用于制备具有增加的天然甜味和风味的发酵食品的乳酸菌组合物的制作方法
本发明涉及用于生产发酵乳制品的组合物,其包含至少一种嗜热链球菌(streptococcusthermophilus(st))菌株,其中所述st菌株发酵半乳糖,其中所述菌株在编码葡糖激酶蛋白的glck基因的dna序列中携带突变,其中所述突变使葡糖激酶蛋白失活或对所述基因的表达具有负作用,和(ii)至少一种德氏乳杆菌保加利亚亚种(lactobacillusdelbrueckiisubsp.bulgaricus(lb))菌株。这种组合物用于生产具有增加的甜味的发酵乳制品。
背景技术:
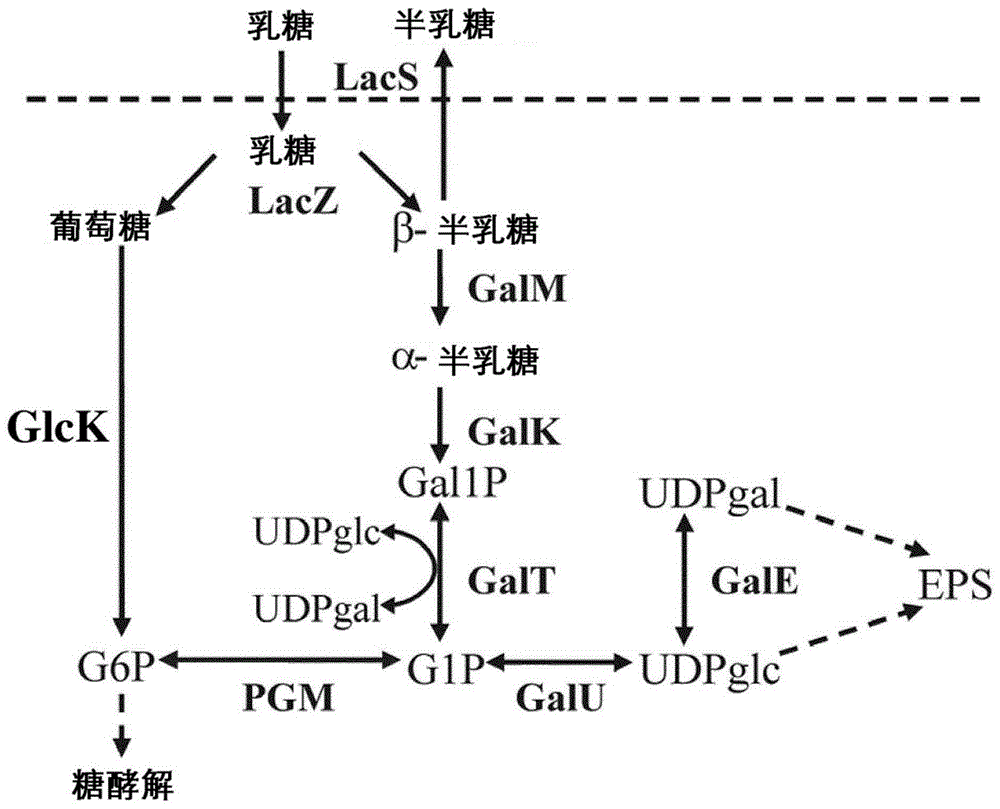
:由于在发酵过程中乳酸菌将乳糖转化为乳酸,纯发酵乳制品通过酸或酸味被识别。因此,它们通常通过添加水果、蜂蜜、糖或人造甜味剂而增甜,以满足消费者对较甜味产品的需求。为了帮助克服在过去20年中变得如此普遍的超重和肥胖问题,食品工业对低热量甜味食品的需求越来越高。甜味通常被认为是令人愉悦的感觉,是由糖和一些其他物质的存在产生的。对糖的感知差异很大。使用蔗糖作为100进行参考,乳糖的甜度为16,半乳糖的甜度为32,葡萄糖的甜度为74(godshall(1988).foodtechnology42(11):71-78)。因此尽管仍然具有大致相同的热量水平,但是葡萄糖感觉的甜是乳糖的4倍以上。发酵食品中的糖经常被可以提供甜度和较低热量摄入的诸如阿斯巴甜、安赛蜜k、三氯蔗糖和糖精等甜味剂替代。然而,人造甜味剂的使用可能导致异味,并且一些研究表明人造甜味剂的消费与诸如饥饿增加、过敏、癌症等缺点有关,这些研究促使消费者偏爱仅含有天然甜味剂或者优选不含添加的甜味剂的发酵乳制品。因此,特别的挑战在于开发天然(内部)甜味高的发酵乳制品。发酵乳制品的酸度在很大程度上取决于存在的乳酸菌和用于制备发酵乳制品的工艺参数。对二糖乳糖的发酵在乳酸菌中进行了大量研究,因为乳糖是乳中的主要碳源。在许多物种中,乳糖在摄取后被β-半乳糖苷酶切割成葡萄糖和半乳糖。葡萄糖被葡糖激酶磷酸化为葡萄糖-6-磷酸,并通过embden-meyerhof-parnas途径(糖酵解)被大多数乳酸菌发酵(图1)。嗜热链球菌(streptococcusthermophilus)是商业嗜热乳发酵最广泛使用的乳酸菌之一,其中该生物通常用作混合起子培养物的一部分,另一组分是乳杆菌(lactobacillussp.),例如用于酸奶的德氏乳杆菌保加利亚亚种(lactobacillusdelbrueckiisubsp.bulgaricus)或用于瑞士型奶酪的瑞士乳杆菌(lactobacillushelveticus)。在许多国家,酸奶的法律定义要求嗜热链球菌和德氏乳杆菌保加利亚亚种一起。这两个物种均产生所需量的乙醛,乙醛是酸奶中的重要风味成分。乳糖和蔗糖比其组分单糖更容易通过嗜热链球菌发酵。在存在过量半乳糖的情况下,当使用嗜热链球菌时,仅乳糖分子的葡萄糖部分发酵,并且半乳糖在发酵乳制品中积累。在高酸浓度限制发酵的酸奶中,游离半乳糖保留,而在瑞士奶酪制造的早期阶段产生的游离半乳糖随后被瑞士乳杆菌发酵。然而,一些研究人员(hutkinsetal.(1986)j.dairysci.69(1):1-8;vail-lancourtetal.(2002)j.bacteriol.184(3);785-793)报道了嗜热链球菌和德氏乳杆菌保加利亚亚种的半乳糖发酵菌株,并且wo2011/026863(chr.hansen)描述了获得发酵半乳糖的嗜热链球菌菌株的方法。为了满足食品工业的要求,提出通过葡萄糖分泌直接向发酵产物提供更多的天然甜味而没有额外热量(内部甜味)的新菌株,特别是嗜热链球菌菌株和德氏乳杆菌保加利亚亚种菌株,具有重要意义。pool等人(2006.metabolicengineering8(5);456-464)公开了乳酸乳球菌(lactococcuslactis)菌株,其中编码葡糖激酶eii(man/glc)和新发现的葡萄糖-ptseii(cel)的基因的缺失完全破坏葡萄糖代谢。构建方法是用于产生所有突变的遗传重组,且因此所得菌株是目前不能用于食品中的基因修饰生物(gmo)。thompson等人(1985.jbacteriol.162(1);217–223)研究了乳酸链球菌(streptococcuslactis)(今天更名为乳酸乳球菌)中的乳糖代谢。在该工作中,使用2-脱氧葡萄糖在甘露糖-pts系统中获得突变体。随后,使用uv诱变诱变该突变体,然后通过复印接种筛选葡萄糖阴性菌落。以这种方式分离出双突变体(甘露糖pts和葡糖激酶)。这种双突变体用于研究与“起子”生物调节乳糖发酵有关的机制。与它们的亲本菌株相比,这些突变体具有几个缺点,这使得它们不适合包含在商业起子培养物中。发酵每摩尔乳糖,突变体的细胞产量是亲本菌株的一半,并且当在乳糖上生长时,突变体的倍增时间显著增加。同样,发酵每摩尔乳糖,乳酸的产率是亲本菌株的一半。没有分析这些菌株在乳中的行为,但预计酸化速率将显著降低。此外,乳酸乳球菌通常不被选择用于乙醛生产,并且其不会有助于满足酸奶法律定义的要求。chervaux等人(2000.appl.andenviron.microbiol.,66,5306-5311)在新的化学成分确定的培养基中研究了德氏乳杆菌保加利亚亚种(lactobacillusdelbrueckiisubsp.bulgaricus)菌株的生理学,并分离了在葡萄糖发酵中具有缺陷的2-脱氧葡萄糖抗性突变体。观察到几种不同的表型,并报道了菌株特异性效应。wo2013/160413公开了具有用于食品天然增甜和降低发酵乳乳糖含量的增强性质的嗜热链球菌菌株和德氏乳杆菌保加利亚亚种菌株,以及筛选和分离这些菌株的方法。wo2015/193449公开了使用乳糖酶和具有葡萄糖代谢缺陷的乳酸菌的组合来生产乳糖浓度极低的发酵乳制品的方法。wo2015/193459公开了生产后酸化水平降低的发酵乳制品的方法,其中一个实施方案使用四种具有葡萄糖代谢缺陷的乳酸菌菌株的混合物,并且其中另一个实施方案使用乳糖缺陷型乳酸菌菌株的混合物。本发明的目的是提供用于生产发酵乳制品的组合物,其包含至少一种葡萄糖缺陷型嗜热链球菌菌株和至少一种德氏乳杆菌保加利亚亚种菌株,其中所述组合物产生具有改善的风味的发酵乳制品。技术实现要素:该目的是通过本发明实现的,本发明涉及用于生产发酵乳制品的组合物,其包含(i)至少一种嗜热链球菌(st)菌株,其中所述st菌株是发酵半乳糖的,其中所述菌株在编码葡糖激酶蛋白的glck基因的dna序列中携带突变,其中所述突变使葡糖激酶蛋白失活或对所述基因的表达具有负作用,和(ii)至少一种德氏乳杆菌保加利亚亚种(lb)菌株,其中所述lb菌株是乳糖缺型的,并且能够代谢非乳糖碳水化合物。众所周知,对于用于生产发酵乳制品、包含嗜热链球菌菌株和德氏乳杆菌保加利亚亚种菌株的传统起子培养物而言,存在形成不希望的异味的风险,例如游离氨基酸,例如谷氨酸。还公知的是,异味是由德氏乳杆菌保加利亚亚种菌株形成的,且形成的异味水平取决于根据许多因素而变化的所述菌株的生长水平,所述因素例如德氏乳杆菌保加利亚亚种菌株的接种水平,生长培养基的类型和发酵的生长条件,以及起子培养物中所用的具体嗜热链球菌菌株和德氏乳杆菌保加利亚亚种菌株。现已发现,对于用于生产具有增加的甜味的发酵乳制品、包含至少一种葡萄糖缺型嗜热链球菌菌株和至少一种德氏乳杆菌保加利亚亚种菌株的起子培养物而言,不希望的异味的形成处于特别高的水平。这与在这种起子培养物中德氏乳杆菌保加利亚亚种菌株生长到特别高的细胞数这一事实一致。令人惊讶地发现,使用乳糖缺陷型德氏乳杆菌保加利亚亚种菌株,例如乳糖缺陷型、葡萄糖阳性德氏乳杆菌保加利亚亚种菌株,可以解决使用具有葡萄糖缺陷型嗜热链球菌菌株的起子培养物的异味问题。还发现,本发明组合物所用的乳糖缺陷型德氏乳杆菌保加利亚亚种菌株不会导致任何后酸化。最后,发现与传统相比,即与乳糖阳性、葡萄糖阳性德氏乳杆菌保加利亚亚种菌株相比,乳糖缺陷型德氏乳杆菌保加利亚亚种与葡萄糖缺陷型嗜热链球菌菌株组合产生非常高的细胞计数。鉴于德氏乳杆菌保加利亚亚种菌株是葡萄糖阳性的并且存在于含有葡萄糖的发酵乳制品中这一事实,从例如乳糖缺陷型葡萄糖阳性德氏乳杆菌保加利亚亚种菌株有可能避免形成异味,并避免后酸化是非常令人惊讶的,其中理论上该菌株在发酵乳制品的储存期间能够继续生长。此外,更令人惊讶的是,有可能从德氏乳杆菌保加利亚亚种菌株避免形成异味,并避免后酸化,同时获得德氏乳杆菌保加利亚亚种菌株的高细胞计数,这是因为通常预期异味和后酸化都将由德氏乳杆菌保加利亚亚种菌株的生长引起。在不受理论束缚的情况下,相信本发明的令人惊讶的效果归因于本发明的乳糖缺陷型德氏乳杆菌保加利亚亚种与传统乳糖阳性德氏乳杆菌保加利亚亚种的代谢差异。附图说明图1是嗜热链球菌中乳糖分解代谢的示意图。glck,葡糖激酶;lacs,乳糖转运蛋白;lacz,β-半乳糖苷酶;galm,变旋酶;galk,半乳糖激酶;galt,半乳糖-1-磷酸尿苷酰转移酶;gale,udp-葡萄糖4差向异构酶;gal1p,半乳糖-1-磷酸。具体实施方式定义如本文所用,术语“乳酸菌”表示革兰氏阳性、微需氧或厌氧细菌,其发酵糖同时产生酸,所述酸包括乳酸作为主要产生的酸、乙酸和丙酸。工业上最有用的乳酸菌见于“乳杆菌目(lactobacillales)”,其包括乳球菌属种(lactococcusspp.)、链球菌属种(streptococcusspp.)、乳杆菌属种(lactobacillusspp)、明串珠菌属种(leuconostocspp.)、片球菌属种(pediococcusspp.)、短杆菌属种(brevibacteriumspp.)、肠球菌属种(enterococcusspp.)和丙酸杆菌属种(propionibacteriumspp.)。通常将乳酸菌,包括乳杆菌属种和嗜热链球菌物种,作为冷冻或冻干培养物供应给乳制品工业用于批量起子繁殖或作为所谓的“直投式”(directvatset,dvs)培养物,用来直接接种到发酵容器或发酵槽中以产生乳制品,例如发酵乳制品。这种培养物通常被称为“起子培养物”或“起子”。