白锈病抗性菊属植物的制作方法
本发明涉及白锈病抗性菊属(chrysanthemum)植物以及其种子、植物部分、植物细胞和子代。本发明还涉及用于鉴定白锈病抗性菊属植物的手段且特别是分子标志物。
菊花(chrysanthemum),也被称为菊花(chrysant(h)),是菊科(asteraceae)菊属的有花植物。该植物原产于亚洲和欧洲东北部,并且包括大量的园艺品种和栽培品种。
几个菊花的属中的经济上重要的菊花(florist’schrysanthemum)在过去被分类于菊属(dendranthema)中。然而目前,花店(florist)的(菊花被重新分类为野菊(chrysanthemumindicum),恢复了这些菊花在菊属中的位置。
天然存在的菊属的种是草本多年生植物。这些菊属的种展示出交替排列的叶片,分为具有锯齿状或有时具有光滑边缘的小叶片。早在公元前15世纪,菊花首先在中国作为有花草药被栽培,并且截止1630年,已经记载了超过500个栽培品种。
目前栽培的菊花与它们野生的亲缘植物相比展示更明显且富有美感的花期。头状花序以多种形式存在,并且可以是雏菊样的或装饰性的,如绒球形(pompon)或纽扣形(button)。该属包含为园艺目的开发的许多杂交种和数以千计的栽培品种。除了传统的黄色之外,其他颜色是可得的,诸如白色、紫色和红色。最重要的杂交种是chrysanthemumxmorifolium,也被称为chrysanthemumxgrandiflorum,其主要源自野菊(chrysanthemumindicum)。
菊花可以分成两个基本的组,花园耐寒型(gardenhardy)和展示型。花园菊是能够在大部分北纬区域越冬的多年生植物。展示品种通常不能够越冬。花园菊可以根据它们在极少(如果有的话)机械辅助诸如用桩支撑(staking)的情况下产生大量小花朵(bloom)的能力和能够耐受风和雨的能力来定义。展示品种通常需要用桩支撑,在相对干燥、凉爽的环境下越冬,并且有时需要增加夜间光照。
白锈病是由担子菌门(basidiomycota)引起的植物中的病害。担子菌门形成真菌样真核微生物的独特的系统发育谱系。它们是丝状、微小、吸附型生物体,其通过有性和无性二者繁殖。担子菌纲(basidiomycetes)兼有腐生性和病原性生活方式,并且包括一些最臭名昭著的植物病原体,引起毁灭性的病害诸如马铃薯、番茄和宽范围的观赏植物的白绢病(southernblight)和小麦的秆锈病(stemrust)。担子菌纲最著名的是产生大子实体,诸如蕈类、马勃(puffball)和檐状菌(bracket),并且是木材和落叶层腐烂中的重要生物体。
在菊花中,白锈病是一种通常由病原性担子菌或真菌堀柄锈菌(pucciniahoriana)引起的病害。菊花的具体症状包括在叶片的上表面上的白锈病斑点。这些斑点最初颜色是浅绿色至黄色,并且直径多达5mm,但随着组织变得坏死,可变成棕色。在叶片的下表面,斑点发展成粉色或白色小隆起物,其随着冬孢子产生而变得突出。该病害通常由温室菊花的受感染的插条和植株,包括切花携带。
直至1963年,堀柄锈菌都局限于中国和日本。然而,此后它在感染的进口的插条上快速传播,并且现在是欧洲的苗圃中的令人担忧且严重的病害。大量致病型堀柄锈菌(p.horiana)是已知的,并且已经显示了致病型堀柄锈菌在接种在不同菊花栽培品种上之后毒力的巨大差异(debacker,2012)。显示在2006年收集于荷兰的堀柄锈菌致病型nl1是毒力最强的一个。
预防性喷洒杀真菌剂是有效的,但昂贵。当气候特别适合白锈病时,即使预防性喷洒也不够有效,并且易感品种非常有可能被感染。被发现有用的活性成分包括氧化萎锈灵(oxycarboxin)、嗪氨灵(triforine)、麦锈灵(benodanil)、三唑酮(triadimefon)、苄氯三唑醇(diclobutrazol)、联苯三唑醇(bitertanol)和丙环唑(propiconazole)。已经建议将蜡蚧轮枝菌(verticilliumlecanii)用于温室菊花的生物防治。
考虑到白锈病对菊花栽培的相当大的损害,本领域中存在对提供新的遗传抗性来源的需求,即,本领域存在对向菊属植物提供持续的白锈病抗性的新的抗性基因的需求。
除了其他目的之外,本发明的一个目的是满足本领域的上述需求。
根据本发明,本发明通过提供如所附权利要求中概述的植物、植物部分、种子和手段来满足该目的。
