黄花苜蓿钙依赖蛋白激酶MfCDPK14蛋白及其应用
本发明属于植物基因工程领域,具体涉及黄花苜蓿钙依赖蛋白激酶mfcdpk14蛋白及其应用。
背景技术:
1、非生物胁迫(如干旱、盐碱、冷害和热害等)对植物的生长发育有不可忽视的影响,导致农作物产量下降,同时也影响林木、果树、花卉以及园艺观赏植物的正常生长和品质。因此,培育出耐逆性强的植物品种成为种植业的重要目标之一。为此,从耐逆性强的植物中分离出耐性基因成为必要之举。
2、植物体内主要存在钙调素(cam)、类钙调素蛋白(cml)、钙调磷酸酶b类似蛋白(cbl)、钙依赖蛋白激酶(cdpk)及钙/钙调素依赖型蛋白激酶(ccamk)等类型的钙感受器(batistic et al.,2012;schulz et al.,2013)。不同于其他只带有钙感受器的小分子蛋白质,cdpk和ccamk是一类植物中特异存在的蛋白激酶。在拟南芥中,4%的基因编码了蛋白激酶(champion et al.,2004;zulawski et al.,2014)。其中参与解码ca2+信号的色氨酸/苏氨酸蛋白激酶主要有三类:cdpk、ccamk及cbl互作蛋白激酶(cipk)(harper et al.,2004),其中cipk需要与cbl蛋白互作才可以参与钙信号的转导(batistic et al.,2009)。cdpk是其中最大的家族,它与ca2+结合后,可以直接把ca2+信号转换成磷酸化信号,通过磷酸化信号直接激活和调控靶蛋白。这一过程并不需要其他蛋白质的参与,这使得cdpk具有双重功能:钙感受器和应答器(harper et al.,2004)。
3、植物响应干旱胁迫最早的反应之一就是细胞质ca2+浓度增加,这一变化可以通过ca2+感受器(例如cdpk)传递至其靶蛋白,进而引起下游系列基因表达和生理生化等适应性变化,提高耐旱性。
4、cdpk还广泛参与植物对干旱胁迫的应答过程,主要包括磷酸化各类离子通道蛋白调控气孔的开闭、与aba信号通路中的转录因子互作调控下游基因表达、调控ros的清除及渗透物质的积累等。其中,气孔关闭是植物应对干旱胁迫的关键反应。当受到外界刺激后,保卫细胞内ca2+浓度发生变化,阴离子与钾离子通道蛋白被激活从而使离子流出,渗透势上升,随之细胞收缩。拟南芥中的多个cdpk被证明参与了该过程的调控。atcpk3/6/21/23在非洲爪蟾卵母细胞中异源表达时可激活s-型阴离子通道slac1和slah3,并且可以在体外磷酸化slac1(brandt et al.,2012;brandt et al.,2015;geiger et al.,2011;geiger etal.,2010;scherzer et al.,2012),但它们间可能存在功能冗余,在cpk3、cpk6及cpk23突变体的离体枝条上只能观测到微弱的气孔表型,而在cpk3/6双突变体以及cpk5/6/11/23四突变体中aba诱导的气孔关闭则显著减弱(brandt et al.,2015;geiger et al.,2010;mori et al.,2006)。相反的,atcpk9/33被鉴定为aba诱导气孔关闭的负调节因子(chen etal.,2019;li et al.,2016)。此外,cdpk还可以通过调控k+通道调控气孔关闭,如cpk10突变体的干旱敏感表型可能是由于保卫细胞内k+通道蛋白的活性增加造成的(zou et al.,2010)。