在描述本发明的上下文中(特别是在以下权利要求的上下文中)使用术语“a(一)”和“an(一)”和“所述(the)”以及类似的指示符应解释为涵盖单数和复数,除非本文另有说明或与上下文明显矛盾。除非另有说明,否则术语“包含(comprising)”、“具有(having)”、“包括(including)”和“含有(containing)”应解释为开放式术语(即,意味着“包括但不限于”)。除非本文另有说明,否则本文中对数值范围的列举仅旨在用作单独提及落入该范围内的每个单独值的简写方法,并且每个单独值并入本说明书中,如同其在本文中单独列举一样。除非本文另有说明或上下文明显矛盾,否则本文所述的所有方法均可以任何合适的顺序进行。除非另外声明,否则本文提供的任何和所有实例或示例性语言(例如,“诸如”)的使用仅旨在更好地说明本发明,而不是对本发明的范围进行限制。不应将说明书中的语言解释为表明任何未要求保护的要素对于本发明的实践是必不可少的。对于本发明,表述“增加甜味”是指与由未在编码葡糖激酶蛋白的glck基因的dna序列中携带突变的母菌株产生的甜味相比,甜味增加,其中所述突变使葡糖激酶蛋白失活或对所述基因表达具有负作用。表述“cfu”表示菌落形成单位(colonyformingunit)。本发明组合物的嗜热链球菌菌株在一些国家,酸奶的法律定义要求存在嗜热链球菌和德氏乳杆菌保加利亚亚种二者。这两个物种均产生期望量的乙醛,酸奶中的重要风味成分。通过使用嗜热链球菌和德氏乳杆菌保加利亚亚种二者进行发酵也可以制备诸如马苏里拉奶酪(mozzarella)和比萨奶酪以及羊奶酪(feta)等奶酪(etal.(2010),于thetechnologyofcheesemaking,2nded.blackwellpublishing,oxford;166-192)。为了满足食品工业的要求,期望开发直接在发酵产品中产生更多天然甜味(内部甜味)而不会贡献额外热量的新菌株,特别是德氏乳杆菌保加利亚亚种菌株和嗜热链球菌菌株。嗜热链球菌是商业乳发酵最广泛使用的乳酸菌之一,其中该生物通常用作混合起子培养物的一部分,另一组分是乳杆菌属种,例如用于酸奶的德氏乳杆菌保加利亚亚种,和用于瑞士型奶酪的瑞士乳杆菌。当使用嗜热链球菌时,乳糖分子的仅葡萄糖部分被嗜热链球菌发酵,并且半乳糖在发酵乳制品中积累。在高酸浓度限制发酵的酸奶中,游离半乳糖保留,而在瑞士奶酪制造的早期阶段产生的游离半乳糖随后被瑞士乳杆菌发酵。在许多用于制作奶酪的起子培养物中发现的乳酸乳球菌也能够消耗由嗜热链球菌产生的半乳糖。为了确保具有尽可能最佳生长性能的嗜热链球菌菌株,本发明人将嗜热链球菌的半乳糖发酵菌株暴露于选择剂2-脱氧葡萄糖。通常,2-脱氧葡萄糖抗性突变体在编码葡糖激酶的基因和为葡萄糖转运编码的基因中具有突变。对2-脱氧葡萄糖具有抗性的分离的突变体chcc16731在其葡糖激酶(glck)基因中具有突变。除了葡糖激酶基因中的突变外,chcc16731和chcc19216都具有意味着分泌的葡萄糖不会再次转运回细胞的突变。令人惊讶的是,单独的这种突变体仍然完全能够酸化乳,尽管酸化到ph5的时间被延迟2-5小时。因此它们在发酵乳应用中是有用的,并且它们保留了母菌株酸化乳的能力,这是酸奶的特征。此外,发现当用所述突变体接种含有0.05%蔗糖的9.5%b-乳并在40℃发酵至少20小时时,所述突变体分泌高水平的葡萄糖。同时,发酵乳中残留的乳糖含量非常低。因此,使用这些菌株生产发酵乳制品对于乳糖不耐受的人可具有重要性。因此如godshall(1988.foodtechnology42(11):71-78)所述计算,最终发酵乳具有增加的内部甜味指数。在本发明的一个方面,嗜热链球菌菌株是嗜热链球菌的半乳糖发酵突变菌株,其中突变菌株在编码葡糖激酶蛋白的glck基因的dna序列中携带突变,其中所述突变使编码的葡糖激酶蛋白失活,或对基因表达具有负作用。用于测量葡糖激酶活性水平或葡糖激酶基因表达水平的方法是容易获知的(porteretal.(1982)biochim.biophys.acta,709;178–186),并且包括具有市售试剂盒的酶测定法和使用容易获得的材料的转录组学或定量pcr。在本发明的优选实施方案中,本发明的嗜热链球菌菌株是2-脱氧葡萄糖抗性的。如本文所用的细菌“菌株”是指在生长或繁殖时保持遗传上不变的细菌。包括大量相同的细菌。本文所用的术语“半乳糖发酵嗜热链球菌菌株”是指能够在m17培养基+2%半乳糖上/中生长的嗜热链球菌菌株。半乳糖发酵的嗜热链球菌菌株在本文中定义为,当由过夜培养物以1%接种并在37℃下温育24小时时,将含有2%半乳糖作为唯一碳水化合物的m17培养液的ph降低至5.5或更低的嗜热链球菌菌株。本文所用术语“突变使葡糖激酶蛋白失活”是指产生“失活的葡糖激酶蛋白”的突变,如果存在于细胞中,则不能发挥其正常功能的葡糖激酶蛋白,以及防止葡糖激酶蛋白形成或导致葡糖激酶蛋白降解的突变。特别地,失活的葡糖激酶蛋白是指,与功能性葡糖激酶蛋白质相比不能促进葡萄糖磷酸化为葡萄糖-6-磷酸,或以显著降低的速率促进葡萄糖磷酸化为葡萄糖-6-磷酸的蛋白。与编码功能性葡糖激酶蛋白的基因相比,编码这种失活的葡糖激酶蛋白的基因在所述基因的开放阅读框(orf)中包含突变,其中所述突变可包括但不限于缺失、移码突变,引入终止密码子或导致(改变蛋白质功能特性的)氨基酸取代的突变,或者降低或消除基因转录或翻译的启动子突变。在优选的实施方案中,突变使葡糖激酶蛋白的活性(葡萄糖磷酸化为葡萄糖-6-磷酸的速率)降低至少50%,例如至少60%,例如至少70%,例如至少80%,例如至少90%。葡糖激酶活性可以通过pool等人(2006.metabolicengineering8;456-464)描述的葡糖激酶酶促测定来确定。本文所用的术语“功能性葡糖激酶蛋白”是指,如果存在于细胞中,则促进葡萄糖磷酸化为葡萄糖-6-磷酸的葡糖激酶蛋白。特别是,功能性葡糖激酶蛋白可以由包含orf的基因编码,所述orf具有对应于seqidno.1中1-966位的序列,或与对应于seqidno.1的1-966位的序列具有至少85%同一性,例如至少90%同一性,例如至少95%同一性,例如至少98%同一性,例如至少99%同一性的序列。两条序列的百分比同一性可以通过使用数学算法测定,例如karlin和altschul的算法(1990.proc.natl.acad.sci.usa87;2264),karlin和altschul描述的改进算法(1993.proc.natl.acad.sci.usa90;5873-5877);myers和miller的算法(1988.cabios4;11-17);needleman和wunsch的算法(1970.j.mol.biol.48;443-453);以及pearson和lipman的算法(1988.proc.natl.acad.sci.usa85;2444-2448)。基于这些数学算法的用于测定核酸或氨基酸序列同一性的计算机软件也是可用的。例如,可以用blastn程序,得分=100,字长=12,进行核苷酸序列的比较。可以用blastx程序,得分=50,字长=3,进行氨基酸序列的比较。对于blast程序的其余参数,可以使用默认参数。许多国家不接受将基因修饰生物(gmo)用于发酵乳制品中。作为替代,本发明提供能够在发酵乳制品中提供所需葡萄糖累积的天然存在的或诱导的突变菌株。因此,在本发明的非常优选的实施方案中,突变菌株是天然存在的突变体或诱导突变体。本文所用的“突变细菌”或“突变菌株”是指在基因组(dna)中包含野生型dna中不存在的一个或多个突变的天然(自发的、天然存在的)突变细菌或诱导的突变细菌。“诱导的突变体”是这样的细菌,在所述细菌中,突变通过人工处理诱导,例如用化学诱变剂、uv-或γ辐射等处理。相比之下,“自发突变体”或“天然存在的突变体”并没有被人诱变。本文中的突变细菌是非gmo(非基因修饰生物),即未经重组dna技术修饰。“野生型菌株”是指如在自然界发现的细菌的非突变形式。诸如“具有增甜特性的菌株”、“能够在发酵乳制品中提供所需的葡萄糖累积的菌株”和“具有增强的食品天然增甜特性的菌株”等术语,在本文中互换使用,来表征在乳制品发酵中使用本发明菌株的有利方面。在优选的实施方案中,当以106-107cfu/ml的浓度接种到9.5%b-乳中,并在40℃下生长至少20小时时,本发明的嗜热链球菌的突变菌株将9.5%b-乳中葡萄糖的量增加至至少5mg/ml。在另一个优选的实施方案中,当以106-107cfu/ml的浓度接种于含有0.05%蔗糖的9.5%b-乳中,并在40℃生长至少20小时时,本发明的嗜热链球菌的突变菌株将含有0.05%蔗糖的9.5%b-乳中葡萄糖的量增加至至少5mg/ml。在本发明的上下文中,9.5%b乳是用重构低脂脱脂乳粉制备至干物质含量为9.5%的水平、并在99℃下巴氏灭菌30min(分钟)、随后冷却至40℃的煮沸的乳。在本发明更优选的实施方案中,突变菌株导致葡萄糖的量增加至至少6mg/ml,例如至少7mg/ml,例如至少8mg/ml,例如至少9mg/ml,例如至少10mg/ml,例如至少11mg/ml,例如至少12mg/ml,例如至少13mg/ml,例如至少14mg/ml,例如至少15mg/ml,例如至少20mg/ml,例如至少25mg/ml。在本发明的另一个实施方案中,嗜热链球菌的突变菌株对2-脱氧葡萄糖具有抗性。在本文中,术语“对2-脱氧葡萄糖具有抗性”的定义是,当在含有20mm2-脱氧葡萄糖的m17培养基平板上划线培养时,在40℃下温育20个小时后,特定突变细菌菌株具有生长成菌落的能力。培养基中2-脱氧葡萄糖的存在会阻止非突变菌株的生长,而突变菌株的生长没有受到影响或没有受到显著影响。在评估抗性时可用作敏感参照菌株的非突变菌株优选包括菌株chcc14994和chcc11976。当用106-107cfu/ml的本发明的嗜热链球菌菌株接种9.5%b-乳,并在40℃下用本发明的嗜热链球菌菌株发酵至少20小时时,本发明的突变嗜热链球菌菌株将葡萄糖分泌到乳中。优选地,当用106-107cfu/ml的本发明的嗜热链球菌菌株接种9.5%b-乳,并在40℃下用所述嗜热链球菌菌株发酵至少20小时时,这种突变菌株单独将分泌至少5mg/ml葡萄糖到b-乳中。