光合作用和/或生物质生成增加的经基因修饰的高等植物、其方法和用途与流程
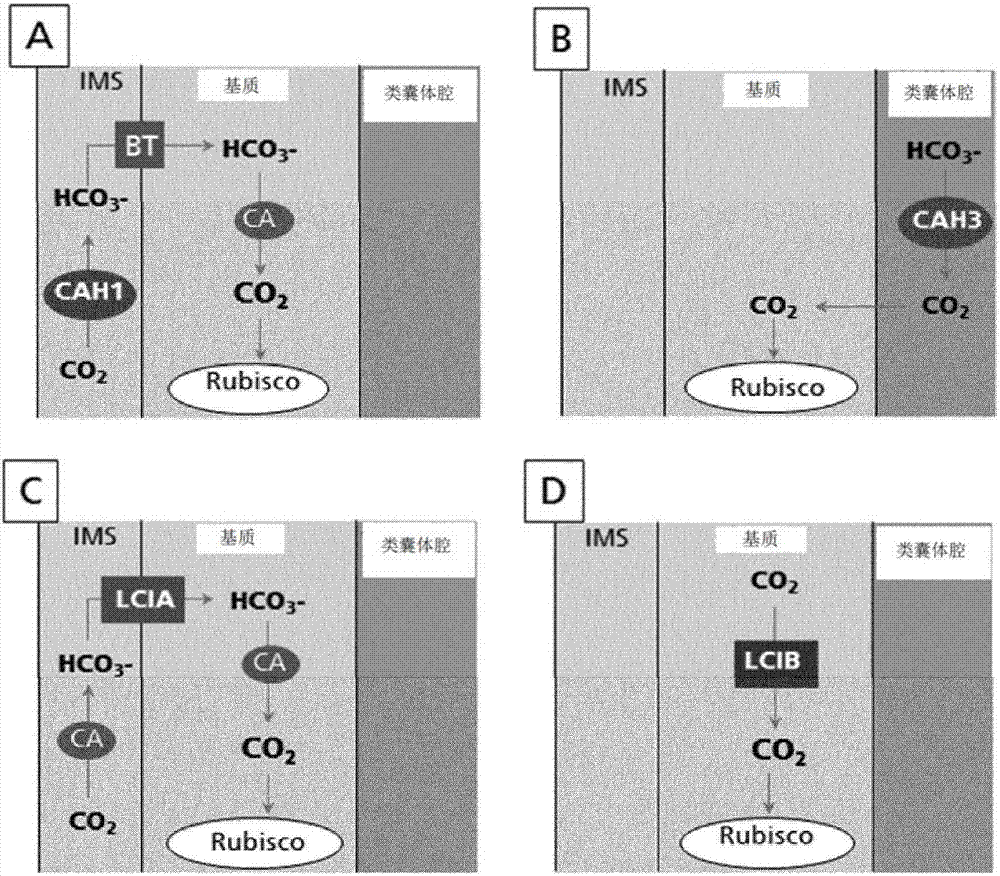
本文提供的技术整体涉及经基因修饰的高等植物,例如c3和c4植物及其后续世代,该经基因修饰的高等植物包括“莱茵衣藻(chlamydomonasreinhardtii)co2浓缩机制”(ccm)的至少一种基因产物(如mrna或蛋白质)的稳定和/或瞬时表达,其中该表达在高等植物的叶绿体的膜间隙和/或一个或多个亚细胞隔室中发生,并且其中该表达增加了经基因修饰的高等植物(t0)的和/或所述经基因修饰的高等植物的t1和/或t2世代或任何后续植物世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。在一个方面,本发明涉及高等植物,其包括衣藻属(chlamydomonas)碳酸氢盐转运蛋白(lcia、lcib)和碳酸酐酶(cah1、cah3)在c3或c4植物的叶绿体的不同亚细胞隔室中的单独或组合表达。在一些方面,lcia、lcib、cah1和cah3在经基因修饰的高等植物的典型定位中表达,在其他方面,它们在经基因修饰的高等植物的非典型定位中表达。一般来讲,本发明涉及改善的谷类、豆类、果类、根部和块茎、油料作物、纤维作物和乔木。此外,本发明涉及此类经基因修饰的高等植物的方法和用途。
背景技术:
:c3碳固定是用于光合作用的碳固定的三种代谢途径之一,其余两种是c4和cam。在c3过程中,二氧化碳和核酮糖二磷酸(rubp,5碳糖)通过以下反应转化为3-磷酸甘油酸:co2+rubp→(2)3-磷酸甘油酸。c3植物来源于中生代和古生代期间,早于c4植物,并且仍代表着大约95%的地球植物生物质。c3植物会经蒸腾作用丧失97%通过其根部吸收的水。单独以c3固定存活的植物(c3植物)往往在其中日光强度适度、温度适度、二氧化碳浓度为约200ppm或更高并且地下水丰富的区域中生长。c3植物的例子包括稻米、小麦、橘树、葡萄植物、咖啡植物、烟草植物、茶植物、花生植物、柠檬树、马铃薯、胡萝卜、番茄、桃树、苹果树、梨树、芒果树和大麦。另外的例子包括燕麦、裸麦、黑小麦、干菜豆、大豆、绿豆、蚕豆、豇豆、豌豆、鹰嘴豆、木豆、兵豆、香蕉、椰子、芋头、薯蓣、甘薯、木薯、甜菜、棉花、黄麻、剑麻、芝麻、向日葵、油菜籽和红花。c3植物不利于在炎热地区生长,因为随着温度的升高,核酮糖-1,5-二磷酸羧化酶/加氧酶(rubisco)将更多的氧掺入rubp。这导致了光呼吸作用的增加,从而导致植物的碳和氮的净损失,因此可限制生长。在干燥区域,c3植物关闭其气孔以减少水分丧失,但这阻止了co2进入叶片,因此减少了叶片中的co2浓缩。这降低了co2:o2比率,因此也增加了光呼吸作用。在另一个方面,对c4和cam植物进行了改进,以使它们能在炎热和干燥地区存活,因此它们优于c3植物。c4碳固定是用于碳固定的三种生物化学机制之一,其余两种是c3和cam光合作用。它因存在于称为c4植物的少数植物中的碳固定的第一个产物中的4碳分子而得名,相比之下,c3植物中为3碳分子产物。c4固定是对更常见的c3碳固定的细化,并且据信是更晚进化得来。c4和cam克服了在所谓的光呼吸作用中rubisco酶浪费于固定氧而非二氧化碳的趋势。这通过使用叶肉细胞中的更有效的固定co2的酶,并且关闭该通过进入维管束鞘细胞的苹果酸或天冬氨酸来进行的碳固定实现。在这些维管束鞘细胞中,rubisco与大气氧隔离,并被苹果酸或草酰乙酸的脱羧作用释放的co2饱和。然而,这些另外的步骤需要更多atp形式的能量。由于该额外能量要求,c4植物能够仅在某些条件下更有效地固定碳,而在其他条件下更常见的c3途径更有效。然而,也可根据本发明修饰c4,以便更进一步改善碳固定。c4植物的例子包括马唐、玉米(玉蜀黍)、苋菜、高粱、粟、甘蔗、香附、马唐、稗、四翅滨藜和藜科植物。衣藻属是由存在于死水和潮湿土壤中,存在于淡水、海水中,甚至雪中(如“雪生藻类”)的单细胞鞭毛藻组成的绿藻属。衣藻属被用作分子生物学,特别是鞭毛运动性和叶绿体动力学、生源论和遗传学的研究的模式生物。水生光合生物诸如莱茵衣藻可调节其光合作用,以通过诱导碳浓缩机制(ccm)来适应co2限制胁迫,该碳浓缩机制包括碳酸酐酶(cah)和无机碳(ci)转运蛋白。碳浓缩机制(ccm)允许莱茵衣藻优化用于光合作用的碳获取。当无机碳(ci;co2和/或hco3-)受到限制时,ccm起到促进co2同化的作用。通过活性ci摄取系统,提高了内部ci水平,然后碳酸酐酶通过积聚的碳酸氢盐的脱水作用为核酮糖-1,5-二磷酸羧化酶/加氧酶(“rubisco”)提供足够的co2。在本发明中,将新鉴定的ccm组分整合进高等植物,以增加所述经基因修饰的高等植物的后续植物世代,特别是t1和/或t2世代和任何其他世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。通过该修饰,高等植物不仅可以更快地生长,在已知的栖息地中更具竞争力,而且甚至可在该植物传统上未占据的气候条件下生长。高等植物的另外例子包括谷类、豆类、果类、根部和块茎、油料作物、纤维作物和乔木。技术实现要素:本发明的一个方面涉及经基因修饰的高等植物及其后续世代,所述经基因修饰的高等植物包括“莱茵衣藻co2浓缩机制”(ccm)的至少一种基因产物的稳定或瞬时表达,其中该表达在植物的叶绿体的膜间隙和/或一个或多个亚细胞隔室中发生。在一个方面,该表达增加了经基因修饰的高等植物(t0)的和/或所述经基因修饰的高等植物的t1和/或t2世代或任何后续世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。在一个方面,所述经基因修饰的高等植物是c3植物,在另一个方面,所述经基因修饰的高等植物是c4植物。另一个方面涉及选自莱茵衣藻的碳酸氢盐转运蛋白lcia、碳酸氢盐转运蛋白lcib、碳酸酐酶cah1和/或碳酸酐酶cah3的“莱茵衣藻co2浓缩机制”(ccm)的基因产物。在又一个方面,与野生型对照相比,cah1、cah3、lcia和lcib中的任一者的表达使所述经基因修饰的高等植物的t1世代生长更快和/或营养期更短。在一个方面,“莱茵衣藻co2浓缩机制”的所述基因产物是莱茵衣藻的碳酸酐酶cah1。在另一个方面,所述碳酸酐酶cah1在经基因修饰的高等植物的叶绿体的非典型定位膜间隙中表达。本发明的一个方面是在叶绿体的非典型隔室中的表达(比如如实例中所示的cah1的非典型表达)增加了经基因修饰的高等植物(t0)的和/或所述经基因修饰的高等植物的t1和/或t2世代或任何后续世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。在本发明的又一个方面,在叶绿体的典型隔室中的表达增加了经基因修饰的高等植物(t0)的和/或所述经基因修饰的高等植物的t1和/或t2世代或任何后续世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。在本发明的又一个方面,与野生型对照相比,在经基因修饰的高等植物的叶绿体的膜间隙中所述碳酸酐酶cah1的表达提高了所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代的光合速率至少8%、叶绿素水平至少7%和/或生物质至少22%。这些植物的叶片数量也增加。在本发明的一个方面,在cah1t2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+22%·鲜重:+26%·干重:+29%在本发明的又一个方面,在cah1t2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+142%/+22%·叶绿素含量:+15%/+13%·叶片数量:+21%/+24%·鲜重:+nd/+26%·干重:+nd/+29在本发明的另一个方面,“莱茵衣藻co2浓缩机制”(ccm)的所述基因产物是莱茵衣藻的碳酸酐酶cah3。在本发明的另一个方面,所述碳酸酐酶cah3在经基因修饰的高等植物的叶绿体的类囊体腔中表达。在本发明的又一个方面,与野生型对照相比,在经基因修饰的高等植物的叶绿体的类囊体腔中所述碳酸酐酶cah3的表达,使得在所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代中,与野生型对照相比,光合速率提高至少8%、叶绿素水平提高至少10%,气孔大小增加最多50%和/或生物质增加至少31%。在本发明的一个方面,在cah3t2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+24%·鲜重:+34%·干重:+31%在本发明的又一个方面,在cah3t2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+78%/+24%·叶绿素含量:+11%/+10%·叶片数量:+26%/+64%·鲜重:+nd/+34%·干重:+nd/+31%在本发明的另一个方面,“莱茵衣藻co2浓缩机制”(ccm)的所述基因产物是莱茵衣藻的碳酸氢盐转运蛋白lcia。在本发明的另一个方面,所述碳酸氢盐转运蛋白lcia在经基因修饰的高等植物的叶绿体的内被膜中表达。在本发明的又一个方面,在营养期结束时,与野生型对照相比,在经基因修饰的高等植物的叶绿体的内被膜中所述碳酸氢盐转运蛋白lcia的表达增加了所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代的光合速率至少8%、叶绿素水平至少15%和/或生物质至少33%。在本发明的一个方面,在lciat2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+33%·鲜重:+54%·干重:+41%在本发明的又一个方面,在lciat2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+93%/+33%·叶绿素含量:+11%/+16%·叶片数量:+17%/+19%·鲜重:+nd/+54%·干重:nd/+41%在本发明的另一个方面,“莱茵衣藻co2浓缩机制”(ccm)的所述基因产物是莱茵衣藻的碳酸氢盐转运蛋白lcib。在本发明的又一个方面,所述碳酸氢盐转运蛋白lcib在经基因修饰的高等植物的叶绿体的基质中表达。在本发明的又一个方面,与野生型对照相比,在经基因修饰的高等植物的叶绿体的基质中所述碳酸氢盐转运蛋白lcib的表达增加了所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代的生物质。在本发明的一个方面,在lcibt2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+23%·鲜重:+30%·干重:+28%在本发明的又一个方面,在lcibt2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+78%/+23%·叶绿素含量:+10%/+15%·叶片数量:+27%/+24%·鲜重:+nd/+30%·干重:+nd/+28%本发明的一个方面还涉及用于制备经基因修饰的高等植物的方法。本发明的一个方面还涉及经基因修饰的高等植物的用途,其中该高等植物的修饰,与野生型对照相比,增加了所述经基因修饰的高等植物所生成的天然和/或转基因产物的产率。本发明的另一个方面是经基因修饰的高等植物的用途,其中经基因修饰的高等植物所产生的天然产物选自植株的花、果实、种子、坚果、叶、茎和/或任何其他适于商业化部分。本发明的又一个方面是经基因修饰的高等植物的用途,其中经基因修饰的高等植物所生成的转基因产物选自核酸、蛋白质、肽和/或通过转基因方式生成的任何其他代谢产物。在本发明的一个方面,上述经基因修饰的高等植物是c3植物,在另一个方面,所述经基因修饰的高等植物是c4植物。本发明的又一个方面是本发明的方法用于增加经基因修饰的高等植物(c3植物或c4植物)的t1和/或t2的和/或任何后续世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征的用途。本发明的又一个方面是本发明的方法用于制备经基因修饰的高等植物的用途,该经基因修饰的高等植物能够在这样的气候和/或营养条件下生长:该条件不允许野生型对照生长或野生型对照仅以低效率生长。在本发明的一些方面,c3植物属于双子叶植物纲(dicotyledons),在其他方面,属于茄目(solanales),在又一个方面,属于茄科(solanaceae),在又一个方面,属于烟草属(nicotiana),在又一个方面,c3植物是烟草植物(nicotianatabacum)。在本发明的一些方面,所公开的用途是增加经基因修饰的高等植物的t1和/或t2的和/或任何后续世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。