具体地,根据第一方面,除了其他目的之外,本发明的该目的通过提供属于菊属的植物来满足的,所述植物抗白锈病且所述植物在其基因组中包含至少一个提供白锈病抗性的基因组区域、或基因或等位基因,所述至少一个提供白锈病抗性的基因组区域、或基因或等位基因与以至少一个拷贝包含于所述抗性植物的基因组中并由seqidno.3表示的核酸序列遗传连锁。考虑到属于菊属的植物的六倍体性质,seqidno.3优选地以至少一个拷贝,诸如2、3、4、5或6个拷贝/抗性植物,存在于抗性植物的基因组中。
在本发明的上下文中,术语“基因组区域(genomicregion)”或“基因组区域(genomicregions)”表示编码一种或更多种提供白锈病抗性的基因诸如一种基因的不同的核的序列。
序列seqidno.3与提供白锈病抗性的基因组区域或基因遗传连锁,或换句话说,seqidno.3是指示提供白锈病抗性的基因组区域或基因的存在的分子标志物。seqidno.3和本文提供的其他序列可以通过将包含白锈病抗性菊属植物(诸如本文鉴定的保藏物(deposits))的基因组dna的样品经受通过用限制酶mse1和ecor1的限制性消化,任选地结合使用基于本文提供的序列开发的引物对的核酸扩增获得。
尽管检测seqidno.3的存在足以确定菊属植物是否抗白锈病,抗性可以另外地通过病害测定诸如以下概述的病害测定来确认。
病害测定可以在使用塑料盖的125cm长×80cm宽×35cm高的密闭的塑料容器内的插条上或小植株上进行。将白锈病感染的接种物植株置于容器内(36株接种物植株/容器,均匀地分布在265个插条中)。可以使用源自堀柄锈菌致病型nl1的分离株;(原始nl1致病型获自荷兰瓦格宁根植物疾病服务机构(theplantenziektenkundigedienst,wageningen,nl))。
由于白锈病要求高的相对湿度和叶片上的水膜,小植株或插条、塑料容器的内侧和盖子优选地用去离子水雾化结合塑料容器底部的湿布。准备实验环境之后,将槽(aquaria)置于18℃的生长室持续4天,用白色塑料覆盖以产生黑暗、潮湿环境。每天使容器通风以确保孢子在整个容器中良好分散。在去除白色塑料之后,用水银灯和son-t灯的组合照射容器18小时/天(6000lux)。
通常,病害症状可以在感染后21至28天评价。根据1(感染)至9(未感染)的量表对植物进行评分,其中1-3的评分指示植物为易感的,4-6指示植物为中度抗性的且7-9指示植物为抗性的。
优选地,本发明的植物在其基因组中还包含另外的提供白锈病抗性的基因组区域、或基因,该另外的提供白锈病抗性的基因组区域、或基因与以至少一个拷贝包含于抗性植物的基因组中并由seqidno.2或seqidno.1表示的核酸序列遗传连锁。考虑到属于菊属的植物的六倍体性质,seqidno.2和seqidno.1优选地以至少一个拷贝,诸如2、3、4、5或6个拷贝/抗性植物,彼此独立地存在于抗性植物的基因组中。
根据特别优选的实施方案,本发明的植物在其基因组中还包含另外的提供白锈病抗性的基因组区域或基因,该另外的提供白锈病抗性的基因组区域或基因与以至少一个拷贝包含于所述抗性植物的基因组中并分别由seqidno.1和seqidno.2表示的核酸序列遗传连锁。考虑到属于菊属的植物的六倍体性质,seqidno.2和seqidno.1优选地以至少1个拷贝,诸如2、3、4、5或6个拷贝/抗性植物,彼此独立地存在于抗性植物的基因组中。
本发明人鉴定了另外的基因组序列,该另外的基因组序列虽然与seqidno.3相似(但不相同),并且存在于白锈病抗性基因组区域、基因座或基因中,但不负责赋予白锈病抗性。因此,与seqidno.3相似(但不相同)的基因组序列被称为易感性等位基因或易感性基因组序列。因此,根据一个优选的实施方案,本发明的植物包含被包含于抗性植物的基因组中的选自由seqidno.4、seqidno.5、seqidno.6、seqidno.7、seqidno.8和seqidno.9组成的组的这些相似(但不相同)的核酸序列的五个拷贝或更少拷贝/植株。考虑到属于菊属的植物的六倍体性质,seqidno.4至seqidno.9优选地以4个拷贝或更少,诸如3、2、1或0个拷贝彼此独立地存在于抗性植物的基因组中。在后一种情况,本发明的植物包含seqidno.3的6个拷贝。
本发明的植物优选地是切花菊(cutchrysanthemum)植物或盆栽菊(potchrysanthemum)植物。切花菊植物和盆栽菊植物在本领域中还分别被称为切花和盆栽植物。有时,还提及花园菊,其为盆栽菊植物的花园检验版本(agardenproofversionofapotchrysanthemumplant)。