虽然酸化至ph5的时间延迟2-5小时,但所述菌株仍能完全酸化乳。最终的发酵乳在发酵乳中含有少于15mg/ml的乳糖。因此,最终的发酵乳具有约2倍或更多的更高内部甜味指数。在又一个实施方案中,本发明的突变菌株可以以其生长模式来表征。这可以由以下发现来说明,即突变菌株在m17培养基+2%半乳糖中的生长速率比在m17培养基+2%葡萄糖中的高。测量生长速率作为以指数生长的培养物在600纳米(od600)下的光密度随时间的发展。在优选的实施方案中,生长速率在m17培养基+2%半乳糖中比在m17培养基+2%葡萄糖中高至少5%,例如高至少10%,例如高至少15%,例如高至少20%。glck基因中的突变在一个优选的实施方案中,所述突变导致编码seqidno.2第249位编码甘氨酸的密码子被编码精氨酸的密码子置换。优选地,glck基因中的突变导致seqidno.1第745位的g被a置换。菌株chcc16731具有这种突变。在优选的实施方案中,所述突变导致编码seqidno.2第72位编码丝氨酸的密码子被编码脯氨酸的密码子置换(未显示)。优选,glck基因中的突变导致seqidno.1第214位的t被c置换(未显示)。在另一个优选的实施方案中,突变导致编码seqidno.2第141位编码苏氨酸的密码子被编码异亮氨酸的密码子取代(未显示)。优选地,glck基因中的突变导致seqidno.1第422位的c被t置换(未显示)。应当强调的是,嗜热链球菌的glck基因可以被glck基因其他位点的其他类型突变失活。降低葡萄糖向细胞内转运的突变在优选的实施方案中,嗜热链球菌菌株携带降低葡萄糖向细胞内转运的突变。本文所用的术语“降低葡萄糖向细胞内转运的突变”是指编码涉及葡萄糖转运的蛋白质的基因中的突变,其导致葡萄糖在细胞环境中累积。当培养基为乳基质时,可以通过本领域技术人员已知的方法容易地测量嗜热链球菌菌株培养基中葡萄糖的水平。在优选的实施方案中,突变使葡萄糖向细胞的转运降低至少50%,例如至少60%,例如至少70%,例如至少80%,例如至少90%。葡萄糖向细胞内的转运可以通过cochu等人(2003.applenvironmi-crobiol69(9);5423-5432)所述的葡萄糖摄取测定来测定。优选,嗜热链球菌菌株在编码葡萄糖转运蛋白组分的基因中携带突变,其中所述突变使葡萄糖转运蛋白失活或对所述基因的表达具有负作用。所述组分可以是葡萄糖转运蛋白中对葡萄糖转运关键的任何组分。例如,预期嗜热链球菌中葡萄糖/甘露糖pts的任何组分的失活都将导致葡萄糖转运蛋白功能失活。本文所用的术语“突变使葡萄糖转运蛋白失活”是指导致“失活的葡萄糖转运蛋白”的突变,“失活的葡萄糖转运蛋白”是如果存在于细胞中,则其不能发挥其正常功能的葡萄糖转运蛋白蛋白质,以及防止葡萄糖转运蛋白蛋白质形成或导致葡萄糖转运蛋白蛋白质降解的突变。特别是,失活的葡萄糖转运蛋白是,与功能性葡萄糖转运蛋白相比不能促进葡萄糖穿过质膜运输或以显著降低的速率促进葡萄糖穿过质膜运输的蛋白。与编码功能性葡萄糖转运蛋白的基因相比,编码这种失活的葡萄糖转运蛋白的基因在所述基因的开放阅读框(orf)中包含突变,其中所述突变可以包括但不限于缺失、移码突变,引入终止密码子或导致改变所述蛋白质功能特性的氨基酸取代的突变,或降低或消除基因转录或翻译的启动子突变。在优选的实施方案中,所述突变使葡萄糖转运蛋白蛋白质的活性(转运葡萄糖的速率)降低至少50%,例如至少60%,例如至少70%,例如至少80%,例如至少90%。葡萄糖转运蛋白活性可以通过cochu等人(2003.applenvironmicro-biol69(9);5423-5432)所述的葡萄糖摄取测定来测定。本文所用的术语“功能性葡萄糖转运蛋白蛋白质”是指,如果存在于细胞中,则促进葡萄糖穿过质膜转运的葡萄糖转运蛋白蛋白质。在本发明的优选实施方案中,本发明的嗜热链球菌菌株在编码葡萄糖/甘露糖磷酸转移酶系统的iidman蛋白质的mann基因的dna序列中携带突变,其中所述突变使所述iidman蛋白失活或对所述基因的表达具有负作用。chcc16731在编码葡萄糖/甘露糖磷酸转移酶系统的iidman蛋白质的mann基因的第79位具有苏氨酸到脯氨酸的变化。优选,mann基因中的突变导致seqidno.3第235位的a被c置换。因此,在优选实施方案中,本发明的嗜热链球菌菌株在编码葡萄糖/甘露糖磷酸转移酶系统的iidman蛋白质的mann基因的dna序列中携带突变,其中所述突变导致seqidno.4第79位苏氨酸置换为脯氨酸。在本发明的另一个优选实施方案中,本发明的嗜热链球菌菌株在编码葡萄糖/甘露糖磷酸转移酶系统的iicman蛋白质的manm基因的dna序列中携带突变,其中所述突变使iicman蛋白质失活或对所述基因的表达具有负作用。在具体优选的实施方案中,所述突变导致编码葡萄糖/甘露糖磷酸转移酶系统的iicman蛋白质的seqidno.6第209位编码谷氨酸的密码子被终止密码子置换(未显示)。优选,突变导致seqidno.5第625位的g被t置换(未显示)。本发明优选的嗜热链球菌菌株在本发明组合物的优选实施方案中,嗜热链球菌菌株选自由以下组成的组:以保藏号dsm32227保藏于德国微生物保藏中心(deutschesammlungvonmikroorganismenundzellkulturen)的嗜热链球菌chcc19216菌株,2014年6月以保藏号dsm28889保藏于德国布伦瑞克因霍芬街7bd-38124(inhoffenstr.7b,d-38124braunschweig)的德国微生物保藏中心(dsmz)的菌株嗜热链球菌chcc16731,以保藏号dsm25850保藏于德国微生物保藏中心的嗜热链球菌chcc15757菌株,以保藏号dsm25851保藏于德国微生物保藏中心的嗜热链球菌chcc15887菌株,以保藏号dsm26722保藏于德国微生物保藏中心的嗜热链球菌chcc16404菌株,以及由其衍生的突变菌株,其中所述突变菌株通过使用保藏菌株之一作为起始材料而获得,并且其中所述突变体保留或进一步改善了所述保藏菌株的质构特性和/或葡萄糖分泌特性。在本发明的上下文中,术语“突变菌株”应理解为通过例如基因工程、辐射和/或化学处理由本发明的菌株(或其母株)衍生的、或可衍生的菌株。“由其衍生的菌株”也可以是自发发生的突变体。“由其衍生的菌株”优选是功能上等同的突变体,例如与其母株具有基本相同或改善的特性的突变体。特别是,术语“突变菌株”是指通过使本发明的菌株经历任何常规使用的诱变处理而获得的菌株,包括用化学诱变剂例如乙烷甲烷磺酸盐(ems)或n-甲基-n'-硝基-n-硝基胍(ntg)、紫外线处理,或是指自发发生的突变体。突变体可能已经受几次诱变处理(单次处理应认为是一诱变步骤,接着是筛选/选择步骤),但目前优选的是,进行不超过20次,或不超过10次,或不超过5次处理(或筛选/选择步骤)。在目前优选的突变体中,与母株相比,细菌基因组中小于1%、小于0.1%、少于0.01%、少于0.001%或甚至少于0.0001%的核苷酸已被另一核苷酸置换或缺失。获得增甜嗜热链球菌菌株的方法在本发明的优选实施方案中,用于研发增甜嗜热链球菌菌株的起始菌株是质构化嗜热链球菌菌株,其可根据本说明书其他部分所述获得。在优选的实施方案中,这种质构化起始菌株选自由以下组成的组:以保藏号dsm22932保藏于德国微生物保藏中心的嗜热链球菌chcc11342菌株,以保藏号dsm22934保藏于德国微生物保藏中心的嗜热链球菌chcc11976菌株,以保藏号dsm24090保藏于德国微生物保藏中心的嗜热链球菌chcc12339菌株,以及由其衍生的菌株。或者,用于研发增甜嗜热链球菌菌株的起始菌株是非质构化嗜热链球菌菌株,在这种情况下,质构化特性则会被引入一旦产生的增甜嗜热链球菌菌株中,即在本说明书其他地方所述的获得质构化菌株的方法中使用增甜菌株作为起始菌株。使嗜热链球菌菌株增甜的方法的第一步是提供半乳糖阳性菌株,即能够使用半乳糖作为碳水化合物源的菌株。半乳糖阳性菌株可以通过包括以下步骤的方法获得:在琼脂平板上,例如含有一定浓度例如2%半乳糖(唯一碳水化合物源)的m17琼脂平板上将待测细菌划线培养,并鉴定能够在所述平板上生长的菌落。2-脱氧葡萄糖以及与在m17培养基+2%葡萄糖中相比,在m17培养基+2%半乳糖中细菌生长模式的测定用于选择在葡糖激酶(glck)基因中具有突变的细菌。一种筛选和分离具有突变的glck基因的嗜热链球菌菌株的方法,包括以下步骤:a)提供半乳糖发酵嗜热链球菌母株;b)从衍生自母株的突变嗜热链球菌菌株池中选择并分离对2-脱氧葡萄糖具有抗性的突变嗜热链球菌菌株池;以及c)如果突变嗜热链球菌菌株在m17培养基+2%半乳糖中的生长速率比在m17培养基+2%葡萄糖中的高,则从对2-脱氧葡萄糖具有抗性的突变嗜热链球菌菌株池中选择并分离所述突变嗜热链球菌菌株。本文中术语“对2-脱氧葡萄糖具有抗性”的定义是,当在含有2%乳糖或2%半乳糖并含有20mm2-脱氧葡萄糖的m17培养基平板上划线培养时,在40℃下温育20小时后,特定突变细菌菌株具有生长成菌落的能力。培养基中2-脱氧葡萄糖的存在会防止非突变菌株的生长,而突变菌株的生长不受影响或未受到显著影响。在抗性评估时可用作敏感参照菌株的非突变菌株包括菌株chcc11976。在实施方案中,所述方法还包括步骤a1):使母株经受诱变,例如使母株经受化学和/或物理诱变剂。在另一个实施方案中,所述方法还包括步骤d):如果嗜热链球菌菌株在m17培养基+2%蔗糖中的生长速率高,但与母株在m17培养基+2%葡萄糖菌中的生长速率相比为零或降低至少0-50%,则从衍生于步骤c)中选择的嗜热链球菌菌株的2-脱氧葡萄糖抗性嗜热链球菌菌株池中选择并分离所述嗜热链球菌菌株。半乳糖发酵嗜热链球菌母株能够在m17培养基+2%半乳糖上/中生长,并且在本文中定义为,当由1%的过夜培养物接种并在37℃温育24小时时,它们具有将含有2%半乳糖作为唯一碳水化合物的m17肉汤中的ph降低至5.5或更低的能力。wo2011/026863(chr.hansena/s(科.汉森有限公司))和wo2011/092300(chr.hansena/s(科.