白菜BrPILS5基因在提高植物花粉耐热性中的应用
本发明涉及生物,具体涉及白菜brpils5基因在提高植物花粉耐热性中的应用。
背景技术:
1、近些年来,随着人口的增长、经济的发展,温室效应越来越突出,高温造成的损失对全球粮食安全构成了重大威胁(peng et al.,2004;charles et al.,2010;chaturvedietal.,2021)。植物是固着生长,不断受到包括热害在内的环境胁迫的挑战,这对植物的生长发育造成严重威胁(ding et al.,2020)。在种子植物中,生殖发育是形成种子、完成世代交替的重要过程,已有研究表明,其是植物生命周期中对高温等环境胁迫最为敏感的发育过程,而其中,雄性生殖发育(主要为花药和花粉发育)对环境胁迫的敏感性又高于雌性生殖发育过程(begcy et al.,2019)。由于环境胁迫对花粉发育产生严重影响,进而影响了农作物种子以及以种子为产品的粮食和蔬菜等的正常生产(de storme and geelen,2014;ghadirnezhad and fallah,2014)。因此,制定出应对环境胁迫的技术措施或培育出抗逆农作物,对确保全球粮食安全至关重要。
2、生长素在花药开裂、花粉成熟、花丝伸长及花粉管生长方面具有重要的调控作用(cecchetti et al.,2008)。生长素作用于植物过程包括合成、代谢、转运及信号转导四大过程,任何一个过程发生不正常变化都会影响植物花粉的发育,例如拟南芥中生长素合成基因yuc2和yuc6双突变形成无花粉的短雄蕊(cheng et al.,2006);水稻中研究发现生长素代谢突变体dao花药不裂和花粉粒发育缺陷(zhao et al.,2013);生长素信号转导基因突变体arf17中花粉壁发育和花药开裂异常(yang et al.,2013),tir1 afb2 afb3三重突变体和tir1 afb1 afb2 afb2四重突变体中花粉早熟(cecchetti et al.,2008);生长素转运突变体pin8引起花粉外壁的发育和花粉萌发异常(dal bosco et al.,2012;ding etal.,2012),abcb1 abcb19双突变体中花粉早熟(cecchetti et al.,2015)。
3、在开花植物中,雄性生殖器官雄蕊的正常发育是成功有性生殖所必需的。在拟南芥中,通常六个雄蕊原基出现在花器官的第三轮,随后分化为雄蕊花丝和花药,在那里发生雄性减数分裂,这一早期阶段之后是一个后期发育阶段,该阶段包括雄蕊花丝的快速伸长,与花药开裂和花粉成熟相协调,越来越多的证据表明,生长素运输对于雄蕊发育的早期和晚期都是必要的(cecchetti et al.,2017)。pin家族的蛋白质是具有极性细胞分布的主要外排载体(galweiler et al.,1998;friml1.et al.,2003;paponov et al.,2005;friml,2010)。pins载体可分为一个大环和一个短环子组。大环pins(pin1、pin2、pin3、pin4和pin7)以一个大的亲水环为特征,定位在质膜上,并通过定位的膜直接转运生长素,短环pins(pin5,pin6,和pin8)不被募集到质膜,但被提议调节细胞质和内质网(er)之间的生长素稳态(mravec et al.,2009;wabnik et al.,2011)。
4、在植物细胞中还存在一个保守的细胞内生长素运输体家族,即pin-likes蛋白质家族(pils)。pils蛋白质家族是近年来被新发现及进行生理功能阐释的一类生长素运输体。该类蛋白质与pin蛋白质在序列同源性上相似性并不高,但在蛋白质高级结构上具有较高的相似性。pils蛋白质家族在拟南芥中存在7个成员,这些成员通过定位在内质网上介导细胞质内的生长素向内质网腔中运输(barbez et al.,2012)。pils蛋白是生长素依赖性调节植物生长所必需的。pils蛋白调节细胞内生长素在内质网的积累,从而调节生长素在细胞核生长素信号传导中的可用性。pils活性可能通过细胞内积累和代谢影响内源性生长素吲哚-3-乙酸(iaa)的水平(barbez et al.,2012;feraru et al.,2012;sauer andkleine-vehn,2019)。但是,目前关于pils蛋白功能的研究还较少。
5、仅有的研究发现,过表达pils1、3、5、6抑制植株的生长(barbez et al.,2012;beziat et al.,2017;feraru et al.,2019;sun et al.,2020),pils6突变体植株长势良好,高温条件(29℃)下,pils6改变根的伸长(与对照相比过表达和突变体中根的长度变短,但是过表达根的长度最短)(feraru et al.,2019)。gasp1(gloomy and shiny pils)编码一种影响生长素信号输出的ring/u-box超家族蛋白。gasp1突变体中的低生长素信号传导与pils5和pils6蛋白丰度的降低相关。高生长素和低生长素条件分别增加和降低pils6蛋白水平。因此,非最佳生长素浓度通过pils6丰度的改变来缓冲,从而导致稳态生长素输出调节(feraru et al.,2022)。
