪接异构体的单克隆抗体f8与il15或il15/il15 rasu形成f8-f8-il15和f8-f8-sd-il15。甚至有学者研发了一种通过使用模块化羊驼单域抗体vhh配体来发现细
胞因子替代激动剂的策略,将vhh和scfv用于人白介素2/15、i型干扰素和白介素10受体,生成了单链双特异性配体的组合矩阵,这些配体表现出多种功能活性,包括通过替代干扰素产生对sars-cov-2的有效抑制。
6.多项研究表明,与血清中的游离il-15相比,il-15sa的半衰期延长(约20h vs约1h),并促进il-15对nk的反应增强约5-50倍和记忆cd8
+
t细胞增殖、细胞因子分泌和细胞毒功能,而单独的il-15则没有。很多情况下,药物的分子量越大,其半衰期越长,延长药物半衰期方法主要有偶联人血清白蛋白(hsa),结合内源hsa,peg化,偶联fc和大分子蛋白质工程等。上述的多种超激动剂则利用了增大融合蛋白分子量或是添加igg1 fc结构域达到体内的半衰期延长,igg的长循环半衰期(≈2-3周)和缓慢清除部分归因于它们与fcrn的独特相互作用。特别是利用偶联hsa和fc结构域,其实fc抗体工程或hsa都是通过与fcrn结合才使药物的半衰期延长。当igg或hsa被极化上皮细胞或内皮细胞吸收时,它们被包装成内体,在穿过细胞时酸化。在弱酸性条件(ph6.0-6.5)下环境中,hsa和igg fc结构域上的组氨酸被质子化并与fcrn结合,其同源轻链β2微球蛋白(β2m)被激活。β2m与fcrn的缔合是与igg和hsa结合所必需的。然后将这些hsa或igg-fcrn-β2m复合物从溶酶体中运输出去,并被循环回到它们进入的膜或转运到相反的膜上,在那里它们在遇到细胞外环境的生理条件(ph7.4)时被释放。靶蛋白重新释放至血液循环中,达到延长半衰期的目的。同时,通过人fcrn、hsa和igg fc的结合晶体结构(pdb 4n0u)进一步明确了三者的结合位点。
7.目前商业化的治疗性抗体药物和临床阶段的抗体药物基本上含有fc结构域。另外,普通重组il-2体内半衰期仅为6.9min,而重组il-2/fc融合蛋白体内循环半衰期则延长了近700倍。全长单克隆抗体(mab)的大尺寸(≈150kda)也引起了一些担忧。专利cn113321740a介绍了一种il15/il15 ra sushi融合了人igg4fc的融合蛋白,显示出高效的细胞因子活性,通过人igg4fc延长了il-15的半衰期,但该融合蛋白在非还原或还原条件下分子量分别在150kda与250kd,80kda与100kda之间。最近的报告表明,全长mab的结构可能使它们在结合具有空间位阻的表位方面的效率降低,以及在有效渗透到组织中并在血管化不良的区域获得靶点。较小的尺寸(≈50kda)和抗her2 fab显示出增强的对正常组织和实体瘤的渗透。此外,有证据表明,较小的部分可以与隐蔽的表位结合。然而,构建功能蛋白(如细胞因子)融合蛋白时要考虑融合子的分子量,其中hsa的分子量约为66kda,fc结构域分子量大约为25-26kda。当与fc结构域或hsa融合,会引起分子量过大、效应功能降低、形成多聚体、抗原介导的特异性清除等不足。
8.为此,我们提出一种新型il-15超级激动剂融合蛋白。
技术实现要素:
9.本发明为了解决如何在取代传统的fc结构域时保持延长融合蛋白体内循环半衰期;且显著降低融合蛋白的分子量,制备生产简单、成本低;更容易通过血管壁,穿透实体瘤,有利于肿瘤的治疗的问题,在于提供一种新型il-15超级激动剂融合蛋白。
10.本发明采用的技术方案如下:
11.本发明第一方面提供一种新型il-15超级激动剂融合蛋白,包括通过连接子多肽组合连接的第一结构单元、第二结构单元和第三结构单元;
12.所述第一结构单元包含il-15;
13.所述第二结构单元包含可溶性il-15受体αsushi-结合结构域;
14.所述第三结构单元包含连接子。
15.进一步地,所述il-15是包括n72d突变的变体il-15蛋白。
16.进一步地,所述连接子的序列如seq id no:1、seq id no:2或seq id no:3所示。