汉森有限公司))中描述了这样的半乳糖阳性菌株。在本上下文中,术语“由其衍生的菌株”应理解为通过例如基因工程、辐射和/或化学处理衍生自或可衍生自质构化或半乳糖发酵嗜热链球菌母株的菌株。“由其衍生的菌株”也可以是自发发生的突变体。优选的是“由其衍生的菌株”是功能上等同的突变体,例如与其母株具有基本相同或改善的特性(例如有关质构或半乳糖发酵)的突变体。特别是,术语“由其衍生的菌株”是指通过使本发明的菌株经受任何常规使用的诱变处理,包括用化学诱变剂例如乙烷甲烷磺酸盐(ems)或n-甲基-n'-硝基-n-硝基胍(ntg)、紫外线处理获得的菌株,或是指自发发生的突变体。突变体可能已经经受几次诱变处理(应当将单次处理理解为一个诱变步骤,接着是筛选/选择步骤),但目前优选进行不超过20次或不超过10次或不超过5次处理(或筛选/选择步骤)。在目前优选的突变体中,与母株相比,细菌基因组中少于1%,少于0.1%,少于0.01%,少于0.001%或甚至少于0.0001%的核苷酸已被另一核苷酸置换或缺失。在本发明中,表述“编码葡糖激酶的glck基因”是指嗜热链球菌的编码具有葡糖激酶活性的蛋白质的任何dna序列,包括seqidno.:1的dna序列编码的具体葡糖激酶。葡糖激酶催化将葡萄糖转化成葡萄糖-6-磷酸的反应,参见图1。本发明组合物的德氏乳杆菌保加利亚亚种菌株术语“乳糖代谢缺陷”和“乳糖缺陷”在本发明的上下文中用于表征部分或完全丧失使用乳糖作为细胞生长或维持细胞活力来源的能力的lab。各lab能够代谢一种或几种选自蔗糖、半乳糖和/或葡萄糖的碳水化合物或另一种可发酵碳水化合物。由于这些碳水化合物不是以足以支持乳糖缺陷型突变体发酵的量天然存在于乳中,因此必须将这些碳水化合物添加到乳中。乳糖缺陷型和部分缺陷型lab可以表征为在含有乳糖和x-gal的培养基上的白色菌落。对于本发明,术语“x-gal”是指5-溴-4-氯-3-吲哚基-β-d-半乳糖吡喃糖苷,它是β-半乳糖苷酶的显色底物,β-半乳糖苷酶将x-gal水解成无色半乳糖和5-溴-4-氯-吲哚酚,它们自发地二聚化形成蓝色颜料。在本发明的具体实施方案中,乳糖缺陷型菌株能够代谢选自由蔗糖、半乳糖和葡萄糖,优选蔗糖组成的组的非乳糖碳水化合物。在本发明的具体实施方案中,乳糖缺陷型菌株能够代谢半乳糖。在本发明组合物的具体实施方案中,lb菌株选自由以下组成的组:于2014年6月12日以保藏号dsm28910保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心的菌株和由其衍生的突变菌株,其中所述突变菌株的进一步特征为具有在含有乳糖和x-gal的培养基上生成白色菌落的能力。乳糖缺陷型菌株可以通过突变从合适的乳糖阳性母株中获得。在uv-诱变后,乳糖缺陷型菌株在合适的培养基上,例如具有1%乳糖和200mg/mlx-gal的mrs琼脂平板上被选择为白色菌落(表明乳糖缺陷表型)。乳糖阳性菌株具有β-半乳糖苷酶活性,并且由于β-半乳糖苷酶的活性,乳糖阳性菌株的菌落呈蓝色。作为用于产生乳糖缺陷型菌株的乳糖阳性母株,可以使用针对wo2011/000879在2007-04-03日以保藏号dsm19252保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心的德氏乳杆菌保加利亚亚种菌株chcc10019。组合物本发明进一步涉及包含104-1012cfu(菌落形成单位)/g嗜热链球菌菌株,例如105-1011cfu/g,例如106-1010cfu/g,或如107-109cfu/g的嗜热链球菌菌株的组合物。在优选的实施方案中,所述组合物的嗜热链球菌菌株不能酸化9.5%的b-乳,定义为当用106-107cfu/ml的嗜热链球菌菌株接种9.5%的b-乳时,并在40℃温育14小时时,导致小于1.0的ph降低,并且所述组合物还包含一定量的可以通过触发嗜热链球菌菌株chcc16404酸化9.5%b-乳的化合物,chcc16404以保藏号dsm26722保藏于德国微生物保藏中心,定义为当用106-107cfu/ml的所述嗜热链球菌菌株接种9.5%b-乳,并在40℃温育14小时时,导致1.0或更高的ph值降低。优选,所述化合物是蔗糖。优选,蔗糖的量为0.000001%-2%,例如0.00001%-0.2%,例如0.0001%-0.1%,例如0.001%-0.05%。在特别优选的实施方案中,所述组合物还包含104-1012cfu/g的德氏乳杆菌保加利亚亚种菌株,例如105-1011cfu/g,例如106-1010cfu/g,或例如107-109cfu/g的德氏乳杆菌保加利亚亚种菌株。在本发明的具体实施方案中,所述组合物含有至少两种st菌株,优选至少三种st菌株,更优选三种st菌株。在本发明的具体实施方案中,所述组合物含有不超过三种lb菌株,优选不超过两种lb菌株,更优选一种lb菌株。在本发明的具体实施方案中,所述组合物含有至少一种选自由以下组成的组的st菌株:于2014年6月4日以保藏号dsm28889保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz)的菌株嗜热链球菌chcc16731,以保藏号dsm25850保藏于德国微生物保藏中心的嗜热链球菌chcc15757菌株,以保藏号dsm26722保藏于德国微生物保藏中心的嗜热链球菌chcc16404菌株,和于2014年6月12日以保藏号dsm28910保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心的lb菌株。德氏乳杆菌保加利亚亚种、嗜热链球菌和其他乳酸菌通常用作起始培养物,在生产各种食品中用作技术目的,例如在乳品工业中,例如用于发酵乳制品。因此,在另一个优选的实施方案中,所述组合物适合作为起子培养物。除了液体起子培养物之外,起子培养物可以作为冷冻或干燥的起子培养物提供。因此,在又一个优选的实施方案中,所述组合物采用冷冻形式、冷冻干燥形式或液体形式。如wo2005/003327所公开的,将某些低温保护剂加入到起子培养物中是有益的。因此,本发明的起子培养物组合物可以包含一种或多种选自以下的低温保护剂:肌苷-5'-单磷酸(imp)、腺苷-5'-单磷酸(amp)、鸟苷-5'-单磷酸(gmp)、尿苷-5'-单磷酸(ump)、胞苷-5'-单磷酸(cmp)、腺嘌呤、鸟嘌呤、尿嘧啶、胞嘧啶、腺苷、鸟苷、尿苷、胞苷、次黄嘌呤、黄嘌呤、次黄嘌呤、乳清苷、胸苷、肌苷和任何此类化合物的衍生物。生产发酵乳制品的方法本发明还涉及生产发酵乳制品的方法,包括用本发明的组合物接种和发酵乳基质。术语“乳(milk)”应理解为通过为任何哺乳动物如奶牛、绵羊、山羊、水牛或骆驼挤奶而获得的乳汁分泌物。在优选的实施方案中,乳是牛奶。术语“乳基质”可以是可根据本发明的方法进行发酵的任何未加工的乳材料和/或加工的乳材料。因此,有用的乳基质包括但不限于包含蛋白质的任何乳或乳样产品的溶液或悬浮液,例如全脂或低脂乳、脱脂乳、酪乳、复原乳粉、炼乳、奶粉(driedmilk)、乳清、乳清渗透物、乳糖、来自乳糖结晶的母液、乳清蛋白质浓缩物或奶油。显然,乳基质可能来源于任何哺乳动物,例如,基本上纯的哺乳动物乳或复原乳粉。优选地,乳基质中的至少部分蛋白质是乳中天然存在的蛋白质,例如酪蛋白或乳清蛋白。然而,部分蛋白质可以是非乳中天然存在的蛋白质。在发酵之前,可以根据本领域已知的方法将乳基质均化并巴氏灭菌。如本文所用,“均化”意指强烈混合以获得可溶性悬浮液或乳液。如果在发酵前进行均化,那么均化可进行以便将乳脂破碎成更小的尺寸,从而乳脂不再与乳分离。这可以通过使乳在高压下通过小孔来完成。如本文所用,“巴氏灭菌”意指处理乳基质以减少或消除活生物例如微生物的存在。优选地,通过将指定温度保持特定时间实现巴氏灭菌。指定温度通常通过加热获得。可以选择温度和持续时间,以杀死或灭活某些细菌,例如有害细菌。随后可以进行快速冷却步骤。本发明的方法中,“发酵”是指通过微生物的作用将碳水化合物转化成醇或酸。优选,本发明方法中的发酵包括将乳糖转化成乳酸。用于生产发酵乳制品的发酵工艺是众所周知的,并且本领域技术人员将知道如何选择合适的工艺条件,例如温度、氧气、微生物的量和特性以及工艺时间。显然,选择发酵条件是为了支持本发明的实现,即获得固体或液体形式的乳制品(发酵乳制品)。本文所用的术语“发酵乳制品”是指食品或饲料产品,其中食品或饲料产品的制备包括用乳酸菌发酵乳基质。本文所用的“发酵乳制品”包括但不限于嗜热发酵乳制品例如酸奶,嗜温发酵乳制品例如酸奶油和酪乳,以及发酵乳清等产品。术语“嗜热菌”在本文中是指在高于35℃的温度下生长最好的微生物。工业上最有用的嗜热细菌包括链球菌属物种和乳杆菌属物种。术语“嗜热发酵”在本文中是指在高于约35℃的温度,例如在约35℃至约45℃下发酵。术语“嗜热发酵乳制品”是指通过嗜热起子培养物的嗜热发酵制备的发酵乳制品,并且包括诸如凝固酸奶、搅拌型酸奶和饮用酸奶(例如养乐多(yakult))等发酵乳制品。术语“嗜温菌”在本文中是指在中等温度(15℃-35℃)下生长最好的的微生物。工业上最有用的嗜温细菌包括乳球菌属物种和明串珠菌属物种。术语“嗜温发酵”在本文中是指在约22℃至约35℃的温度下发酵。术语“嗜温发酵乳制品”是指通过嗜温起子培养物的嗜温发酵制备的发酵乳制品,包括诸如酪乳、酸乳、发酵乳、斯美塔那(smetana)、酸奶油、克非尔(kefir)和新鲜奶酪等发酵乳制品,所述新鲜奶酪例如夸克(quark)、tvarog和奶油奶酪。在优选实施方案中,接种的嗜热链球菌细胞的浓度为每ml乳基质104-109cfu嗜热链球菌细胞,例如每ml乳基质104cfu至108cfu的嗜热链球菌细胞。在本发明方法的另一个优选实施方案中,嗜热链球菌菌株不能酸化9.5%b-乳,定义为当用106-107cfu/ml的所述嗜热链球菌菌株接种9.5%b-乳并在40℃下温育14小时时,导致小于1.0的ph降低,向乳基质中加入一定量的有效地触发嗜热链球菌酸化9.5%b-乳的化合物,定义为当用106-107cfu/ml的所述嗜热链球菌菌株接种9.5%b-乳并在40℃下温育14小时时,导致1.0或更多的ph降低。