在本发明的另一个方面,所公开的用途是制备经基因修饰的高等植物,该经基因修饰的高等植物能够在这样的气候和/或营养条件下生长:该条件不允许野生型对照生长或野生型对照以低效率生长。在本发明的另一个方面,该经基因修饰的高等植物选自:稻米、小麦、橘树、葡萄植物、咖啡植物、烟草植物、茶植物、花生植物、柠檬树、马铃薯、胡萝卜、番茄、桃树、苹果树、梨树、芒果树、大麦、燕麦、裸麦、黑小麦、干菜豆、大豆、绿豆、蚕豆、豇豆、豌豆、鹰嘴豆、木豆、兵豆、香蕉、椰子、芋头、薯蓣、甘薯、木薯、甜菜、棉花、黄麻、剑麻、芝麻、向日葵、油菜籽、红花、马唐、玉米(玉蜀黍)、苋菜、高粱、粟、甘蔗、香附、马唐、稗、四翅滨藜和/或藜科植物。在详细描述本发明之前,应当理解本公开不限于本公开的具体方面。还应当理解,本文所用的术语仅出于描述具体方面的目的,并非旨在进行限制。必须指出,如说明书和所附权利要求所用,单数形式“一个”、“一种”和“所述”包括单数和/或复数形式,除非上下文明确表明相反。还应当理解,在给出由数值界定的参数范围的情况下,该范围被认为包括这些端值。附图说明图1.莱茵衣藻co2浓缩机制(ccm)整合进高等植物,例如c3植物图1示出了不同ccm蛋白在高等植物例如c3植物中的定位。请注意cah1的“非典型”定位。cah1和cah3是指莱茵衣藻的碳酸酐酶。lcia和lcib是指莱茵衣藻的碳酸氢盐转运蛋白。图2.衣藻属碳酸酐酶和碳酸氢盐转运蛋白在烟草叶绿体(t2植株)中的表达制备生成cah1、cah3、lcia或lcib的转基因烟草。此处描述了不同ccm蛋白在高等植物(如烟草植物)的细胞隔室中的定位。缩写“bt”是指内源性烟草碳酸氢盐转运蛋白,缩写“ca”指内源性烟草碳酸酐酶。a:cah1定位于膜间隙(非典型定位)。b:cah3定位于类囊体腔(典型或天然定位)。c:lcia定位于内膜(典型或天然定位)。d:lcib定位于基质(典型或天然定位)。图3.转基因t2植物的转录物表征通过使用基因特异性引物进行rt-pcr(逆转录酶pcr)来确认rna转录物存在于t2植物中。品系1–4是指cah1、cah3、lcia或lcib的转基因品系。使用非转基因烟草作为对照(wt)。nc是rt-pcr的阴性对照。第1–4道分别指cah1或cah3转基因在四种不同cah1或cah3转基因t2品系中的积聚;而第1–2道分别指lcia或lcib转基因在lcia或lcib转基因t2品系中的积聚。图4.转基因t2植株的重组蛋白质表征重组蛋白质的存在通过免疫印迹分析进行。蛋白质的产率(单位μg/g)在右边示出。“fw”意指“鲜重”。品系1–4是指cah1、cah3、lcia或lcib的转基因品系。使用非转基因烟草作为对照(wt)。cah1、lcia和lcib通过tag54特异性抗体检测。cah3使用cah3的特异性抗体检测。第1-4道和第1-5道是指不同的转基因植株。图5.通过共聚焦显微镜观察重组蛋白质的定位图5示出了通过共聚焦显微镜观察到的所有emgfp融合蛋白在经基因修饰的高等植物细胞内的重组蛋白质的烟草叶绿体中的正确定位。图6.重组蛋白质在t2烟草叶绿体中发挥功能图6示出了在经基因修饰的高等植物如烟草的叶绿体中表达的所有重组蛋白质是有功能的。该分析通过测定碳酸酐酶(ca)活性分析进行。该测定发生在5.5周龄植物中,所有值均为平均值+标准偏差(sd);每次分析的植株数量(n=4)。生成cah1和cah3的植株显示出体外ca活性。生成lcia或lcib碳酸氢盐转运蛋白的转基因品系显示出高ca活性。lcia和lcib表达影响内源性植物ca活性。缩写“t.c.”表示“非相关转基因对照”。“wt”意指野生型对照(未经修饰的烟草植物)。图7.转基因t2品系中更高的叶绿素含量图7示出了转基因t2品系中的叶绿素含量增加。使用spad-设备(konicaminolta)测定8.5周龄烟草植物,以测试叶绿素a+b的含量。与野生型和非相关转基因对照相比,所有转基因品系具有显著(p<0.0005)较高的叶绿素含量。缩写“t.c.”表示“非相关转基因对照”。“wt”意指野生型对照(未经修饰的烟草植物)。图8.转基因品系在发育的早期的生长增强图8a:示出了ms平板中的3周龄小株(烟草)。+/-是指ms平板中存在或不存在卡那霉素选择性。与野生型对照相比(wt),在所有转基因植株中,苗的生物质、叶片数量和总叶片面积增加。缩写“t.c.”表示“非相关转基因对照”。图8b:示出了土壤中的4.5周龄植物。与野生型对照相比(wt),在所有转基因植株中,苗的生物质、叶片数量和总叶片面积也增加。图9.烟草t2转基因品系的生长增强图9a示出了具有转基因ccm蛋白的烟草植物的生长增强。每周测定5至8周龄植物的叶片面积。以平均值+/-标准偏差表示。植株的数量(n=6)。图9b示出了具有lcia的8周龄烟草植物的生长增强。可容易地观察到叶片大小较大。图10.t2转基因品系的早开花表型图10示出了t2转基因品系的早开花表型。转基因植物花比野生型和非转基因对照早1.5-2周。所有植物均为8周龄。图11.叶片大小增加图11示出了8周龄植物的叶片大小增加。图12.t2转基因品系的叶片的淀粉含量增加图12示出了与野生型相比,在所有转基因品系的光照期结束时,淀粉水平显著增加。对6周龄植物的叶片进行淀粉分析。图13.转基因植物的光合作用增强。在植物7-8周龄时进行分析。nwt=7;ncah1=8;ncah3=10;nlcia=11;nlcib=3。图14.cah1的核酸序列图15.cah3的核酸序列图16.lcia的核酸序列图17.lcib的核酸序列图18.在缺氮(氮含量少75%)的水耕栽培中生长的t4转基因品系显示出生长增强图19示出了6周龄lcia和lcib转基因烟草植物的生长增强。可容易地观察到,与野生型(wt)和非相关转基因对照(t.c.)相比,lcia和lcib植物的叶片大小较大。具体实施方式本公开涉及经基因修饰的高等植物及其后续世代,所述经基因修饰的高等植物包括“莱茵衣藻co2浓缩机制”(ccm)的至少一种基因产物(如mrna或蛋白质)的稳定或瞬时表达,其中该表达在高等植物的叶绿体的膜间隙和/或一个或多个亚细胞隔室中发生,并且其中该表达增加了经基因修饰的高等植物(t0)的和/或所述经基因修饰的高等植物的t1和/或t2世代或任何其他世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征。在一些方面,还公开了所述经基因修饰的高等植物的更快生长和更早开花。其中术语“高等植物”是指具有维管组织木质部和韧皮部的许多植物。维管植物包括所有种子植物(裸子植物和被子植物)和羊齿植物(包括蕨类、石松门和木贼)。也称为导管植物。在一个方面,该术语涵盖c3植物。在另一个方面,涵盖c4植物。其中术语“稳定表达”涉及基因持续超过数周或数月或甚至数年时间的持续表达(从而持续生成重组蛋白质和/或其他基因产物)。在一个方面,该表达在后续世代,诸如t1、t2和后续世代进行。从而,经基因修饰的高等植物的后代中携带遗传信息。基因可整合进高等植物基因组中和/或叶绿体的遗传信息(拟核)中和/或细胞或细胞器(诸如例如线粒体)中存在的任何其他dna组分中。术语“瞬时表达”是指基因在一段很短的时间期(几小时、几天、几周)内的瞬间表达(因此瞬间生成重组蛋白质和/或其他基因产物)。不受该方法的严格束缚,通常瞬时表达由导入细胞的人工质粒促进。本文的术语“经基因修饰的高等植物”涉及遗传物质已通过基因工程技术改变的高等植物。经基因修饰的高等植物可以是经基因修饰的食品的来源,并且还广泛用于科学研究以及制造商品而非食品。术语经基因修饰的高等植物还涵盖如下定义:“具有通过使用现代生物技术获得的遗传物质的新组合的高等植物”。本文的术语“c3植物”涉及单独以c3固定存活的植物。c3碳固定通过以下反应将二氧化碳和核酮糖二磷酸(rubp,5碳糖)转化为3-磷酸甘油酸:co2+rubp→(2)3-磷酸甘油酸本文的术语“c4植物”涉及使用至少一条潜在的c4碳固定途径的植物。在c4植物中,绕过了c3植物中存在的光呼吸途径。c4植物形成了将co2更有效地递送至rubisco酶的机制。它们利用了特定的叶片解剖结构,其中叶绿体不仅存在于叶片外部的叶肉细胞中,而且还存在于维管束鞘细胞中。co2不在卡尔文(calvin)循环中直接固定到rubisco,而是掺入4碳有机酸,它能够在维管束鞘细胞的叶绿体中再生co2。然后维管束鞘细胞可利用该co2通过常规的c3途径生成碳水化合物。c4途径存在很多变型:·从叶肉细胞转运的4碳酸可以是苹果酸或天冬氨酸。·从维管束鞘细胞转运回的3碳酸可以是丙酮酸或丙氨酸。·在维管束鞘细胞中催化脱羧作用的酶可不同。在玉蜀黍和甘蔗中,该酶是nadp-苹果酸酶;在粟中,该酶是nad-苹果酸酶;在大黍中,该酶是pep羧化激酶。然而,本发明对任何这些变型均有效。本文的术语“植株世代”、“后续世代”、“t0世代”、“t1世代”、“t2世代”涉及在转化事件之后植株的连续世代。亲本转化植株是t0,其直接子代是t1,t1的子代是t2植株等。后续世代可包括t3、t4、t5和/或任何后续世代。然而,特别关注的是t0至t2及其后世代的转基因表达稳定性。本文的术语“莱茵衣藻co2浓缩机制”(ccm)涉及在莱茵衣藻中鉴定的浓缩co2的机制。ccm可基于不同的生物化学机制,诸如c4光合作用和景天酸代谢(cam),基于ci的主动跨膜转运,或基于涉及通过特定细胞隔室的酸化来局部提高co2浓度的过程(giordano等人,2005)。ccm的作用是提高用于rubisco(核酮糖-1,5-二磷酸羧化酶/加氧酶)的co2浓度,该酶负责co2的最初固定。因此,ccm可用于不同co2浓缩机制的领域。因此,为清晰起见,现指出在本发明使用的术语“ccm”永远仅涉及莱茵衣藻的ccm。所提出的浓缩莱茵衣藻中的co2的模型如图1所示。图的左部表示所提出的衣藻属中的ccm模型,而图1的右部表示本发明的将co2浓缩机制的酶整合进高等植物的叶绿体的策略。莱茵衣藻的核编码的碳酸酐酶(cah1和cah3)和碳酸氢盐转运蛋白(clcia和lcib)将在植物叶绿体的不同位置表达,以提高rubisco部位的co2浓度。lcia*表示靶向信号序列的修饰,以允许重组蛋白质整合进类囊体膜。在该模型中,ccm可分为两个阶段。ccm的第一阶段涉及从环境获取无机碳,并将co2和hco3-递送至叶绿体。该部分ccm的组分将包括周质空间中的ca(碳酸酐酶)(cah1和可能地cah8)和细胞质中的ca(cah9),以及原生质膜和叶绿体被膜二者上的hco3-转运蛋白和co2通道。所提出模型的第二部分利用类囊体膜两侧的ph梯度引发叶绿体基质中hco3-水平升高。该部分ccm包括位于叶绿体基质的ca(cah6)和位于类囊体腔内的ca(cah3)以及类囊体膜上的hco3-转运蛋白。应当强调,莱茵衣藻具有严格的c3生物化学,因为与其中转运的碳以有机c4存储的c4途径不同,莱茵衣藻将无机碳特别是hco3-积聚于叶绿体基质中。此外,虽然实验表明,海洋硅藻威氏海链藻(thalassiosiraweisflogii)具有类c4途径,但相同的研究人员推断类c4途径不可能在绿藻中工作(reinfelder等人,2004)。除已显示出作为莱茵衣藻中ccm的一部分的许多ca之外,还讨论了在ccm中发挥作用的一些转运蛋白。这些转运蛋白中的两种lcia和lcib是本发明特别关注的。lcia也称为nar1.2。lcia/nar1.2首先称为亚硝酸盐转运蛋白,并且与细菌亚硝酸盐/甲酸盐转运蛋白家族具有很强的相似性。nar1.2属于由莱茵衣藻中六个nar基因组成的基因家族,意外的是,这些基因与拟南芥属(arabidopsis)或任何其他高等植物的同源性不明显。nar1.2的表达在低co2条件下诱导,并且部分在其他ccm基因表达所需的转录因子cia5的控制下(miura等人,2004)。预期nar1.2定位于叶绿体类囊体或叶绿体被膜,并且具有六个跨膜域。nar1.2在爪蟾(xenopus)卵母细胞中的功能表达显示,与对照相比,nar1.2的存在使进入卵母细胞的碳酸氢盐增加两倍(mariscal等人,2006)。本发明的一部分是,据发现与野生型对照相比,可在生成lcia或lcib碳酸氢盐转运蛋白的转基因品系中测定到更高的碳酸酐酶活性,表明lcia和lcib表达影响了内源性植物碳酸酐酶的活性。lcib与其他生物的蛋白质不具有任何显著同源性,但其预测氨基酸序列与莱茵衣藻基因组中三个基因lcic、lcid和lcie的预测氨基酸序列具有相似性。在低co2条件下lcic和lcid也上调。虽然这些观察结果指向lcib在适应低co2的作用,但预计lcib本身不可能是转运蛋白,因为它缺乏任何疏水跨膜域。因此,lcib被认为具有相当的调节作用或可以是转运ci(无机碳(ci=co2+hco3-))的复合物的一部分(van等人,2001)。术语“膜间隙”涉及线粒体或叶绿体的内膜和外膜之间的区域。膜间隙的主要功能是氧化磷酸化。这与术语“周质空间”形成对比,周质空间是革兰氏阴性菌中两个可选择性渗透屏障,即作为革兰氏阴性菌的内膜(即细胞质膜)和外膜的生物膜所界定的空间。