一种来源于拟南芥的长链非编码RNAT5120及其用途的制作方法
本发明涉及植物基因工程技术领域,具体涉及一种来源于拟南芥的长链非编码rnat5120及其在提高硝态氮同化利用方面的用途。
背景技术:
2002年首次在小鼠cdna文库测序中发现了长链非编码rna(long-noncodingrna,lncrna)的存在。lncrnas是一类长度大于200个核苷酸、无明显开放阅读框、不编码蛋白的rna分子。长链非编码rna广泛存在于真核生物中,大多数lncrnas的转录由rna聚合酶ii完成,其结构与mrna类似,有帽子结构和polya尾巴;还有一部分lncrnas的转录通过rna聚合酶iii/iv/v的作用完成,其主要作为小干扰rna(sirna)的前体rna或参与介导基因转录沉默等(arieletal.,2014,molecularcell55,383-396)。与mrna相比,lncrnas的表达具有更强的组织或细胞特异性,但其表达丰度普遍较低。起初认为lncrnas是生物体转录的副产物,不具有生物学功能,但越来越多的研究表明其在多个生物进程中发挥重要的调节作用。近年来,随着高通量测序和生物信息学技术的快速发展,转录组测序(rna-seq)技术已成为研究lncrnas的重要工具之一(leietal.,2012,bmcbioinformatics13,325-331)。
长链非编码rna在人类和动物中的发现和作用研究非常多,但其在植物生长发育中的功能鉴定相对较少。虽然lncrnas在植物中的研究十分有限,但仍有一部分lncrnas的功能和作用机制得到详细的解析。lncrnas的调控机制十分复杂,其可在转录、转录后、表观遗传的水平上调控蛋白编码基因的表达进而影响植物的生长发育。lncrnas可以通过介导染色质重塑调控相关基因的表达;lncrnas还可以做为mirnas的靶标影响mirnas与靶基因的结合;lncrnas也可与蛋白结合进而改变蛋白的构象或影响蛋白的定位,若其与rna结合蛋白结合则会影响mrna的可变剪切或加工;另外,部分lncrnas可以直接与染色质结合调控基因表达。由于lncrnas调控机制的多样性,使得lncrnas作为新的一类重要的调节子参与到多种多样的生物进程中(heoetal.,2015,journalofplantbiology58,361-365)。
氮素是植物生长发育所需的营养元素之一,氮素的供应直接影响作物的生长与产量。由于植物对氮素的利用效率低,使得大量未被植物吸收利用的氮肥流失到环境中,造成严重的环境污染,因此,亟需加强关于作物吸收利用氮素规律方面的研究、提高作物的氮素利用率、培育高氮效作物新品种。大多数陆生作物如玉米、小麦等都是以吸收硝态氮为主。植物主要通过nrt1和nrt2两个硝态氮转运蛋白家族从外界吸收硝态氮并在体内对硝态氮进行转运分配。近年来的研究发现clc和slac两个蛋白家族在硝态氮的转运和存储中也发挥重要的作用(krappetal.,2010,jexpbot65,789-798)。吸收进入植物体内的硝态氮在硝态氮还原酶(nr)、亚硝态氮还原酶(nir)的作用下被还原为铵,而铵经由gs-gogat途径被还原为有机氮,直接被植物利用维持植物正常的生长发育。