5、干旱胁迫可以引发体内大量的基因发生转录变化,其中一部分是由cdpk介导的。例如,atcpk32与atabf4互作并将其磷酸化对于aba诱导abf4的转录激活具有重要作用(choi et al.,2005),atcpk4可以磷酸化atabf2并激活其转录活性(lu et al.,2013)。此外,atabf1/4还可以作为atcpk4/11的底物,过表达atcpk4/11可以增强胁迫响应基因表达来增强耐旱性,而突变体表现出相反的表型(zhu et al.,2007)。此外,植株还可以通过调控ros的清除及渗透物质的积累来抵御干旱胁迫。拟南芥atcpk8通过磷酸化atcat3并激活atcat3的酶活来提高耐旱性(zou et al.,2015),水稻oscpk10也可以通过增加cat的积累增强耐旱能力(bundo et al.,2017)。另外,枳ptrcdpk10通过与ptrapx互作并使其磷酸化,进而降低ros的积累来参与对渗透和干旱胁迫的响应(meng et al.,2020)。水稻oscpk9通过调控气孔的关闭和提高渗透调节能力来增强耐旱性(wei et al.,2014)。
6、cdpk存在于高等植物、绿藻、卵菌和原生生物中,但不存在于动物和真菌中,而crk仅存在于植物中(hrabak et al.,2003;valmonte et al.,2014)。cdpk包含四个结构域:n端可变域、色氨酸/苏氨酸蛋白激酶域、抑制连接域和类钙调素域(cam-ld)(hrabak etal.,2003)。cdpk家族成员间氨基酸的差异主要集中在n端可变域内,它可能参与cdpk与靶蛋白的互作识别。
7、cdpk和crk在植物中的广泛分布暗示其具有激活多种底物的潜力,目前已经发现并鉴定到了大量的底物,包括各类转录因子、离子通道蛋白、代谢酶及转运蛋白等。研究发现它们通过磷酸化特异的下游底物,在调控植物生长发育及响应各类胁迫等过程中发挥重要作用。
8、苜蓿是优质的豆科牧草,主要种植于我国的干旱和半干旱地区,尤其集中在西北和华北。干旱是苜蓿生产中最主要的逆境因素,严重影响了产量和品质。黄花苜蓿是一种具有强耐旱性的草种,从中克隆抗旱调控基因,并揭示其调节耐旱性的分子机制,是苜蓿育种的重要基因资源。这为农林植物的抗逆分子育种提供了坚实的基础,对于探索苜蓿的耐旱机制以及生产利用和培育耐旱新品系具有重要的指导意义。
技术实现思路
1、为克服现有技术的缺点和不足,本发明的目的是提供一种提高植物耐旱性的黄花苜蓿钙依赖蛋白激酶mfcdpk14蛋白及其编码基因和应用。
2、本发明的目的通过下述技术方案实现:
3、第一方面,本发明请求保护一种黄花苜蓿钙依赖蛋白激酶mfcdpk14蛋白,该蛋白的氨基酸序列为(a)或(b):
4、(a)如seq id no.1所示的氨基酸序列;
5、(b)由seq id no.1所示的氨基酸序列经取代、缺失或添加一个或几个氨基酸而形成的具有同等功能的序列。
6、第二方面,本发明请求保护编码上述黄花苜蓿钙依赖蛋白激酶mfcdpk14蛋白的mfcdpk14基因;该mfcdpk14基因的核苷酸序列为(1)或(2):