根据本发明的优选的实施方案,本发明的属于菊属植物选自由以下组成的组:chrysanthemumxmorifolium;chrysanthemumxgrandiflorum;chrysanthemumxrubellum;chrysanthemumabolinii;chrysanthemumachillaeal.;chrysanthemumalabasicum;chrysanthemumbrachyanthum;花环菊(chrysanthemumcarinatum);chrysanthemumchalchingolicum;白花除虫菊(chrysanthemumcinerariifolium);红花除虫菊(chrysanthemumcoccineum);宽叶菊(chrysanthemumcoreanum);茼蒿(chrysanthemumcoronarium);chrysanthemumdecaisneanum;chrysanthemumdelavayanum;异色菊(chrysanthemumdichrum);chrysanthemumfastigiatum;蓬蒿菊(chrysanthemumfrutescens);chrysanthemumgracile;chrysanthemumgrubovii;chrysanthemumhoraimontanum;chrysanthemumhypoleucum;野菊(chrysanthemumindicuml.);chrysanthemumjunnanicum;chrysanthemumkinokuniense;chrysanthemumkokanicum;chrysanthemumkonoanum;crysanthemummajus;chrysanthemummarginatum;chrysanthemummawei;chrysanthemummaximuml.;chrysanthemummiyatojimense;杭白菊(chrysanthemummorifolium);chrysanthemummultifidum;chrysanthemumnitidum;小叶菊(chrysanthemumparvifolium);chrysanthemumprzewalskii;chrysanthemumpurpureiflorum;chrysanthemumramosum;菱叶菊(chrysanthemumrhombifolium);chrysanthemumroborowskii;南茼蒿(chrysanthemumsegetum);chrysanthemumshihchuanum;chrysanthemumshimotomaii;chrysanthemumtrilobatum;chrysanthemumtripinnatisectum;毛华菊(chrysanthemumvestitum);chrysanthemumvulgare(l.);chrysanthemumyoshinyanthemum;和紫花毛山菊(chrysanthemumzawadskii)。
根据本发明的另一个优选的实施方案,引起本发明的白锈病的致病病原体是堀柄锈菌。堀柄锈菌是一种单主寄生锈菌(anautoeciousrust)。双细胞冬孢子在原位萌发以产生单细胞担孢子,其在气流中分散。已知没有其他孢子。显示高湿度和水膜是冬孢子和担孢子二者萌发所必需的。冬孢子能够在其成熟时马上萌发;担孢子的萌发和释放在4℃与23℃之间发生,并且,在17℃的最佳温度,在3小时内观察到担孢子的释放。担孢子可以在宽的温度范围内萌发,并且,在17℃-24℃,叶片的任一侧可以在2小时内被穿透。因此,仅5小时的潮湿就足以建立新的感染。在叶片内,大量透明的细胞间菌丝通过细胞内吸器产生。潜伏期通常为7-10天,但短期的高温(超过30℃)可以使潜伏期延长至8周。
感染后,在叶片的上表面形成浅绿色至黄色斑点,直径多达5mm。随着衰老,这些斑点的中心变成棕色并坏死。在对应的下表面发现凸起的、浅黄褐色或粉色的蜡状小隆起物(冬孢子堆(telia))。随着上表面上的斑点变得凹陷,这些小隆起物变得非常突出并且当产生担孢子时变得发白。有时,在叶片上表面发现冬孢子堆。被严重攻击的叶片枯萎、垂下茎并逐渐地完全干枯。