硝态氮不仅是植物生长发育所需的营养物质,同时也是重要的信号分子,调控植物体内基因的表达及各个生物进程。硝态氮信号可以被分成两种类型:短期效应和长期效应。短期效应是指植物对硝态氮的初级响应,植物经硝态氮处理后,一些基因的表达会在短期内发生改变如nia1、nir、nrt1.1、nrt2.1等。长期效应是指硝态氮处理较长时间后,对植物生长发育产生的影响,包括种子萌发、根的形态建成、植物开花、气孔运动等。目前已鉴定出多个调控硝态氮信号的基因,如硝态氮感应子nrt1.1、转录因子nlp7、tga1/4、tcp20、可变剪切因子cpsf30、fip1、蛋白激酶cipk8、cipk23等。其中nlp7是重要的硝态氮调控因子,属于nlp(nin-likeproteins)转录因子蛋白家族,含有两个保守的蛋白结构域rwp-rk和pb1,而rwp-rk结构域可以结合到dna元件上进而调控基因的表达。研究表明nlp7可通过与顺式作用元件(如硝态氮响应元件,nre)的结合调控氮素相关基因的表达,进而影响植物对硝态氮的初级响应和同化利用,调控植物的生长发育(marchiveetal.,2013,nat.commun.4(2),1713)。
长链非编码rna作为维持植物正常生长发育重要的调节子,但目前还未有其在硝态氮信号及代谢方面研究的报道。
技术实现要素:
针对上述现有技术,本发明的目的是提供一种来源于拟南芥的长链非编码rnat5120。本发明研究发现过表达长链非编码rnat5120能促进植物对硝态氮信号的初级响应且能提高植物对硝态氮的同化利用效率。
为实现上述目的,本发明采用如下技术方案:
本发明的第一方面,提供一种来源于拟南芥的长链非编码rna,命名为t5120,具有:
1)seqidno.1所示的核苷酸序列;或
2)与seqidno.1所示的核苷酸序列具有90%以上序列同源性,且功能上与seqidno.1所示的核苷酸序列等同。
进一步的,本发明还提供用于扩增上述拟南芥的长链非编码rna的引物对,其核苷酸序列分别如seqidno.2和seqidno.3所示。
本发明的第二方面,提供含有上述长链非编码rna的植物表达载体。
进一步的,本发明还提供包含上述植物表达载体的重组菌。
本发明的第三方面,提供上述长链非编码rna、植物表达载体或重组菌在如下a)-e)至少一项中的用途:
a)增强植物体内硝态氮还原酶的活性;
b)增加植物体内氨基酸的含量;
c)促进植物体内硝态氮同化相关基因的表达;
d)促进植物对硝态氮信号的初级响应;
e)提高植物对硝态氮的同化利用效率。
本发明的第四方面,提供一种提高植物对氮素利用率的方法,包括:将本发明第一方面所述的长链非编码rnat5120或者本发明第二方面所述的植物表达载体导入目标植物或植物组织,并使长链非编码rnat5120过表达。
本发明的第五方面,提供上述长链非编码rna、植物表达载体或重组菌在植物育种中的用途。
所述植物育种为培育高氮效作物品种。
本发明的第六方面,提供一种高氮效作物品种的培育方法,所述培养方法包括:将长链非编码rnat5120转入出发植株中,获得高氮效作物;或者上调出发植株基因组中长链非编码rnat5120的表达,筛选得到氮利用率提高的植株。
优选的,将长链非编码rnat5120转入出发植株中的方法包括:聚乙二醇法、农杆菌介导法或基因枪轰击法。