7、(1)如seq id no.2所示的核苷酸序列;
8、(2)与(1)所述的核苷酸序列具有90%以上的同源性且具有同等功能的核苷酸序列。
9、第三方面,本发明请求保护包含上述mfcdpk14基因的生物材料,该生物材料为表达盒、重组载体、细胞系或重组微生物。
10、进一步,所述的重组载体为重组表达载体,该重组表达载体是通过将上述的mfcdpk14基因与植物表达载体连接得到的。
11、所述植物表达载体包括但不限于双元农杆菌载体以及用于单子叶基因枪转化的载体;所述的双元农杆菌载体包括pbi121、pcambia系列载体;所述植物表达载体还可包含外源基因的3’端非翻译区域,即包含聚腺苷酸信号和任何其它效应mrna加工或基因表达的dna片段;
12、构建含有mfcdpk14基因的重组表达载体时,在其转录起始核苷酸前可使用任何一种强启动子或诱导型启动子;所述强启动子或诱导型启动子包括但不限于泛素(35squtin)启动子和花椰菜花叶病毒(camv)35s启动子,它可单独使用或与其他的植物启动子结合使用;此外,构建含有mfcdpk14基因的重组表达载体时,还可使用增强子,这些增强子区域包括但不限于atg起始密码子和邻接区域。起始密码子必须与编码序列的阅读框相同,以保证整个序列的翻译。翻译控制信号和起始密码子可以是多种不同的来源,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或来自结构基因。
13、为了便于对转基因植株进行鉴定及筛选,可对所使用的植物表达载体进行加工,包括加入或替换植物可选择性标记。可使用的选择性标记包括编码抗除草剂的酶的基因或具有抗性的抗生素标记物;所述的除草剂包括草丁膦、草甘膦等,所述的抗生素包括硫酸卡那霉素、潮霉素、庆大霉素等。从转基因植物的安全性考虑,可以不加任何选择性标记基因,直接以逆境筛选转化植株。
14、本发明的具体实施方式中,所述的植物表达载体选择pcambia3301。
15、所述重组表达载体的构建方法具体包含如下步骤:
16、(1)引物设计
17、引物①为mfcdpk14基因扩增上游引物mfcdpk14-f:
18、5’-atgggttgtcacggcagc-3’;
19、引物②为mfcdpk14基因扩增下游引物mfcdpk14-r:
20、5’-ttaaagtaatggtccctgg-3’;
21、引物③为引入nco i酶切位点的上游引物mfcdpk14-3301-f:
22、5’-cacgggggactcttgaccatggacatgggttgtcacggcagc-3’;
23、引物④为引入bsteiⅰ酶切位点的下游引物mfcdpk14-3301-r:
24、5’-cggggaaattcgagctggtcaccttaaagtaatggtccctgg-3’;
25、(2)mfcdpk14基因片段的获得:
26、以黄花苜蓿的cdna为模板,使用引物①和引物②扩增mfcdpk14基因片段,并与cloning kit载体连接,构建获得peasy-mfcdpk14载体;
27、(3)含有mfcdpk14基因的重组表达载体pcambia3301-mfcdpk14的构建:
28、以peasy-mfcdpk14载体为模板,使用引物③和引物④使mfcdpk14基因片段引入nco i和bsteiⅰ两个酶切位点,并与植物表达载体pcambia3301连接,进而构建获得含有mfcdpk14基因的重组表达载体pcambia3301-mfcdpk14。
29、所述的黄花苜蓿优选为黄花苜蓿(medicago falcata l.)品种海拉尔。
30、本发明的具体实施方式中,采用上述的重组表达载体pcambia3301-mfcdpk14培育表达黄花苜蓿钙依赖蛋白激酶mfcdpk14蛋白的转基因植株,是通过将上述的含有mfcdpk14基因的重组表达载体转化植物组织,并将转化的植物组织培育成转基因植株得到的。
31、所述的含有mfcdpk14基因的重组表达载体转化植物组织可通过使用ti质粒、ri质粒、植物病毒载体、直接的dna转化,显微注射、电穿孔等各种常规或特异的遗传转化方法导入植物细胞或组织,并将转化的植物组织培育成植株。被转化的植物宿主可以是单子叶植物或双子叶植物。所述植物以苜蓿为例。具体包含如下步骤:
32、(1)用电转的方法将重组表达载体pcambia3301-mfcdpk14导入根瘤农杆菌eha105中,获得含有重组表达载体pcambia3301-mfcdpk14的农杆菌阳性菌落;
33、(2)将活化的含有重组表达载体pcambia3301-mfcdpk14的农杆菌eha105浸染苜蓿叶片,经共培养和抗生素筛选,获得转基因苜蓿。
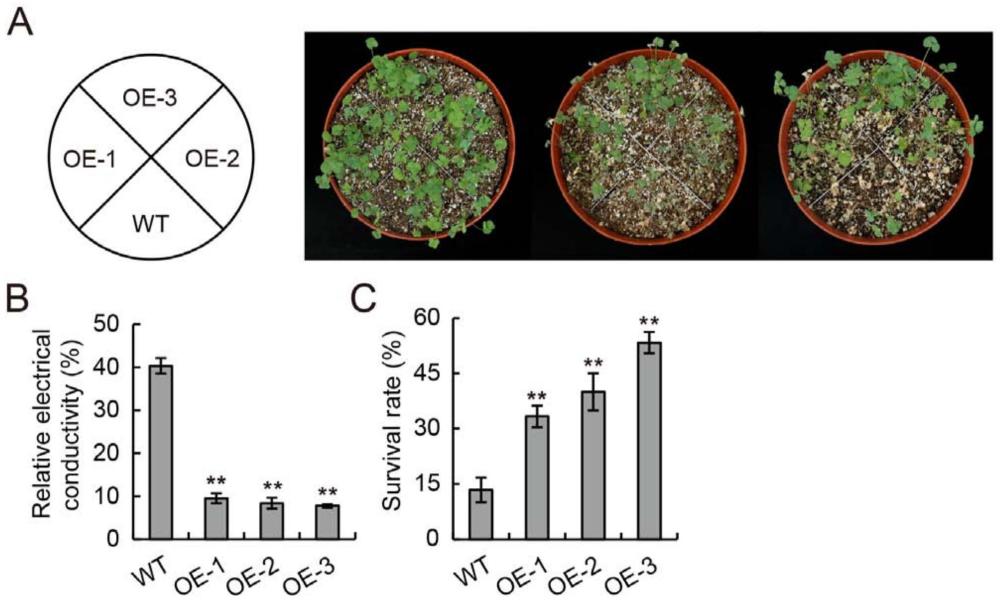
34、第四方面,本发明请求保护上述的黄花苜蓿钙依赖蛋白激酶mfcdpk14蛋白,上述的mfcdpk14基因,或上述述的生物材料在提高植物抗旱性或培育耐旱性提高的转基因植物中的应用。
35、进一步,上述的应用为在目标植物中过表达上述的mfcdpk14基因,或提高上述mfcdpk14蛋白的表达量以提高植物的耐旱性。
36、进一步,在目标植物中过表达所述mfcdpk14基因的过程为:构建如上所述mfcdpk14基因的过表达载体,通过农杆菌介导法将所述的过表达载体转入到目标植物中得到耐旱性提高的转基因植株。
37、第五方面,本发明请求保护一种提高植物抗旱性的方法,在目标植物中过表达上述的mfcdpk14基因,或提高上述mfcdpk14蛋白的表达量以提高植物的耐旱性。
38、上述的植物为单子叶植物或双子叶植物,例如苜蓿,但不限于此。
39、第六方面,本发明请求保护一种黄花苜蓿钙依赖蛋白激酶mfcdpk14启动子,该启动子的核苷酸序列如seq id no.3所示。
40、上述的黄花苜蓿钙依赖蛋白激酶mfcdpk14启动子在如下(1)或(2)中的应用也属于本发明的保护范围:
41、(1)驱动gus表达;
42、(2)调控下游基因响应逆境胁迫的表达;所述的逆境胁迫为干旱胁迫。
43、本发明相对于现有技术具有如下的优点及效果:
44、(1)本发明从黄花苜蓿中克隆了钙依赖蛋白激酶基因mfcdpk14的cdna序列,该mfcdpk14基因的表达受干旱诱导。
45、(2)本发明将获得的mfcdpk14基因与植物表达载体连接,构建了适合于双子叶植物的重组表达载体,并转化双子叶模式植物蒺藜苜蓿,转基因植株明显提高了抗旱性。
46、(3)本发明提供了利用mfcdpk14基因培育耐旱植物的方法。
- 还没有人留言评论。精彩留言会获得点赞!