17.进一步地,所述连接子的序列插入有短fcrn结合肽或人白蛋白源多肽。
18.进一步地,所述融合蛋白的羧基端引入短fcrn结合肽或人白蛋白源多肽,所述短fcrn结合肽通过人fcrn蛋白、hsa蛋白和igg fc蛋白的结合晶体结构确定三者的结合位点。
19.进一步地,所述短fcrn结合肽的序列如seq id no:4、seq id no:5、seq id no:6或seq id no:7所示。
20.进一步地,所述人白蛋白源多肽的序列如seq id no:8序列。
21.本发明第二方面提供一种核酸,编码如上述本发明第一方面任一项所述的一种新型il-15超级激动剂融合蛋白。
22.本发明第三方面提供一种表达载体,包括本发明第二方面所述的一种核酸。
23.本发明第四方面提供一种宿主细胞,其特征在于,包本发明第二方面所述的一种核酸或本发明第三方面所述的一种表达载体。
24.本发明第五方面提供一种制造上述本发明第一方面任一项所述的一种新型il-15超级激动剂融合蛋白的方法,包括以下步骤:
25.步骤s1:在适合表达所述新型il-15超级激动剂融合蛋白的条件下培养权利要求10所述的宿主细胞;
26.步骤s2:收集并纯化所述新型il-15超级激动剂融合蛋白。
27.本发明第六方面提供根据上述本发明第一方面任一项所述的一种新型il-15超级激动剂融合蛋白、根据本发明第二方面所述的一种核酸、根据本发明第三方面所述的一种表达载体、根据本发明第四方面所述的一种宿主细胞、根据本发明第五方面所述的制造一种新型il-15超级激动剂融合蛋白的方法在治疗癌症或自身免疫疾病的用途。
28.本发明的有益效果是:
29.1、本发明通过优化il-15蛋白(il-15n72d)与il-15受体αsushi-结合结构域(il-15rαsu)的连接肽序列,引入fcrn结合多肽,取代传统的fc结构域,却保持了延长融合蛋白体内循环半衰期;且显著降低融合蛋白的分子量,制备生产简单、成本低;更容易通过血管壁,穿透实体瘤,有利于肿瘤的治疗等优点。
30.2、本发明的il-15超级激动剂能高效保进淋巴细胞增殖,诱发产生细胞毒,具有生物学活性。
31.3、本发明的il-15超级激动剂能诱导形成记忆样nk细胞(ciml-nk),这种ciml-nk细胞的活性显著抑制了肿瘤细胞的生长,减轻肿瘤负荷。
附图说明
32.图1为实施例1和实施例4中的il-15超级激动剂蛋白结构示意图;
33.图2为实施例1中的fcrn结合肽对应iggfc蛋白和人血清白蛋白(hsa)的fc受体蛋白结合区域;
34.图3为实施例1中的不同il-15超级激动剂结构示意图;
35.图4为实施例2中的不同il-15超级激动剂蛋白sds-page图;
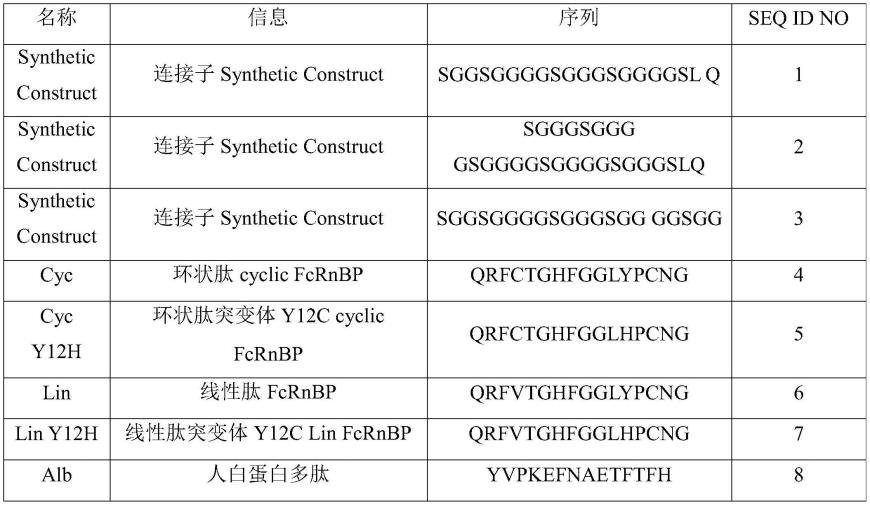
36.图5为实施例3中的小鼠淋巴母细胞系ctll-2在不同浓度il-15超级激动剂的增殖情况;
37.图6为实施例4中的bsa和fcrn-β2m融合蛋白sds-page图,其中m为蛋白maker,泳道1:bsa蛋白,泳道2:fcrn-β2m融合蛋白;
38.图7为实施例4中的elisa分析不同il-15超级激动剂在ph6和7.4下对fcrn的结合能力;
39.图8为实施例4中的不同il-15超级激动剂浓度对a-375的细胞毒性;
40.图9为实施例4中的不同e:t比率的il-15超级激动介导的细胞毒性;
41.图10为实施例5中的不同il-15超级激动剂联合il12、il18对nk细胞的增殖;
42.图11为实施例5中的中的il-15超级激动剂(alb)联合il12、il18增强nk细胞分泌ifn-γ
43.图12为实施例6中的小鼠肺癌模型实验设计;
44.图13为实施例6中的m-nsg小鼠肿瘤进展及体内生物发光成像;
45.图14为实施例6中的m-nsg小鼠肿瘤负荷。
具体实施方式
46.以下对至少一个示例性实施例的描述实际上仅仅是说明性的,决不作为对本发明及其应用或使用的任何限制。基于本发明中的实施例,本领域普通技术人员在没有作出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