本发明的有益效果:
本发明首次发现了长链非编码rna在硝态氮信号及代谢中发挥重要的作用,为提高作物的氮素利用率、培育高氮效作物新品种提供了新的思路与方向。
附图说明
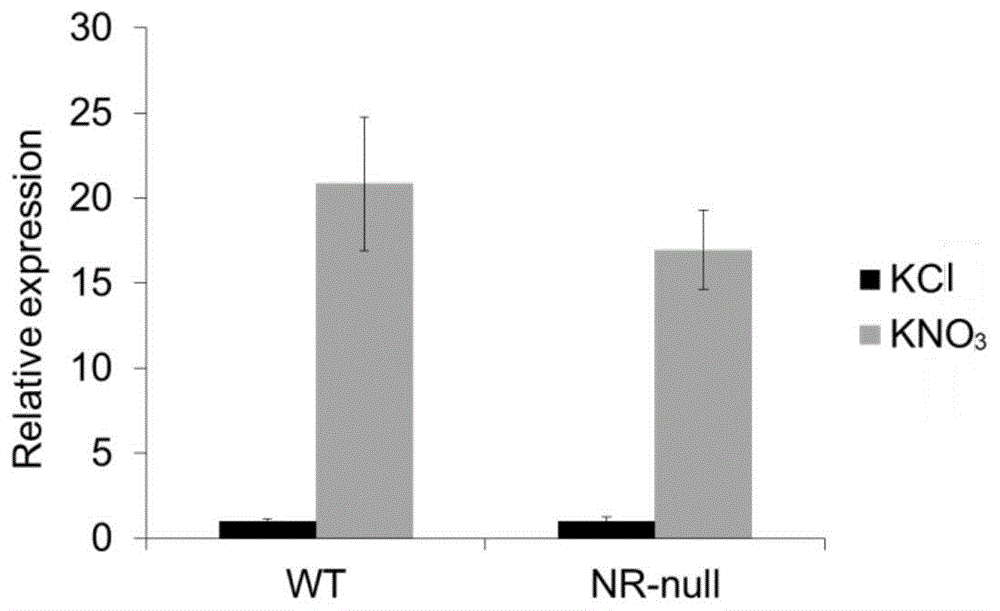
图1:no3-处理后,t5120表达量的检测。将wt、nr-null在只有nh4+为氮源的培养基上生长7天后,分别用10mmkno3和10mmkcl(作为对照)处理2小时,取根并提取rna用于qpcr检测,其中wt:野生型,nr-null:nia1nia2双突变体。
图2:t5120在转基因株系中的表达。wt、t5120-oe-1、t5120-oe-2在1/2ms培养上生长7天,取全苗并提取rna用于qpcr检测,其中wt:野生型,t5120-oe-1、t5120-oe-2:t5120/wt转基因株系。
图3:t5120过表达对no3-同化利用的调控。wt、t5120-oe-1、t5120-oe-2在1/2ms培养基上生长7天,取全苗用于相关实验的检测。(a-c)t5120过表达株系中no3-含量(a)、硝态氮还原酶(nr)活性(b)、氨基酸含量(c)的检测;(d)t5120过表达株系中硝态氮同化基因表达量的检测。其中wt:野生型,t5120-oe-1、t5120-oe-2:t5120/wt转基因株系。
图4:t5120过表达株系中硝态氮响应基因表达量的检测。wt、t5120-oe-1、t5120-oe-2在只有nh4+为氮源的培养基上生长7天后,分别用10mmkno3和10mmkcl(作为对照)处理2小时,取根并提取rna用于qpcr检测,其中wt:野生型,t5120-oe-1、t5120-oe-2:t5120/wt转基因株系。
图5:t5120作用于nlp7的下游调控硝态氮信号。(a)nlp7调控t5120的表达,wt、nlp7-1、nlp7-2、pnlp7::nlp7-gfp/nlp7-1和p35s::nlp7-gfp/nlp7-1在1/2ms培养上生长7天,qpcr检测t5120的表达量;(b)t5120/nlp7-4转基因株系中t5120的表达。wt、nlp7-4和t5120/nlp7-4转基因株系在1/2ms培养上生长7天,取全苗并提取rna用于qpcr检测,其中wt:野生型,t5120/nlp7-4-5、t5120/nlp7-4-11:t5120/nlp7-4转基因株系;(c)nr活性的检测。wt、nlp7-4和t5120/nlp7-4转基因株在1/2ms培养基上生长7天,取全苗用于检测nr活性;(d)qpcr检测wt、nlp7-4和t5120/nlp7-4转基因株系中硝态氮同化相关基因的表达。
具体实施方式
应该指出,以下详细说明都是例示性的,旨在对本申请提供进一步的说明。除非另有指明,本文使用的所有技术和科学术语具有与本申请所属技术领域的普通技术人员通常理解的相同含义。
正如背景技术部分所介绍的,长链非编码rna作为维持植物正常生长发育重要的调节子,但目前还未有其在硝态氮信号及代谢方面研究的报道。基于此,本发明在长链非编码rna调控硝态氮信号及代谢方面进行了深入研究。