6、花粉成熟的主要作用似乎是由内质网定位的pins(如pin8和pin5,可能还有pils5)提供的。众所周知,发育和萌发的花粉粒具有高生长素水平,内质网定位的pins可能调节内质网内部储存的生长素的释放,以控制花粉发育并驱动生长素介导的花粉管伸长(cardarelli and cecchetti,2014)。p35s:pin8-ven和p35s:pils5-gfp植物在补充生长素的培养基上表现出根系生长不敏感,生长素反应降低。这两种表型都表明pin8和pils5对核生长素信号传导的负面影响。此外,pils5的根毛特异性表达导致对细胞长度的抑制(dalbosco et al.,2012)。对于pils5和pin8的功能是否相互关联,以及它们在内质网的活动如何影响花粉功能还有待研究。此外,目前还没有关于pils5基因在植物花粉中的功能及耐热性方面的研究。因此,利用现代生物技术手段研究白菜brpils5基因与植物花粉耐热性的潜在联系,有利于为在白菜及其他开花作物繁种中采取有效的保护措施提高种子产量和质量提供理论支撑。
7、参考文献
8、barbez e,kubes m,rolcik j,beziat c,pencik a,wang b,rosquete mr,zhu j,dobrev pi,lee y,zazimalova e,petrasek j,geisler m,friml j,kleine-vehn j(2012)a novel putative auxin carrier family regulates intracellular auxinhomeostasis in plants.nature 485:119-122
9、begcy k,nosenko t,zhou lz,fragner l,weckwerth w,dresselhaus t(2019)male sterility in maize after transient heat stress during the tetrad stageof pollen development.plant physiol 181:683-700
10、beziat c,barbez e,feraru mi,lucyshyn d,kleine-vehn j(2017)lighttriggers pils-dependent reduction in nuclear auxin signalling for growthtransition.nat plants 3:17105
11、cardarelli m,cecchetti v(2014)auxin polar transport in stamenformation and development:how many actors?front plant sci 5:333
12、cecchetti v,altamura mm,falasca g,costantino p,cardarelli m(2008)auxin regulates arabidopsis anther dehiscence,pollen maturation,and filamentelongation.plant cell 20:1760-1774
13、cecchetti v,brunetti p,napoli n,fattorini l,altamura mm,costantino p,cardarelli m(2015)abcb1 and abcb19 auxin transporters have synergisticeffects on early and late arabidopsis anther development.j integr plant biol57:1089-1098
14、cecchetti v,celebrin d,napoli n,ghelli r,brunetti p,costantino p,cardarelli m(2017)an auxin maximumin the middle layer controls stamendevelopment and pollen maturation in arabidopsis.new phytol 213:1194-1207
15、charles jh,godfray jr,beddington ir,crute lh,lawrence.d,f.j,muir jp,robinson.s,sandy m.thomas,toulmin c(2010)food security:the challenge offeeding 9billion people.science327(5967):812–818
16、chaturvedi p,wiese aj,ghatak a,zaveska drabkova l,weckwerth w,honys d(2021)heat stress response mechanisms in pollen development.new phytol 231:571-585
17、cheng y,dai x,zhao y(2006)auxin biosynthesis by the yucca flavinmonooxygenases controls the formation of floral organs and vascular tissuesin arabidopsis.genes dev 20:1790-1799