优选地,所述化合物是蔗糖。优选,蔗糖的量为0.000001%-2%,例如0.00001%-0.2%,例如0.0001%-0.1%,例如0.001%-0.05%。在另一个优选的实施方案中,发酵乳制品是酸奶或奶酪。通过用嗜热链球菌和德氏乳杆菌保加利亚亚种发酵制备的奶酪的实例包括马苏里拉奶酪和比萨奶酪(etal.(2010),于thetechnologyofcheesemaking,2nded.blackwellpublishing,oxford;166-192)。优选,发酵乳制品是酸奶。在本发明的上下文中,酸奶起子培养物是包含至少一种德氏乳杆菌保加利亚亚种菌株和至少一种嗜热链球菌菌株的细菌培养物。据此,术语“酸奶”是指通过可用包含德氏乳杆菌保加利亚亚种菌株和嗜热链球菌菌株的组合物接种并发酵乳而获得的发酵乳制品。发酵乳制品本发明还涉及包含本发明的组合物的发酵乳制品。在另一个优选的实施方案中,发酵乳制品可以是酸奶、奶酪、酸奶油和酪乳以及发酵乳清。优选,发酵乳制品是酸奶。本发明的组合物的用途本发明的另一方面涉及本发明的组合物用于制备发酵乳制品的用途。以下通过非限制性实施例描述本发明的实施方案。序列表seqidno.1显示了菌株chcc16731的突变葡糖激酶基因的dna序列。seqidno.2显示了由seqidno.1编码的氨基酸序列。seqidno.3显示了菌株chcc16731的突变mann基因的dna序列。seqidno.4显示了由eqidno.3编码的氨基酸序列。seqidno.5显示了菌株chcc16731的manm基因(未突变)的dna序列。seqidno.6显示了由seqidno.5编码的氨基酸序列。实施例材料和方法培养基:对于嗜热链球菌,使用的培养基是本领域技术人员已知的m17培养基。m17琼脂培养基每升h2o具有以下组成:琼脂:12.75g抗坏血酸:0.5g酪蛋白胨(胰蛋白酶的):2.5gβ-甘油磷酸二钠五水合物:19g硫酸镁水合物:0.25g肉膏:5g肉蛋白胨(胃蛋白酶的):2.5g大豆蛋白胨(木瓜蛋白酶的):5g酵母膏:2.5g最终ph7.1±0.2(25℃)m17肉汤每升h2o具有下述组成:抗坏血酸:0.5g硫酸镁:0.25g肉膏:5g肉蛋白胨(胃蛋白酶的):2.5g甘油磷酸钠:19g大豆蛋白胨(木瓜蛋白酶的):5g胰蛋白胨:2.5g酵母膏:2.5g最终ph7.0±0.2(25℃)添加的碳源是无菌乳糖20g/l、葡萄糖20g/l或半乳糖20g/l。如本领域技术人员所知的,m17培养基是被认为适合嗜热链球菌生长的培养基。此外,如本领域技术人员所理解的,在本发明的上下文中,m17浓缩物可以由不同供应商供应,并且对于本文相关的感兴趣的细胞而言,人们(在标准测量不确定度内)独立于特定供应商将得到相同的本文相关的2-脱氧葡萄糖抗性结果。用于培养德氏乳杆菌保加利亚亚种的培养基是mrs-im培养基。mrs-im以琼脂平板或肉汤的形式使用。mrs-im琼脂培养基每升h2o具有以下组成:高压灭菌后在25℃下将ph调节至6.9±0.1。下文实施例中液体培养物所用的的mrs-im肉汤每升h2o具有以下组成:高压灭菌后在25℃下将ph调节至6.9±0.1。首先将碳源即乳糖20g/l或葡萄糖20g/l进行无菌过滤,然后加入到高压灭菌的肉汤中。可以在一定程度上改变上述mrs-im培养基,而不影响培养基支持德氏乳杆菌保加利亚亚种生长的能力。此外,如本领域技术人员将理解,mrs-im浓缩物或上文所述的各种组分可从不同的供应商获得,并用于制备mrs-im培养基。这些培养基将同样被用于下文的实施例中,特别是用于2-脱氧葡萄糖抗性选择测定中。母株嗜热链球菌chcc11976(如wo2011/026863中所述,在galk基因中具有突变的半乳糖发酵菌株)。嗜热链球菌chcc12339(如wo2011/092300中所述的噬菌体抗性和质构化)。嗜热链球菌chcc18948(cgcc12339的在galk基因中具有突变的半乳糖发酵突变体)2-脱氧-葡萄糖抗性菌株嗜热链球菌chcc16165(chcc11976的2-脱氧葡萄糖抗性突变体)。嗜热链球菌chcc16731(chcc16165的增甜和质构化突变体)。嗜热链球菌chcc19216(chcc18948的增甜和质构化突变体)。实施例1:使用2-脱氧葡萄糖分离嗜热链球菌chcc11976的具有增强的葡萄糖分泌的葡糖激酶突变体。为了分离嗜热链球菌菌株chcc11976的突变体,将衍生于单菌落生长的细胞接种到10ml含有2%乳糖的m17肉汤中,并在40℃下生长过夜。第二天,将所述菌株连续稀释铺板于含有2%半乳糖和浓度为20mm的2-脱氧葡萄糖的m17琼脂平板上(chcc11976),并在40℃下温育20小时。首先将抗性菌落在与选择其时相同类型的琼脂平板上重新划线培养。幸存者用于接种含有2%乳糖、2%半乳糖或2%葡萄糖的新鲜m17肉汤,并测量生长。由此,如实施例2中概述的那样,鉴定了能够在半乳糖上比在葡萄糖上生长得快的许多突变体。一种这样的突变体是chcc16165。实施例2:2-脱氧葡萄糖抗性突变体生长模式为了确保选择能够在半乳糖上生长的2-脱氧葡萄糖抗性突变体,使用选自半乳糖发酵菌株集合的两种菌株。尽管这些半乳糖发酵菌株在m17肉汤+2%葡萄糖中仍然比在m17肉汤+2%半乳糖中在指数期生长得快至少10%,但另一方面,chcc11976的2-脱氧葡萄糖抗性突变体衍生物,即chcc16165的特征是在m17培养液+2%半乳糖中比在m17培养液+2%葡萄糖中在指数期生长得更快。指数期的生长在本文中测量为在40℃下以指数生长的培养物在600纳米(od600)下的光密度随时间的发展。如本领域技术人员所知,当培养物处于指数生长时,其可以随着物种而变化。本领域技术人员将知道如何测定指数期的生长,例如,在od6000.1-1.0之间。在分光光度计中测量培养物的光密度(od)。结论基于本实施例2的2-脱氧葡萄糖抗性突变体生长模式-对于特定感兴趣的菌株(例如来自相关商品的一种菌株)-技术人员可以常规测试这种特定感兴趣的菌株是否具有为所选突变体特性的本文相关生长模式。实施例3:选择嗜热链球菌chcc16165的高乳糖发酵和葡萄糖分泌突变体为了分离嗜热链球菌菌株chcc16165的高乳糖发酵和葡萄糖分泌突变体,将衍生自单个菌落生长的细胞接种于10ml含有2%半乳糖的m17肉汤中,并在40℃下过夜生长。第二天,将所述菌株连续稀释平板接种于含有2%蔗糖和浓度为40mm的2-脱氧葡萄糖的m17琼脂平板上,并在40℃下温育20小时。从平板上挑取10个随机菌落,并用于接种含有2%蔗糖的新鲜m17肉汤,并在40℃下温育过夜。将chcc16165突变体转移到含有2%蔗糖的新鲜m17培养基中。实施例4.发酵乳的碳水化合物分析。使实施例3获得的突变体和菌株chcc16165在含有0.01%蔗糖的9.6%b-乳中生长。酸化后,用chcc16165酸化的乳的乳糖浓度为14.9,半乳糖浓度为8.4,葡萄糖浓度为5.7。相比之下,用chcc16165的表现最佳的突变体酸化的乳的乳糖浓度为9.3,半乳糖浓度为10.5,葡萄糖浓度为9.9。所述chcc16165的最佳突变体被命名为chcc16731,当与德氏乳杆菌保加利亚亚种菌株chcc16159和chcc16160(描述于wo2013/160413)组合使用时,使其在乳发酵中进一步经受测试。将chcc16165和chcc16731的过夜培养物与chcc16159和chcc16160中的每一种的组合加入到200mlb-乳样品中,并在40℃下酸化。结果列于表1中:从表1的结果可以看出,chcc16731能够从乳中去除几乎所有的乳糖。此外,chcc16731产生并分泌高水平的半乳糖和葡萄糖到乳中。考虑到蔗糖为100参照,且乳糖的甜度为16,半乳糖的甜度为32,葡萄糖的甜度为74.3,用chcc16731生产的发酵乳制品的甜度大幅增加。实施例5.嗜热链球菌chcc12339的半乳糖阳性突变体的产生使嗜热链球菌chcc12339在40℃下在含有2%乳糖的m17中过夜生长。将过夜培养物平板接种(100μl)于含有2%半乳糖的m17平板上,并在40℃下在厌氧条件下温育。在含有2%半乳糖的m17平板上出现22个菌落。将其中的16个菌落在相同类型的平板上重新划线培养,并且将其中8个菌落转移到含有2%半乳糖的液体m17中,所有这些导致生长,并且将这些培养物中的4个冷冻。将冷冻培养物中的2个涂布在含有2%半乳糖和40mm2-脱氧葡萄糖的m17平板上,并且对于每种培养物,选择所得菌落中的8个。将16种2-脱氧葡萄糖突变体转移到含有2%乳糖和2%半乳糖的液体m17中,并在40℃生长过夜,然后使突变体在含有2%半乳糖的液体m17中生长3个晚上,测量半乳糖和葡萄糖的产量,并追踪其生长。基于半乳糖和葡萄糖的产量,通过向200ml含有0.05%蔗糖的9.5%b-乳中加入2ml培养物,选择7种培养物用于乳酸化测试。3种突变体具有与母株chcc12339类似的酸化特征,并且将这3种突变体划线培养成单个菌落,并温育48小时。随后,将所述菌落转移至含有2%半乳糖的液体m17中,并且相对于ch12339测试3种突变体的生长。选择生长最快的突变体,并通过3次划线培养成单个菌落再次纯化,以产生纯化的命名为chcc18948的半乳糖阳性菌株(chcc12339的半乳糖阳性变体)。实施例6.产生嗜热链球菌chcc18948的葡萄糖分泌增强的2-脱氧葡萄糖突变体将100μl的chcc18949培养物涂布在含有2%半乳糖和20mm或40mm2-脱氧葡萄糖的m17平板上,并在40℃下温育过夜。从20mm平板中选择8个菌落,从40mm平板中选择8个菌落,并使其在含有2%半乳糖和20mm2-脱氧葡萄糖的液体m17中生长。将所得的培养物冷冻并重新划线培养,并且将所得菌落中的12个在m17培养基中开始,并用于酸化b-乳。将2-脱氧葡萄糖突变体的1%的过夜培养物接种到200ml含有0.05%蔗糖的9.5%b-乳中并在40℃下酸化。chcc18948的所有12种2-脱氧葡萄糖突变体都产生具有低乳糖浓度和高半乳糖和葡萄糖浓度的发酵乳制品。选择7种突变体进行单独重新测试并与德氏乳杆菌保加利亚亚种株chcc16159(描述于wo2013/160413)一起测试。