本发明将拟南芥野生型在只有nh4+为氮源的培养基上生长7天,用10mmkno3或10mmkcl(作对照)处理2小时,进行rna-seq测序。通过分析高通量测序结果,发现了一个新的长链非编码rnat5120的表达受no3-的诱导最为显著。本发明进一步利用qpcr技术检测了t5120在拟南芥野生型和硝态氮还原酶突变体(nr-null)中的no3-诱导量,实验结果表明t5120的表达直接受no3-的诱导。
为研究t5120在硝态氮信号及利用方面的功能,发明人首先利用pcr技术克隆了该长链非编码rna的基因片段,以生长7天的幼苗基因组为模板,利用下述引物扩增lncrnat5120的基因片段:
上游引物:5’-acacggatcctgacgtcggcaacatatagc-3’,其核苷酸序列如seqidno.2所示;
下游引物:5’-acacgtcgacgggcggtatattgactcgaa-3’,其核苷酸序列如seqidno.3所示。
在获得上述长链非编码rna之后发明人将其转入受体植物拟南芥中,获得t5120过量表达的转基因株系。在1/2ms培养基上培养转入t5120基因的拟南芥和受体拟南芥,并检测两种植物对硝态氮的吸收利用,结果发现过表达t5120基因能够明显增强硝态氮还原酶的活性、增加植物体内氨基酸的含量、促进硝态氮同化相关基因的表达。发明人还发现经硝态氮处理后,转入t5120基因的拟南芥植株体内响应硝态氮的相关基因表达量显著升高,促进了植物对硝态氮的初级响应。
综上,本发明克隆出一个新的拟南芥长链非编码rna,并将其命名为t5120,其核苷酸序列如seqidno.1所示。该长链非编码rna的表达受硝态氮的强烈诱导;利用该长链非编码rna转入后的拟南芥与受体拟南芥相比,其体内硝态氮含量降低、硝酸盐还原酶活性和氨基酸含量显著升高、硝态氮同化相关基因的表达量显著增加;经硝态氮处理后,其体内响应硝态氮的基因的表达水平明显升高。结果表明,过表达t5120能促进植物对硝态氮信号的初级响应且能提高植物对硝态氮的同化利用效率,由此提出了本发明。
由于核苷酸序列的特殊性,任何seqidno.1所示多核苷酸的变体,只要其与该核苷酸具有90%以上同源性,且具有相同的功能,则均属于本发明保护范围之列。所述多核苷酸的变体是指一种具有一个或多个核苷酸改变的多核苷酸序列。此多核苷酸的变体可以是包括取代变异体、缺失变异体和插入变异体。
基于本发明的拟南芥长链非编码rna的性能,可以用于提高植物对氮素的利用率。在本发明的一种实施方案中,给出了提高植物对氮素利用率的方法,包括下列步骤:
(1)获得拟南芥长链非编码rnat5120的核苷酸序列;
(2)用rcr技术获得t5120基因的片段,并将其连接至植物表达载体上;
(3)将带有目的基因质粒的农杆菌转入目标植物,获得t5120过表达的植物。
发明人还分析了t5120与另一重要的硝态氮调控基因nlp7的作用关系。通过检测t5120在nlp7突变体中的表达量,发现nlp7突变体中t5120的表达显著下降,说明t5120的表达受nlp7的调控;同时,将t5120基因转入nlp7突变体中,构建t5120/nlp7的转基因株系并对其氮素利用率进行检测,发现与受体植株nlp7相比,转入了t5120基因的nlp7植株体内硝态氮还原酶活性升高、硝态氮同化相关基因的表达量明显增加,说明长链非编码rnat5120能促进植物对氮素的同化利用能力。