47.实施例1:核苷酸序列的获得与优化
48.根据已有数据库中蛋白氨基酸序列的信息,查阅il-15(登录号:np_000576)及il-15突变体(il-15n72d)、il-15ra(登录号:np_002180)的序列信息,其中il-15ra的sushi结构域(il-15ra sushi,aa 1-66)。根据需要,插入所述两个蛋白之间的连接子,参见图1,连接子为常规柔性连接子参见表1中如连接子seq id no:1、seq id no:2或seq id no:3;或是在上述三种连接中再插入短fcrn结合肽序列参见表1中seq id no:4、seq id no:5、seq id no:6或seq id no:7,或alb人白蛋白源多肽参见表1中的seq id no:8。进一步在各上述融合子中设计所需羧基端引入短fcrn结合肽序列参见表1中seq id no:4、seq id no:5、seq id no:6或seq id no:7或alb人白蛋白源多肽参见表1中的seq id no:8,其中上述结合肽序列是通过人fcrn、hsa和igg fc的结合晶体结构(pdb 4n0u)进一步明确了三者的结合位点,参见图2。作为对照,参考专利wo2008143794的方法创建il-15n72d:il-15rαsu/fc融合复合物(alt-803),具体的融合蛋白结构示意图参见图3。为了实现融合蛋白的分泌表达,在结构蛋白前面加入高分泌的信号肽序列(tpa6 signal,id no:4),同时在融合蛋白羧基端加入(his)6纯化标签,方便后续对融合蛋白的纯化。
49.将上述各目标氨基酸序列转化为核苷酸序列,并针对可能影响抗体在哺乳动物细胞中表达的一系列参数:密码子偏好性、gc含量(即dna的4种碱基中鸟嘌呤g和胞嘧啶c所占的比率)、cpg岛(即cpg双核苷酸在基因组中密度较高的区域)、mrna的二级结构、拼接位点、前成熟polya位点、内部chi位点(基因组中一段短的dna片段,在该位点附近发生同源重组的几率增加)或者核糖体结合位点、rna不稳定序列、反向重复序列及可能干扰克隆的限制
性酶切位点等进行优化;同时增加了可能会提高翻译效率的相关序列,例如kozak序列、sd序列,以及终止密码子。在融合蛋白的5’端分别设计上根据氨基酸序列优化而得的编码信号肽的核苷酸序列;此外,还对融合蛋白核苷酸序列的3’端分别加上终止密码子。
50.对所有基因进行密码子优化合成这些基因并亚克隆到哺乳动物表达载体pcdna3.1(+)(genscript)中。
51.表1序列信息
[0052][0053]
实施例2:包含il-15/il-15rαsushi和结构域连接子的构建、表达和纯化
[0054]
采用pcdna3.1(+)载体作为表达所述融合蛋白专用载体。pcdna3.1(+)载体含有融合蛋白所使用的启动子cmv promoter、真核筛选标记g418标签和原核筛选标签氨苄西林(ampicilline)。基因合成得到各种融合蛋白的核苷酸序列,用hind iii和xho i对载体和目的片段进行双酶切,回收后通过dna连接酶进行酶连,并转化大肠杆菌感受态细胞dh5α,挑选出阳性克隆并进行质粒提取和酶切验证,并抽提质粒并进行转染;在转染前一天,用无血清free style 293表达培养基(gibco)培养可悬浮培养的hek-293f细胞;在转染当天,将细胞以每毫升1
×
106个细胞的浓度接种到新鲜培养基中,500μg l-1
的质粒dna与2mg l-1
pei(polysciences)混合并在opti prosfm培养基(4%终体积);培养4-5d后,收集上清液并如前所述进行纯化。其中纯化是以利用akta fplc的亲和纯化柱ni-nta进行,并通过鲎试剂盒对纯化的蛋白质进行内毒素检测,确认内毒素水平低于0.01eu ml-1
。如前所述,通过sds-page评估蛋白质纯度,使用nanodrop(thermo scientific)通过280nm处的吸光度测定蛋白质浓度,其中alt-803购于altor bioscience(lot#01062016)。