根据又另一个优选的实施方案,本发明涉及包含源自于2017年5月31日以编号ncimb42762保藏的菊属植物的至少一个提供白锈病抗性的基因组区域或基因的植物。以编号ncimb42762保藏的菊属植物可以通过英国阿伯丁ab219ya巴克斯本克莱斯通村弗格森大厦的国家工业、食品和海洋微生物保藏中心(ncimb)(thenationalcollectionofindustrial,foodandmarinebacteria(ncimb),fergusonbuilding,craibstoneestate,bucksburn,aberdeenab219ya,unitedkingdom)获得。
根据仍另一个优选的实施方案,本发明的植物还包含源自于2016年3月14日以编号ncimb42455保藏的菊属植物的至少一个提供白锈病抗性的基因组区域、或基因或等位基因,或者本发明的植物还包含源自以编号ncimb42455保藏的菊属植物的至少两个提供白锈病抗性的基因组区域、或基因或等位基因。换句话说,本发明的植物优选地堆叠抗白锈病的抗性基因,即包含一个源自保藏物ncimb42762的与seqidno.3遗传连锁的基因组区域、或基因或等位基因,以及一个或两个源自ncimb42455的分别与seqno.1或seqno.2遗传连锁的基因组区域或基因。
根据一个最优选的实施方案,本发明涉及一种抗白锈病病原体堀柄锈菌的植物chrysanthemumxmorifolium。
考虑到本领域中白锈病抗性遗传来源诸如本发明的植物的重要性,根据第二方面,本发明涉及本发明的植物的种子、植物部分或植物细胞。本发明的种子、植物部分或植物细胞在其基因组中包含seqidno.3以及优选地另外的seqidno.2或seqidno.1、最优选地seqidno.2和seqidno.1,并且因此能够提供抗白锈病或者被栽培成抗白锈病并且特别是由堀柄锈菌感染引起的白锈病的植物。
根据第三方面,本发明还涉及本发明的菊属植物的子代。本发明的植物的子代可以通过确定seqidno.3、并且优选另外的seqidno.2或seqidno.1,最优选地seqidno.2和seqidno.1在其基因组中的存在来容易地鉴定。
根据第四方面,本发明涉及seqidno.3用于鉴定白锈病抗性菊属植物的用途。基于这些seqidno.的用于鉴定此类植物的合适的方法是本领域通常已知的,诸如基于基因组dna的核酸扩增和随后的可视化扩增片段的方法,但还可以设想其他技术诸如基于杂交的技术。
本发明将在下文展示的实例中被进一步详细描述。在这些实例中,参考附图,其中:
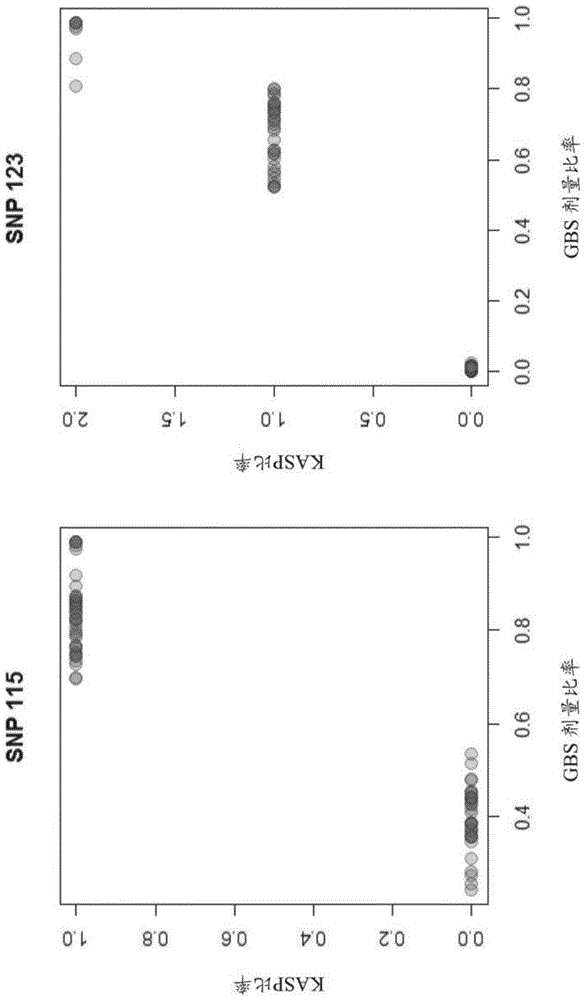
图1:以图形方式示出了两个抗性基因座(snp115和snp123)处的抗性等位基因和易感等位基因的拷贝数之间的估计比率与kasp基因型之间的相关性。对于gbs比率和kasp比率二者,较大的值都表示抗性等位基因的更多的拷贝;
图2:以图形方式示出了f1个体(选择种08041×选择种02033杂交)中两个snp标志物处的基因分型-测序剂量评分和抗性谱之间的关联。在两个标志物处,这种关联非常显著,因为携带更多抗性等位基因拷贝的个体通常抗性更强;
图3:以图形方式示出了f1个体(选择种08041×选择种02033杂交)中两个snp标志物处的kasp基因型评分和抗性谱之间的关联。在这两个标志物处,这种关联非常显著。
图4:示出了f1后代中多重gbs剂量比率分布的直方图。较大的gbs比率表明该个体携带所描述的单倍型(haplotype)的更多“抗性”拷贝;
实施例
实施例1
引言
martin,p.,&firman,i.(1970).resistanceofchrysanthemumcultivarstowhiterust(pucciniahoriana).plantpathology,180-184公开了菊属白锈病抗性植物的若干品种。为了评价与白锈病相关联的基因组序列,即seqidno.1和seqidno.2是否在所公开的菊属栽培品种中被发现,使这些栽培品种经受标志物分析,且结果示于下文表1中:
表1:先前由martin(1970)报道的抗堀柄锈菌或对堀柄锈菌免疫的菊属品种在用堀柄锈菌的nl1分离株接种后的表型。
如可以清楚看出的,martin等人中公开的上述植物中没有一种包含由seqidno.2表示的基因组序列。
实施例2
引言