再次将2-脱氧葡萄糖突变体的1%的过夜培养物接种于200ml含有0.05%蔗糖的9.5%b-乳中,并在40℃下酸化。结果如表2所示:三种最好的突变体是突变体2、3和12,并且在含有0.05%蔗糖的b-乳中,单独且与德氏乳杆菌保加利亚亚种菌株chcc16159、chcc16160和chcc16161(描述于wo2013/160413中)组合被重新测试。结果如表3所示:基于上述数据,选择chch18948突变体3作为最佳突变体,并将其命名为chcc19216。从上述数据可以看出,chcc19216能从乳中去除几乎所有的乳糖。此外,chcc19216产生并分泌高水平的半乳糖和葡萄糖到乳中。考虑到蔗糖为100参照,且乳糖的甜度为16,半乳糖的甜度为32,葡萄糖的甜度为74.3,用chcc19216生产的发酵乳制品的甜度大幅增加。实施例7.chcc16731和chcc19216的质构化性质通过使用以下测定测量剪应力来测试菌株chcc16731和chcc19216的质构化特性:温育7天后,使发酵乳达到13℃,并用装配了钻孔圆盘的棒轻轻搅拌,直至样品均匀。使用以下设置在流变仪(antonpaarphysicaasc/dsr301流变仪(自动进样器),antongmbh,奥地利)上评估样品的流变特性:等待时间(以重建至有所初始的结构)在无振荡或旋转的情况下5分钟振荡(以测量g'和g”,用于计算g*)γ=0.3%,频率(f)=[0.5…8]hz在60s内6个测量点(每10s一个)旋转(以测量300l/s等下的剪应力)和在210s内的21个测量点(每10s一个),上升至300l/s,和在210s内的21个测量点(每10s一个),下降至0.2707l/s对于进一步的分析,选择300l/s下的剪应力。使用表4所列混合物中的菌株chcc16731和chcc19216,使用0.024%的接种体来酸化200ml9.5%b乳样品,直至ph达到4.55。结果如表4所示:lb:德氏乳杆菌保加利亚亚种菌株chcc16159st:嗜热链球菌从结果中可以看出,菌株chcc16731和chcc19216当用作培养混合物中唯一嗜热链球菌时产生剪应力分别为41.3pa和51.5pa的发酵乳制品。当与菌株chcc16731和chcc19216组合使用时,产生剪应力为43.3pa的发酵乳制品。与例如wo2013/160413公开的传统增甜菌株相比,这种水平的剪应力高。实施例8.与具有乳糖阳性、葡萄糖阳性lb的培养物相比,用于本发明培养物的培养组合物的德氏乳杆菌保加利亚亚种(lb)的风味和生长实验计划表5:所测试的培养物的组成本发明的培养物由三种葡萄糖缺陷型嗜热链球菌(st)和一种乳糖缺陷型lb组成。培养物2(比较培养物)由三种葡萄糖缺陷型st和一种乳糖阳性、葡萄糖阳性(传统)lb组成。制备具有表5所示组成的两种培养物,并在发酵中用作起子培养物以在43℃下产生酸奶,直至达到目标ph4.55。比较两种培养物的风味、lb生长、后酸化、碳水化合物和流变学。乳基质乳基质含有4.5%蛋白质、1.0%脂肪和0.1%蔗糖。将0.5%脂肪乳和1.5%脂肪乳混合,以使最终乳基质中的脂肪含量为1%,并添加0.1%蔗糖。使用脱脂乳粉(smp)将蛋白质水平调节至4.5%。水合作用在6℃下进行2小时。此后,将所述乳在65℃下在200/50巴下均化,并在95℃下巴氏灭菌5分钟。酸奶生产参数使用intap在线测量发酵过程中的ph。当酸奶达到ph4.55时,使用带有钻孔盘的棒破坏凝胶。然后将凝胶在后处理单元(ptu)中在2巴、25℃下处理,装入120ml酸奶杯中,并在6℃下储存28天。剪应力的测量使用实施例7所述的方法,不同之处在于样品不用配备有钻孔棒的棒搅拌,因为在ptu处理以获得样品的均匀性期间凝胶已经破裂。菌落形成单位的测量m17用作st的选择性培养基。mrslac(mrs+8%乳糖)用作lb的选择性培养基。由酸奶制成10×稀释系列。将每一稀释液分别平板接种在m17和mrslac琼脂平板上,并在37℃下厌氧培养2天。此后,计数菌落形成单位。酸化培养物1花费10.8小时达到4.55的ph。培养物2花费12.1小时达到4.55的ph。后酸化表6:后酸化ph0天第1天第7天第14天第21天第28天培养物14.554.534.514.544.504.54培养物24.554.514.504.514.504.54lb的生长表7:发酵结束时的培养物生长(菌落形成单位(cfu))从表7中可以看出,本发明培养物的lb细胞计数远高于用传统lb,即乳糖阳性、葡萄糖阳性lb的比较培养物的lb计数。因此,葡萄缺陷型st和乳糖缺陷型lb的组合在发酵结束时产生更高水平的lb。流变学培养物1在发酵结束时在300l/s下的剪应力为41.1pa。培养物2在发酵结束时在300l/s下的剪应力为27.3pa。碳水化合物表8:发酵结束时的碳水化合物水平风味通过品尝小组的评估来评定风味。培养物1没有任何异味。实施例9.与具有葡萄糖缺陷型lb的培养物相比,用于本发明培养物的培养组合物的德氏乳杆菌保加利亚亚种(lb)的风味和生长实验计划表9:所测试的培养物的组成本发明的培养物由三种葡萄糖缺陷型st和一种乳糖缺陷型lb组成。培养物2(比较培养物)由三种葡萄糖缺陷型st和一种乳糖阳性、葡萄糖缺陷型lb组成。制备具有表9所示组成的两种培养物,并在发酵中用作起子培养物以在43℃下产生酸奶,直至达到目标ph4.55。比较这两种培养物的风味(氨基酸水平)、挥发性有机组分、碳水化合物和流变学。乳基质两种乳基质使用脱脂乳粉(smp)或乳清蛋白(wp)来调节,并含有4.5%蛋白质、1%脂肪和0.1%蔗糖。将0.5%脂肪乳和1.5%脂肪乳混合,以使最终乳基质中的脂肪含量为1%,并添加0.1%蔗糖。使用smp或wp将蛋白质水平调节至4.5%。水合作用在6℃下进行2小时。此后,将乳在65℃下在200/50巴下均化,并在95℃下巴氏灭菌5分钟。酸奶生产参数使用cinac在奶瓶(babybottle)中在线测量发酵过程中的ph。当酸奶达到ph4.55时,使用带有钻孔盘的棒破碎凝胶并进行均化。此后,将酸奶储存在奶瓶中。剪应力的测量使用实施例7中描述的方法。流变学smp乳基质中的培养物1在发酵结束时在300l/s下的剪应力为46.2pa。wp乳基质中的培养物1在发酵结束时在300l/s下的剪应力为39.6pa。smp乳基质中的培养物2在发酵结束时在300l/s下的剪应力为42.0pa。wp乳基质中的培养物2在发酵结束时在300l/s下的剪应力为38.4pa。从结果可以看出,与培养物2相比,培养物1具有更高的剪应力。后酸化表10:后酸化lb的生长表11:发酵结束时的培养物生长(菌落形成单位(cfu))从表11中可以看出,本发明培养物的lb细胞计数低于具有乳糖阳性、葡萄糖缺陷型lb的比较培养物的lb计数。碳水化合物表12:发酵结束时的碳水化合物水平氨基酸表13:氨基酸水平含有葡萄糖阴性lb的样品(培养物2)具有量更高的tffa,其指向更高的蛋白水解活性,这反过来可导致令人不愉快的味道。此外,含有葡萄糖阴性lb的酸奶含有量更高的谷氨酸,已知其产生被描述为“肉汤”的异味。缬氨酸是一种苦味氨基酸,在含有葡萄糖阴性lb的酸奶中也以较高的量存在。挥发性有机化合物(voc)voc曲线可以指示样品的风味特征。测量了51种不同的挥发性有机化合物。对于wp乳基质中的培养物1,与wp乳基质中的培养物2相比,51种化合物中有44种(86%)的挥发性有机化合物的量较低。对于wp乳基质中的培养物1,与wp乳基质中的培养物2相比,51种化合物中有34种(67%)的挥发性有机化合物的量较低。总之,与比较培养物相比,本发明培养物的挥发性有机化合物(作为一组化合物观察)含量显著更低。实施例10.用于本发明的培养组合物的德氏乳杆菌保加利亚亚种(lb)的生长和后酸化。实验计划表14:所测试的培养物的组成制备具有表14所示组成的培养物,并在发酵中用作起子培养物以在43℃下产生酸奶,直至达到目标ph4.55。以0.01%、0.02%和0.03%的水平接种培养物。将所产生的酸奶在5℃、13℃和25℃下储存。测试培养物的st和lb生长以及后酸化。乳基质表15:乳基质1和2的组成表16:乳基质3的组成量蛋白质碳水化合物脂肪脱脂乳884.2g1.5%乳1767.1gpromilk802fb10.9gsmp73.9g蔗糖6.0g水257.8g乳基质3000g4.57%5.71%0.92%后酸化表17:后酸化乳基质111333接种0.01%0.02%0.03%0.01%0.02%0.03%5℃第1天4.544.544.534.534.534.525℃第7天4.504.504.494.514.514.515℃第21天4.484.484.494.504.504.505℃第42天4.434.424.454.434.454.4213℃第1天4.504.494.504.514.504.5013℃第7天4.464.464.454.504.494.5013℃第21天4.384.384.384.424.454.4225℃第1天4.434.434.434.454.464.4525℃第7天4.404.404.404.434.444.43从表17可以看出,对于测试的所有接种水平和储存温度而言,后酸化水平非常低,包括13℃和25℃的较高储存温度,其中对于大多数传统起子培养物而言lb的生长水平高并且因此后酸化高。lb和st的生长表18:lb和st的生长乳基质111222接种0.01%0.02%0.03%0.01%0.02%0.03%lbcfu/g5.8e056.5e057.2e053.7e054.7e058.8e05stcfu/g4.2e076.5e077.4e073.8e075.6e077.0e07如表18所示,对于所有接种水平,lb细胞计数在发酵结束时处于高水平。实施例11.与传统起子培养物相比,用于本发明的培养组合物的德氏乳杆菌保加利亚亚种(lb)的生长、后酸化和碳水化合物分析实验计划表19:所测试的培养物的组成制备具有表19所示组成的培养物,并在发酵中用作起子培养物以在43℃下产生酸奶,直至达到目标ph4.55。以0.02%的水平接种培养物。将所产生的酸奶在5℃和13℃下储存。