本文的术语“叶绿体的亚细胞隔室”涉及叶绿体的不同结构实体。而在c3植物中,叶绿体通常为透镜形,直径5–8μm、厚1–3μm,与藻类中存在的叶绿体形状有很大的差异。而例如衣藻属中的叶绿体的形状则像杯子。然而,所有叶绿体具有至少三个膜系统—叶绿体外膜、叶绿体内膜和类囊体系统。作为次级内共生产物的叶绿体可具有另外的围绕这三个膜的膜。在叶绿体外膜和内膜内部的是叶绿体基质,构成大部分叶绿体体积的半凝聚样流体,其中漂浮着类囊体系统(被类囊体腔填充的类囊体膜)。因此,当本发明使用术语“叶绿体的亚细胞隔室”时,其包括至少叶绿体外膜、膜间隙、叶绿体内膜、叶绿体基质、类囊体膜和类囊体腔。本文所用的术语“内被膜”有时指叶绿体内膜。术语“典型定位”和“非典型定位”是指ccm相关蛋白,诸如例如lcia、lcib、cah1和cah3在源细胞或细胞器内的定位。“典型定位”或“天然定位”涵盖ccm相关蛋白的所有定位,类似于在天然来源如莱茵衣藻的叶绿体隔室中的定位。这与“非典型”定位形成对比,在“非典型”定位中ccm相关蛋白定位于叶绿体和/或细胞的隔室,此处天然不存在。然而,莱茵衣藻和高等植物细胞的细胞隔室有时所指之处是不同的。因此,应根据本发明利用下表来定义“相似的隔室”:表1:隔室的比较莱茵衣藻的隔室高等植物的叶绿体的隔室外(细胞)膜质膜/原生质膜周质空间--内(细胞)膜质膜/原生质膜细胞质细胞质叶绿体外膜叶绿体外膜膜间隙膜间隙叶绿体内膜/内被膜叶绿体内膜/内被膜基质叶绿体基质类囊体膜类囊体膜淀粉核--例如:cah1定位于莱茵衣藻的周质空间,因此定位于经基因修饰的植物的叶绿体膜间隙将视为“非典型”。相比之下,cah3、lcia、lcib的定位被视为“典型”或“天然”定位,因为它们分别定位于淀粉核/类囊体腔、叶绿体内被膜或叶绿体基质。术语“增加所述经基因修饰的c3植物和/或c4植物的t1和/或t2世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征”涉及与未经修饰的野生型对照植物相比,任何所述特征的增加。而增加可在至少1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、15%、20%、30%、40%、50%、60%、70%、80%、90%、100%和至多5%、至多10%、至多15%、至多20%、至多25%、至多30%、至多40%、至多50%、至多60%、至多70%、至多80%、至多90%、至多95%、至多99%、至多100%、至多110%、至多120%、至多130%、至多140%、至多150%、至多160%、至多170%、至多180%、至多190%、至多200%、至多250%、至多260%、至多270%、至多280%、至多290%和/或至多300%之间。然而,在一些方面,与未经修饰的野生型对照植物相比,增加可为至少两倍(即100%)、三倍(即200%)、四倍(300%)、五倍(400%)、六倍(500%)和至多七倍(600%)、八倍(700%)、九倍(800%)和十倍(900%)。如无另外说明,本文的测定在植物的营养期结束时进行。在本发明的一些方面,在cah1表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2世代和/或后续世代的光合速率提高至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%、至少10%;叶绿素水平提高至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%、至少10%;生物质增加至少10%、至少20%、至少30%、至少40%、至少50%。在本发明的一些方面,在cah1表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2世代和/或后续世代的光合速率提高在1%-5%、2%-6%、3%-7%、4%-8%、5%-9%、6%-10%、7%-11%、8%-12%、9%-13%、10%-14%之间;叶绿素水平提高在1%-5%、2%-6%、3%-7%、4%-8%、5%-9%、6%-10%、7%-11%、8%-12%、9%-13%、10%-14%之间;生物质增加在10%-20%、20%-30%、30%-40%、40%-50%、50%-60%、60%-70%、70%-80%、80%-90%、90%-100%之间。在本发明的又一个方面,与野生型对照相比,在经基因修饰的高等植物的叶绿体的膜间隙中所述碳酸酐酶cah1的表达提高了所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代的光合速率至少8%、叶绿素水平至少7%和/或生物质至少22%。这些植物的叶片数量也增加。在本发明的一个方面,在cah1t2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+22%·鲜重:+26%·干重:+29%在本发明的又一个方面,在cah1t2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+142%/+22%·叶绿素含量:+15%/+13%·叶片数量:+21%/+24%·鲜重:nd/+26%·干重:nd/+29在本发明的另一个方面,在cah3表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2的和/或任何后续世代的光合速率提高至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%、至少10%,叶绿素水平提高至少5%、至少10%、至少15%、至少20%、至少25%、至少30%,生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。在本发明的一些方面,在cah3表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2世代和/或后续世代的光合速率提高在1%-5%、2%-6%、3%-7%、4%-8%、5%-9%、6%-10%、7%-11%、8%-12%、9%-13%、10%-14%之间;叶绿素水平提高在1%-5%、2%-6%、3%-7%、4%-8%、5%-9%、6%-10%、7%-11%、8%-12%、9%-13%、10%-14%之间;生物质增加在10%-20%、20%-30%、30%-40%、40%-50%、50%-60%、60%-70%、70%-80%、80%-90%、90%-100%之间。在本发明的又一个方面,与野生型对照相比,在经基因修饰的高等植物的叶绿体的类囊体腔中所述碳酸酐酶cah3的表达,使得在所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代中,与野生型对照相比,光合速率提高至少8%、叶绿素水平提高至少10%,气孔大小增加最多50%和/或生物质增加至少31%。在本发明的一个方面,在cah3t2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+24%·鲜重:+34%·干重:+31%在本发明的又一个方面,在cah3t2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+78%/+24%·叶绿素含量:+11%/+10%·叶片数量:+26%/+64%·鲜重:nd/+34%·干重:nd/+31%在本发明的又一个方面在lcia表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2的和/或任何后续世代的光合速率提高至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%、至少10%,叶绿素水平提高至少5%、至少10%、至少15%、至少20%、至少25%、至少30%,和/或生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。在本发明的一些方面,在lcia表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2世代和/或后续世代的光合速率提高在1%-5%、2%-6%、3%-7%、4%-8%、5%-9%、6%-10%、7%-11%、8%-12%、9%-13%、10%-14%之间;叶绿素水平提高在1%-5%、2%-6%、3%-7%、4%-8%、5%-9%、6%-10%、7%-11%、8%-12%、9%-13%、10%-14%之间;生物质增加在10%-20%、20%-30%、30%-40%、40%-50%、50%-60%、60%-70%、70%-80%、80%-90%、90%-100%之间。在本发明的又一个方面,在营养期结束时,与野生型对照相比,在经基因修饰的高等植物的叶绿体的内被膜中所述碳酸氢盐转运蛋白lcia的表达提高了所述经转基因修饰的高等植物的t1和/或t2世代或任何后续世代的光合速率至少8%、叶绿素水平至少15%和/或生物质至少33%。在本发明的一个方面,在lciat2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+33%·鲜重:+54%·干重:+41%在本发明的又一个方面,在lciat2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+93%/+33%·叶绿素含量:+11%/+16%·叶片数量:+17%/+19%·鲜重:nd/+54%·干重:nd/+41%在本发明的一个方面,在lcib表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2的和/或任何后续世代的生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。在本发明的一个方面,在lcib表达的情况下,与未经修饰的野生型对照植物相比,t1和/或t2世代和/或后续世代的生物质增加在10%-20%、20%-30%、30%-40%、40%-50%、50%-60%、60%-70%、70%-80%、80%-90%、90%-100%之间。在本发明的一个方面,在营养期结束时,叶片面积增加23%,鲜重增加30%,干重增加28%,叶片数量增加24%,叶绿素增加15%。在本发明的又一个方面,与野生型对照相比,在经基因修饰的高等植物的叶绿体的基质中所述碳酸氢盐转运蛋白lcib的表达增加了所述经基因修饰的高等植物的t1和/或t2世代或任何后续世代的生物质。在本发明的一个方面,在lcibt2植物的营养期结束时,与野生型对照相比,定义生物质的三个参数增加:·叶片面积:+23%·鲜重:+30%·干重:+28%在本发明的又一个方面,在lcibt2植物的第3周和营养期结束时,与野生型对照相比,百分比增加如下(在第3周的百分比增加/在营养期结束时的百分比增加;nd:不确定):·叶片面积:+78%/+23%·叶绿素含量:+10%/+15%·叶片数量:+27%/+24%·鲜重:nd/+30%·干重:nd/+28%在另一个方面,在lcib表达的经基因修饰的高等植物的t2世代的第6周,叶片面积增加78%,叶片数量增加24%,叶绿素增加10%。在一个方面,在cah3表达的情况下,叶片面积增加约80%,在lcia表达的情况下,增加约154%,在lcib表达的情况下,增加约171%,或在cah1表达的情况下,甚至增加约260%。在另一个方面,在cah1表达的情况下,叶绿素水平提高约6.7%,或在lcia表达的情况下约11.5%。在又一个方面,与野生型植物相比,对于cah1,叶片面积增加22%(p<0.005),对于cah3,24%(p<0.005),对于lcia,33%(p<0.005)以及对于lcib,23%(p<0.005);对于cah1,鲜重(fw,“鲜重”)增加26%(p<0.005),对于cah3,34%(p<0.005),对于lcia,54%(p<0.0005)以及对于lcib,30%(p<0.005);对于cah1,干重(dw)增加29%(p<0.005),对于cah3,31%(p<0.005),对于lcia,41%(p<0.005)以及对于lcib,28%(p<0.005)(参见实例部分的表3)。例如,可观察到以下增加:例如,叶片数量(2-6)和表面积(最多33%)增加;鲜重(26-54%)和干重(28-54%)增加;在cah3和lcia转基因品系中,气孔大小增加(最多51%),气孔数量和密度减少;在cah3植物中,很多茎在早期发育;在lcia品系中,叶片更光滑、更大。技术人员将清楚,本发明的经基因修饰的高等植物可包括cah1、cah3、lcia或lcib的单个转基因,或ccm机制的任何其他成员(如cah6),或这些转基因在一个植株中的许多组合。