[0055]
经sds-page检测如图4所示,所得到的融合蛋白分子量正确,且纯度较高,出现两条分子量比较近的蛋白,可能是由于糖基化水平不同引起的。
[0056]
实施例3:il-15超级激动剂对体外细胞增殖的影响
[0057]
各il15融合蛋白功能评估,是通过添加细胞因子后对细胞增殖的影响。
[0058]
小鼠淋巴母细胞系ctll-2购自atcc,在96孔板中使用补充有10%fbs的prmi-1640接种的小鼠淋巴母细胞ctll-2细胞(15,000个细胞/孔)在37℃下与指定浓度的il15融合蛋
白一起孵育72h。按照产品的说明,使用cell counting试剂盒8(wst-8/cck8)(ab228554)测定细胞增殖,以重组alt-803做对照。如图5所示,能够促进ctll-2细胞的细胞增殖,从而证明il-15活性。与alt-803的活性相比,各il-15超级激动剂的活性有所变化,其中lin-cycy12h活性与alt-803相近,而alb、cyc-cycy12h和alb-cycy12活性略高于alt-803,可能由于多肽结构影响稳定性。
[0059]
实施例4:各il-15超级激动剂与fc功能评估
[0060]
(1)与fcrn的结合活性
[0061]
根据已有数据库中蛋白氨基酸序列的信息,查阅igg receptor fcrn large subunit p51(登录号6wna_a)、beta-2-microglobulin(登录号:1a1m_b)的序列信息,根据需要,插入见图1所述两个蛋白之间的连接子,连接子可以是常规柔性连接子如连接子seq id no:1,在结构蛋白前面加入大肠杆菌细胞周质的信号肽序列(ompa signal,登录号:aaa82946),同时在融合蛋白羧基端加入(his)6纯化标签,方便后续对融合蛋白的纯化,上述元件组合成:ompa signal-linker-fcrn-6*his结构。同时根据大肠密码子偏好性、gc含量(即dna的4种碱基中鸟嘌呤g和胞嘧啶c所占的比率)、mrna的二级结构等进行优化合成,并克隆到pet30a+载体中,挑选出阳性克隆并进行质粒提取和酶切验证,并转化到大肠杆菌表达宿主菌bl21(de3)。用lb培养基37℃培养至大约od600约0.6时加入终浓度0.1mm的iptg至诱导蛋白质表达。低温22℃诱导14h后将细胞沉淀并在1xpbs/1%(v/v)triton x100中超声裂解。其中纯化是以利用akta fplc的亲和纯化柱ni-nta进行。所有纯化的蛋白质都储存在无菌的1xpbs中,并补加终浓度为0.1mm苯甲基磺酰氟(pmsf)蛋白酶抑制剂,在4℃下短期储存(《2周),或在-20℃下用25%(v/v)甘油长期储存。
[0062]
将上述纯化好的fcrn-β2m蛋白用50mm的碳酸盐包被缓冲液(ph9.6)溶解抗原,使抗原浓度为2μg/ml,加100μl/孔到96孔板。第二天弃去包被液后,用pbst洗涤3次,每孔加入150μl 1%bsa 37℃封闭1h。然后将全长抗体或各il-15融合蛋白加入ph7.4或ph6封闭溶液中,平板室温育1-2h。用pbst洗涤3次,通过与1:5000稀释的蛋白质l-hpr温育1h,然后用邻苯二胺(opd)显色作为检测结合的il-15融合蛋白。对于竞争性elisa,根据产品的说明书将il-15融合蛋白与nhs-生物素生物素化,然后用含有人血清白蛋白(sigma-aldrich)或10倍过量的未标记人血清(sigma)的pbst+3%(w/v)bsa稀释,生物素化的il-15融合蛋白用1:10000链霉亲和素-hrp(abcam)和opd法检测。在分光光度仪450nm(a450)处测量吸光度,所有实验三个重复,并用graphpad prism进行统计学分析。
[0063]
结果:纯化了含有fcrn大亚基p51和beta-2-microglobulin组成的完整fcrn-β2m融合蛋白,其蛋白分子量约为43kda,参见图6。通过elisa在ph6.0和7.4下评估不同il-15超级激动剂结合。
[0064]