针对st和lb生长、后酸化和碳水化合物分析测试培养物。yoflexpremium1.0和yoflexyf-l901是商业传统培养物,包含乳糖阳性和葡萄糖阳性的st和lb。乳基质表20:乳基质1的组成量蛋白质碳水化合物脂肪1.5%乳3000gsmp85g乳基质3085g4.20%6.06%1.49%后酸化表21:后酸化样品1234566℃第1天4.494.484.544.544.534.546℃第7天4.454.434.524.524.524.536℃第21天4.444.374.474.474.464.46如表21所示,本发明的培养物具有低水平的后酸化,处于与传统起子培养物相同的水平。lb和st的生长表22:lb和st的生长样品123456lbcfu/g6.1e041.7e053.1e062.8e063.1e062.8e06stcfu/g3.0e084.1e083.2e083.1e083.0e083.1e08如表22所示,本发明培养物的lb细胞计数在发酵结束时比传统培养物水平高。碳水化合物分析表23:碳水化合物分析lod:检测限碳水化合物的所有测定均一式两份进行,表23列出了两次测定的平均值。在酸奶储存7天后进行测定。从表23中可以看出,本发明的培养物产生高水平葡萄糖,并且在储存7天后,葡萄糖以约8mg/g的水平存在。保藏和专家解决方案申请人要求,以下所述的保藏的微生物样品在专利授予日之前只供专家使用。菌株嗜热链球菌chcc11342,在2009年9月8日保藏于德国布伦瑞克因霍芬街7bd-38124(inhoffenstr.7b,d-38124braunschweig)的德国微生物保藏中心(dsmz),保藏号为dsm22932。菌株嗜热链球菌chcc12339,已经在2010年10月14日保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm24090。菌株嗜热链球菌chcc11976,已经在2009年9月8日保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm22934。菌株嗜热链球菌chcc16731,已经在2014年6月4日保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm28889。菌株嗜热链球菌chcc19216,已经在2015年12月8日保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm32227。菌株嗜热链球菌chcc15757菌株,在2012年4月3日保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm25850。菌株嗜热链球菌chcc15887菌株,在2012年4月3日保藏于布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm25851。菌株嗜热链球菌chcc16404菌株,在2012年12月12日保藏于德国布伦瑞克因霍芬街7bd-38124的德国微生物保藏中心(dsmz),保藏号为dsm26722。菌株德氏乳杆菌保加利亚亚种chcc18944,在2014年6月12日保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心,保藏号为dsm28910。菌株德氏乳杆菌保加利亚亚种chcc10019,在2007年04月03日保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心,保藏号为dsm19252。菌株德氏乳杆菌保加利亚亚种chcc16159,在2012年09月06日保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心,保藏号为dsm26420。菌株德氏乳杆菌保加利亚亚种chcc16160,在2012年09月06日保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心,保藏号为dsm26421。菌株德氏乳杆菌保加利亚亚种chcc12813,在2010年09月29日保藏于布伦瑞克因霍芬街7bd-38124的dsmz-德国微生物保藏中心,保藏号为dsm24074。这些保藏是根据有关国际认可的用于专利程序目的微生物保藏的布达佩斯条约进行的。参考文献wo2011/026863wo2011/092300wo2013/160413pooletal.(2006)metabolicengineering8(5);456-464thompsonetal.(1985)j.bacteriol.162(1);217–223chervauxetal.(2000).applandenvironmicrobiol,66,5306-5311cochuetal.(2003).applandenvironmicrobiol,69(9),5423-5432etal.(2010)inthetechnologyofcheesemaking,2nded.blackwellpublishing,oxford;166-192.序列表<110>科·汉森有限公司<120>用于制备具有增加的天然甜味和高质构的发酵食品的乳酸菌<130>p6233pc00<160>6<170>patentinversion3.5<210>1<211>966<212>dna<213>嗜热链球菌(streptococcusthermophilus)<400>1atgagtaagaaactcttaggtattgaccttggtggaacaactgttaagtttggtattttg60actgcagatggtgaagttcaagaaaaatgggctattgaaacaaatacgtttgaaaatggt120agccacattgttcctgacattgtagaatctttgaaacaccgtttggaattgtatggactt180actgctgaagattttattggaattggtatgggatctccaggtgcagttgaccgagaaaat240aaaacagtaacgggtgcctttaacttgaactgggcagaaactcaagaagttggctctgtt300attgaaaaagaacttggtattccattcgctattgataatgatgctaatgtggctgcactg360ggtgaacgttgggttggtgctggtgctaacaatcggaatgttgtctttataacattgggt420acaggtgttggtggcggtgttatcgctgatggtaacttaattcatggtgttgccggtgct480ggtggggaaattggtcacattattgttgaacctgacacaggatttgagtgtacttgcgga540aacaaggggtgtctggaaactgtagcttcagcaacaggtattgtacgtgtagcacatcat600ttggcagaaaaatacgaaggaaactcttctattaaagctgctgtagacaatggtgagttt660gtgacaagtaaagatattatcgtagctgctactgaaggtgataagtttgctgacagcatt720gttgataaagtctctaaatacctcagacttgcaacagcaaacatctcaaacattcttaac780ccagattctgtcgttatcggtggtggtgtttctgccgcaggagaattcttgcgtagtcgt840gttgaaggatactttacacgttatgcattcccacaagttcgccgtacaacaaaagtgaaa900ttagcggagcttggaaatgatgcaggaatcattggagctgctagtcttgcttatagtatt960gacaaa966<210>2<211>322<212>prt<213>嗜热链球菌<400>2metserlyslysleuleuglyileaspleuglyglythrthrvallys151015pheglyileleuthralaaspglygluvalglnglulystrpalaile202530gluthrasnthrphegluasnglyserhisilevalproaspileval354045gluserleulyshisargleugluleutyrglyleuthralagluasp505560pheileglyileglymetglyserproglyalavalasparggluasn65707580lysthrvalthrglyalapheasnleuasntrpalagluthrglnglu859095valglyservalileglulysgluleuglyileprophealaileasp100105110asnaspalaasnvalalaalaleuglygluargtrpvalglyalagly115120125alaasnasnargasnvalvalpheilethrleuglythrglyvalgly130135140glyglyvalilealaaspglyasnleuilehisglyvalalaglyala145150155160glyglygluileglyhisileilevalgluproaspthrglypheglu165170175cysthrcysglyasnlysglycysleugluthrvalalaseralathr180185190glyilevalargvalalahishisleualaglulystyrgluglyasn195200205serserilelysalaalavalaspasnglygluphevalthrserlys210215220aspileilevalalaalathrgluglyasplysphealaaspserile225230235240valasplysvalserlystyrleuargleualathralaasnileser245250255