18、dal bosco c,dovzhenko a,liu x,woerner n,rensch t,eismann m,eimer s,hegermann j,paponov ia,ruperti b,heberle-bors e,touraev a,cohen jd,palme k(2012)the endoplasmic reticulum localized pin8 is a pollen-specific auxincarrier involved in intracellular auxin homeostasis.plant j 71:860-870
19、dal bosco c,dovzhenko a,palme k(2012)intracellular auxin transport inpollen:pin8,pin5 and pils5.plant signal behav 7:1504-1505
20、de storme n,geelen d(2014)the impact of environmental stress on malereproductive development in plants:biological processes and molecularmechanisms.plant cell environ 37:1-18
21、ding y,shi y,yang s(2020)molecular regulation of plant responses toenvironmental temperatures.mol plant 13:544-564
22、ding z,wang b,moreno i,duplakova n,simon s,carraro n,reemmer j,pencika,chen x,tejos r,skupa p,pollmann s,mravec j,petrasek j,zazimalova e,honys d,rolcik j,murphy a,orellana a,geisler m,friml j(2012)er-localized auxintransporter pin8 regulates auxin homeostasis and male gametophyte developmentin arabidopsis.nat commun 3:941
23、feraru e,feraru mi,barbez e,waidmann s,sun l,gaidora a,kleine-vehn j(2019)pils6 is a temperature-sensitive regulator of nuclear auxin input andorgan growth in arabidopsis thaliana.proc natl acad sci u s a 116:3893-3898
24、feraru e,feraru mi,moulinier-anzola j,schwihla m,ferreira da silvasantos j,sun l,waidmann s,korbei b,kleine-vehn j(2022)pils proteins provide ahomeostatic feedback on auxin signaling output.development 149
25、feraru e,vosolsobe s,feraru mi,petrasek j,kleine-vehn j(2012)evolution and structural diversification of pils putative auxin carriers inplants.front plant sci 3:227
26、friml1.j1,anne v,michael s,dolf w,heinz s,thorsten h,remko o,rgens1gj(2003)efflux-dependent auxin gradients establish the apical–basal axis ofarabidopsis.nature 426:146-153
27、friml j(2010)subcellular trafficking of pin auxin efflux carriers inauxin transport.european journal of cell biology 89:231-235
28、galweiler l,guan c,muller a,wisman e,mendgen k,yephremov a,palme k(1998)regulation of polar auxin transport by atpin1 in arabidopsis vasculartissue.science 282:2226-2230
29、ghadirnezhad r,fallah a(2014)temperature effect on yield and yieldcomponents of different rice cultivars in flowering stage.intl j agron 2014:1-4
30、mravec j,skupa p,bailly a,hoyerovák,krecek p,bielach a,petrásek j,zhang j,gaykova v,stierhof yd,dobrev pi,schwarzerovák,rolcík j,seifertovád,luschnig c,benkováe,zazímalováe,geisler m,friml j(2009)subcellularhomeostasis of phytohormone auxin is mediated by the er-localized pin5transporter.nature 459:1136-u1127