fcrn-β2m,以alt-803、hsa作为对照,参见图7,看出环状fcrnbp-fc、线性fcrnbp-fc或fcrnbp-alb表现类似于全长igg(alt-803)或白蛋白(hsa)的结合能力,其kd值在ph值6.0时约为0.3μm-1.3μm,参见表2,其中alb、l-cyc和cyc-cycy12h的kd值接近白蛋白(hsa)的结合能力;而在ph7.4时结合亲和力很低,无法检测到(如lin)。同时,未添加fc受体(fcrn)结合多肽的il-15超级激动剂(l)均无法与fcrn结合,表明这边功能化的il-15超级激动剂确实是由于结合多肽介导的相互作用。
[0065]
表2不同il-15超级激动剂在ph6.0结合fcrn的kd值
[0066]
蛋白kd(μm)alt-8030.9
±
0.1lndcyc1.3
±
0.1alb0.3
±
0.1lin0.06
±
0.05cyc-cycy12h0.4
±
0.03l-cyc0.4
±
0.04hsa0.3
±
0.04
[0067]
(2)细胞毒性试验(51铬释放试验)
[0068]
pbmc制备及扩增:取3位健康志愿者的血液,用ficoll(购自ge)分离pbmc,细胞计数后将细胞密度用x vivo 15(购自lonza公司)培养基调整到1.5
×
105/ml,接种到6孔板中(终体积2ml),然后加入不同浓度的上述融合蛋白,进行体外培养72h后,收获pbmc,重新调整数量测试对人恶性黑色素瘤细胞系a-375的细胞毒性。
[0069]
人恶性黑色素瘤细胞系a-375,购自atcc。准备10%fbs的dulbecco's modified eagle'smedium(dme h-21 4.5g/liter glucose)的完全培养基培养a-375细胞至对数生长状态,取一定细胞数量沉淀后计数,靶细胞(1.0
×
106)用
51
cr放射性标记37℃下标记2h。细胞洗涤两次后,将5000个靶细胞/孔与50,000个效应细胞(e:t=10:1)混合,并添加到96-v底板的每个孔中,终体积为250μl/孔,在5%co2的培养箱中在37℃下孵育4h。在4h孵育结束时,将板在234
×
g下旋转2min,然后使用多通道移液器将50μl从每个孔中的上清液转移到lumaplate-96。使用topcount nxt读数器将每个上清液中存在的
51
cr发射的辐射量确定为每分钟计数(cpm)。
[0070]
结果:参见图8所示,最高浓度(1nm)下,所有11种形式(游离il15或10种不同fcrn结合肽il15/il15rα复合物)在激活淋巴细胞以介导a-375肿瘤溶解方面同样有效;然而,在较低浓度下,与l和alt-803融合蛋白相比,alb、alb-cycy12h和cyc-cycy12h融合蛋白的淋巴细胞活化显著增强。可能由于环状或alb的结合肽相互作用导致更强的信号传导,从而使pbmc衍生的t细胞和nk细胞更具溶细胞性,因此可以预期此类结果。可溶性il15与l或lin融合蛋白之间没有显著差异可以解释为在pbmc群体中将il15呈递给t和nk的辅助细胞(如dc细胞)上存在天然il15rα细胞。
[0071]
另外,在保持1nm的融合蛋白恒定下,测定不同有e:t比率下,其杀伤效果的细胞毒性,与刚才的数据类似,alb、alb-cycy12h和cyc-cycy12h在pbmc中比alt-803更能刺激细胞毒性,参见图9。
[0072]
实施例5:il-15超级激动剂对细胞因子诱导的记忆样nk细胞的诱导,nk细胞分离和扩增
[0073]
取3位健康志愿者的血液,用ficoll(购自ge)分离pbmc,细胞计数后将细胞密度用x vivo15(购自lonza公司)培养基调整到2.0
×
106/ml,接种到6孔板中(终体积2ml),使用cloudz human nk cell expansion kit(cld004,r&d systems)进行体外扩增nk,并在第14d收获细胞,并调整细胞浓度至2.0
×