asnileleuasnproaspservalvalileglyglyglyvalserala260265270alaglyglupheleuargserargvalgluglytyrphethrargtyr275280285alapheproglnvalargargthrthrlysvallysleualagluleu290295300glyasnaspalaglyileileglyalaalaserleualatyrserile305310315320asplys<210>3<211>909<212>dna<213>嗜热链球菌<400>3atggctgaaaaaattcaattatctcaagcggatcgtaaaaaggtttggtggcgctcacaa60ttcttgcaaggtgcatggaactatgaacgtatgcaaaacttgggttgggcttactcactc120attcctgctatcaaaaaactttatactaacaaagaggaccaagccgcagctcttaaacgt180cacttggaattcttcaacactcacccttacgtagctgctcctatcataggggttccctta240gctcttgaagaagaaaaagctaatggtactgaaatcgaagatgcggctatccaaggggtt300aaaatcggtatgatgggtccacttgccggtatcggtgaccctgtcttctggttcacaatt360cgtccaattcttggtgcccttggtgcatcattggcacaagctggtaacattgctggtcca420cttatcttcttcattggttggaaccttatccgcatggccttcttgtggtacactcaagaa480cttggttacaaagcaggttcagaaatcactaaagacatatctggtggtatcttgaaagat540attactaaaggggcatcaatacttggtatgttcatcttggccgtcctcgttgaacgttgg600gtatctgtcgtcttcactgtaaagcttccaggtaaagttttgcctaaaggtgcttatatt660gaatggccaaaaggatatgttactggtgaccaactaaaaactatccttggtcaagtcaac720gataagcttagctttgataagattcaagtcgataccctacaaaaacaattggattcatta780attccaggtttgacgggacttctccttacttttgcatgtatgtggttgcttaagaagaaa840gtttcaccaatcacaatcatcatcggactctttgtagttggtattattgcaagcttcttc900ggaatcatg909<210>4<211>303<212>prt<213>嗜热链球菌<400>4metalaglulysileglnleuserglnalaasparglyslysvaltrp151015trpargserglnpheleuglnglyalatrpasntyrgluargmetgln202530asnleuglytrpalatyrserleuileproalailelyslysleutyr354045thrasnlysgluaspglnalaalaalaleulysarghisleugluphe505560pheasnthrhisprotyrvalalaalaproileileglyvalproleu65707580alaleuglugluglulysalaasnglythrgluilegluaspalaala859095ileglnglyvallysileglymetmetglyproleualaglyilegly100105110aspprovalphetrpphethrileargproileleuglyalaleugly115120125alaserleualaglnalaglyasnilealaglyproleuilephephe130135140ileglytrpasnleuileargmetalapheleutrptyrthrglnglu145150155160leuglytyrlysalaglysergluilethrlysaspileserglygly165170175ileleulysaspilethrlysglyalaserileleuglymetpheile180185190leualavalleuvalgluargtrpvalservalvalphethrvallys195200205leuproglylysvalleuprolysglyalatyrileglutrpprolys210215220glytyrvalthrglyaspglnleulysthrileleuglyglnvalasn225230235240asplysleuserpheasplysileglnvalaspthrleuglnlysgln245250255leuaspserleuileproglyleuthrglyleuleuleuthrpheala260265270cysmettrpleuleulyslyslysvalserproilethrileileile275280285glyleuphevalvalglyileilealaserphepheglyilemet290295300<210>5<211>828<212>dna<213>嗜热链球菌<400>5atgtcagatatgtcaattatttctgcgattttggtcgtagctgttgccttccttgctggt60cttgaaagtatccttgaccaattccaattccaccaaccacttgttgcatgtaccctcatc120ggtgctgccacaggtaacctcactgcaggtatcatgcttggtggttctcttcaaatgatt180acccttgcttgggcaaacatcggtgctgccgtagctcctgacgttgcccttgcatctgtt240gccgctgccatcattttggttaaaggtggtaaatttacagctgaaggtatcggtgttgcg300attgcaatagctatcctgcttgcagttgcaggtctcttcctaactatgcctgttcgtaca360gcatctattgcctttgttcatgctgcagataaagctgcagaacacggaaacatcgctggt420gttgaacgtgcatactacctcgctctccttcttcaaggtttgcgtattgctgtgccagca480gcccttcttcttgccatcccggcccaatctgttcaacatgcccttggcttgatgcctgac540tggctcacccatggtttggttgtcggtggtggtatggtcgtagccgttggttacgccatg600attatcaatatgatggctactcgtgaagtttggccattcttcgccattggttttgctttg660gcagcaattagccaattgacacttatcgctcttagtaccattggtgttgccatcgccttc720atctacctcaacctttctaaacaaggtggcggaaatggtggcggaaatggtggcggaact780tcatctggttcaggcgacccaatcggcgatatcttggaagactactag828<210>6<211>275<212>prt<213>嗜热链球菌<400>6metseraspmetserileileseralaileleuvalvalalavalala151015pheleualaglyleugluserileleuaspglnpheglnphehisgln202530proleuvalalacysthrleuileglyalaalathrglyasnleuthr354045alaglyilemetleuglyglyserleuglnmetilethrleualatrp505560alaasnileglyalaalavalalaproaspvalalaleualaserval65707580alaalaalaileileleuvallysglyglylysphethralaglugly859095ileglyvalalailealailealaileleuleualavalalaglyleu100105110pheleuthrmetprovalargthralaserilealaphevalhisala115120125alaasplysalaalagluhisglyasnilealaglyvalgluargala130135140tyrtyrleualaleuleuleuglnglyleuargilealavalproala145150155160alaleuleuleualaileproalaglnservalglnhisalaleugly165170175leumetproasptrpleuthrhisglyleuvalvalglyglyglymet180185190valvalalavalglytyralametileileasnmetmetalathrarg195200205gluvaltrpprophephealaileglyphealaleualaalaileser210215220glnleuthrleuilealaleuserthrileglyvalalailealaphe225230235240iletyrleuasnleuserlysglnglyglyglyasnglyglyglyasn245250255glyglyglythrserserglyserglyaspproileglyaspileleu260265270gluasptyr275pct/ro/134表当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1