白锈病是造成全世界的作物和花卉育种者的重大经济损失的植物中的病害。白锈病也影响菊属植物,且通常是由真菌堀柄锈菌引起的。感染后,在上表面形成浅绿色至黄色斑点,斑点的直径尺寸可多达5mm。随着植物衰老,这些斑点的中心适时变成棕色并坏死。在叶片的下表面上形成凸起的、浅黄褐色或粉色的小隆起物(孢子堆(telia))。该病害通常携带在温室菊花的受感染的插条和植株,包括切花上。
先前已经鉴定到与影响抗性的分子标志物seqidno.1和seqidno.2遗传连锁的两个基因组区域或基因,并且在这些基因座的每一个处赋予抗性的等位基因存在于品种中,这些品种的实例已经以编号ncimb42455保藏,并且可以通过英国阿伯丁ab219ya巴克斯本克莱斯通村弗格森大厦的国家工业、食品和海洋微生物保藏中心(ncimb)获得。虽然两个基因座都解释了用抗性品种产生的杂交种中的大量变异,但它们无法解释许多其他品种的抗性模式。例如,一些品种尽管缺乏在ncimb42455中鉴定到的基因座处的赋予抗性的等位基因,但完全抗白锈病。这表明,存在影响菊花抗性的尚未知的基因。本文提供的实施例已经鉴定出一个此类的基因组区域、或基因或等位基因。包含本发明的基因组区域或基因和与其遗传相锁的seqidno.3的植物已经以编号ncimb42762保藏并且可以通过英国阿伯丁ab219ya巴克斯本克莱斯通村弗格森大厦的国家工业、食品和海洋微生物保藏中心(ncimb)获得。
方法
杂交方案和实验设置
为了测试snp标志物和抗性谱之间的关联,f1群体通过将来自抗性桑蒂尼菊花(santinichrysanthemum)选择种08041的个体与来自易感多头菊花(spraychrysanthemum)选择种02033的个体杂交来产生。选择种08041是高度抗白锈病的。然后对262个f1个体(cr15-995004)进行白锈病抗性表型分析并且对于每个f1个体,测定6个克隆。在分组设计中进行堀柄锈菌接种,并且如上文描述的,在两个容器中进行(在不同时间)四次试验。对抗性进行评分使得大的值表示抗性(最大值为9)且小的值表示易感性(最小值为1)。复制克隆在测试和容器之间被随机分开。这其中,选择80个克隆用于基因分型。进行选择以最大化被基因分型的样品中存在的表型变异,这应该提高检测snp-表型关联的能力。
选择种08041抗性基因座
使用具有50,000个针对菊花的标志物的affymetrixsnp阵列,对上文提到的f1群体进行基因分型,并且在f1群体的每个个体中进行标志物-性状关联。这产生了9个单核苷酸多态性(snp),解释了菊花中白锈病抗性的大量表型变异:7个snp各自>50%,且2个snp各自超过70%。然后将后两个命名为snp115和snp123的snp作为多重基因分型-测序(gbs)方法的靶点,因为它们显示出与白锈病抗性的最强关联。由于菊花是六倍体,设计了一种方法来估计个体中某一基因座处存在的抗性拷贝数。然后使用这种方法对08041和02033品种之间的f1群体的个体进行基因分型。
对snp115和snp123标志物设计kasp测定。kasp方法是一种比gbs更具成本效益的方法。一个潜在的缺点是,它给出的抗性拷贝数的度量比根据gbs数据估计的剂量比率粗略得多(kasp比率仅可以是0、1或2)。菊花的六倍体性质使得在给定的基因座处的抗性等位基因的数目可以在0和6之间变化。精确了解个体携带的抗性等位基因数目不仅有利于映射(mapping)的目的,也有利于在随后的育种阶段期间选择个体。幸运地,kasp比率显示出与由gbs获得的剂量比率呈强相关(图1),这意味着它们给出了对个体植物携带的抗性单倍型的数目的良好指示。对亲本和所有f1后代二者都使用kasp方法进行基因分型。
用于基因分型的snp115(测定4)和snp123(测定18)的kasp引物是:gxm_wrr_测定4_a(seqidno.10):
gxm_wrr_测定4_b(seqidno.11):
gxm_wrr_测定4_r(seqidno.12):
gxm_wrr_测定18_a(seqidno.13):
gxm_wrr_测定18_b(seqidno.14):
gxm_wrr_测定18_r(seqidno.15):
结果
在源自抗性选择种08041和易感选择种02033之间的杂交的后代的f1克隆内部及它们之间都存在抗性异质性。基于六次重复,一些克隆具有平均抗性值9(完全抗性),而其他克隆具有平均抗性值1(完全易感)。一些克隆显示很小的异质性(重复之间的标准偏差为0),而少数其他克隆显示出较大的异质性水平(标准偏差为3.7)。