如cah1和lcia、cah3和lcib、cah1和cah3、lcia和lcib等。如技术人员可容易地从图2、结果和实验推断的那样,经基因修饰的高等植物可根据个别需要定制。因为那些组合具有另外的作用,具有单个转基因和仅稍微改善的特征的植株可通过添加ccm机制的另外转基因进一步改善。因此,本发明还涵盖所涉及的ccm物类(cah1、cah3、lcia、lcib)的两者、三者或全部四者的组合,或甚至与ccm机制的其他成员(如cah6)的组合。本文涉及的植物特征增加的计算可通过下式进行:从上式可显而易见,只要在经基因修饰的植物和未经修饰的野生型对照之间测试相同特征,并且将相同的测试系统用于测试,百分比增加与所测定特征的单位无关。可增加的特定特征包括经基因修饰的高等植物本身(t0)或所述经基因修饰的高等植物的t1或所述经基因修饰的高等植物的t2世代和/或任何其他世代的光合速率、光合碳固定、叶绿素水平和/或生物质。如上所述,本文的特征测定通常在植物的营养期结束时进行。在脱离此规则之处,将明确说明。例如,“光合速率”可通过co2的摄取、o2的生成、碳水化合物的生成和/或干重(如收集植物、干燥至恒重并称重)的增加测定。例如,“光合碳固定”可通过痕量nah14co3的摄取测定。然后通过过滤颗粒物和通过测定其放射性或通过酸化和鼓泡技术(其测定颗粒和溶解产物二者)确定该组合。例如,“叶绿素水平”可通过分光光度法、高效液相色谱法(hplc)和/或荧光测定法测定。例如,“生物质”可通过叶片面积、全植株重量(干重,如收集植物、干燥至恒重并称重)和/或叶片数量测定。技术人员理解,光合速率增加也可导致生物质增加和/或碳固定增加,反之亦然。不同特征的测定也可根据实例部分所述的测试进行(不受这些测试的束缚)。合适的测试为例如:种子重量和数量、代谢物分析、碳水化合物含量诸如淀粉含量或糖含量、新鲜和干燥生物质等。在一个方面,莱茵衣藻ccm的任何成员可用于增加植物特征。然而,在另一个方面,ccm的基因产物选自莱茵衣藻的碳酸氢盐转运蛋白lcia、碳酸氢盐转运蛋白lcib、碳酸酐酶cah1和/或碳酸酐酶cah3。在一些方面,ccm的基因产物是在经基因修饰的高等植物的叶绿体的膜间隙稳定或瞬时表达的莱茵衣藻的碳酸酐酶cah1。在一些方面,与相同龄期、相同栽培品种和在相同条件下生长的未经修饰的野生型对照植物相比,叶绿体的膜间隙中的此类表达使叶绿素水平提高至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少9%或至少10%;和/或使生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。在一些方面,ccm的基因产物是在经基因修饰的高等植物的叶绿体的类囊体腔稳定或瞬时表达的莱茵衣藻的碳酸酐酶cah3。在一些方面,与相同龄期、相同栽培品种和在相同条件下生长的未经修饰的野生型对照植物相比,叶绿体的膜间隙中的此类表达使光合速率增加至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少7%、至少9%或至少10%;使叶绿素水平提高至少10%、至少11%、至少12%、至少13%、至少14%、至少15%、至少16%、至少17%、至少18%、至少19%或至少20%;和/或使生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。在一些方面,与野生型对照相比,经基因修饰的高等植物生长得更快和/或营养期更短。在一些方面,ccm的基因产物是在经基因修饰的高等植物的叶绿体的膜间隙稳定或瞬时表达的莱茵衣藻的碳酸氢盐转运蛋白lcia。在一些方面,与相同龄期、相同栽培品种和在相同条件下生长的未经修饰的野生型对照植物相比,叶绿体的内被膜中的此类表达使光合速率增加至少1%、至少2%、至少3%、至少4%、至少5%、至少6%、至少7%、至少8%、至少7%、至少9%或至少10%;使叶绿素水平提高至少10%、至少11%、至少12%、至少13%、至少14%、至少15%、至少16%、至少17%、至少18%、至少19%或至少20%;和/或使生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。在一些方面,ccm的基因产物是在经基因修饰的高等植物的叶绿体的膜间隙稳定或瞬时表达的莱茵衣藻的碳酸氢盐转运蛋白lcib。在一些方面,与相同龄期、相同栽培品种和在相同条件下生长的未经修饰的野生型对照植物相比,叶绿体的基质中的此类表达使生物质增加至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少100%。本发明还涉及制备本文所述的经基因修饰的高等植物的方法。此外,本发明还涉及使用经基因修饰的高等植物,来提高与野生型对照相比,所述经基因修饰的高等植物所生成的天然和/或转基因产物的产率的用途。此类“天然产物”可以是花、种子、果实、植物、坚果、油、蛋白质、肽、核酸、代谢物诸如醇、抗氧化剂、有机酸、多元醇、氨基酸、维生素和/或核苷酸、木材、纤维、叶片或所述经修饰的有机植物增量生成的并且可从该植物收集的任何其他物质。通常此类天然产物是植物的具有商业用途部分。技术人员理解,本发明可应用于c3以及c4植物二者。c3植物的例子包括稻米、小麦、橘树、葡萄植物、咖啡植物、烟草植物、茶植物、花生植物、柠檬树、马铃薯、胡萝卜、番茄植物、桃树、苹果树、梨树、芒果树和大麦;从此类植物收集的天然产物可以是例如稻粒、小麦粒、橘果、葡萄、咖啡豆、烟草叶片、茶叶和茶芽、花生、柠檬、马铃薯、胡萝卜、番茄、桃、苹果、梨、芒果或大麦粒等。另外的例子包括燕麦、裸麦、黑小麦、干菜豆、大豆、绿豆、蚕豆、豇豆、豌豆、鹰嘴豆、木豆、兵豆、香蕉、椰子、芋头、薯蓣、甘薯、木薯、甜菜、棉花、黄麻、剑麻、芝麻、向日葵、油菜籽和红花。另外的例子包括谷类、豆类、果类、根部和块茎、油料作物、纤维作物和乔木。就c4植物而言,例子包括来源于马唐、玉米(玉蜀黍)、苋菜、高粱、粟、甘蔗、香附、马唐、稗、四翅滨藜和藜科植物的任何产物。然而,此类天然产物还包括例如来源于稻米的支链淀粉、来源于马铃薯的淀粉、来源于烟草的尼古丁、来源于咖啡或茶植物的咖啡因等。在一些方面,可从根据本发明的经基因修饰的高等植物制备生物柴油和/或生物乙醇。“转基因产物”包括非野生型植物天然生成,而是由于通过除基因修饰(其使植物特征增加)之外的转基因和/或生物技术方法将包含一个或多个编码基因产物的基因的核酸引入植物而生成的任何产物。此类转基因产物包括核酸、蛋白质、肽和/或通常非该植物生成的任何其他代谢物诸如醇、抗氧化剂、有机酸、多元醇、氨基酸、维生素和/或核苷酸。在一些方面,转基因产物可包括药物活性物质,也称为活性成分(ai)、活性药物成分(api)或有时也成为植物药物。作为活性成分(ai),包括具有生物活性的任何物质。此类成分可用于动物或人类药物,或可以例如是杀虫剂、杀真菌剂、抗生素等等。例如,在一个方面,疫苗、酶、抗体和/或激素由经修饰的植物生成。本发明还涉及增加经基因修饰的高等植物的t1和/或t2的和/或任何后续世代的选自光合速率、光合碳固定、叶绿素水平和/或生物质的一个或多个特征的方法的用途。此类用途可包括增加经基因修饰的高等植物的抗性或耐久性能力,以增加在其天然环境中的可持续性。在其他方面,该方法用于增加经基因修饰的高等植物的抗性或耐久性能力,以允许该植物在野生型对照植物难以生存的条件下生长。在一些方面,涵盖了该方法用于制备经基因修饰的高等植物的用途,该经基因修饰的高等植物能够在这样的气候和/或营养条件下生长:该条件不允许野生型对照生长。在其他方面,经基因修饰的高等植物属于双子叶植物纲,属于茄目,属于茄科和/或属于烟草属。在一个方面,经基因修饰的高等植物是烟草植物(nicotianatabacum)。术语“核酸分子”或“核酸”旨在表示cdna、基因组dna、合成dna或rna、肽核酸(pna)或lna来源的任何单链或双链核酸分子。术语“突变”是指单个或多个核苷酸三联体的置换或替换、一个或多个密码子的插入或缺失、不同基因之间的同源或异源重组、另外的编码序列融合至编码序列的任一端、或插入另外的编码序列或这些方法的任何组合,它们产生编码所需蛋白质的多核酸序列。因此,术语“突变”还指一个或多个上述变化修饰的多核酸序列编码的多肽序列的所有变化。本公开还涉及包含本公开的核苷酸分子的载体。术语“载体”包括能够转运与其连接的另外的核酸的核酸分子。一种类型的载体是“质粒”,它是指可连接另外的dna片段的环状双链dna环。另一种类型的载体是其中另外的dna片段可连接至病毒基因组的病毒载体。某些载体能够在它们被引入的宿主细胞中自主复制(如,具有细菌复制起点和附加体哺乳动物载体的细菌载体)。其他载体(如,非附加体哺乳动物载体)可在引入宿主细胞时整合进宿主细胞的基因组,从而与宿主基因组一起复制。此外,某些载体能够引导它们可操作地连接的基因的表达。此类载体在本文中称为“重组表达载体”(或简称“表达载体”)。一般来讲,重组dna技术所用的表达载体通常是质粒的形式。在本说明书中,“质粒”和“载体”可互换使用,因为质粒是最常用形式的载体。然而,本公开旨在包括这些具有等同功能的其他形式的表达载体,诸如病毒载体(如,复制缺陷逆转录病毒、腺病毒和腺相关病毒)。一般来讲,本领域的技术人员能够很好地构建载体和设计方案,用于重组基因表达。载体构建体和表达系统是本领域熟知的,并且可适于结合本文提供的重组融合多肽的使用。例如,转基因植物制备是已知的,并且构建体生成和植物制备可根据本领域已知的技术改进。在一些方面,植物中的瞬时表达系统是所期望的。一般来讲,本领域已知的标准方法可用于根据本发明培养或栽培植物、植物细胞和/或植物组织(如克隆植物、克隆植物细胞、克隆根、克隆根系、芽、发芽幼苗、植物等),分别用于生物质制备或重组多肽和代谢物制备。在本公开的一些方面,分离例如用于疫苗产物、酶、抗体、激素等等的重组多肽是所期望的。当根据本公开从表达蛋白质的植物组织或它们的一部分如根、根细胞、植物、植物细胞制备该蛋白质时,本领域已知的方法可用于任何部分或完整的分离的植物体。当希望从表达产物的一些或所有植物细胞或组织分离该表达产物时,可利用任何可用的纯化技术。本领域的普通技术人员熟悉一系列广泛的分级和分离工序(参见例如scopes等人,proteinpurificationprinciplesandpractice,第3版,janson等人,1993,proteinpurificationprincipleshighresolutionmethods,andinventions,wiley-vch,1998,springer-verlag,ny,1993和roe,proteinpurificationtechniques,oxforduniversitypress,2001,每个参考文献以引用的方式并入本文)。本领域的技术人员将认识到,获得所需的重组融合多肽产物的方法是提取。可提取植物体(如,根、叶片等),以从残留的生物质移除所需的产品,从而提高产物的浓缩和纯度。可在缓冲液中提取植物。例如,可以按重量计一比一的比率将植物体转移至一定量的冰冷水(已使用如磷酸盐缓冲液缓冲)中。可根据需要加入蛋白酶抑制剂。可通过剧烈混合或研磨破坏植物体,同时悬浮于缓冲液中,并通过过滤或离心移除所提取的生物质。可通过另外的步骤进一步纯化溶液中获得的产物,或通过冷冻干燥或沉淀转化为干粉。可通过在压机中压制植物或根,或在它们通过间隔紧密的辊时压碎来进行提取。根据本领域熟知的方法收集和加工来源于压碎的植物或根的流体。通过压制提取允许以更浓缩的形式释放产物。在一些方面,可通过色谱法进一步纯化多肽,该色谱法包括但不限于阴离子交换色谱(q柱)或超滤。包含his标签的多肽可根据标准方法使用镍交换色谱纯化。在一些方面,所制备的蛋白质或多肽不从植物组织分离,而是以活植物(如,发芽幼苗)的背景提供。在一些方面,当植物为可食用时,直接提供包含表达蛋白或多肽的植物组织用于消耗。因此,本公开提供包含表达蛋白或多肽的可食用的秧苗生物质(如可食用的发芽幼苗)。另外的加工步骤诸如纯化步骤可通过许多本领域已知的工序进行,该工序包括但不限于色谱(如,离子交换、亲和、疏水、色谱聚焦和体积排阻)、电泳工序(如,制备性等电聚焦)、差别溶解(如,硫酸铵沉淀)或提取。此外,所关注的分离和纯化多肽可进一步加工,诸如例如配制成组合物,如药物组合物。在本发明的语境中,术语“工艺”可与术语“方法”或“工序”互换使用,特别指根据本公开的用于纯化重组制备的融合蛋白的过程步骤的任何组合和/或顺序。在本发明的语境中,术语“多步骤”工艺用于描述用于纯化蛋白质和/或肽的工艺,其包括一系列昂贵的和/或费力的和/或耗时的和/或技术复杂的纯化步骤。作为一般原则,与本公开的方法描述的具有一个或仅具有几个步骤工艺相比,多步骤工艺包括用于宿主细胞的初始破坏和用于从宿主细胞蛋白和污染物第一次粗分离异源蛋白的至少两个明显分开的过程步骤。通常,这些步骤在至少一个纯化步骤之后再次进行。虽然多步骤工艺和本公开涉及的工艺还可包括多个另外的纯化步骤,但如果需要更高纯度的所关注的蛋白质,这不是本公开所述的工艺必须的。