综上所述,本发明所提供的长链非编码rnat5120的表达受no3-的强烈诱导,在植株体内过表达该长链非编码rna能增强植物对硝态氮的初级响应,促进植物对硝态氮的同化利用。
为了使得本领域技术人员能够更加清楚地了解本申请的技术方案,以下将结合具体的实施例详细说明本申请的技术方案。
本发明实施例中所用的试验材料均为本领域常规的试验材料,均可通过商业渠道购买得到。未注明详细条件的实验方法是按照常规试验方法或按照供应商所建议的操作说明书进行的。
实施例1:硝态氮响应lncrnat5120的鉴定
将拟南芥野生型在2.5mmnh4suc培养基上生长7天,用10mmkno3或10mmkcl(作对照)处理2小时,进行rna-seq测序。分析高通量测序结果,筛选能响应硝态氮的lncrnas,其中lncrnat5120的表达受no3-处理后变化最明显。为验证rna-seq测序的结果,将野生型和nr-null(nia1nia2)突变体在上述培养条件下生长并进行相应的处理,取根并提取rna,利用试剂盒将rna反转录成cdna,以cdna为模板,进行qpcr实验,结果发现在no3-处理后的野生型和nr-null突变体中t5120的表达明显升高,说明t5120的表达直接受no3-的诱导(图1)。
其中进行qpcr实验所用的引物序列如下:
上游引物:5’-tgaaccacttgacacatgcg-3’,其核苷酸序列如seqidno.4所示;
下游引物:5’-agtgccatattttgagtaccaca-3’,其核苷酸序列如seqidno.5所示。
2.5mmnh4suc培养基配方:
实施例2:lncrnat5120植物表达载体的构建
将拟南芥野生型在1/2ms培养基上生长7天,取全苗并提取基因组dna。以提取的dna为模板,利用高保真dna聚合酶扩增t5120的基因片段,pcr所用的引物序列为:上游引物:5’-acacggatcctgacgtcggcaacatatagc-3’,其核苷酸序列如seqidno.2所示;下游引物:5’-acacgtcgacgggcggtatattgactcgaa-3’,其核苷酸序列如seqidno.3所示。将pcr产物在1%的琼脂糖中进行电泳,根据dnamarker找到目的条带并将此胶块切下。利用胶回收试剂盒(购自omega公司)回收目的dna,对回收的目的dna及植物表达载体pzp211进行酶切(bamhi和sali),然后将酶切的dna片段和载体用t4dna连接酶连接。随后利用热激法将连接产物转化至大肠杆菌mach1感受态中,于37℃摇床,230rpm震荡培养1小时,将摇好的菌液涂布于含spe+抗性的lb平板上,37℃倒置培养12小时。通过菌落pcr技术筛选成功转入重组载体的阳性克隆,挑取阳性克隆的菌体于5mllb培养液中培养并提取质粒用于测序。将测序结果与t5120的序列进行比对,确认无误后将该质粒转化至农杆菌gv3101感受态中,放置于28℃摇床,230rpm震荡培养3小时,将菌液均匀地涂布于含spe+抗性的lb平板上,28℃倒置培养36小时。菌落pcr鉴定成功转入重组质粒的农杆菌并将菌种在-80℃中保存。
1/2ms培养基配方:
注:定容后,用koh调ph值为5.7.