与菊花的白锈病抗性最强关联的两个snp是重叠群72689上的72689_15854_115(下文称为snp115)和72689_15854_123(下文称为snp123),该重叠群映射在番茄1号染色体(96.77mb)上。
被证明赋予白锈病抗性的单倍型的dna序列在下文示出。为了便于说明,snp115(g)和snp123(t)以粗体突出显示:
seqidno.3:
还鉴定出与数据集中的易感性相关的六种单倍型。snp115和snp123以粗体突出显示并且发现与抗性单倍型不同的核苷酸的位置被加下划线。
在两个基因座处的gbs剂量(图2)和kasp基因型(图3)与克隆的抗性评分之间存在明显且非常显著的关联(在所有情况下均p<1*10-16)。就snp123而言,其中基因型和抗性之间的关联显示出可以潜在地暗示该基因座处的显性的模式。基因型评分为1的个体与具有kasp基因型评分为2的个体具有一样的抗性,而基因型分类为0的个体通常是非常易感的(除了一个异常值,图2)。然而,被归类为具有kasp基因型为零的个体也可能携带抗性单倍型的拷贝,因此需要更多的研究以阐明抗性规模是否随抗性拷贝数叠加,或者抗性单倍型是否相比于易感单倍型是占优势的。对于两个snp标志物,存在一些基因型比基于其基因型预测的抗性更强,而有些基因型抗性更弱(图2)。这些情况可以通过实验室或实验中的样品交换(sampleswap)来解释,或者通过抗性等位基因和赋予抗性的实际(未鉴定的)基因之间的不完全ld来解释,因为鉴定的“抗性”单倍型虽接近该基因(如通过个体携带的抗性等位基因的拷贝数与其抗性谱之间的非常显著的关联所确认的),但并不是完全连锁的是可能的。结果是,由于新近的和/或过去的重组事件,抗性单倍型的一些拷贝未完全连接基因的抗性拷贝。另外的可选解释是,在08041或02033品种中存在具有小效应的其他基因的变体,或者遗传变体的外显率是不完全的。
一个有趣的观察是,对于snp123,一些个体具有2的kasp基因型,而抗性更强的亲本(选择种08041)仅具有1的kasp基因型(图3)。这是令人惊讶的,因为这表明一些f1后代在该区域具有比选择种08041更多的抗性单倍型的拷贝,这与预期相反。因此,我们更详细地研究了f1中gbs剂量比率的分离。
我们发现snp123处的剂量比率的分布极不寻常(图4)。并且当亲本的剂量比率是未知的时,我们使用模拟来测试我们是否可以重现这些分布。我们模拟了各自在感兴趣的基因座处具有7种可能的基因型之一(aaaaaa、baaaaa、bbaaaa等)的亲本之间的杂交。然后,对于49对成对杂交中的每一个,我们随机抽取配子,假设多体遗传导致后代的六倍体基因型。该过程随机重复总计50次。
如果我们观察模拟的分布,我们观察到对于snp115,剂量比率的经验分布最接近地类似于其中抗性最强的亲本有5个抗性等位基因拷贝并且易感亲本具有2个抗性单倍型拷贝的情况的分布。对于snp123,观察到的剂量比率的分布与亲本对的任何模拟组合都不匹配。这种差异是否具有生物学的或技术上的解释值得在未来进行研究,从对亲本系进行基因分型开始。
结论
对影响白锈病(堀柄锈菌)抗性的基因及其变体的精确了解对于培育抗性更强的菊花品种是关键。先前已经鉴定到两个主要的效应基因座(seqidno.1和seqidno.2)。
据推测,一定存在其他能够赋予菊花白锈病抗性的基因。本文提供的结果明确地显示,两个密切连锁的snp标志物处的变体可以强烈地预测抗性,表明这些snp及发现其一个的单倍型与致病基因(causalgenes)呈紧密连锁。
序列表
<110>多盟集团公司
<120>白锈病抗性菊属植物
<130>4/2vs58/17p
<150>pct/ep2016/073672
<151>2016-10-04
<160>15
<170>bissap1.3.6
<210>1
<211>192
<212>dna
<213>菊属(chrysanthemum<genus>)
<220>
<223>29tgr2周围的序列
<400>1
ttaactcaaaaatatgactacaaatcaattttcaggactttttttcgatacttccctctt60
tggtaccggtaccgtattagtggtaccgattttttttgggctcaattcatggtacaggca120
ccgtaccgtgtattgggagtcggtaccgtttcgatacggtaccggtacggttccgattcg180