本公开的生成实际上不含污染性宿主细胞蛋白的基本上纯的重组融合蛋白的工艺可优选地以基本上单个过程步骤进行。方法和实例以下提供了本公开的实例、材料和方法。应当理解这些实例仅为了说明的目的,并且不应解释为以任何方式限制本公开。本文引述的所有公开、专利和专利发明全文据此以引用方式并入,用于所有目的。实例1:莱茵衣藻cah1、cah3、lcia和lcibcdna的合成莱茵衣藻的cah1(genbank:d90206.1)、cah3(genbank:u73856)、lcia(genbank:ay612639.1)和lcib(genbank:ab168093.1)的编码序列通过化学dna合成获得(genscript,piscataway,usa)。在合成之前,每个cdna是根据烟草密码子使用而进行密码子优化的,以使表达产量最大化。此外,使用泛型算法同时为一大组竞争参数而优化合成cdna,诸如mrna二级结构、隐蔽剪接位点、密码子和基序重复和均一gc含量。合成cah3、lcia和lcibcdna包含莱茵衣藻的天然转运肽序列,使重组cah3、lcia和lcib蛋白分别靶向类囊体腔、叶绿体内膜和基质。合成cah1cdna包含拟南芥(arabidopsisthaliana)的tic22转运肽序列,使重组cah1蛋白靶向莱茵衣藻cah1的非典型定位内膜间隙。每个合成构建体包含cah1、cah3、lcia或lcibcdna的上游和tag54编码序列下游的欧芹(petrosilium)查耳酮合酶基因的5’-非翻译区。通过限制性酶位点ecori和xbai将四个合成cdna单独转移至载体puc18。实例2:cah1、cah3、lcia和lcib分别亚克隆至植物表达载体为评估莱茵衣藻cah1、cah3、lcia或lcib分别在烟草cv.petithavanasr1植株中对碳酸酐酶活性和生物质生成的体内作用,将每个编码cah1、cah3、lcia或lcib的cdna单独插入单独或融合至emgfpcdna的植物表达载体ptra。转基因表达由具有重复增强子区的花椰菜(cauliflower)花叶病毒35s启动子驱动。ptra质粒包含烟草rb7基因(gi3522871)和ppcv002的nptii盒的支架结合区(konzetshell,1986),用于在含卡那霉素的琼脂平板上选择转基因植物。指定的最终构建体为:·ptra-35s-attic22-tp:cah1,ptra-35s-attic22-tp:cah1-gfp,·ptra-35s-crtl-tp:cah3,ptra-35s-crtl-tp:cah3-gfp·ptra-35s-crim-tp:lcia,ptra-35s-crim-tp:lcia-gfp·ptra-35s-crs-tp:lcib,ptra-35s-crs-tp:lcib-gfp以下公开了植物表达载体的例子ptra-35s-attic22-tp:cah1的序列(8882bp):实例3:烟草植物转化和再生使用genepulserii电穿孔系统(biorad,hercules,ca,usa)根据制造商的说明将植物表达载体引入根癌农杆菌(agrobacteriumtumefaciens)gv3101(pmp90rk,gmr,kmr,rifr)细胞。4–5周龄野生型烟草(n.tabacumcv.petithavanasr1)植株的叶盘通过携带上述双元载体的重组根癌农杆菌的感染来转化(dietze等人,1995)。愈伤组织在含50mg/l卡那霉素的murashige-skoog培养基上生长,并且植株从抗性愈伤组织再生。将可能的转基因植物转移至温室中的土壤,然后自交产生t1世代。在温室的13l反应釜的de73标准土壤中栽培转基因烟草植物,使用16h自然光光周期和22℃白天/20℃白天/黑夜温度。使用pcr筛选最多25个存在整合进基因组的转基因的转基因t0品系(即cah1-1至cah1-25;cah1-gfp-1至cah1-gfp25;cah3-1至cah3-25;cah3-gfp-1至cah3-gfp25;lcia-1至lcia-25;lcia-gfp-1至lcia-gfp-25;lcib-1至lcib-25和lcib-gfp-1至lcib-gfp-25)。pcr分析确认所有的分析转基因品系中存在转基因。转录物和重组蛋白质的积聚分别使用rt-pcr和免疫印迹分析展示。对于免疫印迹分析,将5周龄烟草植物的完全展开的上部叶片在液氮下碾磨成细粉,使用两体积提取缓冲液(50mmtris-hclph8、100mmnacl、10mm二硫苏糖醇(dtt)、5mm乙二胺四乙酸(edta)和0.1%(v/v)tween-20)提取可溶性总蛋白(tsp)。在4℃下以8500×g离心提取物20min,并用于免疫印迹分析。使用兔抗tag54单克隆抗体(rab-tag54;200ng/ml)检测重组cah1、cah3、lcia和lcib蛋白。此外,还使用兔抗cah3多克隆抗体(在1×pbs中1:2000稀释)(antibody-online,aachen,germany)检测cah3蛋白。重组蛋白质的带强度使用aida软件(raytest,straubenhardt,germany)定量,标准物为已知浓度的细菌亲和纯化的scfv。虽然全部转基因t0品系均生成lcib重组蛋白质,但仅25%和75%的再生品系显示出可检测水平的cah1或lcia蛋白,预期分子大小分别为51.6kda和36.5kda。分析的t0植物中未检测到重组cah3蛋白生成,很明显是由于低积聚水平和/或检测抗体的低灵敏度。重组cah1-gfp、cah3-gfp、lcia-gfp和lcib-gfp蛋白在所有分析的转基因t0品系中均生成。选择在孟德尔(mendelian)分离后具有最高水平重组蛋白质积聚的转基因cah1、lcia和lcib品系来建立t1世代。由于cah3积聚是基于免疫印迹检测水平的,我们选择在孟德尔(mendelian)分离后的三个pcr阳性t0品系来建立cah3t1世代。通过pcr确认每个构建体的全部二十五个分析的t1植株中cah1、cah3、lcia和lcib插入序列的存在。此外,75%的cah3转基因t1品系中cah3转录物的存在也通过rt-pcr分析确认。重组cah1、cah3、lcia和lcib的积聚类似于亲代t0品系。t2世代中插入序列存在和重组蛋白质积聚也通过rt-pcr(图3)和免疫印迹(图4)确认。虽然t2世代中cah3的积聚增加至1μg/g鲜重,但重组cah1、lcia和lcib的积聚类似于亲代品系(cah1:5μg/g鲜重;lcia:100μg/g鲜重;lcib40μg/g鲜重)。实例4:碳酸酐酶活性的分析为研究cah1和cah3碳酸酐酶在植物中是否有活性,积聚最高水平重组蛋白质的6周龄转基因t2植株中的总碳酸酐酶活性根据wilbur和anderson(1948)描述的电位测定方法确定。将20μl系列稀释(50-375个单位)的牛红细胞的碳酸酐酶溶液(sigmaaldrich,germany)或来自转基因和野生型对照的植物提取物加入1.48ml20nmveronal缓冲液(727mmnacl、9.12mm二乙基巴比妥钠、15.63mm5,5’-二乙基巴比妥酸)(ph8.3),在反应室内维持于4℃。通过加入0.5ml冰冷的co2饱和水引发反应,并测定ph从8.3降至8.0所需的时间。wa活性单位定义如下:wa单位=tc/t-1,tc和t是分别在不存在和存在酶溶液的情况下ph下降所需的时间。由于内源性碳酸酐酶的活性,野生型烟草植物和非相关转基因对照(生成人类抗体m12的转基因t2烟草植物)在相同条件下的分析显示出显著的背景活性(图6)。从cah1和cah3植物制备的提取物产生的碳酸酐酶活性比野生型植物显著更高(分别为208%和188%)(图6),表明工程化cah1和cah3具有碳酸酐酶活性。此外,在分别生成重组lcia或lcib的转基因植物的叶片提取物中也观察到碳酸酐酶活性的显著增加(183%和179%),表明莱茵衣藻碳酸氢盐转运蛋白的组成型表达使得内源性烟草碳酸酐酶的活性增加。因此,我们推断工程化重组cah1、cah3、lcia和lcib在转基因t1和t2植物中发挥作用。实例5:转基因烟草植物的光合活性重组cah1、cah3、lcia和lcib对转基因烟草植物的光合性能的作用通过监测气体交换参数确定。使用li-6400系统(li-cor,badhomburg,germany)对7-8周龄t1和t2烟草植物的完全展开的上部叶片进行气体交换测定。使用以下参数:光子通量密度1,000mmolm–2s–1、反应室温度26℃、流速150mmols–1、相对湿度60–70%。co2补偿点(γ)通过测定400、300、200、100、80、60和40ppmco2下的光合速率来确定。表观co2补偿(γ)从a/ci曲线通过回归分析在曲线的线性范围内推导。相同植物的测定在4h光照后的后续两个不同的天进行。如图13所示,在环境条件(400ppmco2、21%o2、24℃)下,所选择的cah1、cah3、lcia和lcib产生品系的表观光合速率(a)分别提高12.4%、14%、9%和10%,暗示转基因品系中光合速率的提高反映了在ribisco附近co2的可获取性更佳。转基因cah3和lcia产生植物的特征也是表观co2补偿点显著(p<0.01)降低(分别为3%和2%),表明光合速率更高。cah1和lcib产生t1和t2品系还显示出co2补偿点降低的趋势。此外,在cah3和lcia产生品系中气孔导度分别显著增加27%和33%,表明进入植物细胞的co2水平很高。在图13中,示出了转基因植物中的光合作用增强。在植物7-8周龄时进行分析。nwt=7;ncah1=8;ncah3=10;nlcia=11;nlcib=3。实例6:转基因植物中的叶片淀粉分析为确定莱茵衣藻碳酸酐酶和碳酸氢盐转运蛋白对初级碳代谢的作用,我们评估了转基因t2品系和野生型植物积聚光合作用终产物的能力。为确定淀粉水平,在两个不同的时间点,即在光照期开始时和结束时收集50mg叶片材料。在液氮中碾磨冰冻的叶片材料并重悬于80%(v/v)乙醇中。提取物在80℃下混合10min,并以4000g离心20min。将沉淀物分别重悬于80%(v/v)和50%(v/v)乙醇中,然后在80℃下混合并如上所述离心。用90%(v/v)乙醇洗涤所得的沉淀物,重悬于400μl0.2koh中,在95℃下温育1h。最后,将样品与70μl1m乙酸混合,通过酶学方法测定淀粉含量。在清晨,所有品系的叶片包含低水平的淀粉,但在一天结束时观察到与野生型对照相比,转基因品系中淀粉水平显著增加(在lcia产生t2品系中,增加最多2.67倍,p<;0.005)。在所有转基因品系中测定的淀粉增加较多,反映了co2同化的改善,这不仅足以支持生长,而且在叶片中积聚储存化合物,然后固定该储存化合物以提供夜间生长所需的碳。实例7:积聚莱茵衣藻cah1、cah3、lcia或lcib的植物的表型评估每周通过测定叶片数量、植株高度和叶片面积来监测叶绿体中积聚cah1、cah3、lcia或lcib的t1和t2世代转基因植物的生长,所述叶片面积根据下式计算:叶片面积(cm2)=3.73×(长度×宽度/100)+0.011×(长度×宽度/100)2。与非转基因对照植物和非相关转基因对照相比,产生一种重组蛋白质的转基因烟草t2品系在完全生长期显示出叶片面积显著增加(表2)。此外,转基因植物在发育早期具有更多的叶片(多于2-6片),生长更快(在5周龄,叶片面积:cah1:增加260%、cah3:增加69%、lcia:增加154%和lcib:增加171%)。更快速生长使得种子的产生更早(与野生型和非相关转基因对照相比,1.5-2周)。表2*:p<0.005;**:p<0.0005;***:p<0.00005;n.s.无显著性当植物达到8周龄时,监测t2世代的干重和鲜重。如表3所示,在营养期结束时,所有转基因品系显示出鲜重和干重显著增加26%至54%的范围。表3注:1:在营养期结束时*:p<0.005;**:p<0.0005实例8:氮饥饿研究使野生型纯合lcia和lcib转基因t4品系在包含100%氮的ms培养基上水耕栽培生长4周,并在低氮下再生长2周(氮比正常ms培养基少75%)。与野生型植株相比,6周龄lcia和lcib转基因植物(在氮饥饿之后2周)显示出苗的鲜重和干重显著增加(图18;表4)。有趣的是,我们观察到根的鲜重和干重也显著增加(lcia分别为:32%、17%;lcib分别为:56%和45%;表4)。在这个时间点,测定硝酸还原酶活性,硝酸还原酶是硝酸盐同化最重要的酶(可用于绿色植物在土壤中生长的主要氮形式)。与野生型植物相比,转基因lcia和lcib植物显示出硝酸还原酶活性显著增加(lcia:48%和lcib:45%)(图19)。苗或根鲜重和干重,或野生型和非相关转基因对照之间的硝酸还原酶活性无显著差异,暗示所观察到的显著变化仅仅是由于lcia和lcib表达。这些数据表明,烟草叶绿体中lcia或lcib的组成型表达可能提高了lcia和lcib转基因植物中氮同化的效率。