lb液体培养基配方:
实施例3:转基因阳性植株的鉴定
挑取-80℃保存的农杆菌菌体于250mllb培养液中,放置于28℃摇床,震荡培养16小时。利用浸花法将农杆菌侵染至拟南芥野生型植株。待植株成熟后,收取种子,并将种子铺在含k+抗性的1/2ms平皿上,筛选阳性苗即转基因株系t1代。将筛选的t1代种子移植至蛭石中培养,收获t2代种子。将t2代种子在含k+抗性的1/2ms平皿上培养,发现其存活率为3/4,将存活的t2植株移至蛭石中培养,单株收获t3代种子。将t3代种子在含k+抗性的1/2ms平皿上培养,筛选纯合的转基因株系并移至蛭石中扩繁。
实施例4:t5120转基因植株的功能鉴定
利用qpcr技术检测上述实验获得的纯合转基因株系中t5120的表达量,获得t5120过表达的转基因株系(图2)。首先对转基因株系的硝态氮含量、硝酸还原酶活性、氨基酸含量进行检测,结果发现,t5120过表达株系中硝态氮含量降低、nr活性和氨基酸含量升高,接下来又利用qpcr技术检测了t5120过表达株系中负责硝态氮同化的相关基因的表达,发现部分同化基因的表达在t5120过表达株系中的表达量显著升高(图3)。同时,利用qpcr技术检测了t5120过表达株系在no3-处理后,硝态氮响应基因nia1、nir、nrt2.1的表达,发现这些基因的no3-诱导量在t5120过表达株系中明显增加(图4)。以上这些实验结果表明lncrnat5120能促进植物对硝态氮的同化及响应。
实施例5:t5120与nlp7作用关系研究
将野生型、nlp7突变体、nlp7/nlp7转基因株系在1/2ms培养基上生长7天,取材后利用qpcr检测这些株系中t5120的表达,结果表明nlp7可以调控t5120的表达(图5a);为进一步研究两者的作用关系,我们将实施例2中获得的能表达lncrnat5120的重组质粒转入nlp7-4突变体中,通过qpcr检测发现在t5120/nlp7-4的转基因株系中t5120的表达显著升高(图5b)。将wt、nlp7-4、t5120/nlp7-4株系在1/2ms培养基上生长7天,检测硝态氮还原酶活性及相关同化基因的表达,发现在nlp7-4突变体中过表达t5120后,转基因株系的nr活性及no3-同化基因的表达量都显著高于nlp7-4突变体,但仍低于wt(图5c和图5d)。以上结果表明t5120能部分恢复nlp7-4突变体在硝态氮响应及同化方面的缺陷,在硝态氮信号调控中t5120作用于nlp7的下游发挥作用。
以上所述仅为本申请的优选实施例而已,并不用于限制本申请,对于本领域的技术人员来说,本申请可以有各种更改和变化。凡在本申请的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本申请的保护范围之内。
sequencelisting
<110>山东农业大学
<120>一种来源于拟南芥的长链非编码rnat5120及其用途
<130>2019
<160>5
<170>patentinversion3.5
<210>1
<211>589
<212>dna
<213>拟南芥(arabidopsisthaliana)
<400>1
ttagggaaacaacgacgaccctttcgtgaccctttgagccactttcactacatttacatt60
tattcatttctccttacaatatacatattctcctttttaacatgtactatgcattcacaa120
accatgtcgtactatataaaaatatacattgacgacacaaatcaaataagtatatagatt180
ttagtatgtatttatcttgaaatatatggaaaacaaccatgtcgtactgcacaaaataat240
ttgatggaaacaaattatcatacatacaaagcaatggccagaatatcccaaacattagct300
caccgtcaatttaaacacaattatcctggagataatcataattctaatccctattgtggt360
gttttaaaattctcatgttgattttgtctaaaagcagcacttttgctatcatataaacat420
atttaagcagcacttttcatttcatataacgaaataatgaaccacttgacacatgcggct480
atatttttttaaccatatgatatttgccatattggtatatagttggacacttttactgtt540
tatagtcaaagtcagtgtggtactcaaaatatggcacttacaaaagttg589
<210>2
<211>30
<212>dna
<213>人工序列
<400>2
acacggatcctgacgtcggcaacatatagc30
<210>3
<211>30
<212>dna
<213>人工序列
<400>3
acacgtcgacgggcggtatattgactcgaa30
<210>4
<211>20
<212>dna
<213>人工序列
<400>4
tgaaccacttgacacatgcg20
<210>5
<211>23
<212>dna
<213>人工序列
<400>5
agtgccatattttgagtaccaca23
- 还没有人留言评论。精彩留言会获得点赞!