31、paponov ia,teale wd,trebar m,blilou k,palme k(2005)the pin auxinefflux facilitators:evolutionary and functional perspectives.trends in plantscience 10:170-177
32、peng sp,jianliang huang,john e.sheehy,rebecca c.laza,romeom.visperas,xuhua zhong,grace s.centeno,gurdev s.khush,cassman akg(2004)riceyields decline with higher night temperaturefrom global warming.proc natlacad sci usa 101:9971–9997
33、sauer m,kleine-vehn j(2019)pin-formed and pin-likes auxin transportfacilitators.development 146
34、sun l,feraru e,feraru mi,waidmann s,wang w,passaia g,wang zy,wabnikk,kleine-vehn j (2020)pin-likes coordinate brassinosteroid signaling withnuclear auxin input in arabidopsis thaliana.curr biol 30:1579-1588 e1576
35、wabnik k,kleine-vehn j,govaerts w,friml j(2011)prototype cell-to-cellauxin transport mechanism by intracellular auxin compartmentalization.trendsin plant science 16:468-475
36、yang j,tian l,sun mx,huang xy,zhu j,guan yf,jia qs,yang zn(2013)auxinresponse factor17 is essential for pollen wall pattern formation inarabidopsis.plant physiol 162:720-731
37、zhao z,zhang y,liu x,zhang x,liu s,yu x,ren y,zheng x,zhou k,jiang l,guo x,gai y,wu c,zhai h,wang h,wan j(2013)arole for a dioxygenase in auxinmetabolism and reproductive development in rice.developmental cell 27:113-122
技术实现思路
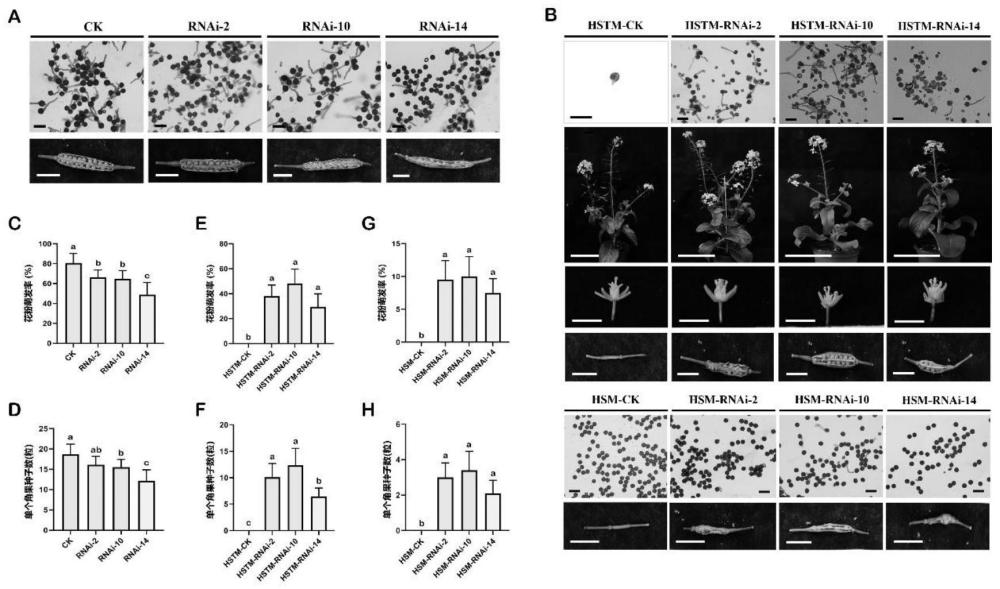
1、本发明提供了白菜brpils5基因在提高植物花粉耐热性中的应用,在受体植物中过表达抑制白菜brpils5基因表达的人工mirna片段,发现抑制白菜brpils5基因的表达能够提高花粉发育早期、发育晚期的耐热性,挽救高温条件下造成的完全雄性不育,为选育植物抗性新品种提供了宝贵资源。
2、本发明具体采用的技术方案如下:
3、白菜brpils5基因在提高植物花粉耐热性中的应用,所述应用为抑制表达白菜brpils5基因提高花粉发育期在高温胁迫下植物花粉的抗性,所述抑制表达白菜brpils5基因是共抑制白菜brpils5a和brpils5b基因,其中,brpils5a基因编号为braa09g010950.3c,基因序列如seq id no.1所示,brpils5b基因编号为braa07g003460.3c,基因序列如seq idno.2所示。
4、进一步地,所述花粉发育期为花粉发育早期或/和花粉发育晚期。
5、进一步地,所述植物花粉为双子叶被子植物花粉或单子叶被子植物花粉。
6、更进一步地,所述植物花粉为十字花科植物花粉。
7、更进一步地,所述植物花粉为白菜花粉。
8、进一步地,应用时,将抑制白菜brpils5基因表达的人工mirna片段连入植物过表达载体中,构建得到重组过表达载体,然后将重组过表达载体转化到受体植物中。
9、更进一步地,所述植物过表达载体为pcambia1300。
10、更进一步地,将重组过表达载体转化到受体植物中,操作如下:将重组过表达载体转化农杆菌,然后用获得的重组农杆菌侵染受体植物。
11、本发明的有益效果:
12、本发明通过分别克隆白菜brpils5基因和抑制其表达的人工mirna片段,并在十字花科植物白菜中进行遗传转化,结果表明过量表达抑制白菜brpils5基因表达的人工mirna片段,能提高花粉发育早期、发育晚期的耐热性,挽救高温条件下造成的完全雄性不育,说明白菜brpils5基因在高温条件下提高植物制种过程中种子的产量和质量具有一定的应用潜力。将该基因应用于白菜或其它开花植物中抗性育种,具有良好的应用前景。
- 还没有人留言评论。精彩留言会获得点赞!