ataccggaattc192
<210>2
<211>180
<212>dna
<213>菊属(chrysanthemum<genus>)
<220>
<223>24hs1r1周围的序列
<400>2
gaattcctatacgaaggttttgtagatgtgtctccgagcgagtttgatcctaactcacga60
ctagtaatttatgtccataaaaccgataacttgaacattccctcttgttcttgtagatgg120
ggatgacttcactgagtctccattcttctggcatcttatcactggaaaaaatcttgttaa180
<210>3
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>3
agcaaaacatgcagtgatttatcctcgttaagtgtatggaaaatcgacaccagggtgc58
<210>4
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>4
accaaaacaggcagtgatttatcctcgttaagtgtatggaagatcgacaccagagtgc58
<210>5
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>5
agcaaaacaggcagtgatttatcctcgttaagtgtatggaagatcgacaccagagtgc58
<210>6
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>6
accaaaacaggcagtgatttatcctcgttaagtgtacggaagatcgacagcagggtgc58
<210>7
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>7
accaaaacaggcagtgatttatcctcgttaagtgtacggaagatagacagcagggtgc58
<210>8
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>8
agcaaaacaggcagtgatttatcctcgttaagtgtatggaagatcgacaccagggtgg58
<210>9
<211>58
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>9
accaaaacaggcagtgatttatcctcgttaagtgtatggaagatcgacaccagggtgg58
<210>10
<211>43
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>10
gaaggtgaccaagttcatgctcaagtcttgtacaaycaaggag43
<210>11
<211>43
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>11
gaaggtcggagtcaacggattcaagtcttgtacaaycaaggac43
<210>12
<211>23
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>12
tacacttaacgaggataaatcac23
<210>13
<211>42
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>13
gaaggtgaccaagttcatgcttacaaccaaggascaaaacat42
<210>14
<211>42
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>14
gaaggtcggagtcaacggatttacaaccaaggascaaaacag42
<210>15
<211>23
<212>dna
<213>菊属(chrysanthemum<genus>)
<400>15
gtcgattttccatacacttaacg23
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!