106/ml,并使用rhil-12(10ng/ml)+rhil-18(50ng/ml)+rhil-15(50ng/ml)预活化16h,或用不同融合蛋白替换rhil-15,并以低剂量rhil-15
(1ng/ml)作为对照。离心收集细胞,和pbs洗2遍,制备成不同融合蛋白组合的记忆样nk细胞,并进行预处理或是流式标记。其中细胞因子刺激16h后,加入20ul cell activation cocktail(brefeldin a和莫能菌素)处理5h,进行cd3、cd56、cd16染色。对细胞内的ifn-γ染色之前,将细胞固定和破膜。
[0074]
其中预处理和清洗后的nk细胞,用低剂量rhil-15(1ng/ml)完全培养基x vivo 15维持培养7d,并采用cfse标记nk细胞进行流式检测,以评估不同il-15超级激动剂对nk细胞增殖的影响。在bd facsariaiii流式细胞仪上采集细胞,并使用flowjo 9.3.2版(tree star)软件进行分析。
[0075]
结果显示:采用不同il-15超级激动剂与il12、il18诱导形成的记性样nk细胞(ciml-nk),其结果与上述对小鼠淋巴母细胞系ctll-2的增殖情况类似,相对于纯il15激活,用il12/18/il15超级激动剂激活诱导强烈的增殖;另外,alb和cyc-cycy12h诱导nk细胞增殖高于alt-803,参见图10。
[0076]
另外,通过流式分析了nk(cd3-cd56
+
)的不同表面标记物的表达情况,如nkg2c、nkg2d、cd45ro和cd107a
+
等,不同的il-15超级激动剂诱导所形成的nk表型差异不大。但有一个共同的表征,就是上调ifn-γ的表达量,大概提高50倍左右,参见图11。
[0077]
实施例6:il-15超级激动剂在体内诱导记忆样nk细胞抗肿瘤活性
[0078]
人肺腺癌荧光素酶标记细胞a549-luc,购自atcc。准备10%fbs的rpmi 1640的完全培养基培养a549-luc细胞至对数生长状态,取一定细胞数量沉淀后计数,并调整细胞浓度至2.0
×
106/ml。
[0079]
8-12周雄性和雌性nod-prkdc
scid il2rg
em1
/smoc m-nsg小鼠购买于上海南方模式生物科技股份有限公司,并且在室内适应7d,然后进行研究。在第-1d,小鼠通过尾静脉注射a549-luc总细胞量约为4.0
×
105,在第0、2和4d每天经尾静脉输注不同来源nk细胞,组别包括了溶媒、10.0
×
106对照(1ng/ml il-15中的nk细胞)或用il-12/15/18激活的nk细胞(10ng/ml il-12、50ng/ml il-15和50ng/ml il-18)或18/12/il15融合蛋白(38nm)经尾静脉注射给予小鼠(来自2个独立实验,每组总共2只小鼠),试验设计及给药方式如图12所示。在注射a549-luc细胞后,通过给腹腔注射d-荧光素钾盐(d-luciferin pbs中150mg/kg,翌圣生物科技(上海)股份有限公司),3min后将小鼠放入麻醉室进行麻醉,成像前以低流量(1.0l/min)o2作载气行4%异氟醚诱导小鼠麻醉,并进行体内成镜以确认a549-luc细胞植入。在第0、3、13和19d通过体内成像监测肿瘤负荷,通过采集总光子通量(photons/s)评估效果,并在第20d用co2对小鼠实施安乐死,并收获组织并制备病理切片。
[0080]
结果显示:通过测量肿瘤生物发光随时间的变化来监测肿瘤生长,参见图13,与用溶媒或对照il15 nk细胞治疗相比,输注记忆样nk细胞改善了肿瘤控制;用il12/18/il-15超级激动剂诱导的ciml-nk在第19d表现出增强的肿瘤控制,参见图14,其中cyc-cycy12h、alb组达显著好于il12/18/15组。同时,alb-cych12h抑制效果最为明显。这些数据表明用il-12/18/il-15超级激动剂激活nk细胞可以诱导ciml-nk细胞表型,类似于由il12/18/15组合,其在体内发挥增强的肿瘤靶标的控制。
[0081]
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1