表4*:p<0.05图18.在缺氮(氮含量少75%)的水耕栽培中生长的t4转基因品系显示出生长增强。图18示出了6周龄lcia和lcib转基因烟草植物的生长增强。可容易地观察到,与野生型(wt)和非相关转基因对照(t.c.)相比,lcia和lcib植物的叶片大小更大。图19.在氮饥饿条件下,水耕栽培生长的植物的硝酸还原酶活性以微摩尔no2/克鲜重(fw)/分钟表示。数据为平均值±sd(n=5)。*p<0.05参考文献所有引述的参考文献的内容,包括文献、公布的专利和公布的专利发明,如在整个本发明中引述的那样,据此明确地以引用的方式并入。dietzeetal.,1995,agrobacterium-mediatedtransformationofpotato(solanumtuberosum).inipotrykus,gspangenberg,eds,genetransfertoplants,springerlaboratorymanual.springer,berlin,pp24-29giordano,m.,j.beardall,andj.a.raven.2005.co2concentratingmechanismsinalgae:mechanisms,environmentalmodulation,andevolution.annu.rev.plantbiol.56:99-131.reinfelder,j.r.,a.m.l.kraiepiel,andf.m.m.morel.2004.theroleofthec4pathwayincarbonaccumulationandfixationinamarinediatom.plantphysiol.135:2106-2111jansonetal,1993,proteinpurificationprincipleshighresolutionmethods,andinventions,wiley-vch,1998,springer-verlag,ny,1993konzetshell,1986thepromoteroftl-dnagene5controlsthetissue-specificexpressionofchimaericgenescarriedbyanoveltypeofagrobacteriumbinaryvector.molgengenet204:383-396mariscal,v.,p.moulin,m.orsel,a.j.miller,e.fernández,anda.galván.2006.differentialregulationofthechlamydomonasnar1genefamilybycarbonandnitrogen.protist157:421-433miura,k.,t.yamano,s.yoshioka,t.kohinata,y.inoue,f.taniguchi,e.asamizu,y.nakamura,s.tabata,k.t.yamato,k.ohyama,andh.fukuzawa.2004.expressionprofiling-basedidentificationofco2-responsivegenesregulatedbyccm1controllingacarbon-concentratingmechanisminchlamydomonasreinhardtii.plantphysiol.135:1595-1607roe,proteinpurificationtechniques,oxforduniversitypress,2nded.,2001scopesetal,proteinpurificationprinciplesandpractice,springer,3rded.van,k.,y.wang,y.nakamura,andm.h.spalding.2001.insertionalmutantsofchlamydomonasreinhardtiithatrequireelevatedco2forsurvival.plantphysiol.127:607-614wilburandanderson(1948)electrometricandcolorimetricdeterminationofcarbonicanhydrase.jbiolchem176147-154序列表<110>弗劳恩霍夫应用研究促进协会<120>光合作用增加的经基因修饰的高等植物<130>fra-pa15-ep<160>5<170>patentinversion3.5<210>1<211>1254<212>dna<213>chlamydomonasreinhardtii<400>1atggagtcaagcgttaagccaaacccattcctctcattttcttcttttattcatcaccaa60tgtactagattcagtagcgatttgagtgctagaatcgaagatacaaagaggtttgctgag120actcttgcaacaagaaggttttctttgcctactccacctccattcgcttccgtttccatg180gggtgtatctataagttcggtactagcccagattccaaagcaacagtgtctggagatcat240tgggatcacggacttaatggtgaaaactgggagggaaaagatggagctggtaatgcatgg300gtgtgcaagacaggtagaaagcaatcaccaattaatgttccacaatatcaggtgttggat360ggaaagggttcaaaaattgctaatggtcttcaaactcagtggagttaccctgatttgatg420tctaacggaacatcagttcaagttattaataacggacatactatacaagttcagtggaca480tacaactacgctggtcatgcaactattgctatcccagcaatgcacaatcaaacaaacagg540attgttgatgtgcttgaaatgagacctaatgatgctgcagatagggttactgctgtgcca600acacagtttcacttccattcaactagtgaacatcttttggcaggaaagatctatccttta660gagctccatatagttcaccaagtgactgaaaagcttgaggcttgtaaaggaggttgcttt720agtgttacaggaatccttttccagttggataatggtccagataacgaactccttgaacct780atattcgctaatatgccaagtagagaaggtactttcagcaacttacctgcaggaactaca840attaagctcggagagcttttgccatctgatagggattatgttacttacgaaggatctctt900actacacctccatgttctgagggtttactctggcatgtgatgacacaacctcagagaatt960tcttttggacagtggaatagatacagattagctgttggtctcaaagaatgcaactcaact1020gagacagctgcagatgctggacatcaccatcaccatagaaggcttttgcacaatcatgca1080cacttagaagaggttcctgctgcaactagcgaaccaaagcattattttagaagagttatg1140cttgctgagtctgcaaatccagatgcatacacatgcaaagctgttgcattcggtcaaaac1200tttaggaaccctcagtatgctaacggaagaactatcaagctcgctaggtatcat1254<210>2<211>930<212>dna<213>chlamydomonasreinhardtii<400>2atgaggtccgcagtgctccaaaggggtcaggcaagaagagtttcttgtagagtgagggct60gatggatctggtgttgattcacttccaagcacttccgcttcttcaagtgcaagacctttg120atcgatagaaggcaacttttgactggagctgcagcttcagttataacatttgtgggatgt180ccatgccctctttgtaagccaggtgaagctaaagcagctgcatggaattatggagaggtt240gcaggtccacctacttggaagggagtgtgcgctacaggaaagagacagtccccaattaat300atccctcttaacacatctgcacctaaggttgatgctgaaatgggagagtttgatttcgct360tacggttcattcgaaaaatgcgatgttttgaatactggacatggtacaatgcaagttgtg420aattttccagctggaaacttagcattcattggtaacatggaacttgaattactccagttt480catttccacgcacctagcgaacatgctatggatggaagaagatatgctatggaggcacat540cttgttcacaagaataagagtactggacacttagcagtgctcggtattatgctcccagga600ggtgctgatcaaaaccctgctctttctacagcattggaagttgctccagaggtgcctttg660gcaaagaaaccatcacctaagggtatcaatccagttatgcttttgcctaagaaaagtaaa720gctggaactagaccatttgtgcactaccctggtagtttgactacaccaccttgtagcgag780ggagttgattggtttgtgttcatgcaaccaattaaggttcctgattctcagatcctcgat840ttcatgaggttcgtgggagataacaaaacatatgctactaacacaagaccattacaactc900ctcaactcaagactcgtggaatatgaactc930<210>3<211>1008<212>dna<213>chlamydomonasreinhardtii<400>3atgcagactacaatgacaagaccatgtttagcacagccagttcttagatccagggtgttg60agatctcctatgagggttgtggctgcatccgctccaactgcagttactacagttgtgaca120tctaatggtaacggtaatggtcatttccaagctgcaactacacctgttccacctacacca180gctcctgttgcagtgtcagctccagtgagggctgttagtgtgttgactccacctcaggtt240tatgaaaacgcaattaatgtgggagcatacaaagctggtcttactcctttggctacattt300gttcaaggaatacaggctggtgcatatatcgcattcggagcttttcttgcaatttcagtt360ggtggtaacatacctggagtggctgcagctaatccaggtcttgctaagcttttgttcgca420ttagtttttccagtgggactcagtatggttactaactgtggagctgaacttttcactggt480aatacaatgatgcttacatgcgctttgattgagaagaaagcaacttggggtcaattactc540aaaaactggtctgtttcatacttcggaaattttgttggttcaatagctatggtggcagct600gttgtggcaactggatgtttaactacaaacacactccctgtgcagatggctactttaaag660gcaaatctcggttttacagaagttcttagtaggtccatactttgtaattggttggtttgt720tgcgctgtgtggtctgcatcagcagctacatctttgcctggaagaatccttgctttgtgg780ccatgcatcactgctttcgttgcaattggtcttgaacattcagttgctaacatgtttgtg840attcctttaggaatgatgctcggtgcagaggttacttggagtcaatttttctttaataac900cttatcccagttactttgggaaatacaatcgctggtgtgcttatgatggctatagcatac960agtatctccttcggatcattgggaaagtctgcaaaaccagctacagca1008<210>4<211>1347<212>dna<213>chlamydomonasreinhardtii<400>4atgttcgcactcagttcaagacagacagcaaggtccgcatgtagagcatcttgtccatgc60gcttcatgcaggggtgtggctagtgcaccagttagagcaacatatgctgcaaggcctgtt120aagaaatctgctgcatcagttgtggttaaggctcaagctgcatctactgctgtggcacca180gttgagaatggtgctgcacctgctgttgcacataagagaacatttgctcagaggcatagt240gaattgattaaacactttccttccaccatgggcgttgatgatttcatgggtagagtggag300gttgctcttgcaggatttggtttcactggagataatacaattgcaatgactaacctctgt360agggatgaagtgacacaagttcttaaggataaaatcgaggctatttttggatcttcattc420aatactaacggtcttggtggtgtgttgacatgtggagttactggtatgaaagctggattg480tctcattcaccagtgtgcaatggtggaagagaaagatatgttttctttgcatttcctcac540attgctattaattctgaaggagagatgggtgcactttcaagacctggtaggcctaagcaa600agttgtgcttgcggagcacttttggctattcttaatgctttcaaagtggatggtgttgaa660aagtcatgtaaagtgccaggagttcacgatccacttgatcctgagttaacaatactccaa720cagagattggctagaagggtgaggtatgaaaagttagatgtttccaaacttgatttgcca780ggtttgacttctgttgctgaaagaactataacagatgatcttgaatatttgatagagaag840gcagttgatccagctgttgcagattacgctgtgataacaggagttcaaatccataattgg900ggtaaagaacttagtgcatccggagatgcttctattgagtttgttgctccagcaaagtgc960tatacagtggttaacggtcttaaaacttacatagatttgccacaggttcctgctttatca1020cctagacaaatccagacaatggctcaagcaagtcttaatggtttcgaaccaaagcacatc1080cagcctggaatgaggggtagtgtgatttccgaagttcctttagagtatctcgttactaaa1140ttaggaggttcccaactcatggaggatggaaactcttacgcaccagtttttgctagttcc1200gattcattcgaatggcctacatggcagagtagaattaggcttgataataacccaaacaga1260ttactctctgtggagagggatgctaacgcacctactatggaatcaccagagcctgttcac1320cctagttttgaggcacctaagaataag1347<210>5<211>8882<212>dna<213>artificialsequence<220><223>vectorconstructptra-35s-attic22-tp:cah1<400>5aattccgcccctagaaatatttgcgactcttctggcatgtaatatttcgttaaatatgaa60gtgctccatttttattaactttaaataattggttgtacgatcactttcttatcaagcgtt120actaaaatgcgtcaatctctttgttcttccatattcatatgtcaaaatctatcaaaattc180ttatatatctttttcgaatttgaagtgaaatttcgataatttaaaattaaatagaacata240tcattatttaggtgtcatattgatttttatacttaattactaaatttggttaactttgaa300agtgtacatcaacgaaaaattagtcaaacgactaaaataaataaatatcatgtgttatta360agaaaattctcctataagaatattttaatagatcatatgtttgtaaaaaaaattaatttt420tactaacacatatatttacttatcaaaaatttgacaaagtaagattaaaataatattcat480ctaacaaaaaaaaaccagaaaatgctgaaaacccggcaaaaccgaaccaatccaaaccga540tatagttggtttggtttgattttgatataaaccgaaccaactcggtccatttgcacccct600aatcataatagctttaatatttcaagatattattaagttaacgttgtcaatatcctggaa660attttgcaaaatgaatcaagcctatatggctgtaatatgaatttaaaagcagctcgatgt720ggtggtaatatgtaatttacttgattctaaaaaaatatcccaagtattaataatttctgc780taggaagaaggttagctacgatttacagcaaagccagaatacaaagaaccataaagtgat840tgaagctcgaaatatacgaaggaacaaatatttttaaaaaaatacgcaatgacttggaac900aaaagaaagtgatatattttttgttcttaaacaagcatcccctctaaagaatggcagttt960tcctttgcatgtaactattatgctcccttcgttacaaaaattttggactactattgggcg1020cggggcgcgcccggtccaaagaccagagggctattgagacttttcaacaaagggtaatat1080cgggaaacctcctcggattccattgcccagctatctgtcacttcatcgaaaggacagtag1140aaaaggaagatggcttctacaaatgccatcattgcgataaaggaaaggctatcgttcaag1200atgcctctaccgacagtggtcccaaagatggacccccacccacgaggaacatcgtggaaa1260aagaagacgttccaaccacgtcttcaaagcaagtggattgatgtgatacatggtggagca1320cgacactctcgtctactccaagaatatcaaagatacagtctcagaagaccagagggctat1380tgagacttttcaacaaagggtaatatcgggaaacctcctcggattccattgcccagctat1440ctgtcacttcatcgaaaggacagtagaaaaggaagatggcttctacaaatgccatcattg1500cgataaaggaaaggctatcgttcaagatgcctctaccgacagtggtcccaaagatggacc1560cccacccacgaggaacatcgtggaaaaagaagacgttccaaccacgtcttcaaagcaagt1620ggattgatgtgatatctccactgacgtaagggatgacgcacaatcccactatccttcgca1680agacccttcctctatataaggaagttcatttcatttggagaggacacgctgaattcacta1740cacagattagattcatcgaaagattcatcaagaagaagaaaactatggagtcaagcgtta1800agccaaacccattcctctcattttcttcttttattcatcaccaatgtactagattcagta1860gcgatttgagtgctagaatcgaagatacaaagaggtttgctgagactcttgcaacaagaa1920ggttttctttgcctactccacctccattcgcttccgtttccatggggtgtatctataagt1980tcggtactagcccagattccaaagcaacagtgtctggagatcattgggatcacggactta2040atggtgaaaactgggagggaaaagatggagctggtaatgcatgggtgtgcaagacaggta2100gaaagcaatcaccaattaatgttccacaatatcaggtgttggatggaaagggttcaaaaa2160ttgctaatggtcttcaaactcagtggagttaccctgatttgatgtctaacggaacatcag2220ttcaagttattaataacggacatactatacaagttcagtggacatacaactacgctggtc2280atgcaactattgctatcccagcaatgcacaatcaaacaaacaggattgttgatgtgcttg2340aaatgagacctaatgatgctgcagatagggttactgctgtgccaacacagtttcacttcc2400attcaactagtgaacatcttttggcaggaaagatctatcctttagagctccatatagttc2460accaagtgactgaaaagcttgaggcttgtaaaggaggttgctttagtgttacaggaatcc2520ttttccagttggataatggtccagataacgaactccttgaacctatattcgctaatatgc2580caagtagagaaggtactttcagcaacttacctgcaggaactacaattaagctcggagagc2640ttttgccatctgatagggattatgttacttacgaaggatctcttactacacctccatgtt2700ctgagggtttactctggcatgtgatgacacaacctcagagaatttcttttggacagtgga2760atagatacagattagctgttggtctcaaagaatgcaactcaactgagacagctgcagatg2820ctggacatcaccatcaccatagaaggcttttgcacaatcatgcacacttagaagaggttc2880ctgctgcaactagcgaaccaaagcattattttagaagagttatgcttgctgagtctgcaa2940atccagatgcatacacatgcaaagctgttgcattcggtcaaaactttaggaaccctcagt3000atgctaacggaagaactatcaagctcgctaggtatcataaacatctgaaagattgggaac3060atctggaagaatttgcggccgctcatcaccatcaccatcactagctcgagctcggatcct3120ctagagtccgcaaaaatcaccagtctctctctacaaatctatctctctctatttttctcc3180agaataatgtgtgagtagttcccagataagggaattagggttcttatagggtttcgctca3240tgtgttgagcatataagaaacccttagtatgtatttgtatttgtaaaatacttctatcaa3300taaaatttctaattcctaaaaccaaaatccagtgaccgggcggccggccgcccctagaaa3360tatttgcgactcttctggcatgtaatatttcgttaaatatgaagtgctccatttttatta3420actttaaataattggttgtacgatcactttcttatcaagcgttactaaaatgcgtcaatc3480tctttgttcttccatattcatatgtcaaaatctatcaaaattcttatatatctttttcga3540atttgaagtgaaatttcgataatttaaaattaaatagaacatatcattatttaggtgtca3600tattgatttttatacttaattactaaatttggttaactttgaaagtgtacatcaacgaaa3660aattagtcaaacgactaaaataaataaatatcatgtgttattaagaaaattctcctataa3720gaatattttaatagatcatatgtttgtaaaaaaaattaatttttactaacacatatattt3780acttatcaaaaatttgacaaagtaagattaaaataatattcatctaacaaaaaaaaacca3840gaaaatgctgaaaacccggcaaaaccgaaccaatccaaaccgatatagttggtttggttt3900gattttgatataaaccgaaccaactcggtccatttgcacccctaatcataatagctttaa3960tatttcaagatattattaagttaacgttgtcaatatcctggaaattttgcaaaatgaatc4020aagcctatatggctgtaatatgaatttaaaagcagctcgatgtggtggtaatatgtaatt4080tacttgattctaaaaaaatatcccaagtattaataatttctgctaggaagaaggttagct4140acgatttacagcaaagccagaatacaaagaaccataaagtgattgaagctcgaaatatac4200gaaggaacaaatatttttaaaaaaatacgcaatgacttggaacaaaagaaagtgatatat4260tttttgttcttaaacaagcatcccctctaaagaatggcagttttcctttgcatgtaacta4320ttatgctcccttcgttacaaaaattttggactactattgggcgggtggagggggatcaga4380ttgtcgtttcccgccttcagtttaaactatcagtgtttgacaggatatattggcgggtaa4440acctaagagaaaagagcgtttattagaataatcggatatttaaaagggcgtgaaaaggtt4500tatccgttcgtccatttgtatgtgtacatcaccgacgagcaaggcaagaccgagcgcctt4560tccgacgctcaccgggctggttgccctcgccgctgggctggcggccgtctatggccctgc4620aaacgcgccagaaacgccgtcgaagccgtgtgcgagacaccggccgccggcgttgtggat4680acctcgcggaaaacttggccctcactgacagatgaggggcggacgttgacacttgagggg4740ccgactcacccggcgcggcgttgacagatgaggggcaggctcgatttcggccggcgacgt4800ggagctggccagcctcgcaaatcggcgaaaacgcctgattttacgcgagtttcccacaga4860tgatgtggacaagcctggggataagtgccctgcggtattgacacttgaggggcgcgacta4920ctgacagatgaggggcgcgatccttgacacttgaggggcagagtgctgacagatgagggg4980cgcacctattgacatttgaggggctgtccacaggcagaaaatccagcatttgcaagggtt5040tccgcccgtttttcggccaccgctaacctgtcttttaacctgcttttaaaccaatattta5100taaaccttgtttttaaccagggctgcgccctgtgcgcgtgaccgcgcacgccgaaggggg5160gtgcccccccttctcgaaccctcccggcccgctaacgcgggcctcccatccccccagggg5220ctgcgcccctcggccgcgaacggcctcaccccaaaaatggcagcgctggcagtccttgcc5280attgccgggatcggggcagtaacgggatgggcgatcagcccgacaagctacccctatttg5340tttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaat5400gcttcaataatattgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttat5460tcccttttttgcggcattttgccttcctgtttttgctcacccagaaacgctggtgaaagt5520aaaagatgctgaagatcagttgggtgcacgagtgggttacatcgaactggatctcaacag5580cggtaagatccttgagagttttcgccccgaagaacgttttccaatgatgagcacttttaa5640agttctgctatgtggcgcggtattatcccgtattgacgccgggcaagagcaactcggtcg5700ccgcatacactattctcagaatgacttggttgagtactcaccagtcacagaaaagcatct5760tacggatggcatgacagtaagagaattatgcagtgctgccataaccatgagtgataacac5820tgcggccaacttacttctgacaacgatcggaggaccgaaggagctaaccgcttttttgca5880caacatgggggatcatgtaactcgccttgatcgttgggaaccggagctgaatgaagccat5940accaaacgacgagcgtgacaccacgatgcctgtagcaatggcaacaacgttgcgcaaact6000attaactggcgaactacttactctagcttcccggcaacaattaatagactggatggaggc6060ggataaagttgcaggaccacttctgcgctcggcccttccggctggctggtttattgctga6120taaatctggagccggtgagcgtgggtctcgcggtatcattgcagcactggggccagatgg6180taagccctcccgtatcgtagttatctacacgacggggagtcaggcaactatggatgaacg6240aaatagacagatcgctgagataggtgcctcactgattaagcattggtaactgtcagacca6300agtttactcatatatactttagattgatttaaaacttcatttttaatttaaaaggatcta6360ggtgaagatcctttttgataatctcatgaccaaaatcccttaacgtgagttttcgttcca6420ctgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcg6480cgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccgga6540tcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagataccaaa6600tactgtccttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcc6660tacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtg6720tcttaccgggttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaac6780ggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactgagatacct6840acagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatcc6900ggtaagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctg6960gtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatg7020ctcgtcaggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcct7080ggccttttgctggccttttgctcacatggactctagctagaggatcacaggcagcaacgc7140tctgtcatcgttacaatcaacatgctaccctccgcgagatcatccgtgtttcaaacccgg7200cagcttagttgccgttcttccgaatagcatcggtaacatgagcaaagtctgccgccttac7260aacggctctcccgctgacgccgtcccggactgatgggctgcctgtatcgagtggtgattt7320tgtgccgagctgccggtcggggagctgttggctggctggtggcaggatatattgtggtgt7380aaacaaattgacgcttagacaacttaataacacattgcggacgtttttaatgatcgaata7440ctaacgtctctaccagatatcagcttgcatgccggtcgatctagtaacatagatgacacc7500gcgcgcgataatttatcctagtttgcgcgctatattttgttttctatcgcgtattaaatg7560tataattgcgggactctaatcataaaaacccatctcataaataacgtcatgcattacatg7620ttaattattacatgcttaacgtaattcaacagaaattatatgataatcatcgcaagaccg7680gcaacaggattcaatcttaagaaactttattgccaaatgtttgaacgatctgcttgactc7740tagctagagtccgaaccccagagtcccgctcagaagaactcgtcaagaaggcgatagaag7800gcgatgcgctgcgaatcgggagcggcgataccgtaaagcacgaggaagcggtcagcccat7860tcgccgccaagctcttcagcaatatcacgggtagccaacgctatgtcctgatagcggtcc7920gccacacccagccggccacagtcgatgaatccagaaaagcggccattttccaccatgata7980ttcggcaagcaggcatcgccctgggtcacgacgagatcctcgccgtcgggcatccgcgcc8040ttgagcctggcgaacagttcggctggcgcgagcccctgatgctcttcgtccagatcatcc8100tgatcgacaagaccggcttccatccgagtacgtcctcgctcgatgcgatgtttcgcttgg8160tggtcgaatgggcaggtagccggatcaagcgtatgcagccgccgcattgcatcagccatg8220atggatactttctcggcaggagcaaggtgagatgacaggagatcctgccccggcacttcg8280cccaatagcagccagtcccttcccgcttcagtgacaacgtcgagcacagctgcgcaagga8340acgcccgtcgtggccagccacgatagccgcgctgcctcgtcttggagttcattcagggca8400ccggacaggtcggtcttgacaaaaagaaccgggcgcccctgcgctgacagccggaacacg8460gcggcatcagagcagccgattgtctgttgtgcccagtcatagccgaatagcctctccacc8520caagcggccggagaacctgcgtgcaatccatcttgttcaatcatgcctcgatcgagttga8580gagtgaatatgagactctaattggataccgaggggaatttatggaacgtcagtggagcat8640ttttgacaagaaatatttgctagctgatagtgaccttaggcgacttttgaacgcgcaata8700atggtttctgacgtatgtgcttagctcattaaactccagaaacccgcggctgagtggctc8760cttcaacgttgcggttctgtcagttccaaacgtaaaacggcttgtcccgcgtcatcggcg8820ggggtcataacgtgactcccttaattctccgctcatgatcgatatccattgaagagcaag8880ct8882当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1